Introduction
Breast cancer is the most commonly diagnosed cancer
and the leading cause of cancer-associated mortality among women
worldwide (1). In 2012, ~2 million
females globally were diagnosed with breast cancer (2). In the USA, the American Cancer Society
estimates diagnoses of approximately 2,350 new men cases and around
231,000 new cases of invasive breast cancer among women in 2015
(3). As a result, breast cancer
prevention and therapeutic intervention are significant objectives
for a number of researchers. These measures require an
understanding of the biology of the disease during its development
and progression. To gain this knowledge, basic clinical research is
necessary. Due to the scarcity of human tissue samples and the
ethical issues regarding conducting such research in humans,
researchers require in vivo and in vitro human breast
cancer (HBC) models (1). The most
commonly used in vitro model are cell lines; additionally,
an organotypic culture model has been recently developed. While
cell lines are the most widely used model for pre-clinical
research, they have limitations, which make them poorly
representative of real cancers (4).
In vivo models include xenografts, as well as syngeneic and
genetically engineered mice, cats and dogs (5). Rodent models are considered the most
useful model of breast cancer, but they also have limitations
(5). This review will show how dogs,
according to several aspects and studies is considered the optimal
model for human breast cancer.
Cell lines as a model for HBC
Established breast cancer cell lines are used as
in vitro models in the majority of breast cancer research
laboratories. There are a number of advantages to their use,
including: Cell homogeneity, low cost, uncomplicated handling, and
straightforward removal from frozen stocks. In addition, cell lines
replicate indefinitely. Several breast cancer cell lines are
hormone-dependent, which allows for the analysis of estrogen and
progesterone-regulated signaling pathways in breast cancer. MCF-7
is considered to be an ideal model for hormone response studies due
to its particular hormone sensitivity as a result of the expression
of estrogen receptors (ER) (6). In
addition, the effects of the chemical inhibition of signaling
pathways and altered gene function on the tumorigenicity of the
cells may be examined by transplanting them as xenografts into
appropriate, immunocompromised animals. However, breast cancer cell
line usage has a number of disadvantages, which include: A
propensity for genotypic drift; and phenotypic selection due to the
continuous culturing of rapidly growing, specific clones from the
subpopulations (7,8).
Furthermore, the culturing of breast cancer cells
from primary tumors via the extraction of viable tumor cells from
the surrounding stroma is challenging (4,9); however,
five cell lines were recently isolated from a ductal carcinoma
in situ (DCIS) excision from a Singaporean patient of
Chinese origin (10), which
represents a rich resource for cell physiology and drug discovery
studies for early stage breast cancer. In comparison to primary
tumors, metastatic tumors provide large numbers of viable, detached
tumor cells with little or no contamination from tumor stroma
cells, particularly metastatic effusions (11). Long-term culturing success is low,
even from samples isolated from metastatic tumors (4,11,12). The majority of available breast cancer
cell lines were isolated from pleural effusions (12). Other cells have been obtained from
less typical origins, including: MDA-MB-361, which originated from
a brain metastasis (12,13); HH375 and HH315, which originated from
supraclavicular and abdominal lymph node metastases, respectively
(14); LCC15-MB, which was obtained
from a femoral metastasis (15);
MALL, which originated from bone marrow aspirate (16); and MAST and ZR-75-1 cells, which were
extracted from ascitic fluid (17,18).
Furthermore, it has been demonstrated that certain breast cancer
cell lines have a tropism for certain metastatic sites (19). MA-11 cells frequently metastasize to
the brain, whereas MT-1 cells frequently metastasize to the bone
and bone marrow in mice (19).
Cell lines are considered to be relatively
unrepresentative models of the tumors from which they were
obtained, despite their essential role in the majority of aspects
of cancer biology research (20). The
majority of breast cancer lines have a metastatic origin and their
association with primary tumors is an important issue (8). In addition, the pure and clonal
population of any breast cancer cell line is expected to
inadequately reflect the presumed heterogeneity of breast tumors,
regardless of its metastatic or primary origin (8,21). This
heterogeneity reflects the tumor progression through pathological
and clinical phases, starting as an atypical hyperplasia, followed
by carcinoma in situ, invasive carcinoma, and metastatic
disease (22). These stages are
mediated by the various genetic and phenotypic changes in an
individual cell, followed by clonal selection and expansion, which
leads to intratumoral diversity (22). The culture conditions of the cells may
eliminate a number of types of cancer cells that were originally
present in the tumor samples. For example, certain cell lines are
unable to grow well on plastic and require specific tumor
environments for specific factors (23). These cells are unlikely to be included
in the panel of currently available breast cancer cell lines.
Furthermore, cancer cells are genetically unstable, and may undergo
specific genotypic and phenotypic alterations as a result of
long-term culturing and storage. Over 20 years ago, a
cross-contamination of breast cancer cell lines with HeLa cells was
identified, which had produced false cell lines (8). Accordingly, it is imperative that cell
lines are well characterized prior to research use (8). Whether or not the few frequently used
breast cancer cell lines precisely reflect intertumoral
heterogeneity has been extensively debated (20).
Studies recently established and characterized
unique, novel breast cancer cell lines, including from primary
lesions (24–26). The process of selecting cells to fit
the criteria of a bona fide continuous cell line is
intensive. The criteria include: A tendency toward
anchorage-independent growth; altered cytomorphology; increased
growth; increased clonogenicity; reduced serum dependency; changes
in ploidy; tumorigenicity in nude mice; and an unlimited lifespan
(27). Established cell lines will be
continued to be used as models for breast cancer due to their
distinct advantages. However, it is necessary that researchers are
aware of the limitations of cell lines, and the possible effects on
experiments and results. Cell lines isolated from primary breast
tumors are now available through conventional cell line
repositories and are worth consideration as research tools
(8).
Animal models of HBC
Rodent models
Mice and rats are the most important models for HBC,
and has enabled scientists to understand the fundamental events
that cause breast cancer initiation, development and progression
(28). In addition, mouse models
offer a number of opportunities for studying breast cancer
treatment and prevention (29).
Inbred mice
Inbred mice were first used as a unique model for
studying susceptibility to specific cancer types, including breast
cancer, in 1920 (30). The use of
inbred strains demonstrated that certain hormonal, environmental
and genetic factors, including mouse mammary tumor virus (MMTV),
have important roles in determining the susceptibility to breast
cancer (30). Inbred mice have a
carbon-copy genetic background, so they are expected to generate
analogous responses to similar treatments and exposures (31). Gathering data from standardized
collections around the world allows cancer researchers to
investigate virtually any phenotype of interest, and utilize the
depth and breadth of knowledge on that model system.
Transgenic and genetically engineered
mice models (GEMMs)
In 1980, GEMMs were developed through the removal of
specific tumor suppressor genes or the overexpression of specific
oncogenes, in a germline-specific manner (32). These models have been used to assess
signal transduction pathways, pathobiology and gene expression
signatures. They have also been used to evaluate the efficacy of
chemoprevention strategies (33).
Using GEMMs, researchers have the ability to examine the effect of
single or multiple genes on the evolution of tumors in the breast.
GEMMs have conditional alleles that induce the mutations necessary
for specific cancer types (34). In a
number of cases, GEMMs more faithfully, compared with other animal
models, represent the full biochemical, proteomic, genetic,
phenotypic and histological characteristic features of specific
human tumors (35). In 2007, a novel
approach using the avian leucosis virus receptor A as a transgene
targeted to the mammary gland was published (36). These models are advantageous as they
mimic the condition of initiated cells within a field of normal
cells. Furthermore, if a means to modify the vector to infect
particular subpopulations is developed, the phenotypes arising from
certain cell subtypes can be traced more accurately (36).
Carcinogen-induced mice models
Several rodent studies have demonstrated that
exposure to cancer-causing agents, including radiation or chemicals
produces specific tumors, have effects of these agents resulting in
uncontrolled cell growth due to mutations and alterations (37,38). These
models recapitulate the multi-stage and time-dependent development
of tumor pathogenesis in the reaction to tumor-promoting agents and
etiologically relevant environmental carcinogens (39). Researchers generated these models in
outbred rodent breeds of various genetic backgrounds, and reported
a high frequency of organ-specific cancer types with highly
reproducible phenotypes. Furthermore, these models have
histopathological, biochemical and molecular features that resemble
the developmental consequences of specific human tumors, which make
them clinically and biologically important (39,40).
As with traditional xenograft models developed in
syngeneic hosts and GEMMs, carcinogen-induced tumor models allow
the assessment and modulation of the function of humoral and
cellular immune components in tumor immune surveillance and evasion
mechanisms. The participating role of chronic and acute
inflammatory processes in cancer development and progress is
clearly represented in this model (41,42). The
carcinogen-induced tumor model is particularly relevant given the
epidemiological and experimental association between human tumor
development and chronic inflammation (43). In addition, these models have been
used in chemoprevention studies to assess the therapeutic effects
on tumor development and growth (29,44). A
number of experimental agents were determined to be useful in the
management or prevention of certain human tumors in their analogous
organ-specific, carcinogen-induced cancer models (45–48).
The molecular, phenotypic and histological
similarities, the general predilection for metastasis, and the
utility in inflammatory and immune response studies make these
models beneficial translational biology systems for drug discovery
and early non-clinical drug development stages.
Xenograft mice models
Currently, mice are used in xenograft experiments,
in particular severe combined immunodeficiency mice and nude mice.
These two strains naturally induce single gene mutations that
compromise the immune system. HBC cell lines are relatively easily
transplanted into these animal models. For invasive breast cancer,
the reported success is 7–20% (49).
