Introduction
Glioma is the most prevalent and most aggressive
primary brain tumor in adults, accounting for >50% of all tumors
of the central nervous system (1).
Although a combination of microsurgery, radiotherapy and
chemotherapy have been widely used to treat glioma, the prognosis
of patients with malignant glioma remains poor, with a median
survival time in the range of 12 to 15 months (2). Dozens of molecules and signal pathways
have been established to be involved in the genetic alterations and
chromosome aberrations that occur during tumorigenesis. Therefore,
the development of efficient treatment therapies, based on an
improved understanding of the pathophysiological and molecular
properties of malignant glioma, are required.
Human corticotropin-releasing factor (hCRF), a
41-residue neuropeptide, is produced in the hypothalamus and is an
important component in the regulation of the
hypothalamic-pituitaryadrenal axis. CRF receptors constitute a
family of G-protein-coupled receptors, of which there are two major
classes, CRFR1 and CRFR2 (3,4). CRFR1 binds CRF with a higher affinity
compared with CRFR2 and is expressed primarily in the brain and
pituitary gland. The activation of CRFR1 exerts numerous central
and peripheral effects associated with pathological disease
(5). In addition to the expression of
CRFRs in the central nervous system, receptors are identified in
the immune, cardiovascular and reproductive systems, suggesting
that the CRF receptors may mediate numerous diverse functions
(6,7).
Previous studies have established that CRF and its
receptors may contribute to the pathogenesis of neurodegenerative
disorders, including brain injury, ischemia, Alzheimer's disease
and Parkinson's disease (8–11). CRF-family proteins and their receptors
have been detected in a number of tumor types. Numerous epithelial
tumors, epithelial tumor cell lines and neuroendocrine tumor cell
lines have been demonstrated to express CRF receptors (12,13). CRFR1
has been detected in the MCF7 breast cancer cell line, in human
melanomas and in melanoma cell lines (14–16). Even
though the expression of CRF and its receptors has been described
in different types of tumors and cancer cells, the role of these
peptides has not been previously investigated in human glioma and
malignant glioma cell lines. However, evidence suggests that hCRF
may represent a novel therapeutic target in peritumoral brain edema
(17).
In the present study, the expression levels of CRFR1
in human glioma tissues and U87 glioblastoma cell lines were
assessed, and its association with pathological grades determined.
The U87 glioma cell line was subsequently treated with hCRF to
assess the action of the ligand on cell proliferation, cell cycle
and apoptosis. hCRF was identified to affect cell proliferation,
the cell cycle and apoptosis of U87 glioma cells via targeting p53,
demonstrating CRFR1 may be a putative therapeutic target for human
glioma treatment.
Materials and methods
Human sample collection
Human glioma samples were obtained from the
Department of Neurosurgery, The Second Affiliated Hospital of Hebei
Medical University (Hebei, China) in 2014. Of the 35 samples, 18
were from male patients and 17 from females, with a mean age ±
standard deviation (SD) 45.15±12.35 years (age range, 3–62). The 35
tumor samples were divided into 4 groups (18–20):
Medulloblastoma (n=5); grade II (n=8); grade III (n=12); and grade
IV (n=10). All specimens were obtained during initial surgery and
patients were not administered preoperative radiotherapy,
chemotherapy or immunotherapy. Diagnoses were established using
histology staining by two pathologists, according to the WHO
classification guidelines (21).
Normal cerebral tissues, used as controls, were obtained from 13
patients suffering from severe brain injury who underwent
intracranial decompression. All samples were stored at −80°C in
liquid nitrogen immediately following resection. Data collection
and analysis were approved by the ethics committee of The Second
Affiliated Hospital of Hebei Medical University.
Cells and cell culture
Human U87 glioma cells were purchased from the
Chinese Academy of Sciences Cell Bank (Shanghai, China). The cells
were maintained in Dulbecco's modified Eagle's medium (HyClone,
Logan, UT, USA) enriched with 10% fetal bovine serum (HyClone; GE
Healthcare Life Sciences, Shanghai, China), 100 U/ml penicillin and
100 µg/ml streptomycin at a 37°C with 5% CO2.
hCRF treatment
hCRF was purchased from Shanghai Shenggong Biology
Engineering Technology Service, Ltd. (Shanghai, China). U87 cells
were divided into 4 groups, including the control group, U87 cells
treated with 3 doses of hCRF (10−7, 10−8 and
10−9 mol/l). All cells in this and the remaining
experiments were harvested at 24, 48 and 72 h. The experiment was
repeated 3 times.
Immunohistochemistry (IHC)
staining
IHC staining of tumor tissue was performed following
the standard procedures (22). In
brief, paraffin-embedded tumor tissue sections (5-µm) on
poly-L-lysine-coated slides were heated for 30 min at 60°C, dewaxed
in xylene and rehydrated in an ethanol series of 100 (10 min), 95
(5 min), 80 (5 min) and 75% (5 min), then submerged in water,
followed by a 5-min microwave (400 W) antigen retrieval set.
Endogenous peroxidase was inactivated with 3%
H2O2 for 20 min and sections were incubated
in 5% bovine serum albumin (BSA) solution for 20 min to block
non-specific binding. Subsequently, sections were incubated
overnight at 4°C with goat anti-CRFR1 polyclonal antibodies (cat.
no., ab59023; 1:100; Abcam, Cambridge, UK). The membraned were
washed 3 times in PBS for 5 min, followed by a biotinylated
secondary anti-goat antibody (dilution, 1:200; cat. no., sc-2350;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) incubation for 30
min. 3,3′-Diaminobenzidine was used to reveal the IHC reaction and
the slides were counterstained with hematoxylin, and examined using
a Leica DM1000 microscope (Leica, Microsystems GmbH, Wetzlar,
Germany). Primary antibodies were substituted for phosphate
buffered saline (PBS) in the negative control.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the tumor tissues and
U87 glioma cells using Trizol reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
protocol. Total RNA (2 µg) was reverse-transcribed with M-MLV
reverse transcriptase (Invitrogen Thermo Fisher Scientific, Inc.,)
and oligo (dT) primers (Sangon Biotech Co., Ltd., Shanghai, China).
cDNA (1 µl) was used for qPCR, which was performed detect CRFR1,
p53 using SYBR Green Master mix (Takara, Bio, Inc., Otsu, Japan)
according to the manufacturer's protocol. Sequences of CRFR1, p53
and GAPDH primers were as follows: CRFR1 forward,
5′-TACGACAATGAGAAGTGCTGGT-3′ and reverse,
5′-GGACGATGTAAAGGAAGAT-3′; p53 forward,
5′-CGCCTGAGGTTGGCTCTGACTGTA-3′ and reverse,
5′-GTCTCTCCCAGGACAGGCACAAAC-3′; GAPDH (internal control) forward,
5′-ACGAGGCCCAGAGCAAGAGA-3′ and reverse, 5′-AGGTGTGGTGCCAGATTTTC-3′.
qPCR was performed with an initial denaturation step at 95°C for 30
sec, followed by 40 cycles of 95°C for 5 sec and 60°C for 40 sec.
