Introduction
Colorectal cancer (CRC) is one of the most prevalent
causes of cancer-associated mortality worldwide (1,2).
Clinically, 5-fluorouracil (5-FU)-based chemotherapy serves as the
initial first-line chemotherapeutic drug of choice in patients with
CRC; however, the response rates to 5-FU are ~15% in patients with
advanced CRC (3). Although novel
therapeutic approaches, including combination chemotherapy of 5-FU
and oxaliplatin (FOLFOX) or irinotecan (FOLFIRI), have presented
promising potential, the majority of the patients continue to
exhibit inadequate responses (4,5). The
failure of chemotherapy in CRC patients is primarily attributed to
therapeutic resistance following a long period of treatment
(6). These observations emphasize
that chemotherapeutic resistance is a major obstacle in the
treatment of CRC, and that the molecular mechanisms underlying this
issue remain unclear.
Chemotherapeutic resistance poses a major challenge
in cancer chemotherapy. Chemoresistance of tumor cells develops
primarily due to acquired resistance de novo, following an
initially sensitive response, and/or in combination with
pre-established cell-intrinsic mechanisms (7). A number of well-established mechanisms
are reported to be involved in cancer drug resistance, including
failure of the drug to reach or enter the target cell, formation of
efflux pumps on the surface of tumor cells, increased expression of
anti-apoptotic proteins, induction of drug-detoxifying mechanisms
and upregulation of DNA repair enzymes (8,9). However,
there is an absence of integral understanding of acquired drug
resistance in the context of CRC. Such insight may be crucial for
the development of an effective therapeutic approach to overcome
chemoresistance and to improve clinical therapeutic response in
CRC.
MicroRNAs (miRNAs/miRs) are small non-coding RNAs
composed of 19–25 nucleotides, which are involved in numerous
biological processes, including survival, apoptosis, the cell cycle
and gene regulation (10,11). miRNAs regulate downstream target gene
expression at a post-transcriptional level by binding with specific
sequences in the 3′-untranslated region (3′-UTR) of downstream
target genes, leading to mRNA degradation and/or translational
inhibition (12). Several lines of
evidence have indicated that aberrant expression of miRNAs is
involved in the progression and metastasis of different types of
cancer (13–18). Furthermore, research has indicated
that upregulation or downregulation of a certain miRNA (let-7a) was
directly correlated with the response to chemotherapeutic agents
(19). Emerging evidence has
demonstrated that miRNAs are key regulators in the chemotherapeutic
resistance of CRC. Ectopic expression of miR-125a/b restored
paclitaxel sensitivity through downregulation of aldehyde
dehydrogenase 1 in HT29 cells (20).
Furthermore, Zhang et al (21)
revealed that miR-587 promoted the resistance of CRC cells to 5-FU
via inhibiting protein phosphatase 2 scaffold subunit Aβ. Notably,
silencing miR-587 re-sensitized CRC cells to 5-FU. These
observations indicated that miRNAs may function as
tumor-suppressors or inducers in CRC.
The present study demonstrated that let-7f-5p
expression was elevated in chemotherapy-resistant CRC tissues
compared with chemotherapy-sensitive tissues. Furthermore,
upregulating let-7f-5p increased the expression of the
anti-apoptotic proteins, B-cell lymphoma 2 (Bcl-2) and B-cell
lymphoma-extra large (Bcl-xL), and decreased the activity of
caspase-3 and caspase-9 in CRC cells. Conversely, downregulating
let-7f-5p decreased the expression of Bcl-2 and Bcl-xL, and
increased the activity of caspase-3 and caspase-9 in CRC cells. Of
note, the results of the present study indicated that let-7f-5p
promotes chemotherapy-resistance via directly repressing the
expression of several pro-apoptotic proteins, including tumor
protein p53 (TP53), TP53-inducible nuclear protein 1,
TP53-inducible nuclear protein 2 and caspase-3. Therefore, the
present study identified a novel mechanism by which let-7f-5p
enhances the resistance of CRC cells to chemotherapeutics,
indicating that therapy targeting let-7f-5p in combination with
chemotherapeutics may become an effective therapeutic strategy
against CRC.
Materials and methods
The Cancer Genome Atlas (TCGA)
analysis
The miRNA dataset of colorectal cancer was
downloaded from the TCGA and obtained the expression value(s) of
the corresponding genes from the Level 3 data of each sample. The
unit of miRNA expression levels in TCGA is Reads per kilobase of
exon model per million mapped reads (RKPM). Analysis of the log2
value of each sample was performed using Excel 2010 and GraphPad 5
(GraphPad Software, Inc., La Jolla, CA, USA). Furthermore,
statistical analysis of the miRNA expression levels within
colorectal cancer tissues was performed using paired t-test or
unpaired t-test.
Cell lines and cell culture
The human CRC HCT116 and SW480 cell lines were
obtained from Type Culture Collection Chinese Academy of Sciences
(Shanghai, China) and were cultured in RPMI-1640 medium (Thermo
Fisher Scientific, Inc., Waltham, MA, USA), supplemented with
penicillin G (100 U/ml), streptomycin (100 mg/ml) and 10% fetal
bovine serum (Thermo Fisher Scientific, Inc.). HCT116 and SW480
cells were incubated in 10 µmol/l 5-FU for 24 h, as previously
described (22,23). The cells were incubated at 37°C in a
humidified atmosphere with 5% CO2 and were routinely
sub-cultured using 0.25% (w/v) trypsin-ethylenediaminetetraacetic
acid solution (Thermo Fisher Scientific, Inc.).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using an RNA
Isolation kit (Qiagen, Inc., Valencia, CA, USA), according to the
manufacturer's protocol. mRNA and microRNA (miRNA) were reverse
transcribed from total RNA using the RevertAid First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. cDNA was amplified and quantified on the
CFX96 system (Bio-Rad Laboratories, Inc., Hercules, CA, USA) using
iQ SYBR-Green (Bio-Rad Laboratories, Inc.). The thermocycling
conditions were as follows: 95°C for 3 min, then 35 cycles of 95°C
for 5 sec, 60°C for 15 sec, and 72°C for 15 sec followed by 72°C
for 10 min. The primer sequences are provided in Table I. Primers for U6 and let-7f-5p were
synthesized and purified by Guangzhou RiboBio Co., Ltd. (Guangzhou,
China). U6 or GAPDH were used as endogenous controls. Relative fold
expression was calculated using the 2−ΔΔCq method
(24).
