Introduction
Colorectal cancer (CRC) is one of the most common
types of cancer worldwide (1). The
changes in the Chinese diet and lifestyle have contributed to CRC
in becoming the third leading cause of cancer mortality in China
(2). Despite recent advances in the
surgical treatment, chemoprevention has emerged as an indispensable
option for the treatment of CRC (3).
The poor prognosis of patients with CRC is primarily due to primary
tumor spread and metastasis (4).
Therefore, cancer metastasis is one of the important causes of
mortality in patients with CRC (5).
Plant-derived compounds are a major source for
cancer therapy. Triterpenoids have attracted much attention for
their effective antitumor activities. Asiatic acid (AA; 2α, 3β,
23-trihydroxy-12-ursen-28-oic acid,
C30H48O5) (4,6,7), a naturally occurring pentacyclic
triterpenoid, is present in a variety of plants, including
Centella asiatica, Purnella vulgaris, Nepeta hindostana,
Eucalyptus perriniana and Psidium guajava. AA was
primarily used as a dermatological topical agent to prevent
ultraviolet A-mediated photo-aging and wounding healing (8). Previous studies suggested that AA
possessed a wide range of pharmacological properties, including
antihepatofibric, neuroprotective, anti-inflammatory, anti-diabetic
and antitumor activities (9). As for
antitumor effect, AA was reported to induce apoptosis in hepatic
and breast cancer cell lines and other tumor cells (10,11). The
efficacy of AA was possibly attributed to the inhibition of nuclear
factor-κB, p38 mitogen-activated protein kinase and extracellular
signal-regulated kinase, as well as the change of Bcl-2 and caspase
family proteins. Despite the aforementioned widely described
antitumor properties, further studies are required to investigate
the anti-metastatic potential of AA and to investigate the
molecular mechanisms of apoptosis induction and metastasis
inhibition.
In the present study, apoptosis and antitumor
effects of AA on human colon cancer cells (SW480 and HCT116), was
evaluated, and this revealed the role of phosphoinositide 3-kinase
(PI3K)/protein kinase B (Akt) signaling pathway in the underlying
molecular mechanism.
Materials and methods
Materials
RPMI-1640 with 10% heat-inactivated fetal bovine
serum (FBS) were purchased from Biological Industries (Kibbutz
Beit-Haemek, Israel). Rapamycin was purchased from Selleck
Chemicals (Houston, TX, USA). Antibodies were purchased from Cell
Signaling Technology (Danvers, MA, USA). Hoechst 33342 dye was
purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Fluorescein isothiocyanate (FITC)-labeled Annexin V and propidium
iodide (PI) were purchased from Nanjing KeyGen Biotech. Co., Ltd.
(Nanjing, China).
AA samples
AA previously isolated from the urban of Centella
asiatica (Umbelliferae) was used (12). These compounds used for the present
study were analyzed by high-performance liquid chromatography and
were >97% pure. AA was dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich; Merck KGaA) to make a stock solution at a
concentration of 1 mg/ml, which was further diluted to the
appropriate concentration with culture medium prior to each
experiment.
Cell line and cell culture
The human colon cancer cells (SW480 and HCT116) were
obtained from the Cancer Research Institute, Southern Medical
University (Guangzhou, China). The cells were incubated in
RPMI-1640 added with either 10% heat-inactivated FBS (both from
Biological Industries), at 37°C in a humidified atmosphere
containing 5% CO2.
Cell viability assay
Cell viability was measured by MTT assay; cells
without AA treatment were used as the control group.
(Sigma-Aldrich; Merck KGaA). Subsequently, the cells
(3×103) were seeded into 96-well plates (Nest
Biotechnology Co., Ltd., Wuxi, China) and allowed to adhere
following culture. After the cells were treated with 0, 10, 20, 30,
40 and 50 µg/ml of AA for various time-points from 24 to 72 h. A
total of 15 µl MTT solution was added to each well for an
additional 4 h at 37°C, and 150 µl DMSO was also added into each
well, followed by incubation at 37°C for 10 min with gentle
shaking. The absorbance value of each well was measured at 490 nm
using a spectrophotometer (ELx800; BioTek Instruments, Inc.,
Winooski, VT, USA). The experiments were performed in
triplicate.
Assessment of cell morphology
Abnormalities in cell morphology were observed using
an optical microscope. Briefly, 5×105 cells were seeded
into each well of 6-well plates (Nest Biotechnology Co., Ltd.) and
allowed to adhere following culture. Subsequently, the cells were
treated with 0, 15 and 25 µg/ml of AA for 24 h at 37°C, and
observed under an optical microscope (magnification, ×100; Nikon
TE2000; Nikon Corporation, Tokyo, Japan).