Data were analyzed using GraphPad Prism 4.0 software (GraphPad
Software, Inc., La Jolla, CA, USA). The 2−ΔΔCq method
was used to quantitate the relative gene expression (23). PCR experiments were repeated 3
times.
Western blot analysis
Western blot analysis was performed to evaluate the
expression levels of CRFR1 in glioma tissues and U87 cells and the
protein levels of p53 were evaluated in U87 cells. Total proteins
were extracted and the protein concentration was determined using a
bicinchoninic acid assay kit (Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China). Equal amounts of protein (50
mg) were separated using 10% SDS-PAGE, transferred onto
polyvinylidene fluoride membranes (Roche Diagnostics GmbH,
Mannheim, Germany) by a transfer apparatus at 200 mA for 50 min.
The membrane was blocked with 5% skimmed milk for 2 h, and
incubated with indicated primary antibodies overnight at 4°C,
including goat anti-CRFR1 polyclonal antibodies (1:1,000; cat. no.,
ab59023; Abcam), rabbit anti-p53 polyclonal antibodies (cat. no.,
ab1431; 1:1,000; Abcam) and rabbit anti-β-actin monoclonal antibody
(1:1,000; cat. no., RB-9421-P0; Affymetrix, Inc., Santa Clara, CA,
USA). Then membranes were incubated with horseradish peroxidase
conjugated anti-rabbit or anti-goat IgG (1:5,000; cat. nos. sc-2370
and sc-2350; Santa Cruz Biotechnology, Inc.) for 2 h at room
temperature. Blots were detected using an enhanced
chemiluminescence kit and the densitometry signals were quantified
using Image Lab (Image Lab v4.1; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The immunoreactive bands of all proteins were
normalized to the intensity of corresponding bands for β-actin.
Results were analyzed using the National Institutes of Health Image
v1.41 software (Bethesda, MD, USA).
Cell proliferation assay using cell
counting for quantification
The U87 cells (control group) or treated with hCRF
(10−7 mol/l) were cultured for 48 h and were collected
in the logarithmic growth phase. The cells were then resuspended,
seeded in triplicate in 96-well plates at a density of 2,000
cells/well and cultured at 37°C in an incubator with a 5%
CO2 atmosphere. After 24 h, images of the cell plates
were captured under a Nikon 90I light microscope (Nikon, Japan) at
×100, magnification and the images were quantified using Cellomics
ArrayScan VTI (Thermo Fisher Scientific, Inc.) once a day for 3
days and then a cell growth curve was generated based on data
obtained using the Cellomics ArrayScan VTI.
5-bromo-2-deoxyuridine (BrdU) cell
proliferation assay
The U87 cells treated with hCRF (10−7
mol/l) were cultured for 48 h. The cells were then resuspended,
seeded in 96-well plates at 1×106 cells per 200 µl, and
then cultured for 24–72 h. The cell proliferation was assessed
using a BrdU Cell Proliferation ELISA kit (Roche Applied Science,
Rotkreuz, Switzerland) according to the manufacturer's protocol.
During the final 2–24 h, BrdU reagents were diluted at a ratio of
1:100 and added to the cells (10 µl/well). Then, the cells were
fixed using 200 µl FixDenat per well for 30 min and blocked with 5%
BSA for 30 min at room temperature. Anti-BrdU-POD monoclonal
antibody (dilution, 1:100; cat. no. 11444611001; Roche Applied
Science, Penzberg, Germany) and added (100 µl/well) to the cells
for 90 min at room temperature. Following 3 washes with washing
buffer, substrate solution (100 µl/well) (provided in the ELISA
kit) was added and the cells were incubated for 5–30 min in the
dark. The color reaction was activated with 10%
H2SO4 for 30 min and the BrdU quantity was
determined at a wavelength of 450 nm using an ELx800 Absorbance
Mircoplate Reader (BioTek Instruments, Inc., Winooski, VT,
USA).
Flow cytometry
For cell-cycle analysis, the control and hCRF group
cells in a log phase of growth were harvested at a density of
5×105 cells and washed with PBS twice, fixed with
ice-cold 70% ethanol for >1 h. Subsequent to washing with PBS,
the cells were treated with RNase A, 0.2% Triton X-100 and 50 µg/ml
propidium iodide stain (PI; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) at 4°C for 30 min. The cell cycle profiles were then
analyzed using flow cytometry. The experiments were performed in
triplicate. The apoptosis analysis was performed in control and
hCRF group cells by staining with the Annexin V V-APC Apoptosis
Detection kit (BioVision, Inc., Milpitas, CA, USA) according to the
manufacturer's protocol. In brief, cells were harvested at a
density of 1×106 cells/ml, and 5×105 cells
were plated into a 6-well plate and treated with 0.1% trypsin and
0.02% EDTA, and were washed twice with PBS at 4°C prior to
collection. The cells were mixed with staining buffer and 100 µl of
this cell suspension were mixed with 5 µl of annexin V solution at
room temperature in the dark for 10–15 min. The level of cell
apoptosis was analyzed using flow cytometry (FACSCalibur). All
assays were performed in triplicate.
Statistical analysis
Data were analyzed using the statistical package SAS
version 8 software (SAS, Inc., NC, USA) and all experiments were
repeated >3 times. All the data were expressed as mean ± SD.
Significant differences between the groups were analyzed using a
χ2 test and the one-way analysis of variance, followed
by the Student-Newman-Keuls test. P<0.05 was considered to
indicate a statistically significant difference.