 | Table I.Primers used in reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used in reverse
transcription-quantitative polymerase chain reaction.
| Primer |
| Sequence
(5′-3′) |
|---|
| TP53 | Forward |
GCTCGACGCTAGGATCTGAC |
|
| Reverse |
GCTTTCCACGACGGTGAC |
| TP53INP1 | Forward |
TATGCTGCCCCATTTCATTT |
|
| Reverse |
CTGTGCATAACTCCTGCCCT |
| TP53INP2 | Forward |
TGAAGAAGAGGCTGGAGAGG |
|
| Reverse |
CTCCCAGCTGTTTGGATCAC |
| Caspase-3 | Forward |
TCGCTTCCATGTATGATCTTTG |
|
| Reverse |
CTGCCTCTTCCCCCATTCT |
| Bcl-2 | Forward |
GTGGATGACTGAGTACCTGAACC |
|
| Reverse |
AGACAGCCAGGAGAAATCAAAC |
| Bcl-xL | Forward |
GGTATTGGTGAGTCGGATCG |
|
| Reverse |
TGCTGCATTGTTCCCATAGA |
| GAPDH | Forward |
ATTCCACCCATGGCAAATTC |
|
| Reverse |
TGGGATTTCCATTGATGACAAG |
Transfection
let-7f-5p mimics, inhibitors and respective control
RNAs, including siRNA scramble and negative controls, (50 nmol/l)
were synthesized and purified by Guangzhou RiboBio Co., Ltd.
Transfection of miRNA, siRNAs and plasmids was performed using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.), 48 h after transfection, cells were harvest and
the indicated experiments were performed, according to the
manufacturer's protocol.
Western blot analysis
Nuclear/cytoplasmic fractionation was performed
using a Cell Fractionation kit (Cell Signaling Technology, Inc.,
Danvers, MA, USA) according to the manufacturer's protocol, and the
whole cell lysates were extracted using radioimmunoprecipitation
assay buffer (Cell Signaling Technology, Inc.). The cell lysates
were loaded with 10% loading buffer (Beyotime Institute of
Biotechnology, Haimen, China) and were heated for 5 min at 100°C.
The protein concentrations were determined using the BCA Protein
Assay kit (Invitrogen; Thermo Fisher Scientific, Inc.). Equal
quantities of denatured protein samples (40 µl) were resolved on
10% SDS-polyacrylamide gels, prior to being transferred onto
polyvinylidene difluoride membranes (Roche Diagnostics, Basel,
Switzerland). Following 1 h blocking with 5% dry skimmed milk in
Tris-buffered saline/0.05% Tween-20 at room temperature, membranes
were incubated with primary antibodies against Bcl-2 at a dilution
of 1:1,000 (cat. no. ab194583; Abcam, Cambridge, UK) or Bcl-xL at a
dilution of 1:1,000 (cat. no. ab32370; Abcam), overnight at 4°C,
followed by incubation with a horseradish peroxidase-conjugated
secondary antibody for 1 h at room temperature. Proteins were
visualized using Pierce™ Fast Western Blot kit (Thermo Fisher
Scientific, Inc.). The membranes were stripped and reprobed with an
anti-α-tubulin antibody at a dilution of 1:1,000 overnight at 4°C
(cat. no. T6199; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) as
the loading control.
Caspase-9 or −3 activity assays
Activity of caspase-9 or −3 was analyzed by
spectrophotometry using a caspase-9 colorimetric assay kit (cat.
no. KGA403) or acaspase-3 colorimetric assay kit (cat. no. KGA202;
Nanjing KeyGen Biotech, Co. Ltd., Nanjing, China), according to the
manufacturer's protocol. In brief, 5×106 cells were
washed with cold phosphate-buffered saline, re-suspended in Lysis
Buffer (from the two kits) and incubated on ice for 30 min. The 50
µl cell suspension was mixed with 50 µl Reaction Buffer (from the
two kits) and 5 µl caspase-3/-9 substrate, prior to being incubated
at 37°C for 4 h. The absorbance was measured at 405 nm, and
bicinchoninic acid protein quantitative analysis was used as the
reference to normalize each of the experimental groups.
Flow cytometric analysis
Flow cytometric analysis of apoptosis was performed
using the fluorescein isothiocyanate (FITC)-Annexin V Apoptosis
Detection kit I (BD Biosciences, Franklin Lakes, NJ, USA), and was
performed, according to the manufacturer's protocol. In brief,
cells were dissociated with trypsin and were resuspended at
1×106 cells/ml in binding buffer (Thermo Fisher
Scientific, Inc.) with 50 µl/ml FITC-Annexin V and 50 µl/ml
propidium iodide (PI). The cells were subsequently incubated for 15
min at room temperature, prior to being analyzed using a Gallios
flow cytometer (Beckman Coulter, Inc., Brea, CA, USA). The inner
mitochondrial membrane potential (Δψm) of the cells was detected by
flow cytometry using a MitoScreen JC-1 staining kit (cat no.
551302; BD Biosciences), according to the manufacturer's protocol.
In brief, cells were dissociated with trypsin and resuspended at
1×106 cells/ml in assay buffer (BD Biosciences), prior
to being incubated at 37°C for 15 min with 10 µl/ml JC-1. Prior to
flow cytometric analysis, cells were washed twice in assay buffer
(BD Biosciences), part of the MitoScreen JC-1 staining kit. Flow
cytometry data were analyzed using FlowJo 7.6 software (FlowJo LLC,
Ashland, OR, USA).
Luciferase assay
A total of 4×104 cells were seeded in
triplicate onto 24-well plates and were cultured at 37°C for 24 h.