This range depends on a number of factors, including the
implantation site, strain, hormonal supplementation and the age of
the mice. However, pre-invasive breast cancer samples, including
DCIS, were reported to be more successful in developing xenografts
(50,51).
Rats as a model
The rat is considered to be the first domesticated
laboratory mammal used as a model for HBC research, used as early
as 1912. Rats are easier to use in research than mice, as they are
larger in size (52,53). A number of comparative studies have
illustrated the histological similarities between HBC and rat
mammary tumors (50,51). The incidence of breast cancer in rat
models is either spontaneous, or generated by the transgenesis of
activated oncogenes or the induction of mutations in tumor
suppressor genes (54). Chemical
carcinogens have been used to induce breast cancer in the rat
mammary gland for >50 years, most commonly
N-nitroso-N-methylurea or 7,12-dimethylbenz(a)anthracene (55–57). These
chemically-induced tumor models have been frequently utilized in
the investigation of hormone-dependent breast tumors, and the role
of pregnancy in preventing breast tumors (58–60). It
was identified that chemically-induced tumors in rats are
hormone-dependent adenocarcinomas, in contrast to the majority of
mammary tumors that arise in GEMMs (51).
Limitations of rodents as models for
breast cancer
Although rodents are the most frequently used animal
models in cancer research, there are a number of limitations to
their role. Firstly, there are significant differences between the
biology of rodents and their tumors, and that of humans and their
tumors. There are immanent differences in the developmental
programs of rodents and humans. Size is a notable difference, which
affects the number, differentiation and maturation of transformed
cells (61). In addition, observable
tumors in mice require rapid growth progression relatively, as the
lifespan of rodents is relatively short compared with that of
humans. Mice can develop severe malignant lesions with multiple
genetic alterations within 6–18 months (61). Furthermore, rodent cells are much
easier, compared with human cells, to transform with chemical
carcinogens or oncogene transfection in vitro, possibly due
to their relatively poor control of genetic stability, less
effective DNA repair, or altered regulation of gene expression
(62).
Another notable difference is in the tumor pathology
and biology observed between rodents and humans. Fewer genetic
changes are required for rodent cell transformation in
vitro, and potentially also in vivo (37). For example, ~50% of HBCs are
hormone-dependent at diagnosis, whereas the majority of mouse
tumors are hormone-independent, with notably low ER and
progesterone receptor (PR) levels (61).
Although rodents are intrinsically more susceptible
than humans to carcinogenesis, in wild-type rodents, sporadic
cancers are relatively rare (1). In
the past, mouse breeds prone to mammary tumors were produced by
vertical transmission through milk or Bittner factor, which was
later demonstrated to be due to MMTV (63,64). By
contrast, viruses are not convincingly involved in HBC oncogenesis,
except as probable cofactors (64).
Furthermore, rodent cells are easily immortalized (65,66).
Primary human cells and mouse cells also exhibit considerable
differences in telomerase regulation and telomere dynamics.
Telomeres are considerably longer in laboratory mice (40–60 kb)
compared with that of humans (10 kb), and telomerase is highly
expressed in the tissues of adult mice (67). Cancer cells in humans avoid
replicative senescence by maintaining telomeres through telomerase
reactivation and other mechanisms, which may not be analogous to
tumor development in mice (67).
Although the morphological patterns of lesions in
humans and rodents initially appear to be similar, the detailed
features of the majority of mouse tumors are not similar to HBCs,
and cannot be classified as the same pathological grades and types
(68,69). Another major difference is that small
animals, including rats and mice, consume higher oxygen levels on a
per-cell basis, compared with large animals (70). This results in distinct cellular
microenvironments, particularly in comparatively hypoxic and
avascular tumors, in which hypoxia-induced genes may influence
differentiation and growth to an extent that may be
unrepresentative of these processes in human tumor development
(71).
Despite the usefulness of GEMMs in breast cancer
research, it is difficult to regulate the extent of GEMM gene
expression due to multiple copies of one gene, which may be
inserted into the genome (72).
Another limitation of these models is that the removal of tumor
suppressor genes, if not initially intended to the mammary gland,
may have lethal effects on the neonate or fetus (29). GEMMs also exhibit fundamental
differences at the organism and cell levels. GEMMs usually
reproduce highly specific tumor formations, follow all lesion
progression aspects and primarily their design depends on the
awareness of tumor genetics of human (62,72).
During the use of transgenic mice, the type and degree of genetic
abnormalities, which create disease, must be determined in
association with those that create the disease in humans, in order
to evaluate if they disagree and therefore meaning that they are
unsatisfactory models (62).
Asynchronous tumor development in the transgenic host is considered
a major disadvantage of GEMMs (73).
These models are frequently characterized by low penetrance,
miscellaneous tumor frequency, and latent growth properties and
tumor development (69,71). Another challenging problem in
establishing GEMMs of certain cancer types is the scarcity of
tissue-specific promoters, which manage transgene expression in the
adult cells of the targeted tissue (74). Furthermore, GEMMs incur significant
cost, time and effort for the development and maintenance required
to produce analytically meaningful data (32). In addition, as in the majority of
rodents, the metastatic patterns vary from human cases (32). In humans, breast cancer usually
spreads via the lymphatic system, beginning in local lymph nodes,
and spreading to the bone, liver, brain and the lung (75). Whilst in rodents, mammary tumors
metastasize particularly to the lung via the blood (76).
In spite of the important role of carcinogen-induced
models, they still have basic restraints for regular usage in the
early stages of drug discovery (77).
Even though these models require minimal initial cost for
implementation, they require significant time and high cost for
maintenance (77).
While the rat model serves an important role in
breast cancer research, there are a number of disadvantages to its
use. One limitation is that rat mammary tumors rarely metastasize
(51). Compared with mice, the
limited availability of rat genetic engineering
strategies-specifically nuclear transfer for gene knockouts-has
reduced the extensive use of rats as genetic models for the disease
(78). A mutagenesis strategy for
effectively generating breast cancer-associated (BRCA) 1 and BRCA2
gene mutants was developed (79);
however, this method still requires more efficient technologies for
the complete gene knockouts used in breast cancer
investigation.
Cats as a model
A previous study focused on feline mammary tumors to
identify the similarities between feline and human disease
(80). Studies examining the
similarities between human premalignant lesions and feline mammary
intraepithelial lesions (IELs) observed that the latter are
characterized by a spontaneous onset and high prevalence, similar
to humans, as well as all the morphological features of human
pre-invasive breast tumors (81).
These lesions were also distinguished by the loss of PR and ER
expression, promoting the cat as a model for pre-invasive ER- and
PR-negative breast tumors (82).
Weijer et al (83) reported
that, as in humans, feline tumor lesion size, method of growth and
histological grade are associated with disease prognosis.
Additionally, it was indicated that feline mammary carcinomas are
able to provide a suitable model for comparative studies to examine
therapeutic interventions, and serve as an intermediate model for
the study of mammary tumor histogenesis and etiology (83–86).
Limitation of cats as a model for
breast cancer
Despite the similarities determined between human
and feline disease that make the cat as a good model for HBC
studies, there is significant time used to gather the minimum
number of cats required for a single study within a limited time
period (80). Another factor that may
prevent the results of a study from being homogeneous, is that the
study is reliant on pet owners when applying postoperative
treatment in cases that are not in hospitals and following up the
study (81). Additionally, the high
cost required for studies on cats is considered a large
disadvantage (85).
Dogs as a model
In recent decades, clinical and molecular
similarities have been identified between HBC and canine mammary
tumors (CMT) (87). The clinical
similarities include the spontaneous tumor incidence, onset age,
hormonal etiology and the identical course of the disease.