Results
High CRFR1 expression is associated
with malignant phenotypes of human glioma
In the present study, the expression pattern of
CRFR1 was evaluated in 35 glioma tissues and 13 normal brain
tissues using IHC staining. CRFR1 expression was significantly
increased in glioma tissues, but minimally detected in the normal
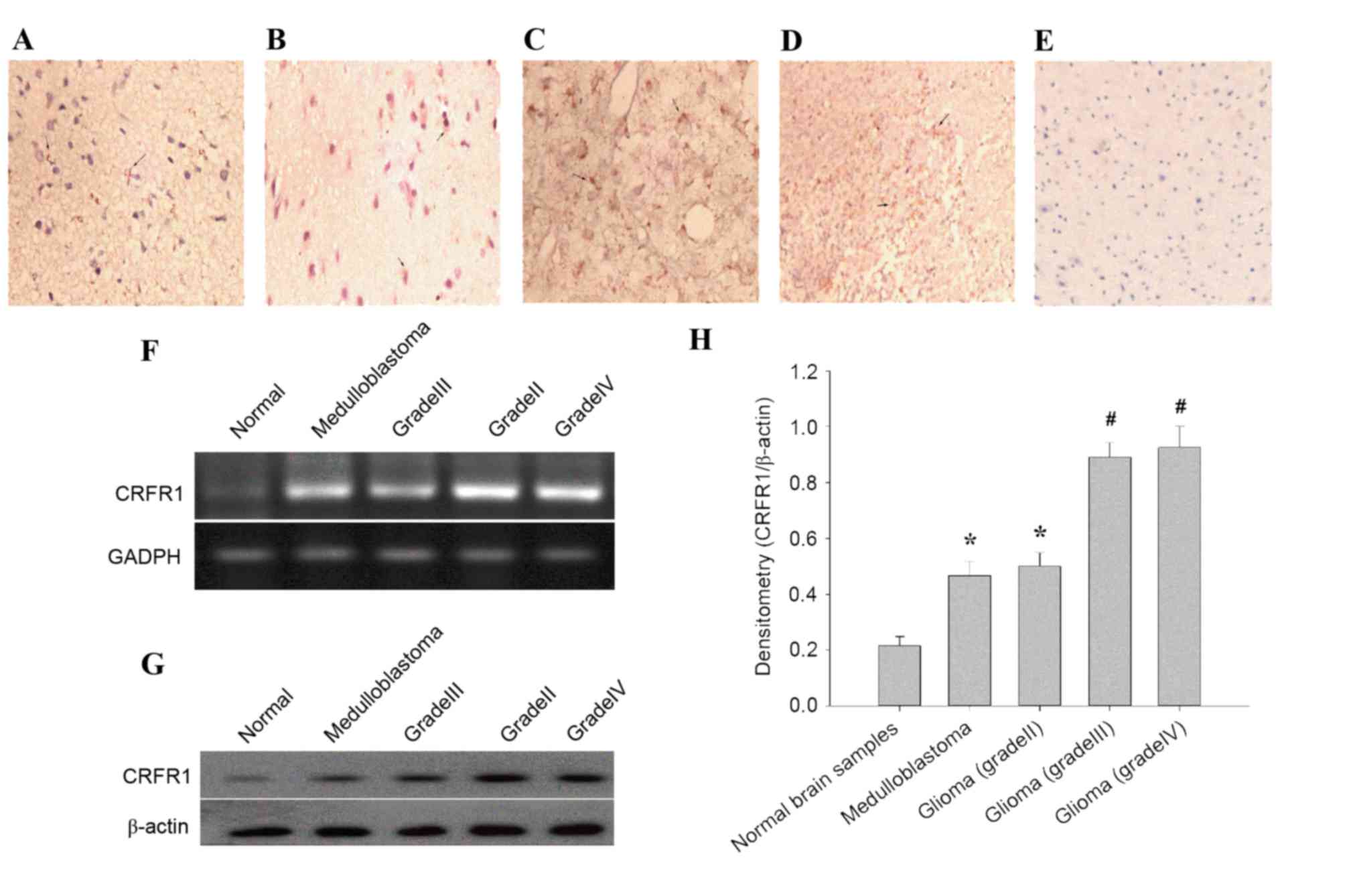
brain specimens (Fig. 1A-E). In
glioma cells positive for immunostaining, the labeling was
primarily cytoplasmic as observed using light microscopy. The
positive expression rate of CRFR1 was significantly different
(χ2=17.0817, P<0.0001) between the human glioma
(74.29%, 26/35) and the normal brain tissues (7.69%, 1/13; Table I). Furthermore, the association
between CRFR1 expression and clinicopathological characteristics
were analyzed in the current study. There was no significant
association observed between CRFR1 expression and age, gender and
position (all P>0.05). Notably, a significant difference
(χ2=6.4852, P=0.0396) was identified between the
low-grade (grade II, 2/8, 25%) and the high-grade (medulloblastoma,
4/5, 80%; grade III and IV, 16/22, 72.72%) glioma (Table II). Semi-quantitative RT-PCR results
demonstrated that CRFR1 mRNA levels were significantly higher in
human glioma compared with the normal cerebral tissues (Fig. 1F). In addition, the protein levels of
CRFR1 was determined using western blot analysis, the results
demonstrated that CRFR1 protein levels were significantly increased
in human glioma compared with the normal cerebral tissues (Fig. 1G and H). These results suggested that
increased CRFR1 expression is associated with high-grade
glioma.
 | Table I.Expression of CRFR1 in human gliomas
and normal brain samples using immunohistochemical staining. |
Table I.
Expression of CRFR1 in human gliomas
and normal brain samples using immunohistochemical staining.
|
|
| Expression status
of CRFR1 |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
| Group | Cases, n | Negative | Positive | CRFR1-positive,
% |
χ2-value | P-value |
|---|
| Glioma tissues | 35 | 9 | 26 | 74.29 | 17.0817 | <0.0001 |
| Normal brain
tissues | 13 | 12 | 1 |
7.69 |
|
|
| Table II.Association between CRFR1 expression
and clinicopathological parameters assessed by immunohistochemical
staining. |
Table II.