Cells were transfected with 100 ng control pmirGLO,
pmirGLO-TP53-3′UTR, -TP53INP1-3′UTR, -TP53INP2-3′UTR, or
-caspase3-3′-UTR luciferase plasmid, plus 5 ng pRL-TK
Renilla plasmid (Promega Corporation, Madison, WI, USA),
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Luciferase and Renilla signals were measured 36 h after
transfection using a Dual Luciferase Reporter assay kit (Promega
Corporation) at room temperature, according to the manufacturer's
protocol.
Statistical analysis
All values are presented as the mean ± standard
deviation. Significant differences were determined using GraphPad
5.0 software (GraphPad Software, Inc., La Jolla, CA, USA). The
Mann-Whitney U test and χ2 tests were used to determine
statistical differences between two groups. One-way analysis of
variance, followed by the Student-Newman-Keuls test, was used to
determine statistical differences among multiple groups. P<0.05
was considered to indicate a statistically significant difference.
All the experiments were repeated three times.
Results
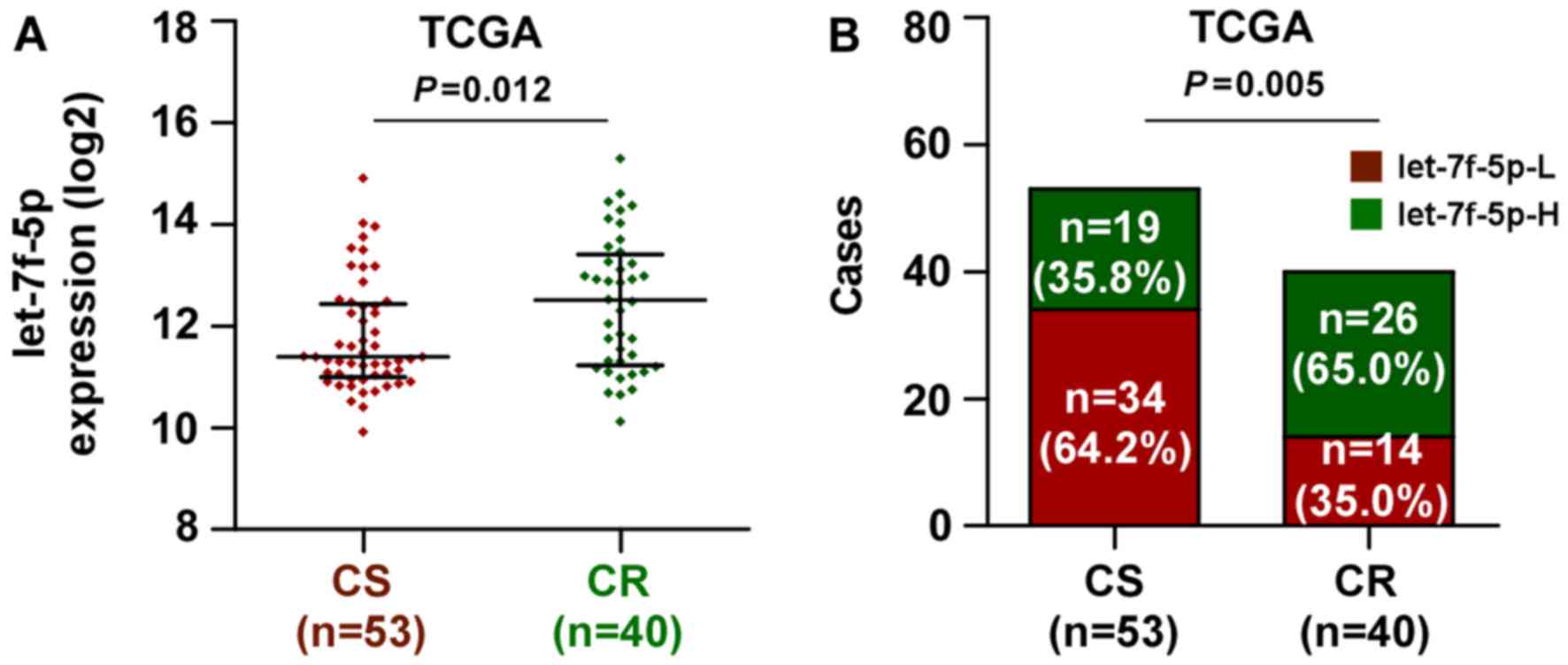
let-7f-5p is upregulated in
chemoresistant CRC
The miRNA sequencing datasets of CRC were analyzed
using TCGA, which identified that let-7f-5p expression was elevated
in chemoresistant CRC tissues, compared with chemosensitive tissues
(Fig. 1A). Notably, the present study
demonstrated that the expression level of let-7f-5p was upregulated
in patients with CRC exhibiting poor chemotherapeutic response,
compared with those with a positive chemotherapeutic response
(Fig. 1B). These observations
revealed that let-7f-5p may contribute toward chemoresistance in
CRC.