Furthermore, the factors that affect the clinical outcome,
including the tumor size, clinical stage and lymph node invasion,
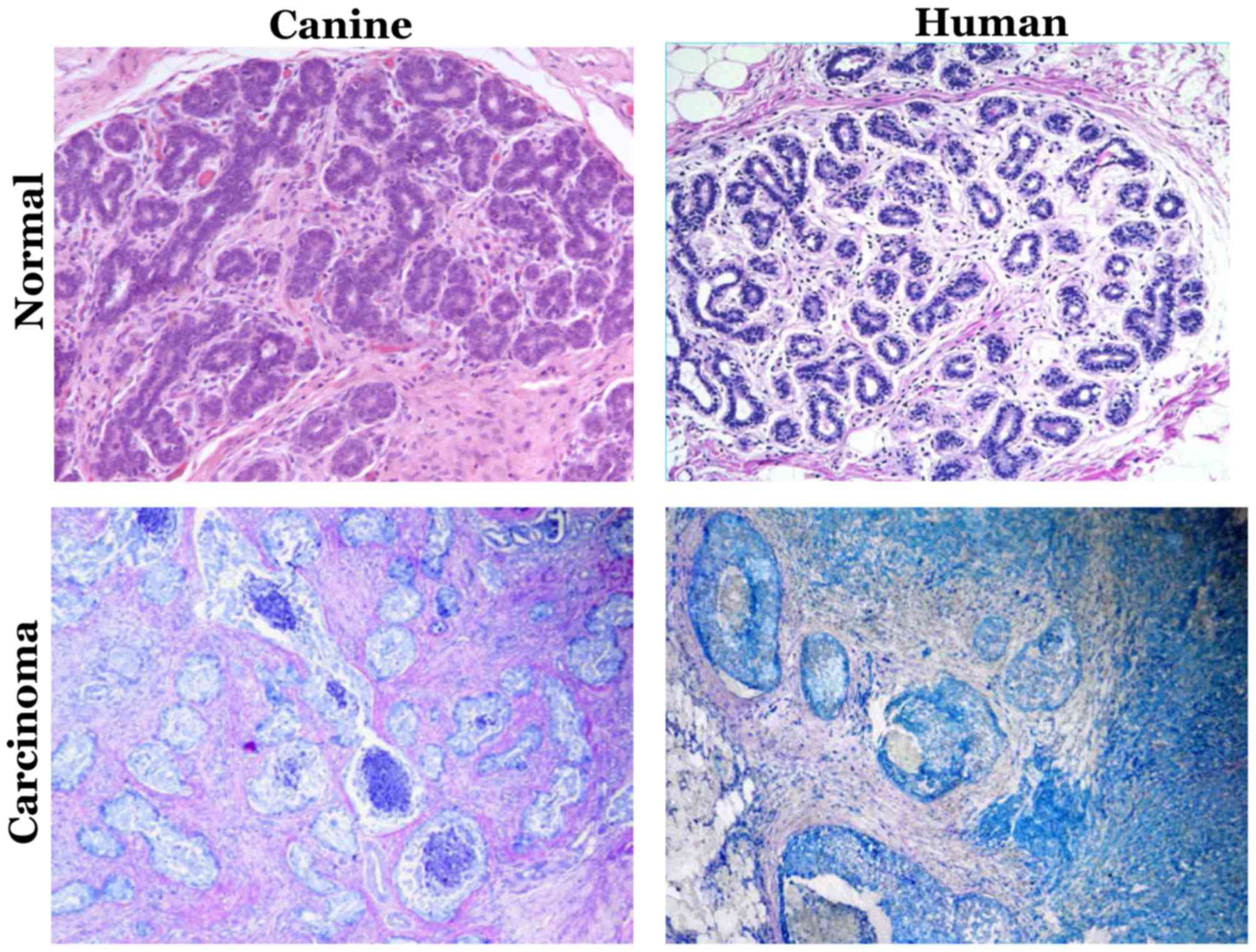
are identical (88). Fig. 1 depicts the histopathological
similarities of mammary intraepithelial lesions between canine
mammary tumor and human breast cancer. The molecular
characteristics, including steroid receptor, epidermal growth
factor (EGF), proliferation markers, metalloproteinase and
cyclooxygenase overexpression, and p53 mutation, also mimic HBC
(88). Table I demonstrated the similarities between
canine mammary tumor and human breast cancer.
 | Table I.Similarities between canine mammary
tumor and human breast cancer. |
Table I.
Similarities between canine mammary
tumor and human breast cancer.
| Similarity
features | Humans | Dogs |
|---|
| Occurrence | Spontaneous | Spontaneous |
| Onset age | Median age, 62
years | Median age, ~10.5
years (10.5-year-old dog, equivalent to a 65.5 year old woman) |
| Course of the
disease | Identical in human
and dog | Identical in human
and dog |
| Size of the
tumor | Similar in human
and dog | Similar in human
and dog |
| Clinical
stages | Identical in both
species | Identical in both
species |
| Invasion to lymph
nodes | Identical in human
and dog | Identical in human
and dog |
| Most common
spontaneous malignancy | Mammary
neoplasia | Mammary
neoplasia |
| Estrogen
dependency | Long exposure to
estrogen increases the risk of tumor occurrence | Non-spayed dogs
have a fourfold higher danger of tumor occurrence than spayed dogs
<2 years old |
| Most common
histological type | Invasive ductal
carcinoma | Carcinomas |
| Premalignant
lesions | Prevalent | Prevalent |
| Molecular
markers | A number of genes
were identified to perform a critical role in carcinogenesis of
mammary tumors | It was determined
that these genes have identical role in carcinogenesis of canine
mammary tumors |
| Mammographic
abnormalities | Dog and human
mammary neoplasm have similar microcalcifications and
macrocalcifications | Dog and human
mammary neoplasm have similar microcalcifications and
macrocalcifications |
The reported incidence of HBC and CMT has increased
recently (89–91), due to the frequent widespread usage of
mammography screening, and the increased awareness of dog owners
(88). The higher incidence of CMT
provides an increased sample size for clinical trials in
comparative medicine.
As in human females, the most common spontaneous
malignancy in female dogs is mammary neoplasia (92), and premalignant lesions are prevalent
in canine mammary glands (93). In
females, breast cancer development is affected by estrogen;
similarly, non-spayed female dogs <2 years old have a 4-fold
higher risk of tumor occurrence compared with spayed dogs of the
same age (94–96).
Morphological and histological features
of canine mammary cancer
i) Molecular biology
During puberty, steroid hormones are essential for
mammogenesis (97); during pregnancy,
the complete development of lobules and alveoli is induced by the
stimulation of progesterone and estrogen (97). In CMT and HBC, ER expression is
associated with the pathological features of the disease and the
degree of tumor differentiation (98).
In CMT, it was identified that larger tumors with
skin ulcerations and reduced prognosis exhibit a reduced level of
ERα expression (99). In humans, ERβ
has a greater associated with benign tumors, and its higher
expression in a tumor of a lower malignancy grade is an indicator
of a relatively good prognosis (99).
The expression of ERβ in HBC has been associated with an extended
survival time in patients with ERα-negative tumors treated with
tamoxifen (100). Divergences in ER
expression have a significant effect on endocrine therapy;
therefore, selective ER modulators that act as full agonists for
ERβ and antagonists for ERα may be effective in CMT treatment,
particularly as a chemoprevention strategy (101).
PR expression has been demonstrated as an important
indicator of breast cancer recurrence in humans (102). A negative PR status is associated
with a reduced prognosis (101,103). A
combination of PR and ER expression status has also been associated
with overall survival. ER−/PR− cases are
associated with the worst survival rates, followed by
ER−/PR+, ER+/PR− and
ER+/PR+ (42).
In CMT, ER and PR expression are common in normal
and benign tissues. It was observed that
ERα+/PR+ was the most common status of benign
tumors, whereas an ERα−/PR+ status was twice
as common as ERα+/PR+ in malignant cancer
(77). Malignant tumors tend to lose
hormonal dependency as the disease progresses towards metastasis,
therefore these tumors and their metastases have a tendency to be
ER−/PR− (95).
In HBC, the ER−/PR− status is associated with
a poorer prognosis (104).
Insulin-like growth factor-1 (IGF-1) and growth
hormone (GH) have been investigated in human breast carcinogenesis
(45,105). Previously, it was identified that
the inhibition of GH/IGF-1 may prevent pre-neoplastic breast
disease and cancer (46). In dogs,
progestin stimulates the mammary gland tissue to locally produce
GHs (48). It was assumed that the
epithelial hyperplasia that was observed following the
administration of progestin and the subsequent biosynthesis by the
mammary gland was mediated by autocrine GHs (47,105). The
full-length canine GH receptor (GHR) sequence in CMT has expanded
homology with the GHR sequence of humans and various other species
(106). This evidence may support
the utility of GH in comparative pathology. According to novel
data, an association between GH and progesterone levels in CMT
homogenates has been identified, indicating that the
progesterone/GH axis has an evident role in CMT (107). The GH/IGF-1 axis is also significant
in CMT prognosis (108).
Pre-surgical serum concentrations of the IGF-1 content of the
malignant lesions and tissue GH levels are associated with a
reduction in the post-surgical survival rate (109). Another study demonstrated that the
concentrations of serum IGF-1 in female dogs with malignant tumors
were higher compared with in healthy controls (110). The predictive value of IGF-1 and GH
in mammary tumors was established for the first time in veterinary
and human medicine (111). This
evidence has facilitated novel therapeutic approaches, including
the use of pegvisomant as a GH-competitive antagonist (108,111).
Prolactin is characterized by its lactogenic
activity, as well as functioning as a growth factor in breast
tumors (112). It has been observed
in breast cancer that prolactin supplementation promotes the
survival of mammary tissue, and that the clinical condition of
patients may improve following hypophysectomy, indicating a
hormonal dependency on prolactin (112). HBC cells and tumors co-express sex
steroid hormone and prolactin receptors, which are cross-regulated
(113). This may clarify the
collaboration between sex steroid hormones and prolactin during
neoplastic growth and mammary development, in particular between
prolactin and progesterone (113).
It is proposed that prolactin and steroid hormones act as local
growth factors, stimulating malignant tumor proliferation (109). In addition, it was determined that
clinical tumor features were associated with prolactin levels in
tissue homogenates, demonstrating an association with steroid
hormone levels (114). Further
studies are required to understand the effect of prolactin on
prognosis in veterinary medicine.
ii) Molecular markers
In recent decades, a number of molecular markers
associated with tumor development have been identified. Of the
number of genes identified to be responsible for breast cancer
occurrence and development in humans, a number were also
demonstrated to serve an essential role in the carcinogenesis of
mammary tumors in dogs (115). These
markers include BRCA gene mutations, EGF receptor (EGFR), Ki-67,
human epidermal growth factor receptor (HER2)/neu, p53, p63, matrix
metalloproteinases, phosphatase and tensin homolog, heat-shock
proteins, mucins, maspin, Sialyl Lewis X antigen and
cyclooxygenase-2 (88). Uva et
al (116) examined the oncogene
expression common between human and canine breast cancer, and
reported significant similarities in the signaling networks that
regulate mammary cancer biology. Germline mutations in BRCA genes
have been associated with a relatively high risk (4-fold) of
mammary tumor development in certain breeds of dogs (117,118).