Association between CRFR1 expression
and clinicopathological parameters assessed by immunohistochemical
staining.
|
|
| CRFR1 expression,
n |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
| Clinicopathological
characteristics | Cases, n | Negative | Positive | CRFR1-positive
tumors, % |
χ2-value | P-value |
|---|
| Age, years |
|
|
|
| 0.0204 | 0.8864 |
|
<45 | 24 | 6 | 18 | 75.00 |
|
|
|
≤45 | 11 | 3 | 8 | 72.73 |
|
|
| Gender |
|
|
|
| 0.0826 | 0.7738 |
|
Male | 18 | 5 | 13 | 72.22 |
|
|
|
Female | 17 | 4 | 13 | 76.47 |
|
|
| Position |
|
|
|
| 0.0374 | 0.8467 |
|
Supratentorial | 28 | 7 | 21 | 75.00 |
|
|
|
Subtentorial | 7 | 2 | 5 | 71.43 |
|
|
| Glioma grade |
|
|
|
| 6.4852 | 0.396 |
|
Medulloblastoma | 5 | 1 | 4 | 80.00 |
|
|
| II | 8 | 6 | 2 | 25.00 |
|
|
|
III+IV | 22 | 6 | 16 | 72.72 |
|
|
CRFR1 is expressed in human U87 glioma
cells
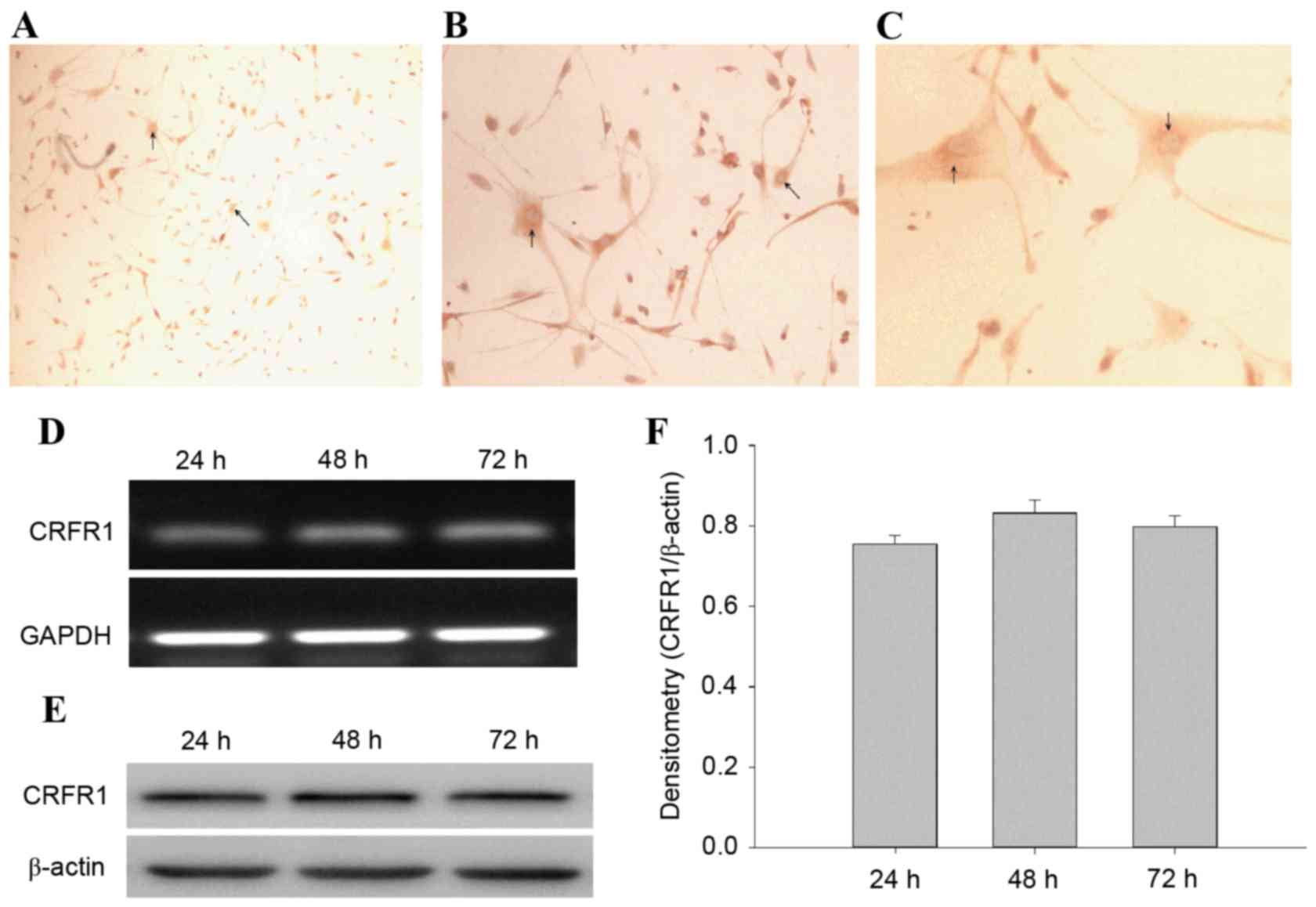
The CRFR1-positive cells were observed and the
immunostaining was primarily cytoplasmic (Fig. 2A-C). CRFR1 mRNA and protein levels
were determined using semi-quantitative RT-PCR and western blot
analysis at 24, 48 and 72 h. The results revealed that the CRFR1
mRNA and protein was expressed in U87 cells at each time point
(Fig. 2D and E). However, there was
no significant difference between the levels of CRFR1 expression
between the 3 time points (Fig.
2F).