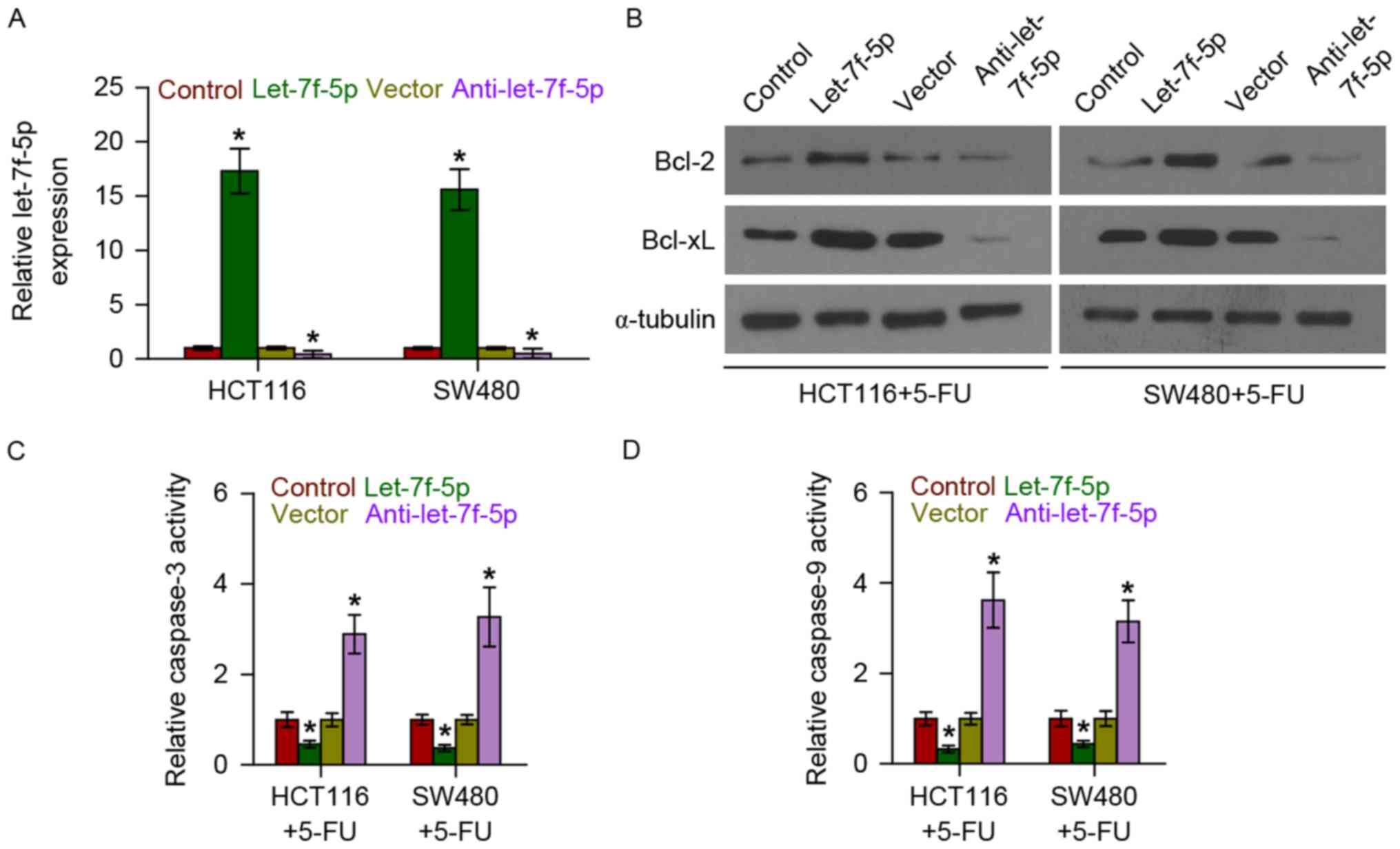
Overexpression of let-7f-5p increases
the expression of anti-apoptotic proteins Bcl-2 and Bcl-xL in CRC
cells
The role of let-7f-5p in the chemoresistance of CRC
was additionally examined following treatment with the first-line
drug 5-FU. let-7f-5p was upregulated and downregulated following
transfection with let-7f-5p mimics and inhibitors, respectively, in
CRC HCT116 and SW480 cells (Fig. 2A).
The effect of let-7f-5p on the expression of anti-apoptotic
proteins, Bcl-2 and Bcl-xL, was further examined. The results
demonstrated that upregulating let-7f-5p increased Bcl-2 and Bcl-xL
expression, while downregulating let-7f-5p decreased their
expression (Fig. 2B). Therefore,
these results demonstrated that let-7f-5p promoted the
chemoresistance of CRC cells by enhancing the expression of
anti-apoptotic proteins, Bcl-2 and Bcl-xL.
Overexpression of let-7f-5p decreases
the activity of caspase-3 and caspase-9 in CRC cells
The effect of let-7f-5p on the activity of caspase-3
and-9 was examined. As demonstrated in Fig. 2C and D, upregulating let-7f-5p
decreased the activity of caspase-3, while silencing let-7f-5p had
the opposite effect. Overexpression of let-7f-5p consistently
repressed the activity of caspase-9, while silencing of let-7f-5p
increased the activity of caspase-9. Therefore, these observations