An association between the expression of mutated BRCA1 or BRCA2,
and RAD51 recombinase (RAD51), a DNA-repairing protein that is,
when upregulated, associated with the development of mammary
tumors, was also reported in dogs (119). Nieto et al (120) revealed that the expression of BRCA1
was reduced in malignant CMT lesions. Reverse
transcription-quantitative polymerase chain reaction on
laser-microdissected tissue samples of normal mammary gland
epithelia, simple canine adenomas and adenocarcinomas free from
non-neoplastic epithelial and stromal cells were used to study the
differential mRNA expression of RAD51, BRCA1 and BRCA2 (121). In that study, the expression of
BRCA1 was not clearly associated with the histological criteria of
malignancy, in contrast to human tumors; however, BRCA2 and BRCA1
were overexpressed in 50% of lymph node metastases, whereas RAD51
was overexpressed in 50% of primary tumors and 80% of lymph node
metastases. According to these conflicting results, a reduction in
expression in more malignant cells may be expected. The conclusion
was that this overexpression was induced by the inhibition of
proliferation due to a negative regulatory loop in which BRCA2 was
produced by cellular proliferation (122).
EGFR, also known as HER1, is considered a molecular
marker in triple-negative breast cancer, including ER−,
PR− and HER2/neu− (123). In CMT, cases with positive EGFR
expression are associated with reduced disease-free and overall
survival rates. Additionally, it was reported that benign and
hyperplastic lesions consistently express EGFR in the myoepithelial
cells of humans and dogs (124). It
has been indicated that in tissues with high concentrations of
progesterone and EGF, a mechanism involving the angiogenic effects
of EGF and the induction of proliferation by progesterone may exist
in CMT (125).
HER2/neu is another important prognostic factor
(126). HER2/neu+
tumor-specific therapy has yielded promising treatment results
(126). In the study by Rungsipipat
et al (127), out of 79 CMT
cases, HER2/neu was expressed in a ~19% adenocarcinomas and ~50%
benign tumors, including simple and complex adenomas. In a study of
malignant CMT and HBC, the rate of HER2/neu expression was similar
(20 and 30%) (128). Kerns et
al (128) detected the
co-expression of HER2/neu and p53 mutation, which induced malignant
cell behavior. Similar to humans, four phenotypic subtypes have
been identified in malignant CMT, according to the
immunohistochemical expression of HER2 and ER: HER2 overexpressing
(ER−/HER2+, 8.3%); basal-like
(ER−/HER2−, 29.2%); luminal A
(ER+/HER2−, 44.8%); and luminal B
(ER+/HER2+, 13.5%) (129,130).
Individuals diagnosed with the basal-like subtype have reduced
survival rates (131). This
indicates the molecular heterogeneity of HBC and CMT, providing
further evidence that CMT is a potential model for the study of
HBC.
The Ki-67 antigen is a nuclear protein highly
expressed in proliferating cells prior to mitosis (132). Ki-67 labeling has been identified in
different types of human and canine tumors (133). For humans, immunohistochemical
determination of Ki-67 expression in mammary carcinomas has been
indicated as a prognostic marker for relapse-free survival time
(134). Proliferating cell nuclear
antigen (PCNA) and other proliferation markers have been studied in
combination with Ki-67 (135). The
reliable prognostic value of PCNA in other types of malignancy has
been described by several studies (135); however, it was determined to be a
poor prognostic indicator for human mammary carcinoma (135). The immunohistochemical expression of
Ki-67 and PCNA has also been studied in CMT; the expression of
Ki-67 has been associated with a poor prognosis (133). PCNA was observed to be more
frequently expressed in malignant CMT than in normal mammary
glands, hyperplasia and benign tumors (136). Similar to HBC, the expression of
Ki-67 in benign and malignant tumors was reported to be
considerably associated with the presence of remote metastases, as
well as associated with the histological criteria of malignancy and
the PCNA index (137).
Following DNA damage, the p53 tumor suppressor gene
has an important role in regulating cell growth. Deregulated cell
proliferation occurs following mutations of this gene, which causes
tumor formation and progression (138). In HBC, p53 gene mutation leads to
the accumulated nuclear expression of the p53 protein. This event
has prognostic value, and is usually indicative of a relatively
poor survival time (139). In CMT,
the expression of mutant p53 has also been identified as a valuable
prognostic marker (140). The
frequency of p53 mutation in CMT is ~20%, which is analogous to
that detected in HBC (141). In
humans and dogs, mutations in the conserved domains of p53 appear
to have an essential role in the carcinogenesis of mammary glands
(142). It was identified that p53
mutations in CMT occur within the second, fourth and fifth exons,
and Ala125Val point mutations are common between human and canine
p53 genes (143). A study of
aggressive CMT carcinomas indicated an association between p53
mutations and the aggressive type (143), which has also been concluded in HBC
studies (143). Additionally, as the
arrangement of p53 gene products and coding exons are similar in
both species, there may be a shared therapeutic target (141).
The p63 gene is from the p53 gene family. p63 is
involved in the regeneration of epithelial stem cells and is not
considered a tumor suppressor gene (144). It was demonstrated to be
overexpressed in the basal epithelial cells of various human cancer
types (144). The p63 gene may act
as a tumor promoter in specific pathological conditions (145). In humans, p63 is overexpressed in
the breast carcinoma basal phenotype, in which it regulates the
growth and differentiation of stratified epithelia (146). In humans, p63 is a specific and
sensitive indicator for myoepithelial cells (147). Strongly positive p63 staining
indicates myoepithelial differentiation in papillary carcinomas
(147). p63 has been determined to
be a strong myoepithelial cell indicator in dogs, and is important
for the differentiation of myoepithelial or basal cells from
stromal myofibroblasts (148).
Furthermore it was determined that the overexpression of p63 was
associated with stem and myoepithelial cells, which may explain the
histogenesis of CMT (149).
iii) Mammography and ultrasound
imaging
In order to confirm that the dog is a model that
represents HBC in every aspect, x-ray mammography has been used to
define the similarities between human and dog specimens. X-ray
mammography is currently used to detect human DCIS by identifying
the presence of microcalcifications (150). Such microcalcifications are
identified in 42–72% of DCIS cases (150,151).
The existence of microcalcifications alone is used to diagnose ~90%
of nonpalpable DCIS (152).
Mammogram data are classified by a Breast Imaging Reporting and
Data System (153) (BI-RADS),
according to the morphology and distribution of calcifications,
into three types: Typical benign, intermediate concern, and higher
expectation of malignancy (153).
The detection of DCIS has improved due to mammography use; however,
whether the detection of DCIS by mammography leads to a decrease in
the breast cancer mortality rate has yet to be fully investigated
(154). Thus, they assessed the dog
as a model for studying pre-invasive and invasive breast tumors.
The study identified mammographic abnormalities in canine mammary
pre-invasive lesions, benign tumors and malignant tumors that were
similar to the abnormalities associated with HBC. These
abnormalities include the pattern, presence and distribution of
microcalcifications and macrocalcifications (154). BI-RADS categorization was thus
considered a precise method for the detection of mammary malignant
lesions in canines, with high specificity and sensitivity (155).
In a study of 88 mammary glands excised from 40
female dogs of different breeds, mammographic features, including
microcalcifications, macrocalcifications, mass margin and shape,
and their association with the histological diagnosis, were
identified in 78/88 mammary glands (155). The remaining 10 mammary glands
exhibited a normal mammographic appearance (155). Calcification was identified in 42/78
glands (54%), of which 19 glands (24%) had macrocalcifications and
23 glands (29%) contained microcalcifications. Lesions with
microcalcifications (29%) were amorphous/indistinct (5/23, 22%),
linear branching (4/23, 17%) or pleomorphic regional and diffuse
(14/23, 61%), and the majority were associated with atypical IELs
and malignant tumors. On the other hand, 11/19 lesions with
macrocalcifications (57%) were ‘popcorn-like’, with regional
distribution in four glands (24%) and diffuse distribution in seven
glands (36%). The other 8/19 (42%) were round, with regional
localization. Lesions with macrocalcifications were diagnosed as
malignant and benign tumors, primarily of the complex type or IELs
without atypia. In addition, mass margins and shape were evaluated
by mammography in 54/78 (69%) glands. Of the benign lesions, 21/26
(81%) were oval or round masses and 12/26 (46%) had circumscribed
margins. Of the malignant lesions, 12/25 (48%) were irregular in
shape, and 22/25 (88%) exhibited indistinct margins. The ultrasound
abnormalities of 46/78 mammary lesions corresponded with
histological data. Using ultrasound, calcifications were identified
in 41% of the lesions and 53% were associated with malignant
tumors. An oval shape was observed in 25/40 (62.5%) benign or
malignant tumors. Of the benign lesions, 78% featured a
well-circumscribed margin, whereas 80% of malignant lesions
featured irregular margins (155).
The study indicated that dog and human mammary
neoplasms form similar microcalcifications and macrocalcifications.
It also indicated a similar process of cancer pathogenesis between
humans and dogs. Furthermore, it was determined that the
sonographic and mammographic features of benign and malignant
canine mammary IELs were associated with the histopathological
data. The BI-RADS categorization was concluded to be an excellent
predictor of malignant canine mammary lesions. In addition, the
study indicated that dogs are suitable models for HBC (155).
Limitations of dog as a model for
breast cancer
Using pets as a model for studying has the same
limitations as in case of cats, including high cost and time
consumption, and reliance on the owner of the pet post-treatment
(156). These are considered
significant limitations for using dogs as a model for HBC.
Conclusion and future perspectives
CMT resembles human mammary tumors in every aspect.