hCRF inhibits the proliferation of U87
glioma cells
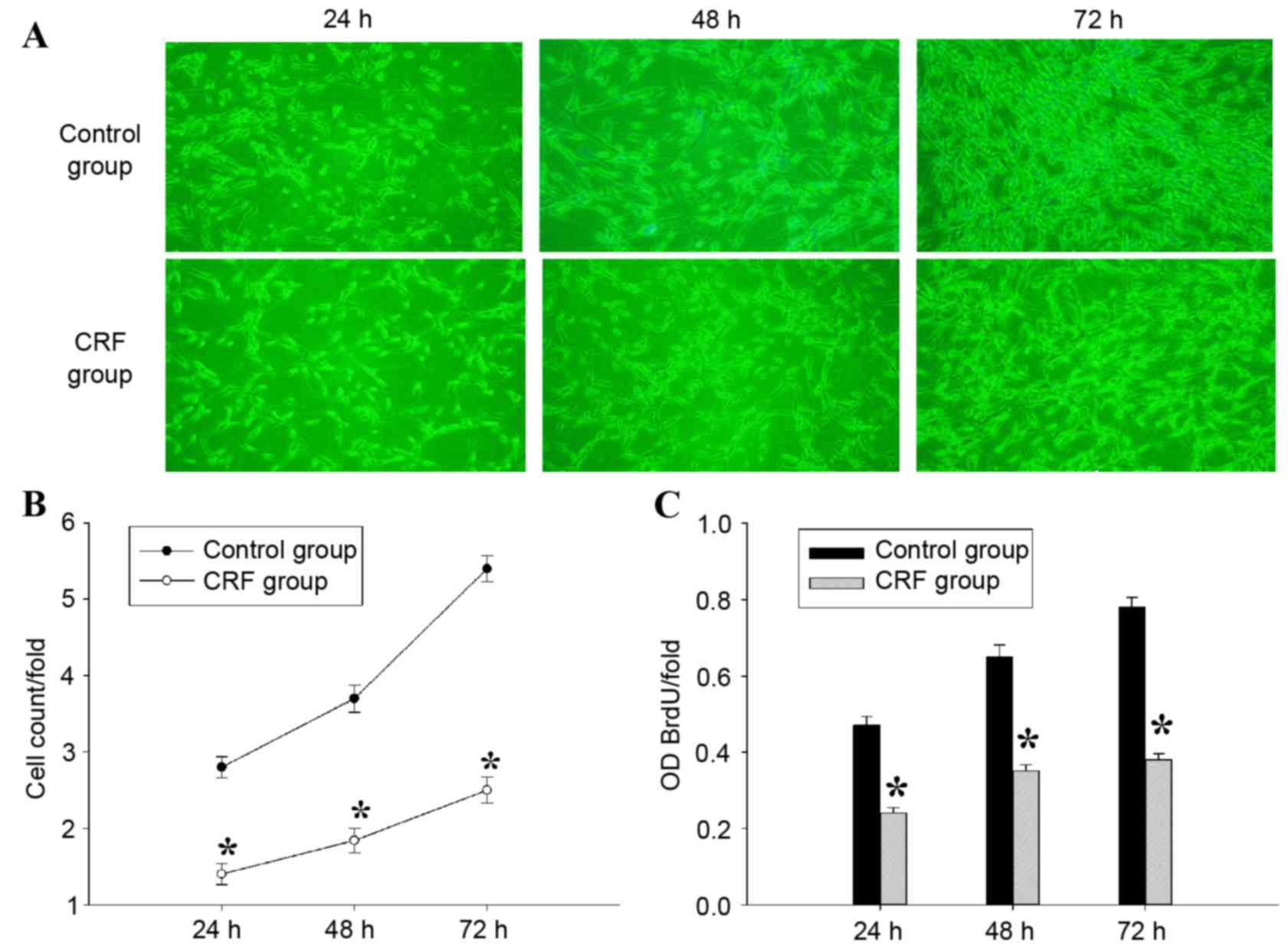
In order to determine the effect of hCRF on cell
growth, U87 cells were treated with or without hCRF
(10−7 mol/l) and cultured for 24, 48 and 72 h. The cell
number was measured using Cellomics ArrayScan VTI every day for 3
days, and a significant inhibition of cell proliferation was
observed in U87 cells from 24 to 72 h following treatment with hCRF
(Fig. 3A and B). To confirm the
suppressive effect of hCRF on the proliferation of U87 cells
further, a BrdU incorporation assay was performed to assess cell
proliferation status. In this assay, cell proliferation was
evaluated by measuring a DNA synthesis marker, which was quantified
by the BrdU incorporation ratio. Consistent with the results of
cell counting, the quantity of newly synthesized DNA decreased
significantly at 24, 48 and 72 h following treatment with hCRF,
compared with the control group (Fig.
3C). These results indicated that cell proliferation and DNA
synthesis were suppressed by hCRF in human U87 glioma cells.
hCRF induces cell cycle arrest and
promotes apoptosis in human U87 glioma cells
To investigate the underlying mechanisms of the
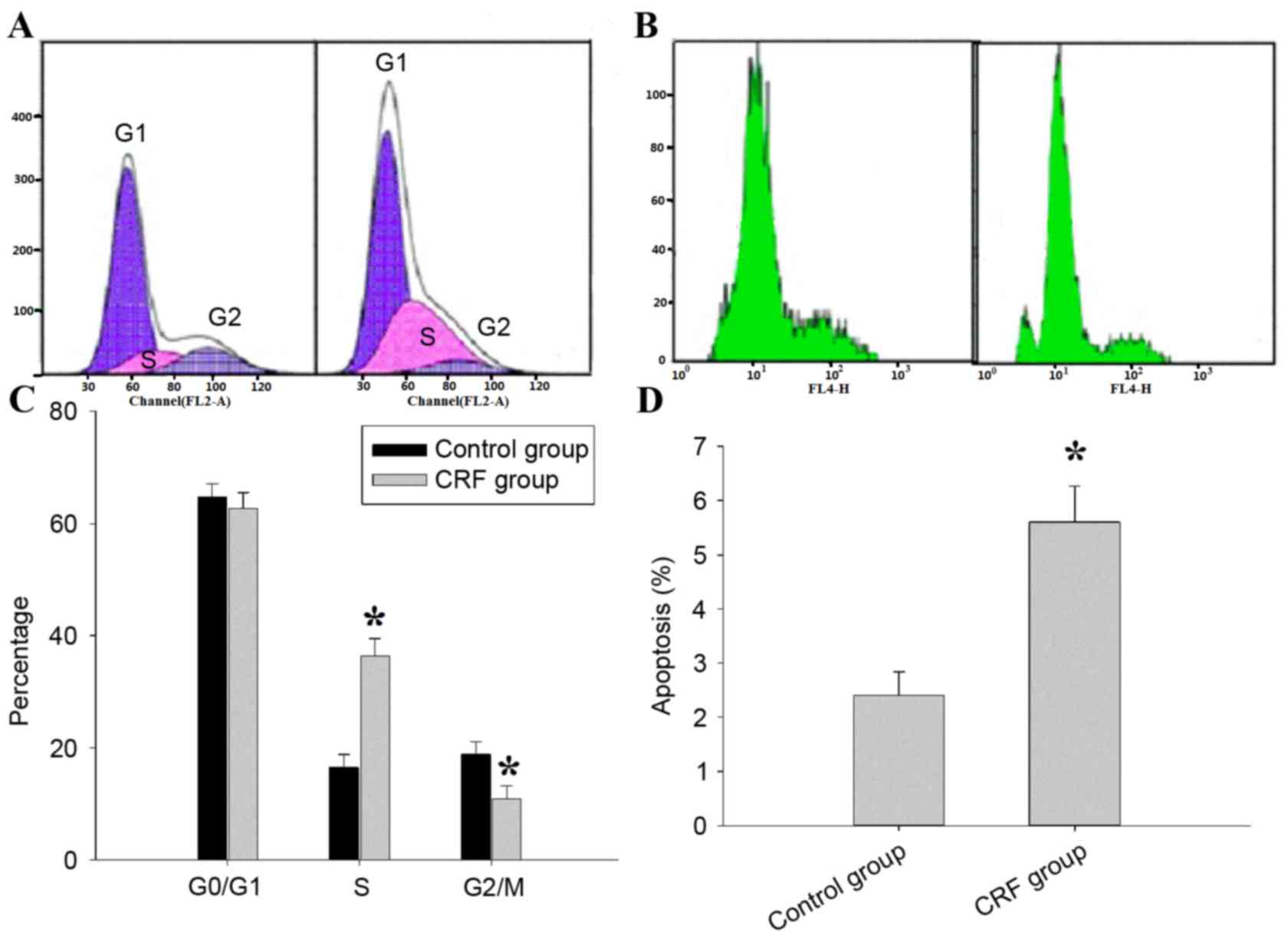
inhibitory effect of hCRF on U87 cell growth, the role of hCRF in
cell cycle progression of the U87 was assessed using PI-FACS
(Fig. 4A). In the control group,
65.32±1.15, 16.53±0.78 and 18.81±0.45% cells were found in the
G0/G1, S and G2/M phases,
respectively; in the hCRF group, 56.18±0.87, 34.62±0.49 and
10.87±0.68% cells were in the G0/G1, S and
G2/M phases, respectively. U87 cells that underwent hCRF
treatment exhibited an increased percentage of cells in the S phase
compared with control (*P<0.01) and concomitantly with a
significant decrease in G2/M phase cells (*P<0.01).