suggested that let-7f-5p promotes the chemoresistance of CRC cells
by inhibiting the activity of caspase-3 and caspase-9 in CRC
cells.
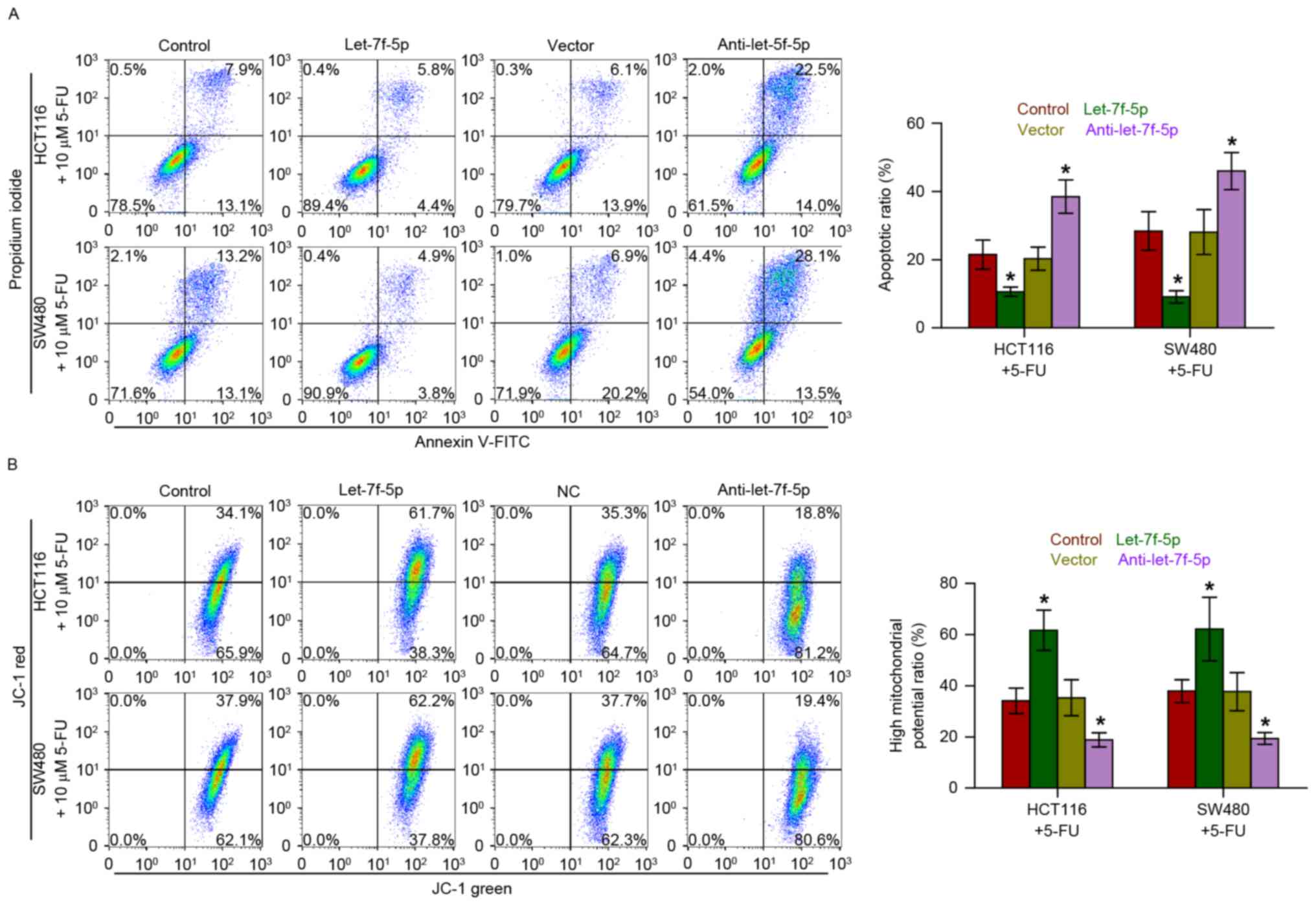
Overexpression of let-7f-5p decreases
the apoptotic rate and increases the mitochondrial potential of CRC
cells
Annexin V-FITC/PI staining demonstrated that
overexpression of let-7f-5p decreased the apoptotic rate of HCT116
and SW480 cells treated with 5-FU, while silencing let-7f-5p had
the opposite effect (Fig. 3A).
Furthermore, the effect of let-7f-5p on the mitochondrial potential
of CRC cells was examined, and the results indicated that
upregulating let-7f-5p enhanced the mitochondrial potential of
HCT116 and SW480 cells following treatment with 5-FU; however,
silencing let-7f-5p exhibited the opposite effects on the
mitochondrial potential of CRC cells (Fig. 3B). Therefore, these observations
demonstrated that let-7f-5p promotes the chemoresistance of CRC
cells to 5-FU by decreasing the apoptotic rate and increasing the
mitochondrial potential of HCT116 and SW480 cells.
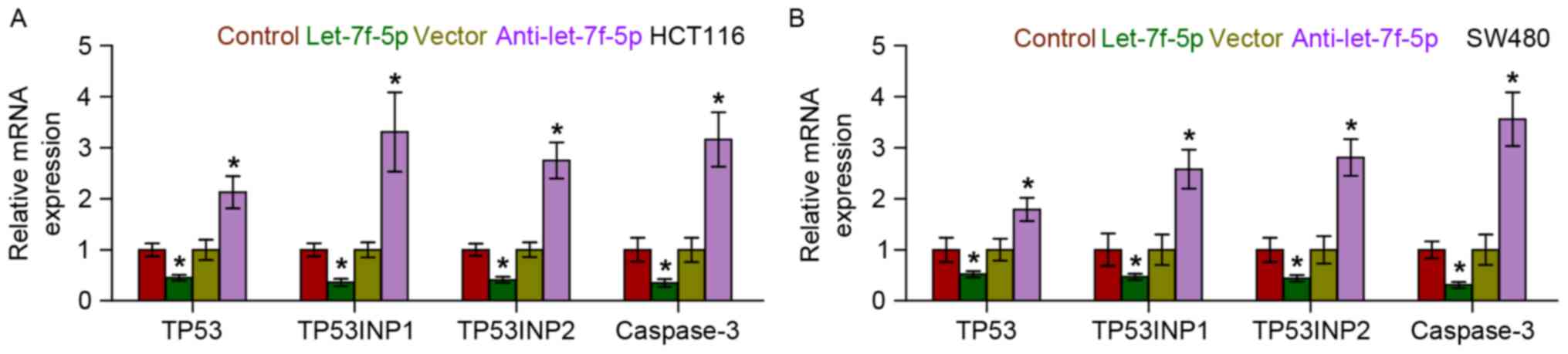
let-7f-5p targets multiple
pro-apoptotic proteins in CRC cells
Using the publicly available algorithms, TargetScan
and miRanda, it was identified that multiple pro-apoptotic
proteins, including TP53, TP53INP1, TP53INP2 and caspase-3, may be
potential targets of let-7f-5p (Fig.