Invasive mammary tumors, as well as pre-invasive lesions, are
similar to those in human in histopathological, molecular and
imaging characteristics, and clinical outcomes (90). Mammary tumors develop in dogs
spontaneously without genetic or chemical manipulation with an
intact immune system. Recently, there has been increasing interest
in the development of immunotherapy and immunoprevention strategies
to treat or prevent cancer (157).
Dogs are an excellent resource for testing these modalities in a
preclinical setting, as remarkable similarities exist between
humans and dogs with regard to tumor infiltrating lymphocytes
(TIL), including the association between TIL numbers and mammary
tumor aggressiveness, the association between the
CD4+/CD8+ T-cell ratio and survival rate, the
promotion of tumor progression by Th2 cells, and the association
between Treg cell numbers and poor prognostic factors (158).
Furthermore, the development of prophylactic
vaccines for cancer faces multiple challenges, including the low
cancer incidence in humans, the years it takes for the efficacy of
the vaccine to be determined and the cost of human clinical trials.
The dog model addresses these challenges, as the published work has
demonstrated that CMTs are so prevalent in non-spayed female dogs,
that 50% of randomly screened dogs (n=150) have premalignant
lesions in one or two mammary glands (159). Dogs with premalignant lesions may
progress to invasive cancer in one year (160), allowing for the testing of the
efficacy of a vaccine in a short time. Additionally, although dogs
are more outbred compared with laboratory rodents, certain breeds
are at an increased risk of developing mammary tumors. Given the
high homology between the canine genome sequence and its human
counterpart, the dog model offers an excellent resource to explore
prevention strategies for triple-negative breast cancer in females,
particularly high-risk BRCA1/2 mutation carriers.
Acknowledgements
The authors would like to thank previous and
current members of the Dr Sulma Mohammed lab for their help and
improvement of this work.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
SMA wrote the first draft and SM edited the final
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ghoncheh M, Pournamdar Z and Salehiniya H:
Incidence and mortality and epidemiology of breast cancer in the
world. Asian Pac J Cancer Prev. 17:43–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis CE, Bray F, Ferlay J,
Lortet-Tieulent J, Anderson BO and Jemal A: International variation
in female breast cancer incidence and mortality rates. Cancer
Epidemiol Biomarkers Prev. 24:1495–1506. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
American Cancer Society: Cancer Facts
& Figures. American Cancer Society, Inc.; Atlanta, GA: 2015
|
|
4
|
Gazdar AF, Kurvari V, Virmani A, Gollahon
L, Sakaguchi M, Westerfield M, Kodagoda D, Stasny V, Cunningham HT,
Wistuba II, et al: Characterization of paired tumor and non-tumor
cell lines established from patients with breast cancer. Int J
Cancer. 78:766–774. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Holen I, Speirs V, Morrissey B and Blyth
K: In vivo models in breast cancer research: Progress, challenges
and future directions. Dis Model Mech. 10:359–371. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Levenson AS and Jordan VC: MCF-7: The
first hormone-responsive breast cancer cell line. Cancer Res.
57:3071–3078. 1997.PubMed/NCBI
|
|
7
|
Osborne CK, Hobbs K and Trent JM:
Biological differences among MCF-7 human breast cancer cell lines
from different laboratories. Breast Cancer Res Treat. 9:111–121.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Burdall SE, Hanby AM, Lansdown MR and
Speirs V: Breast cancer cell lines: Friend or foe? Breast Cancer
Res. 5:89–95. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Amadori D, Bertoni L, Flamigni A, Savini
S, De Giovanni C, Casanova S, De Paola F, Amadori A, Giulotto E and
Zoli W: Establishment and characterization of a new cell line from
primary human breast carcinoma. Breast Cancer Res Treat.
28:251–260. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yong JW, Choong ML, Wang S, Wang Y, Lim SQ
and Lee MA: Characterization of ductal carcinoma in situ cell lines
established from breast tumor of a Singapore Chinese patient.
Cancer Cell Int. 14:942014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Meltzer P, Leibovitz A, Dalton W, Villar
H, Kute T, Davis J, Nagle R and Trent J: Establishment of two new
cell lines derived from human breast carcinomas with HER-2/neu
amplification. Br J Cancer. 63:727–735. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cailleau R, Olivé M and Cruciger QV:
Long-term human breast carcinoma cell lines of metastatic origin:
Preliminary characterization. In vitro. 14:911–915. 1978.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Engel LW and Young NA: Human breast
carcinoma cells in continuous culture: A review. Cancer Res.
38:4327–4339. 1978.PubMed/NCBI
|
|
14
|
Nayak SK, Kakati S, Harvey SR, Malone CC,
Cornforth AN and Dillman RO: Characterization of cancer cell lines
established from two human metastatic breast cancers. In vitro Cell
Dev Biol Anim. 36:188–193. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kurebayashi J, Otsuki T, Tang CK, Kurosumi
M, Yamamoto S, Tanaka K, Mochizuki M, Nakamura H and Sonoo H:
Isolation and characterization of a new human breast cancer cell
line, KPL-4, expressing the Erb B family receptors and
interleukin-6. Br J Cancer. 79:707–717. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rye PD, Norum L, Olsen DR, Garman-Vik S,
Kaul S and Fodstad O: Brain metastasis model in athymic nude mice
using a novel MUC1-secreting human breast-cancer cell line, MA11.
Int J Cancer. 68:682–687. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zoli W, Roncuzzi L, Flamigni A, Gruppioni
R, Sensi A, Zini N, Amadori D and Gasperi-Campani A: A new cell
line from human infiltrating ductal carcinoma of the breast:
Establishment and characterization. J Cancer Res Clin Oncol.
122:237–242. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Engel LW, Young NA, Tralka TS, Lippman ME,
O'Brien SJ and Joyce MJ: Establishment and characterization of
three new continuous cell lines derived from human breast
carcinomas. Cancer Res. 38:3352–3364. 1978.PubMed/NCBI
|
|
19
|
Engebraaten O and Fodstad O: Site-specific
experimental metastasis patterns of two human breast cancer cell
lines in nude rats. Int J Cancer. 82:219–225. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lacroix M and Leclercq G: Relevance of
breast cancer cell lines as models for breast tumours: An update.
Breast Cancer Res Treat. 83:249–289. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nelson-Rees WA, Daniels DW and
Flandermeyer RR: Cross-contamination of cells in culture. Science.
212:446–452. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rivenbark AG, O'Connor SM and Coleman WB:
Molecular and cellular heterogeneity in breast cancer: Challenges
for personalized medicine. Am J Pathol. 183:1113–1124. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Weiswald LB, Bellet D and Dangles-Marie V:
Spherical cancer models in tumor biology. Neoplasia. 17:1–15. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ethier SP, Mahacek ML, Gullick WJ, Frank
TS and Weber BL: Differential isolation of normal luminal mammary
epithelial cells and breast cancer cells from primary and
metastatic sites using selective media. Cancer Res. 53:627–635.
1993.PubMed/NCBI
|
|
25
|
Wistuba II, Behrens C, Milchgrub S, Syed
S, Ahmadian M, Virmani AK, Kurvari V, Cunningham TH, Ashfaq R,
Minna JD and Gazdar AF: Comparison of features of human breast
cancer cell lines and their corresponding tumors. Clin Cancer Res.
4:2931–2938. 1998.PubMed/NCBI
|
|
26
|
Latimer JJ, Nazir T, Flowers LC, Forlenza
MJ, Beaudry-Rodgers K, Kelly CM, Conte JA, Shestak K,
Kanbour-Shakir A and Grant SG: Unique tissue-specific level of DNA
nucleotide excision repair in primary human mammary epithelial
cultures. Exp Cell Res. 291:111–121. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Freshney RI: Animal cell culture: a
practical approach. IRL Press (Oxford University Press); Oxford:
1992
|
|
28
|
Kim IS and Baek SH: Mouse models for
breast cancer metastasis. Biochem Biophys Res Commun. 394:443–447.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Allred DC and Medina D: The relevance of
mouse models to understanding the development and progression of
human breast cancer. J Mammary Gland Biol Neoplasia. 13:279–288.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lewis MT and Porter WW: Methods in mammary
gland biology and breast cancer research: An update. J Mammary
Gland Biol Neoplasia. 14:3652009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Perez C, Parker-Thornburg J, Mikulec C,
Kusewitt DF, Fischer SM, Digiovanni J, Conti CJ and Benavides F:
SKHIN/Sprd, a new genetically defined inbred hairless mouse strain
for UV-induced skin carcinogenesis studies. Exp Dermatol.
21:217–220. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Borowsky A: Special considerations in
mouse models of breast cancer. Breast Dis. 28:29–38. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen Q and Brown PH: Novel agents for the
prevention of breast cancer: Targeting transcription factors and
signal transduction pathways. J Mammary Gland Biol Neoplasia.
8:45–73. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huijbers IJ, Krimpenfort P, Berns A and
Jonkers J: Rapid validation of cancer genes in chimeras derived
from established genetically engineered mouse models. Bioessays.
33:701–710. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ruggeri BA, Camp F and Miknyoczki S:
Animal models of disease: Pre-clinical animal models of cancer and
their applications and utility in drug discovery. Biochem
Pharmacol. 87:150–161. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Du Z and Li Y: RCAS-TVA in the mammary
gland: An in vivo oncogene screen and a high fidelity model for
breast transformation? Cell Cycle. 6:823–826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Balmain A and Harris CC: Carcinogenesis in
mouse and human cells: Parallels and paradoxes. Carcinogenesis.