The level of cell apoptosis was assessed using annexin V staining
and flow cytometry (Fig. 4B). Cell
apoptosis was significantly increased in hCRF group when compared
with the control group (5.41±0.28 vs. 2.64±0.15, *P <0.01). The
proportion of cells in S phase increased from 16.53 to 34.62% in
the hCRF-treated group, whereas the proportion in the G2/M phase
decreased from 18.81 to 10.87% (Fig.
4C). Therefore, the present results indicated that hCRF
treatment led to the inhibition of the S-M transition. Apoptosis
was observed in 5.68% of U87 cells treated with hCRF, while only
being observed in 2.45% of U87 cells in the control group (Fig. 4D). These data suggest that hCRF may
promote cell apoptosis in U87 cells.
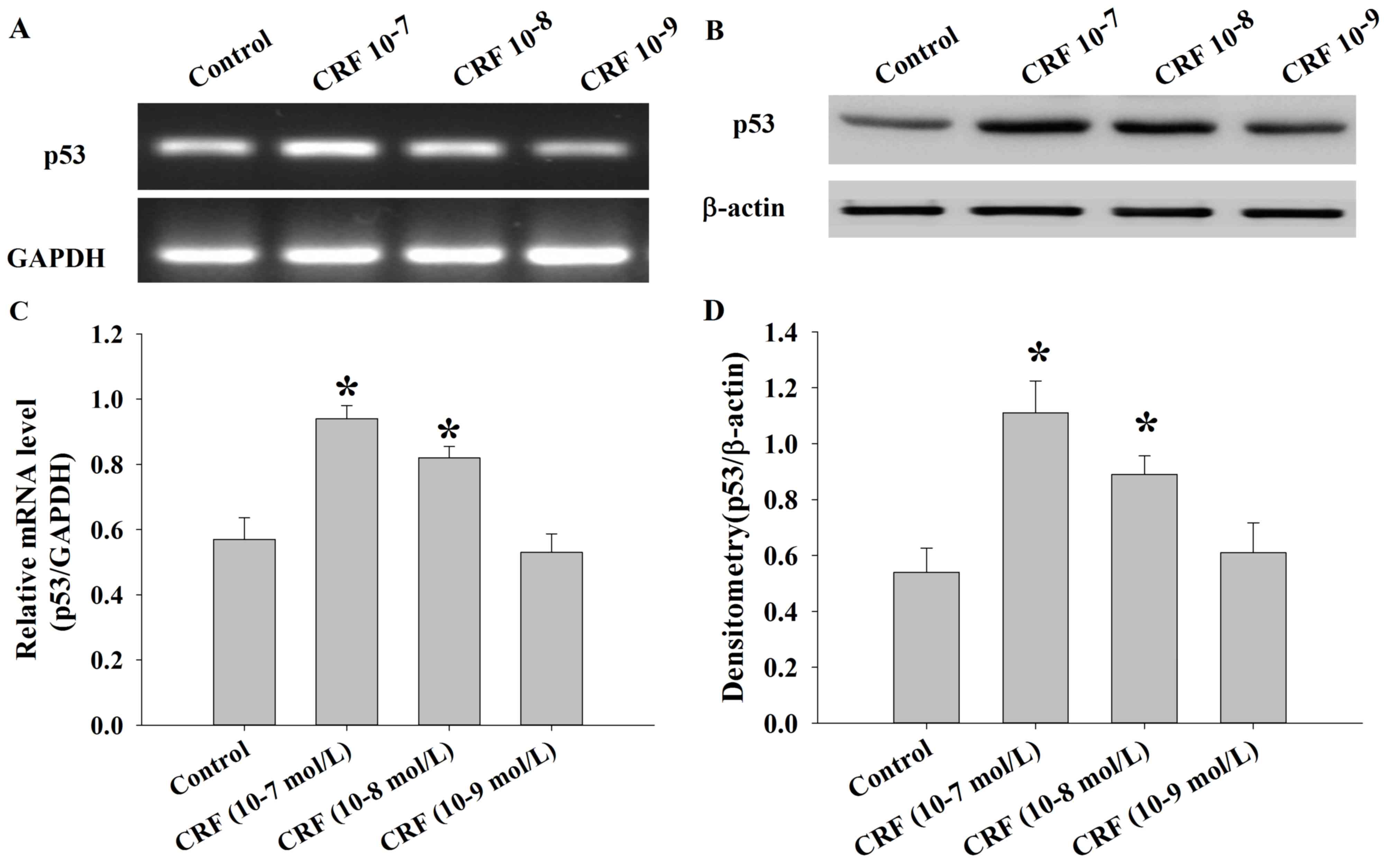
hCRF upregulates the expression of p53
in human U87 glioma cells
The levels of p53 mRNA and protein were determined
in U87 cells following treatment with control or hCRF (at
10−7, 10−8 and 10−9 mol/l) group
using semi-quantitative RT-PCR and western blot analysis. The
results demonstrated that the expression of p53 mRNA was
significantly upregulated in the hCRF treatment group compared with
the control group (*P<0.01; Fig. 5A
and B). Consistent with these results, the levels of p53
protein were also significantly increased in the groups treated
with higher hCRF concentrations (10−7 and
10−8 mol/l) compared with the control or low hCRF
concentration (10−9 mol/l) group (*P<0.01; Fig. 5C and D).