4). RT-qPCR analysis revealed that let-7f-5p overexpression
reduced the mRNA expression levels of TP53, TP53INP1, TP53INP2 and
caspase-3. By contrast, silencing of let-7f-5p increased the
expression of TP53, TP53INP1, TP53INP2 and caspase-3, suggesting
that let-7f-5p negatively regulated the mRNA expression levels of
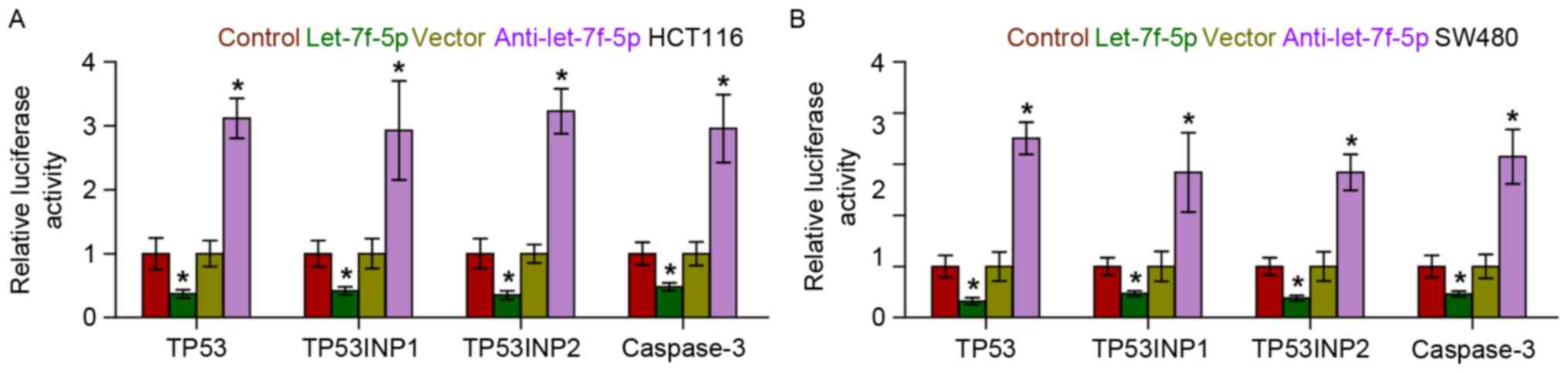
TP53, TP53INP1, TP53INP2 and caspase-3 (Fig. 5). Furthermore, a luciferase assay
revealed that let-7f-5p overexpression decreased the reporter
activity driven by the 3′-UTRs of these transcripts, while
silencing let-7f-5p increased this activity (Fig. 6). Taken together, the results of the
present study indicated that let-7f-5p directly targets TP53,
TP53INP1, TP53INP2 and caspase-3, which further promotes
chemotherapeutic resistance in CRC.
Discussion
The results of the present study demonstrated that
let-7f-5p expression was elevated in chemotherapy-resistant CRC
tissues, compared with chemotherapy-sensitive tissues. Furthermore,
upregulating let-7f-5p increased the expression of the
anti-apoptotic proteins, Bcl-2 and Bcl-xL, and decreased the
activity of caspase-3 and caspase-9 in CRC cells, while
downregulating let-7f-5p had the opposite effect. Notably, the
observations of the present study indicated that let-7f-5p promotes
chemotherapy resistance via directly repressing the expression of a
number of pro-apoptotic proteins, including TP53, TP53INP1,
TP53INP2 and caspase-3. Therefore, the results of the present study
indicate that overexpression of let-7f-5p exhibits an important
function in the chemoresistance of CRC.
The transcription factor, p53 is the most
characterized tumor suppressor gene and has been reported to be
involved in the development and progression of several types of
cancer via controlling cell cycle checkpoints, and promoting
apoptosis and senescence (25,26). Loss
or mutation of the p53 gene has been identified to be positively
associated with the recurrent incidence of chemotherapeutic
resistance in different types of cancer (27). In CRC, p53 is mutated in ~50% of
patients and these mutations of are considered to be inactivating,
leaving to TP53 being incapable of regularly exerting its
functions, including its pro-apoptotic role (28,29). The
present study identified that let-7f-5p decreased the apoptosis of
CRC cells induced by 5-FU and conferred the resistance of these
cells to 5-FU via inhibiting TP53 expression. Furthermore, the
results further demonstrated that let-7f-5p simultaneously targets
TP53INP1, TP53INP2 and caspase 3, which cooperatively attenuated
the apoptosis induced by 5-FU and promoted chemoresistance in CRC
cells. TP53INP1 and TP53INP2, which act as pro-apoptotic and
anti-proliferative proteins, have been reported to positively
regulate p53, and to stimulate its ability to induce apoptosis and
to regulate the cell cycle (30). A
previous study reported that miR-182 increased drug resistance in
cisplatin-treated hepatocellular carcinoma cells via inhibiting
TP53INP1 expression (31). Caspase-3
has been reported to be involved in the activation cascade of
caspases responsible for apoptosis, and functions by cleaving and
activating caspases-6, −7 and −9, which serve important roles in
the chemoresistance of different types of cancer (32–34).
Through the use of luciferase assays, the present study identified
that let-7f-5p simultaneously repressed the activity of TPp53,
TP53INP1, TP53INP2 and caspase-3 by binding to the 3′-UTRs of their
mRNAs, resulting in a decrease in the apoptosis induced by 5-FU in
CRC cells and the development of chemoresistance.
Members of the Let-7 family have been identified to
function as tumor suppressors through regulating multiple oncogenic
signals (35). For example, Di Fazio
et al (36) reported that
let-7b exhibited an important tumor-suppressive role via inhibiting
the expression of high-mobility group AT-hook 2, a nuclear
non-histone transcriptional co-factor with known oncogenic
properties, in liver cancer cell lines. Furthermore, emerging
evidence has demonstrated that members of the let-7 family of
miRNAs are downregulated in a number of types of human cancer, and
that low let-7 expression has been correlated with resistance to
chemotherapeutics (37,38). Downregulation of let-7 was also
associated with chemoresistance in breast cancer cells (39). In addition, Sugimura et al
(38) reported that low expression of
let-7b and let-7c was significantly associated with a poor response
to chemotherapy in esophageal squamous cell carcinoma, and that
transfection of let-7c restored sensitivity to cisplatin and
increased the rate of apoptosis following exposure to cisplatin
in vitro assays. Additionally, Tsang and Kwok (34) reported that induced expression of
let-7a increased the resistance of human squamous carcinoma A431
cells to chemotherapeutic drugs, including interferon-γ,
doxorubicin and paclitaxel. These observations revealed that the
members of the let-7 family may function as oncomiRs and
tumor-suppressive miRNAs, depending on the type of tumor. The
present study reported that, as a member of the let-7 family,
let-7f-5p was elevated in chemoresistant CRC tissues, compared with
chemosensitive tissues. Upregulation of let-7f-5p decreased the
apoptosis induced by 5-FU in vitro assays, while silencing
let-7f-5p had the opposite effect. These results, in conjunction
with those of other studies, indicated that the pro-cancer and
anticancer roles of the let-7 family are environmentally and
tumor-type dependent.