21:371–377. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Barrett JC: Mechanisms of multistep
carcinogenesis and carcinogen risk assessment. Environ Health
Perspect. 100:9–20. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Steele VE, Moon RC, Lubet RA, Grubbs CJ,
Reddy BS, Wargovich M, McCormick DL, Pereira MA, Crowell JA,
Bagheri D, et al: Preclinical efficacy evaluation of potential
chemopreventive agents in animal carcinogenesis models: Methods and
results from the NCI chemoprevention drug development program. J
Cell Biochem Suppl. 20:32–54. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Abel EL, Angel JM, Kiguchi K and
DiGiovanni J: Multi-stage chemical carcinogenesis in mouse skin:
Fundamentals and applications. Nat Protoc. 4:1350–1362. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tsukamoto T, Mizoshita T and Tatematsu M:
Animal models of stomach carcinogenesis. Toxicol Pathol.
35:636–648. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Takahashi M, Hori M, Mutoh M, Wakabayashi
K and Nakagama H: Experimental animal models of pancreatic
carcinogenesis for prevention studies and their relevance to human
disease. Cancers (Basel). 3:582–602. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Grivennikov SI, Greten FR and Karin M:
Immunity, inflammation, and cancer. Cell. 140:883–899. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shoushtari AN, Michalowska AM and Green
JE: Comparing genetically engineered mouse mammary cancer models
with human breast cancer by expression profiling. Breast Dis.
28:39–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pichon MF, Broet P, Magdelenat H, Delarue
JC, Spyratos F, Basuyau JP, Saez S, Rallet A, Courriere P, Millon R
and Asselain B: Prognostic value of steroid receptors after
long-term follow-up of 2257 operable breast cancers. Br J Cancer.
73:1545–1551. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Schernhammer ES, Holly JM, Hunter DJ,
Pollak MN and Hankinson SE: Insulin-like growth factor-I, its
binding proteins (IGFBP-1 and IGFBP-3), and growth hormone and
breast cancer risk in the nurses health study II. Endocr Relat
Cancer. 13:583–592. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kleinberg DL, Wood TL, Furth PA and Lee
AV: Growth hormone and insulin-like growth factor-I in the
transition from normal mammary development to preneoplastic mammary
lesions. Endocr Rev. 30:51–74. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Selman PJ, Mol JA, Rutteman GR, van
Garderen E and Rijnberk A: Progestin-induced growth hormone excess
in the dog originates in the mammary gland. Endocrinology.
134:287–292. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mehta RR, Graves JM, Hart GD, Shilkaitis A
and Das Gupta TK: Growth and metastasis of human breast carcinomas
with Matrigel in athymic mice. Breast Cancer Res Treat. 25:65–71.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gandhi A, Holland PA, Knox WF, Potten CS
and Bundred NJ: Effects of a pure antiestrogen on apoptosis and
proliferation within human breast ductal carcinoma in situ. Cancer
Res. 60:4284–4288. 2000.PubMed/NCBI
|
|
51
|
Chan KC, Knox WF, Gee JM, Morris J,
Nicholson RI, Potten CS and Bundred NJ: Effect of epidermal growth
factor receptor tyrosine kinase inhibition on epithelial
proliferation in normal and premalignant breast. Cancer Res.
62:122–128. 2002.PubMed/NCBI
|
|
52
|
Shanks N, Greek R and Greek J: Are animal
models predictive for humans? Philos Ethics Humanit Med. 4:22009.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ericsson AC, Crim MJ and Franklin CL: A
brief history of animal modeling. Mo Med. 110:201–205.
2013.PubMed/NCBI
|
|
54
|
Russo J, Gusterson BA, Rogers AE, Russo
IH, Wellings SR and van Zwieten MJ: Comparative study of human and
rat mammary tumorigenesis. Lab Invest. 62:244–278. 1990.PubMed/NCBI
|
|
55
|
Russo J and Russo IH: Atlas and histologic
classification of tumors of the rat mammary gland. J Mammary Gland
Biol Neoplasia. 5:187–200. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Szpirer C: Cancer research in rat models.
Methods Mol Biol. 597:445–458. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Russo IH and Russo J: Developmental stage
of the rat mammary gland as determinant of its susceptibility to
7,12-dimethylbenz[a]anthracene. J Natl Cancer Inst. 61:1439–1449.
1978.PubMed/NCBI
|
|
58
|
Thompson HJ, Adlakha H and Singh M: Effect
of carcinogen dose and age at administration on induction of
mammary carcinogenesis by 1-methyl-1-nitrosourea. Carcinogenesis.
13:1535–1539. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Thompson HJ and Meeker LD: Induction of
mammary gland carcinomas by the subcutaneous injection of
1-methyl-1-nitrosourea. Cancer Res. 43:1628–1629. 1983.PubMed/NCBI
|
|
60
|
Russo J, Balogh GA, Heulings R, Mailo DA,
Moral R, Russo PA, Sheriff F, Vanegas J and Russo IH: Molecular
basis of pregnancy-induced breast cancer protection. Eur J Cancer
Prev. 15:306–342. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Nandi S, Guzman RC and Yang J: Hormones
and mammary carcinogenesis in mice, rats, and humans: A unifying
hypothesis. Proc Natl Acad Sci USA. 92:3650–3657. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Holliday R: Neoplastic transformation: The
contrasting stability of human and mouse cells. Cancer Surv.
28:103–115. 1996.PubMed/NCBI
|
|
63
|
Holt MP, Shevach EM and Punkosdy GA:
Endogenous mouse mammary tumor viruses (mtv): New roles for an old
virus in cancer, infection and immunity. Front Oncol. 3:2872013.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wong M, Pagano JS, Schiller JT, Tevethia
SS, Raab-Traub N and Gruber J: New associations of human
papillomavirus, Simian virus 40, and Epstein-Barr virus with human
cancer. J Natl Cancer Inst. 94:1832–1836. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Newbold RF: Genetic control of telomerase
and replicative senescence in human and rodent cells. Ciba Found
Symp. 211:177–197. 1997.PubMed/NCBI
|
|
66
|
Greenberg RA, Allsopp RC, Chin L, Morin GB
and DePinho RA: Expression of mouse telomerase reverse
transcriptase during development, differentiation and
proliferation. Oncogene. 16:1723–1730. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Cabuy E, Newton C, Roberts T, Newbold R
and Slijepcevic P: Identification of subpopulations of cells with
differing telomere lengths in mouse and human cell lines by flow
FISH. Cytometry A. 62:150–161. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Cardiff RD, Anver MR, Gusterson BA,
Hennighausen L, Jensen RA, Merino MJ, Rehm S, Russo J, Tavassoli
FA, Wakefield LM, et al: The mammary pathology of genetically
engineered mice: The consensus report and recommendations from the
Annapolis meeting. Oncogene. 19:968–988. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Cardiff RD: Validity of mouse mammary
tumour models for human breast cancer: Comparative pathology.
Microsc Res Tech. 52:224–230. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Savage VM, Allen AP, Brown JH, Gillooly
JF, Herman AB, Woodruff WH and West GB: Scaling of number, size,
and metabolic rate of cells with body size in mammals. Proc Natl
Acad Sci USA. 104:4718–4723. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Helczynska K, Kronblad A, Jögi A, Nilsson
E, Beckman S, Landberg G and Påhlman S: Hypoxia promotes a
dedifferentiated phenotype in ductal breast carcinoma in situ.
Cancer Res. 63:1441–1444. 2003.PubMed/NCBI
|
|
72
|
Walrath JC, Hawes JJ, Van Dyke T and
Reilly KM: Genetically engineered mouse models in cancer research.
Adv Cancer Res. 106:113–164. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Gomez-Cuadrado L, Tracey N, Ma R, Qian B
and Brunton VG: Mouse models of metastasis: Progress and prospects.
Dis Model Mech. 10:1061–1074. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Frese KK and Tuveson DA: Maximizing mouse
cancer models. Nat Rev Cancer. 7:645–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Rahman M and Mohammed S: Breast cancer
metastasis and the lymphatic system. Oncol Lett. 10:1233–1239.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Kucherlapati R: Genetically modified mouse
models for biomarker discovery and preclinical drug testing. Clin
Cancer Res. 18:625–630. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Olive KP and Tuveson DA: The use of
targeted mouse models for preclinical testing of novel cancer
therapeutics. Clin Cancer Res. 12:5277–5287. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Huang G, Ashton C, Kumbhani DS and Ying
QL: Genetic manipulations in the rat: Progress and prospects. Curr
Opin Nephrol Hypertens. 20:391–399. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Cheung A, Young L, Chen P, Chao C, Ndoye
A, Barry P, Muller W and Cardiff R: Microcirculation and metastasis
in a new mouse mammary tumor model system. Int J Oncol. 11:69–77.
1997.PubMed/NCBI
|
|
80
|
Wiese DA, Thaiwong T, Yuzbasiyan-Gurkan V
and Kiupel M: Feline mammary basal-like adenocarcinomas: A
potential model for human triple-negative breast cancer (TNBC) with
basal-like subtype. BMC Cancer. 13:4032013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
De Maria R, Olivero M, Iussich S, Nakaichi
M, Murata T, Biolatti B and Di Renzo MF: Spontaneous feline mammary
carcinoma is a model of HER2 overexpressing poor prognosis human
breast cancer. Cancer Res. 65:907–912. 2005.PubMed/NCBI
|
|
82
|
Burrai GP, Mohammed SI, Miller MA, Marras
V, Pirino S, Addis MF, Uzzau S and Antuofermo E: Spontaneous feline
mammary intraepithelial lesions as a model for human estrogen
receptor- and progesterone receptor-negative breast lesions. BMC
Cancer. 10:1562010. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Weijer K, Head KW, Misdorp W and Hampe JF:
Feline malignant mammary tumors. I. Morphology and biology: Some
comparisons with human and canine mammary carcinomas. J Natl Cancer
Inst. 49:1697–1704. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Li W, Xiao C, Vonderhaar BK and Deng CX: A
role of estrogen/ERalpha signaling in BRCA1-associated
tissue-specific tumor formation. Oncogene. 26:7204–7212. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Lin SC, Lee KF, Nikitin AY, Hilsenbeck SG,
Cardiff RD, Li A, Kang KW, Frank SA, Lee WH and Lee EY: Somatic
mutation of p53 leads to estrogen receptor alpha-positive and
-negative mouse mammary tumors with high frequency of metastasis.