Discussion
Glioma are the most prevalent, aggressive type of
tumor observed in the central nervous system and the prognosis
remains poor due to their biological characteristics (24). CRF is a major hypothalamic
stress-induced neuropeptide that is also expressed in peripheral
tissues. CRF and its receptors have been detected in a number of
tumor types. In a previous study using an animal tumor model, CRF
has been established to inhibit tumor proliferation (25). This action is mediated by the specific
CRF receptors, CRFR1 and CRER2 (26).
The aim of the present study was to determine the expression of
CRFR1 in human glioma tissue and the human U87 glioma cell line,
and to evaluate the potential effect of CRF on human U87 glioma
cell proliferation, apoptosis and the cell cycle.
Previous studies have demonstrated that CRFR1 is
expressed in tumor cells, and CRF and urocortin may reduce tumor
cell growth via their interaction with CRFR1 (13,25). A
number of malignancies have been established to express high levels
of CRFR1 and CRFR2 and to be sensitive to the suppressive effects
of CRF and its agonists; however, there is little currently
understood about the role of CRF in glioma. To the best of our
knowledge, the present study is the first to investigate the
specific expression of CRFR1 in human glioma cells. The increased
expression of CRFR1 is associated with an increased glioma grade,
which provides additional support for the involvement of CRF in the
development of glioma. However, CRFR1 expression was minimally
detected in normal brain tissue. Following analysis of U87 glioma
cells, the expression of the CRFR1 gene and protein were
significantly increased in U87 glioma cells at all time points
following CRF treatment. These results suggested that the role of
CRF in glioma is distinct and essential.
In order to further characterize the effect of CRF
in glioma, U87 cells were treated with hCRF [10−7 mol/l;
the dosage was determined in accordance with a previous study
(25)]. Compared to the control
group, U87 cells that underwent hCRF treatment exhibited reduced
cell proliferation, were arrested in S-phase and demonstrated an
increased level of apoptosis. These results suggest that CRF may be
involved in the regulation of cell cycle checkpoints in U87 cells
and may demonstrate that CRF promotes a marked reduction in
replication initiation in glioma cells. Therefore, CRF has an
important role in promoting U87 cell growth and is associated with
U87 cell cycle phase distribution. Using primary neuronal cultures
from the cerebellum, cerebral cortex and hippocampus, low CRF
concentrations (10−8 mol/l) have been reported to exert
a brain region-specific neuroprotective effect on amyloid-ß 25–35
toxicity in the cerebellum and hippocampus (27). However, a higher CRF concentration
(10−7 mol/l) led to the additional protection of
cortical neurons (25). CRF was
reported to act on CRFR1 to inhibit the proliferative effects of
estrogens on MCF7 cells in a paracrine and autocrine manner
(16) and urocortin inhibited the
proliferation of melanoma cells in vitro and in vivo,
also through CRFR1 (28). Moroz et
al (29) reported that hCRF was
more effective compared with dexamethasone or temozolomide in the
treatment of U87 xenografts, with certain patients reporting an
increase in long-term survival and only mild toxicity. Therefore,
the results of the present study are in accordance with these
previous studies, as hCRF inhibits the proliferation and promotes
the apoptosis of U87 cells in vitro via CRFR1.
p53 is one of the most extensively studied molecules
in cancer research and molecular biology. p53 has numerous
anticancer functions and serves vital roles in DNA repair,
apoptosis and inhibition of angiogenesis (30). Previous studies have reported that the
wild type p53 gene is present in ~70% of primary glioblastoma
(31), however U87 possesses wild
type p53 (http://p53.free.fr/Database/Cancer_cell_lines/p53_cell_lines.html).
Xu et al (32) reported that
histone acetyltransferase inhibitor II inhibits proliferation and
induces apoptosis via the caspase-dependent pathway in human U87
and U251 glioma cell lines, potentially by activating the p53
signaling pathway (32). Another
study identified that the CRFR1-triggered ERK1/2 pathway is
involved in the p53 activation and the suppression of the apoptotic
Bax gene in rat liver cells following hypoxia (33). The present study also hypothesizes
that the p53 signaling pathway may be influenced by CRFR1 in U87
cells. The results of the current study demonstrated that the
expression of p53 mRNA and protein were significantly increased in
U87 cells treated with hCRF (10−7 and 10−8
mol/l). It is therefore possible that hCRF may inhibit
proliferation and induce cell cycle arrest and apoptosis in U87
glioma cells in a CRFR1-mediated p53-dependent mechanism. Notably,
in a phase 1 clinical trial, patients who received hCRF reported
improved neurological symptoms and physical results, with little or
no toxicity (17).
In conclusion, the results of the present study
suggest that hCRF may inhibit cell proliferation and apoptosis in
U87 cells via a CRFR1-mediated p53 signaling pathway. Data from
previous studies and the present study demonstrate that hCRF is
innocuous in humans and that CRFR1 may be a key regulator of human
glioma, suggesting it is a novel and attractive therapeutic target
for anticancer therapy.
Acknowledgments
Not applicable.
Funding
The present study was supported by a grant from the
Outstanding medical talents plan of 2015 Hebei Province (grant no.
361004).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YF performed the experiments. XL and QW assisted the
first author in tumor specimen collection, IHC staining, cell
culture and related experiments (cell proliferation, flow
cytometry, RNA extraction, RT-qPCR, western blotting). FH and HZ
analyzed the results and data. LW was responsible for designing the
study, summarizing the results and writing the article. XS
performed the analysis and offered assistance to the corresponding
author in the manuscript writing and its English-language
editing.
Ethics approval and consent to
participate
Data collection and analysis were approved by the
ethics committee of The Second Affiliated Hospital of Hebei Medical
University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stupp R, Tonn JC, Brada M and
Pentheroudakis G; ESMO Guidelines Working Group: High-grade
malignant glioma: ESMO Clinical Practice Guidelines for diagnosis,
treatment and follow-up, : Ann Oncol. 21 Suppl:S190–S193. 2010.
View Article : Google Scholar
|
|
2
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen R, Lewis KA, Perrin MH and Vale WW:
Expression cloning of a human corticotropin-releasing-factor
receptor. Proc Natl Acad Sci USA. 90:8967–8971. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Perrin M, Donaldson C, Chen R, Blount A,
Berggren T, Bilezikjian L, Sawchenko P and Vale W: Identification
of a second corticotropin-releasing factor receptor gene and
characterization of a cDNA expressed in heart. Proc Natl Acad Sci
USA. 92:2969–2973. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dautzenberg FM and Hauger RL: The CRF
peptide family and their receptors: Yet more partners discovered.