Previous studies have implicated the altered
expression of let-7f-5p in several diseases, including major
depression, myasthenia gravis, Alzheimer's disease (40–42), and
neoplastic conditions, including laryngeal carcinoma, CRC and
thyroid cancer (43,44). Notably, Pathak et al (45) reported that let-7f-5p was highly
upregulated in the human colon cancer HCT116 and SN38 cell lines
following radiation treatment in a p53-directed manner, indicating
that let-7f-5p may contribute to the chemoresistance of CRC.
However, the specific role of let-7f-5p in CRC remains unclear. The
results of the present study also demonstrated that let-7f-5p
expression was elevated in chemoresistant CRC tissues compared with
chemosensitive tissues. Furthermore, ectopic expression of
let-7f-5p had the effect of reducing the apoptosis induced by 5-FU,
while silencing let-7f-5p enhanced the apoptosis induced by 5-FU.
Additionally, silencing let-7f-5p enhanced sensitivity to 5-FU.
Importantly, the results demonstrated that let-7f-5p overexpression
promotes chemotherapeutic resistance in CRC via directly repressing
the expression of several pro-apoptotic proteins, including TP53,
TP53INP1, TP53INP2 and caspase-3.
In summary, the results of the present study
revealed that let-7f-5p serves an important role in the
chemotherapy resistance of CRC via directly repressing expression
of TP53, TP53INP1, TP53INP2 and caspase-3. Therefore, improved
understanding of the specific role of let-7f-5p in the pathogenesis
of CRC would facilitate an increase in the available knowledge of
CRC development, which will assist in the development of novel
therapeutic measures against CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Gansu Province (Lanzhou, China; grant no.
096RJZA096).
Availability of data and materials
The datasets generated and analyzed in the present
study are included in this published article.
Authors' contributions
YT and CC developed ideas and drafted the
manuscript. HY and HW conducted the experiments and contributed to
the analysis of data. HT and YP contributed to the analysis of
data. BL and XD contributed to the analysis of data and revised the
manuscript. YY and ZQ analysed and interpreted the data and agreed
to be accountable for all aspects of the work in ensuring that
questions related to the accuracy or integrity of any part of the
work are appropriately investigated and resolved. All authors
contributed to revise the manuscript and approved the final version
for publication.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing
interests.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ricci-Vitiani L, Fabrizi E, Palio E and De
Maria R: Colon cancer stem cells. J Mol Med (Berl). 87:1097–1104.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnston PG and Kaye S: Capecitabine: A
novel agent for the treatment of solid tumors. Anticancer Drugs.
12:639–646. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Giacchetti S, Perpoint B, Zidani R, Le
Bail N, Faggiuolo R, Focan C, Chollet P, Llory JF, Letourneau Y, et
al: Phase III multicenter randomized trial of oxaliplatin added to
chronomodulated fluorouracil-leucovorin as first-line treatment of
metastatic colorectal cancer. J Clin Oncol. 18:136–147. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Douillard JY, Cunningham D, Roth AD,
Navarro M, James RD, Karasek P, Jandik P, Iveson T, Carmichael J,
Alakl M, et al: Irinotecan combined with fluorouracil compared with
fluorouracil alone as first-line treatment for metastatic
colorectal cancer: A multicenter randomised trial. Lancet.
355:1041–1047. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Longley DB and Johnston PG: Molecular
mechanisms of drug resistance. J Pathol. 205:275–292. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gonzalez-Angulo AM, Morales-Vasquez F and
Hortobagyi GN: Overview of resistance to systemic therapy in
patients with breast cancer. Adv Exp Med Biol. 608:1–22. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gottesman MM: Mechanisms of cancer drug
resistance. Annu Rev Med. 53:615–627. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu X and Xiao H: MiRNAs modulate the drug
response of tumor cells. Sci China C Life Sci. 9:797–801. 2009.
View Article : Google Scholar
|
|
10
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ren D, Wang M, Guo W, Huang S, Wang Z,
Zhao X, Du H, Song L and Peng X: Double-negative feedback loop
between ZEB2 and miR-145 regulates epithelial-mesenchymal
transition and stem cell properties in prostate cancer cells. Cell
Tissue Res. 358:763–778. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo W, Ren D, Chen X, Tu X, Huang S, Wang
M, Song L, Zou X and Peng X: HEF1 promotes epithelial mesenchymal
transition and bone invasion in prostate cancer under the
regulation of microRNA-145. J Cell Biochem. 114:1606–1615. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Simerzin A, Zorde-Khvalevsky E, Rivkin M,
Adar R, Zucman-Rossi J, Couchy G, Roskams T, Govaere O, Oren M,
Giladi H and Galun E: The liver-specific microRNA-122*, the
complementary strand of microRNA-122, acts as a tumor suppressor by
modulating the p53/mouse double minute 2 homolog circuitry.
Hepatology. 64:1632–1636. 2016. View Article : Google Scholar
|
|
16
|
Ren D, Wang M, Guo W, Zhao X, Tu X, Huang
S, Zou X and Peng X: Wild-type p53 suppresses the
epithelial-mesenchymal transition and stemness in PC-3 prostate
cancer cells by modulating miR-145. Int J Oncol. 42:1473–1481.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang M, Ren D, Guo W, Wang Z, Huang S, Du
H, Song L and Peng X: Loss of miR-100 enhances migration, invasion,
epithelial-mesenchymal transition and stemness properties in
prostate cancer cells through targeting Argonaute 2. Int J Oncol.
45:362–372. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sahu N, Stephan JP, Cruz DD, Merchant M,
Haley B, Bourgon R, Classon M and Settleman J: Functional screening
implicates miR-371-3p and peroxiredoxin 6 in reversible tolerance
to cancer drugs. Nat Commun. 7:123512016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K,
Guo J, Zhang Y, Chen J, Guo X, et al: Characterization of microRNAs
in serum: A novel class of biomarkers for diagnosis of cancer and
other diseases. Cell Res. 18:997–1006. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen J, Chen Y and Chen Z: MiR-125a/b
regulates the activation of cancer stem cells in
paclitaxel-resistant colon cancer. Cancer Invest. 31:17–23. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Y, Talmon G and Wang J: MicroRNA-587
antagonizes 5-FU-induced apoptosis and confers drug resistance by
regulating PPP2R1B expression in colorectal cancer. Cell Death Dis.