Cancer Res. 64:3525–3532. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Jang JW, Boxer RB and Chodosh LA:
Isoform-specific ras activation and oncogene dependence during MYC-
and Wnt-induced mammary tumorigenesis. Mol Cell Biol. 26:8109–8121.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Owen LN: A comparative study of canine and
human breast cancer. Invest Cell Pathol. 2:257–275. 1979.PubMed/NCBI
|
|
88
|
Queiroga FL, Raposo T, Carvalho MI, Prada
J and Pires I: Canine mammary tumours as a model to study human
breast cancer: Most recent findings. In vivo. 25:455–465.
2011.PubMed/NCBI
|
|
89
|
Glass AG, Lacey JV Jr, Carreon JD and
Hoover RN: Breast cancer incidence, 1980–2006: combined roles of
menopausal hormone therapy, screening mammography, and estrogen
receptor status. J Natl Cancer Inst. 99:1152–1161. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Pollan M, Pastor-Barriuso R, Ardanaz E,
Argüelles M, Martos C, Galcerán J, Sánchez-Pérez MJ, Chirlaque MD,
Larrañaga N, Martínez-Cobo R, et al: Recent changes in breast
cancer incidence in Spain, 1980–2004. J Natl Cancer Inst.
101:1584–1591. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Dobson JM, Samuel S, Milstein H, Rogers K
and Wood JL: Canine neoplasia in the UK: Estimates of incidence
rates from a population of insured dogs. J Small Anim Pract.
43:240–246. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Misdorp W: Armed Forces Institute of
Pathology (U.S.); American Registry of Pathology.; WHO
Collaborating Center for Worldwide Reference on Comparative
Oncology: Histological classification of mammary tumors of the dog
and the cat. Washington: Armed Forces Institute of Pathology in
cooperation with the American Registry of Pathology and the World
Health Organization Collaborating Center for Worldwide Reference on
Comparative Oncology. Comp Oncol. 7:591999.
|
|
93
|
Antuofermo E, Miller MA, Pirino S, Xie J,
Badve S and Mohammed SI: Spontaneous mammary intraepithelial
lesions in dogs-a model of breast cancer. Cancer Epidemiol
Biomarkers Prev. 16:2247–2256. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Schneider R: Comparison of age, sex, and
incidence rates in human and canine breast cancer. Cancer.
26:419–426. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Rutteman GR, Misdorp W, Blankenstein MA
and van den Brom WE: Oestrogen (ER) and progestin receptors (PR) in
mammary tissue of the female dog: different receptor profile in
non-malignant and malignant states. Br J Cancer. 58:594–599. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Fisher ER, Sass R and Fisher B: Pathologic
findings from the national surgical adjuvant breast project.
Correlations with concordant and discordant estrogen and
progesterone receptors. Cancer. 59:1554–1559. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Lamote I, Meyer E, Massart-Leen AM and
Burvenich C: Sex steroids and growth factors in the regulation of
mammary gland proliferation, differentiation, and involution.
Steroids. 69:145–159. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Elston CW: Classification and grading of
invasive breast carcinoma. Verh Dtsch Ges Pathol. 89:35–44.
2005.PubMed/NCBI
|
|
99
|
de las Mulas Martín J, Ordás J, Millán MY,
Chacón F, De Lara M, de los Monteros Espinosa A, Reymundo C and
Jover A: Immunohistochemical expression of estrogen receptor beta
in normal and tumoral canine mammary glands. Vet Pathol.
41:269–272. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Gruvberger-Saal SK, Bendahl PO, Saal LH,
Laakso M, Hegardt C, Edén P, Peterson C, Malmström P, Isola J, Borg
A and Fernö M: Estrogen receptor beta expression is associated with
tamoxifen response in ERalpha-negative breast carcinoma. Clin
Cancer Res. 13:1987–1994. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Saji S, Hirose M and Toi M: Clinical
significance of estrogen receptor beta in breast cancer. Cancer
Chemother Pharmacol. 56 Suppl 1:S21–S26. 2005. View Article : Google Scholar
|
|
102
|
Joensuu K, Leidenius M, Kero M, Andersson
LC, Horwitz KB and Heikkilä P: ER, PR, HER2, Ki-67 and CK5 in early
and late relapsing breast cancer-reduced CK5 expression in
metastases. Breast Cancer (Auckl). 7:23–34. 2013.PubMed/NCBI
|
|
103
|
Gelbfish GA, Davidson AL, Kopel S,
Schreibman B, Gelbfish JS, Degenshein GA, Herz BL and Cunningham
JN: Relationship of estrogen and progesterone receptors to
prognosis in breast cancer. Ann Surg. 207:75–79. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Mouttet D, Laé M, Caly M, Gentien D,
Carpentier S, Peyro-Saint-Paul H, Vincent-Salomon A, Rouzier R,
Sigal-Zafrani B, Sastre-Garau X and Reyal F: Estrogen-receptor,
progesterone-receptor and HER2 status determination in invasive
breast cancer. Concordance between immuno-histochemistry and
MapQuant™ microarray based assay. PLoS One. 11:e01464742016.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Krajcik RA, Borofsky ND, Massardo S and
Orentreich N: Insulin-like growth factor I (IGF-I), IGF-binding
proteins, and breast cancer. Cancer Epidemiol Biomarkers Prev.
11:1566–1573. 2002.PubMed/NCBI
|
|
106
|
van Garderen E, de Wit M, Voorhout WF,
Rutteman GR, Mol JA, Nederbragt H and Misdorp W: Expression of
growth hormone in canine mammary tissue and mammary tumors.
Evidence for a potential autocrine/paracrine stimulatory loop. Am J
Pathol. 150:1037–1047. 1997.PubMed/NCBI
|
|
107
|
van Garderen E, van der Poel HJ,
Swennenhuis JF, Wissink EH, Rutteman GR, Hellmén E, Mol JA and
Schalken JA: Expression and molecular characterization of the
growth hormone receptor in canine mammary tissue and mammary
tumors. Endocrinology. 140:5907–5914. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Divisova J, Kuiatse I, Lazard Z, Weiss H,
Vreeland F, Hadsell DL, Schiff R, Osborne CK and Lee AV: The growth
hormone receptor antagonist pegvisomant blocks both mammary gland
development and MCF-7 breast cancer xenograft growth. Breast Cancer
Res Treat. 98:315–327. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Queiroga FL, Pérez-Alenza MD, Silvan G,
Peña L, Lopes CS and Illera JC: Crosstalk between GH/IGF-I axis and
steroid hormones (progesterone, 17beta-estradiol) in canine mammary
tumours. J Steroid Biochem Mol Biol. 110:76–82. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Queiroga FL, Pérez-Alenza D, Silvan G,
Peña L, Lopes CS and Illera JC: Serum and intratumoural GH and
IGF-I concentrations: Prognostic factors in the outcome of canine
mammary cancer. Res Vet Sci. 89:396–403. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Yin D, Vreeland F, Schaaf LJ, Millham R,
Duncan BA and Sharma A: Clinical pharmacodynamic effects of the
growth hormone receptor antagonist pegvisomant: Implications for
cancer therapy. Clin Cancer Res. 13:1000–1009. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Hobbs JR and Salih H: Prolactin dependence
in human breast cancer. Proc R Soc Med. 66:8661973.PubMed/NCBI
|
|
113
|
Ormandy CJ, Hall RE, Manning DL, Robertson
JF, Blamey RW, Kelly PA, Nicholson RI and Sutherland RL:
Coexpression and cross-regulation of the prolactin receptor and sex
steroid hormone receptors in breast cancer. J Clin Endocrinol
Metab. 82:3692–3699. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Queiroga FL, Pérez-Alenza MD, Silvan G,
Peña L, Lopes C and Illera JC: Role of steroid hormones and
prolactin in canine mammary cancer. J Steroid Biochem Mol Biol.
94:181–187. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Visan S, Balacescu O, Berindan-Neagoe I
and Catoi C: In vitro comparative models for canine and human
breast cancers. Clujul Med. 89:38–49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Uva P, Aurisicchio L, Watters J, Loboda A,
Kulkarni A, Castle J, Palombo F, Viti V, Mesiti G, Zappulli V, et
al: Comparative expression pathway analysis of human and canine
mammary tumors. BMC Genomics. 10:1352009. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Egenvall A, Bonnett BN, Ohagen P, Olson P,
Hedhammar A and von Euler H: Incidence of and survival after
mammary tumors in a population of over 80,000 insured female dogs
in Sweden from 1995 to 2002. Prev Vet Med. 69:109–127. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Rivera P, Melin M, Biagi T, Fall T,
Häggström J, Lindblad-Toh K and von Euler H: Mammary tumor
development in dogs is associated with BRCA1 and BRCA2. Cancer Res.