Trends Pharmacol Sci. 23:71–77. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Audhya T, Jain R and Hollander CS:
Receptor-mediated immuno-modulation by corticotropin-releasing
factor. Cell Immunol. 134:77–84. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Clifton VL, Owens PC, Ribinson PJ and
Smith R: Identification and characterization of a
corticotropin-releasing hormone receptor in human placenta. Eur J
Endocrinol. 133:591–597. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Roe SY, McGowan EM and Rothwell NJ:
Evidence for the involvement of corticotropin-releasing hormone in
the pathogenesis of traumatic brain injury. Eur J Neurosci.
10:553–559. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Strijbos PJ, Relton JK and Rothwell NJ:
Corticotropinreleasing factor antagonist inhibits neuronal damage
induced by focal cerebral ischaemia or activation of NMDA receptors
in the rat brain. Brain Res. 656:405–408. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lezoualc'h F, Engert S, Berning B and Behl
C: Corticotropinreleasing hormone-mediated neuroprotection against
oxidative stress is associated with the increased release of
non-amyloidogenic amyloid b precursor protein and with the
suppression of nuclear factor-jB. Mol Endocrinol. 14:147–159. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baram TZ and Hatalski CG:
Neuropeptide-mediated excitability: A key triggering mechanism for
seizure generation in the developing brain. Trends Neurosci.
21:471–476. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gutknecht E, Hauger RL, Van der Linden I,
Vauquelin G and Dautzenberg FM: Expression, binding, and signaling
properties of CRF2 (a) receptors endogenously expressed in human
retinoblastoma Y79 cells: Passage-dependent regulation of
functional receptors. J Neurochem. 104:926–936. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schoeffter P, Feuerbach D, Bobirnac I,
Gazi L and Longato R: Functional, endogenously expressed
corticotropin-releasing factor receptor type 1 (CRF1) and CRF1
receptor mRNA expression in human neuroblastoma SH-SY5Y cells.
Fundam Clin Pharmacol. 13:484–489. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sato H, Nagashima Y, Chrousos GP,
Ichihashi M and Funasak Y: The expression of
corticotropin-releasing hormone in melanoma. Pigment Cell Res.
15:98–103. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Funasaka Y, Sato H, Chakraborty AK, Ohashi
A, Chrousos GP and Ichihashi M: Expression of proopiomelanocortin,
corticotropinreleasing hormone (CRH), and CRH receptor in melanoma
cells, nevus cells, and normal human melanocytes. J Investig
Dermatol Symp Proc. 4:105–109. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Graziani G, Tentori L, Muzi A, Vergati M,
Tringali G, Pozzoli G and Navarra P: Evidence that
corticotropin-releasing hormone inhibits cell growth of human
breast cancer cells via the activation of CRH-R1 receptor subtype.
Mol Cell Endocrinol. 264:44–49. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Moliterno JA, Henry E and Pannullo SC:
Corticorelin acetate injections for the treatment of peritumoral
brain edema. Expert Opin Investig Drugs. 18:1413–1419. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Muleci A, Cervoni L and Delfini R:
Medulloblastomas in children and in adults:a comparative study.
Acta Neurochir (Wien). 119:62–67. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Roberts RO, Lynch CF, Jones MP and Hart
MN: Medulloblastoma: A population-based study of 532 cases. J
Neuropathol Exp Neurol. 50:134–144. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sarkar C, Pramanik P, Karak AK,
Mukhopadhyay P, Sharma MC, Singh VP and Mehta VS: Are childhood and
adult medulloblastomas different? A comparative study of
clinicopathological features, proliferation index and apoptotic
index. J Neurooncol. 59:49–61. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hao J, Wang Z, Wang Y, Liang Z, Zhang X,
Zhao Z and Jiao B: Eukaryotic initiation factor 3C silencing
inhibits cell proliferation and promotes apoptosis in human glioma.
Oncol Rep. 33:2954–2962. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ohgaki H and Kleihues P: Epidemiology and
etiology of gliomas. Acta Neuropathol. 109:93–108. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Graziani G, Tentori L, Portarena I,
Barbarino M, Tringali G, Pozzoli G and Navarra P: CRH inhibits cell
growth of human endometrial adenocarcinoma cells via CRH-receptor
1-mediated activation of cAMP-PKA pathway. Endocrinology.
143:807–813. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Grigoriadis DE, Lovenberg TW, Chalmers DT,
Liaw C and De Souze EB: Characterization of corticotropin-releasing
factor receptor subtypes. Ann N Y Acad Sci. 780:60–80. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bayatti N, Zschocke J and Behl C: Brain
region-specific neuroprotective action and signaling of
corticotropin-releasing hormone in primary neurons. Endocrinology.
144:4051–4060. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Carlson KW, Nawy SS, Wei ET, Sadee W,
Filov VA, Rezsova VV, Slominski A and Quillan JM: Inhibition of
mouse melanoma cell proliferation by corticotropin-releasing
hormone and its analogs. Anticancer Res. 21:1173–1179.
2001.PubMed/NCBI
|
|
29
|
Moroz MA, Huang R, Kochetkow T, Shi W,
Thaler H, de Stanchina E, Gamez I, Ryan RP and Blasberg RG:
Comparison of corticotropin- releasing factor, dexamethasone, and
temozolomide: Treatment efficacy and toxicity in U87 and C6
intracranial gliomas. Clin Cancer Res. 17:3282–3292. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee YJ, Chung DY, Lee SJ, Jhon Ja G and
Lee YS: Enhanced radiosensitization of p53 mutant cells by
oleamide. Int J Radiat Oncol Biol Phys. 64:1466–1474. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ohgaki H, Dessen P, Jourde B, Horstmann S,
Nishikawa T, Di Patre PL, Burkhard C, Schüler D, Probst-Hensch NM,
Maiorka PC, et al: Genetic pathways to glioblastoma: A
population-based study. Cancer Res. 64:6892–6899. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu LX, Li ZH, Tao YF, Li RH, Fang F, Zhao
H, Li G, Li YH, Wang J, Feng X and Pan J: Histone acetyltransferase
inhibitor II induces apoptosis in glioma cell lines via the p53
signaling pathway. J Exp Clin Cancer Research. 33:1082014.
View Article : Google Scholar
|
|
33
|
Zhao Y, Wang MY, Hao K, Chen XQ and Du JZ:
CRHR1 mediates p53 transcription induced by high altitude hypoxia
through ERK 1/2 signaling in rat hepatic cells. Peptides. 44:8–14.
2013. View Article : Google Scholar : PubMed/NCBI
|