6:e18452015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang X, Chen Y, Hao L, Hou A, Chen X, Li
Y, Wang R, Luo P, Ruan Z, Ou J, et al: Macrophages induce
resistance to 5-fluorouracil chemotherapy in colorectal cancer
through the release of putrescine. Cancer Lett. 381:305–313. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Du C, Huang D, Peng Y, Yao Y, Zhao Y, Yang
Y, Wang H, Cao L, Zhu WG and Gu J: 5-Fluorouracil targets histone
acetyltransferases p300/CBP in the treatment of colorectal cancer.
Cancer Lett. 400:183–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-(delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vousden KH and Prives C: Blinded by the
light: The growing complexity of p53. Cell. 137:413–431. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Goh AM, Coffill CR and Lane DP: The role
of mutant p53 in human cancer. J Pathol. 223:116–126. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alam SK, Yadav VK, Bajaj S, Datta A, Dutta
SK, Bhattacharyya M, Bhattacharya S, Debnath S, Roy S, Boardman LA,
et al: DNA damage-induced ephrin-B2 reverse signaling promotes
chemoresistance and drives EMT in colorectal carcinoma harboring
mutant p53. Cell Death Differ. 23:707–22. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Deschoemaeker S, Di Conza G, Lilla S,
Martín-Pérez R, Mennerich D, Boon L, Hendrikx S, Maddocks OD, Marx
C, Radhakrishnan P, et al: PHD1 regulates p53-mediated colorectal
cancer chemoresistance. EMBO Mol Med. 7:1350–1365. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Muller PA and Vousden KH: p53 mutations in
cancer. Nat Cell Biol. 15:2–8. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Okamura S, Arakawa H, Tanaka T, Nakanishi
H, Ng CC, Taya Y, Monden M and Nakamura Y: p53DINP1, a
p53-inducible gene, regulates p53-dependent apoptosis. Mol Cell.
8:85–94. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qin J, Luo M, Qian H and Chen W:
Upregulated miR-182 increases drug resistance in cisplatin-treated
HCC cell by regulating TP53INP1. Gene. 538:342–347. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Harrington HA, Ho KL, Ghosh S and Tung KC:
Construction and analysis of a modular model of caspase activation
in apoptosis. Theor Biol Med Model. 5:262008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sidi S, Sanda T, Kennedy RD, Hagen AT,
Jette CA, Hoffmans R, Pascual J, Imamura S, Kishi S, Amatruda JF,
et al: Chk1 suppresses a caspase-2 apoptotic response to DNA damage
that bypasses p53, Bcl-2, and caspase-3. Cell. 133:864–877. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tsang WP and Kwok TT: Let-7a microRNA
suppresses therapeutics-induced cancer cell death by targeting
caspase-3. Apoptosis. 13:1215–1222. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chang CJ, Hsu CC, Chang CH, Tsai LL, Chang
YC, Lu SW, Yu CH, Huang HS, Wang JJ, Tsai CH, et al: Let-7d
functions as novel regulator of epithelial-mesenchymal transition
and chemoresistant property in oral cancer. Oncol Rep.
26:1003–1010. 2011.PubMed/NCBI
|
|
36
|
Di Fazio P, Montalbano R, Neureiter D,
Alinger B, Schmidt A, Merkel AL, Quint K and Ocker M:
Downregulation of HMGA2 by the pan-deacetylase inhibitor
panobinostat is dependent on hsa-let-7b expression in liver cancer
cell lines. Exp Cell Res. 318:1832–1843. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu C, Xie S, Song C, Huang L and Jiang Z:
Lin28 mediates cancer chemotherapy resistance via regulation of
miRNA signaling. Hepatogastroenterology. 61:1138–1141.
2014.PubMed/NCBI
|
|
38
|
Sugimura K, Miyata H, Tanaka K, Hamano R,
Takahashi T, Kurokawa Y, Yamasaki M, Nakajima K, Takiguchi S, Mori
M and Doki Y: Let-7 expression is a significant determinant of
response to chemotherapy through the regulation of IL-6/STAT3
pathway in esophageal squamous cell carcinoma. Clin Cancer Res.
18:5144–5153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wu J, Li S, Jia W, Deng H, Chen K, Zhu L,
Yu F and Su F: Reduced Let-7a is associated with chemoresistance in
primary breast cancer. PLoS One. 10:e01336432015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Maffioletti E, Cattaneo A, Rosso G, Maina
G, Maj C, Gennarelli M, Tardito D and Bocchio-Chiavetto L:
Peripheral whole blood microRNA alterations in major depression and
bipolar disorder. J Affect Disord. 200:250–258. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Punga T, Bartoccioni E, Lewandowska M,
Damato V, Evoli A and Punga AR: Disease specific enrichment of
circulating let-7 family microRNA in MuSK+ myasthenia gravis. J
Neuroimmunol. 292:21–26. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Satoh J, Kino Y and Niida S: MicroRNA-Seq
data analysis pipeline to identify blood biomarkers for Alzheimer's
disease from public data. Biomark Insights. 10:21–31. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Damanakis AI, Eckhardt S, Wunderlich A,
Roth S, Wissniowski TT, Bartsch DK and Di Fazio P: MicroRNAs let7
expression in thyroid cancer: Correlation with their deputed
targets HMGA2 and SLC5A5. J Cancer Res Clin Oncol. 142:1213–1220.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lu ZM, Lin YF, Jiang L, Chen LS, Luo XN,
Song XH, Chen SH and Zhang SY: Micro-ribonucleic acid expression
profiling and bioinformatic target gene analyses in laryngeal
carcinoma. Onco Targets Ther. 7:525–533. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pathak S, Meng WJ, Nandy SK, Ping J,
Bisgin A, Helmfors L, Waldmann P and Sun XF: Radiation and SN38
treatments modulate the expression of microRNAs, cytokines and
chemokines in colon cancer cells in a p53-directed manner.
Oncotarget. 6:44758–44780. 2015. View Article : Google Scholar : PubMed/NCBI
|