69:8770–8774. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Ochiai K, Morimatsu M, Tomizawa N and
Syuto B: Cloning and sequencing full length of canine Brca2 and
Rad51 cDNA. J Vet Med Sci. 63:1103–1108. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Nieto A, Pérez-Alenza MD, Del Castillo N,
Tabanera E, Castaño M and Peña L: BRCA1 expression in canine
mammary dysplasias and tumours: Relationship with prognostic
variables. J Comp Pathol. 128:260–268. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Klopfleisch R and Gruber AD: Increased
expression of BRCA2 and RAD51 in lymph node metastases of canine
mammary adenocarcinomas. Vet Pathol. 46:416–422. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Rajan JV, Marquis ST, Gardner HP and
Chodosh LA: Developmental expression of Brca2 colocalizes with
Brca1 and is associated with proliferation and differentiation in
multiple tissues. Dev Biol. 184:385–401. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Rakha EA, El-Sayed ME, Green AR, Lee AH,
Robertson JF and Ellis IO: Prognostic markers in triple-negative
breast cancer. Cancer. 109:25–32. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Gama A, Gärtner F, Alves A and Schmitt F:
Immunohistochemical expression of Epidermal Growth Factor Receptor
(EGFR) in canine mammary tissues. Res Vet Sci. 87:432–437. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Queiroga FL, Perez-Alenza D, Silvan G,
Peña L and Illera JC: Positive correlation of steroid hormones and
EGF in canine mammary cancer. J Steroid Biochem Mol Biol. 115:9–13.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Ross JS, Fletcher JA, Linette GP, Stec J,
Clark E, Ayers M, Symmans WF, Pusztai L and Bloom KJ: The Her-2/neu
gene and protein in breast cancer 2003: Biomarker and target of
therapy. Oncologist. 8:307–325. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Rungsipipat A, Tateyama S, Yamaguchi R,
Uchida K, Miyoshi N and Hayashi T: Immunohistochemical analysis of
c-yes and c-erbB-2 oncogene products and p53 tumor suppressor
protein in canine mammary tumors. J Vet Med Sci. 61:27–32. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Kerns BJ, Pence JC, Huper G, Kinney RB and
Iglehart JD: c-erbB-2 expression in breast cancer detected by
immunoblotting and immunohistochemistry. J Histochem Cytochem.
38:1823–1830. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Paredes J, Lopes N, Milanezi F and Schmitt
FC: P-cadherin and cytokeratin 5: Useful adjunct markers to
distinguish basal-like ductal carcinomas in situ. Virchows Arch.
450:73–80. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Abd El-Rehim DM, Pinder SE, Paish CE, Bell
J, Blamey RW, Robertson JF, Nicholson RI and Ellis IO: Expression
of luminal and basal cytokeratins in human breast carcinoma. J
Pathol. 203:661–671. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Gama A, Alves A and Schmitt F:
Identification of molecular phenotypes in canine mammary carcinomas
with clinical implications: Application of the human
classification. Virchows Arch. 453:123–132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Gerdes J, Lemke H, Baisch H, Wacker HH,
Schwab U and Stein H: Cell cycle analysis of a cell
proliferation-associated human nuclear antigen defined by the
monoclonal antibody Ki-67. J Immunol. 133:1710–1715.
1984.PubMed/NCBI
|
|
133
|
Peña LL, Nieto AI, Pérez-Alenza D, Cuesta
P and Castaño M: Immunohistochemical detection of Ki-67 and PCNA in
canine mammary tumors: Relationship to clinical and pathologic
variables. J Vet Diagn Invest. 10:237–246. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Railo M, Nordling S, von Boguslawsky K,
Leivonen M, Kyllönen L and von Smitten K: Prognostic value of Ki-67
immunolabelling in primary operable breast cancer. Br J Cancer.
68:579–583. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Thomas M, Noguchi M, Kitagawa H, Kinoshita
K and Miyazaki I: Poor prognostic value of proliferating cell
nuclear antigen labelling index in breast carcinoma. J Clin Pathol.
46:525–528. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Funakoshi Y, Nakayama H, Uetsuka K,
Nishimura R, Sasaki N and Doi K: Cellular proliferative and
telomerase activity in canine mammary gland tumors. Vet Pathol.
37:177–183. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Kumaraguruparan R, Prathiba D and Nagini
S: Of humans and canines: Immunohistochemical analysis of PCNA,
Bcl-2, p53, cytokeratin and ER in mammary tumours. Res Vet Sci.
81:218–224. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Velculescu VE and El-Deiry WS: Biological
and clinical importance of the p53 tumor suppressor gene. Clin
Chem. 42:858–868. 1996.PubMed/NCBI
|
|
139
|
Beenken SW, Grizzle WE, Crowe DR, Conner
MG, Weiss HL, Sellers MT, Krontiras H, Urist MM and Bland KI:
Molecular biomarkers for breast cancer prognosis: Coexpression of
c-erbB-2 and p53. Ann Surg. 233:630–638. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Lee CH and Kweon OK: Mutations of p53
tumor suppressor gene in spontaneous canine mammary tumors. J Vet
Sci. 3:321–325. 2002.PubMed/NCBI
|
|
141
|
Chu LL, Rutteman GR, Kong JM, Ghahremani
M, Schmeing M, Misdorp W, van Garderen E and Pelletier J: Genomic
organization of the canine p53 gene and its mutational status in
canine mammary neoplasia. Breast Cancer Res Treat. 50:11–25. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Veldhoen N, Watterson J, Brash M and
Milner J: Identification of tumour-associated and germ line p53
mutations in canine mammary cancer. Br J Cancer. 81:409–415. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Muto T, Wakui S, Takahashi H, Maekawa S,
Masaoka T, Ushigome S and Furusato M: p53 gene mutations occurring
in spontaneous benign and malignant mammary tumors of the dog. Vet
Pathol. 37:248–253. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Moll UM and Slade N: p63 and p73: Roles in
development and tumor formation. Mol Cancer Res. 2:371–386.
2004.PubMed/NCBI
|
|
145
|
McKeon FD: p63 and p73 in tumor
suppression and promotion. Cancer Res Treat. 36:6–12. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Matos I, Dufloth R, Alvarenga M, Zeferino
LC and Schmitt F: p63, cytokeratin 5, and P-cadherin: Three
molecular markers to distinguish basal phenotype in breast
carcinomas. Virchows Arch. 447:688–694. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Stefanou D, Batistatou A, Nonni A,
Arkoumani E and Agnantis NJ: p63 expression in benign and malignant
breast lesions. Histol Histopathol. 19:465–471. 2004.PubMed/NCBI
|
|
148
|
Desantis CRS and Jemal A: Breast cancer
facts & figures 2015–2016. Am Cancer Soc. 44:2015.
|
|
149
|
Gama A, Alves A, Gartner F and Schmitt F:
p63: A novel myoepithelial cell marker in canine mammary tissues.
Vet Pathol. 40:412–420. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Elmore JG, Armstrong K, Lehman CD and
Fletcher SW: Screening for breast cancer. JAMA. 293:1245–1256.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Morgan MP, Cooke MM and McCarthy GM:
Microcalcifications associated with breast cancer: An epiphenomenon
or biologically significant feature of selected tumors? J Mammary
Gland Biol Neoplasia. 10:181–187. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Muttarak M, Kongmebhol P and Sukhamwang N:
Breast calcifications: Which are malignant? Singapore Med J.
50:907–914. 2009.PubMed/NCBI
|
|
153
|
Burnside ES, Sickles EA, Bassett LW, Rubin
DL, Lee CH, Ikeda DM, Mendelson EB, Wilcox PA, Butler PF and D'Orsi
CJ: The ACR BI-RADS experience: Learning from history. J Am Coll
Radiol. 6:851–860. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Ernster VL and Barclay J: Increases in
ductal carcinoma in situ (DCIS) of the breast in relation to
mammography: A dilemma. J Natl Cancer Inst Monogr. 151–156. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Mohammed SI, Meloni GB, Parpaglia Pinna
ML, Marras V, Burrai GP, Meloni F, Pirino S and Antuofermo E:
Mammography and ultrasound imaging of preinvasive and invasive
canine spontaneous mammary cancer and their similarities to human
breast cancer. Cancer Prev Res (Phila). 4:1790–1798. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Cekanova M and Rathore K: Animal models
and therapeutic molecular targets of cancer: Utility and
limitations. Drug Des Devel Ther. 8:1911–1921. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Decker WK, da Silva RF, Sanabria MH,
Angelo LS, Guimarães F, Burt BM, Kheradmand F and Paust S: Cancer
immunotherapy: Historical perspective of a clinical revolution and
emerging preclinical animal models. Front Immunol. 8:8292017.
View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Estrela-Lima A, Araújo MS, Costa-Neto JM,
Teixeira-Carvalho A, Barrouin-Melo SM, Cardoso SV, Martins-Filho
OA, Serakides R and Cassali GD: Immunophenotypic features of tumor
infiltrating lymphocytes from mammary carcinomas in female dogs
associated with prognostic factors and survival rates. BMC Cancer.
10:2562010. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Akiyama F and Horii R: Therapeutic
strategies for breast cancer based on histological type. Breast
Cancer. 16:168–172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Sorenmo KU, Kristiansen VM, Cofone MA,
Shofer FS, Breen AM, Langeland M, Mongil CM, Grondahl AM, Teige J
and Goldschmidt MH: Canine mammary gland tumours; a histological
continuum from benign to malignant; clinical and histopathological
evidence. Vet Comp Oncol. 7:162–172. 2009. View Article : Google Scholar : PubMed/NCBI
|