Colony formation assay
Cell proliferation was detected by colony formation
assay. The cells were seeded into 6-well plates (Nest Biotechnology
Co., Ltd.) at 2×102 cells/well. Following incubation for
24 h at 37°C, the medium was replaced by fresh medium with 0, 15
and 20 µg/ml AA respectively. When there were visible colonies,
incubation was stopped and the cells were stained. The number of
colonies was under an optical microscope.
Migration assay
For migration assay, 1×102 cells in 200
µl RPMI-1640 were seeded into upper Transwell chamber with a 8.0-µm
pore polycarbonate membrane insert (Nest Biotechnology Co., Ltd.).
RPMI-1640 (600 µl) with 10% FBS was added into the lower chamber as
a chemoattractant. After the cells were incubated for 8 h at 37°C,
the insert was washed with PBS, and the cells on the upper surface
of the insert were gently removed by a cotton swab. The cells
adhered to the lower surface were fixed by methanol, stained using
0.1% crystal violet at room temperature for 20 min and counted
under an optical microscope in 6–8 predetermined fields. For each
experiment, three independent filters were analyzed.
Analysis of apoptosis
The cells were distributed into control and
treatment groups with AA at 0, 15 and 20 µg/ml for 24 h. Following
washing with PBS for 1–2 times, 5 µg/ml Hoechst 33342 dye was added
to the cells. The cells were subsequently incubated in the dark for
5 min at room temperature. Then, the cells were observed with a
fluorescence microscope (Nikon TE2000; Nikon Corporation) and were
captured at 350 nm (excitation) and 460 nm (emission).
For analysis of apoptotic rate, flow cytometry (BD
Biosciences, Franklin Lakes, NJ, USA) was used. Each cell line was
maintained at a density of 2×105 cells in 6-well plates.
Following treatment with AA, the cells were harvested, rinsed with
ice-cold PBS, and were treated for 30 min with FITC-labeled Annexin
V and PI (Nanjing KeyGen Biotech. Co. Ltd.) for 10 min at room
temperature in the dark. Binding buffer (400 µl) was added to each
tube according to the supplier's protocols, and the cells were
analyzed with a flow cytometer within 1 h.
The cells were distributed into 6-well plates, and
each cell group had three wells. Following incubation for 12 h at
37°C, the cells were treated with AA from 0 to 25 µg/ml,
respectively. To evaluate the DNA content, the nuclei were fixed in
methanol, stained with 15 µg/ml of 4, 60-diamidino-2-phenylindole
and 100 µg/ml DNase free RNase A for 30 min at 37°C, rinsed twice
with PBS. The DNA content of labeled cells was analyzed using
fluorescence-activated cell sorting cytometry assay (BD
Biosciences). All experiments were performed in triplicate.
Western blotting
The cells were harvested and lysed. 20 µg of
protein/lane was separated by 10% SDS PAGE and transferred to
polyvinylidene fluoride (PVDF) membranes. The membranes were
incubated with the primary antibodies (all rabbit polyclonal
antibodies were used at dilution, 1:1,000) including
anti-epithelial (E-) cadherin (catalog no. 2500), anti-neural (N-)
cadherin (catalog no. 13116), anti-vimentin (catalog no. 12020),
anti-phosphorylated (-p) PI3K (catalog no. 3811), anti-PI3K
(catalog no. 3821), anti-p Akt (Ser473; catalog no.
5012), anti-Akt (catalog no. 4059), anti-p-mammalian target of
rapamycin (mTOR; catalog no. 2974), anti-mTOR (catalog no. 2972),
anti-p-p70S6K (catalog no. 9205), anti-p70S6K (catalog no. 9202),
anti-programmed cell death 4 (Pdcd4; catalog no. 9535S),
anti-β-actin (catalog no. 12620S) and anti-GAPDH (catalog no.
2118S) (all purchased from Cell Signaling Technology, Inc.)
overnight at 4°C, until they were blocked with 5% non-fat dried
milk for 1 h at room temperature. Then, the immunoreactive bands
were visualized by enhanced chemiluminescence using horseradish
peroxidase-conjugated immunoglobulin G secondary antibodies
(anti-rabbit; catalog no. 7074P2; dilution, 1:10,000; Cell
Signaling Technology, Inc.). The bands were visualized by using an
ECL western blot kit (Kangwei Biotech Co., Ltd., Beijing, China).
The images were captured with ChemiDoc™ CRS and Molecular Imager
(Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
All data are expressed as the mean value ± standard
error from at least three independent experiments. The comparisons
of different groups were performed using Student's t-test and
one-way of variance (ANOVA). Statistical analyses were performed
with the SPSS 13.0 statistical software package (SPSS Inc.,
Chicago, IL, USA). All statistical tests were two-sided, and
P<0.05 was considered to be statistically significant.
Results
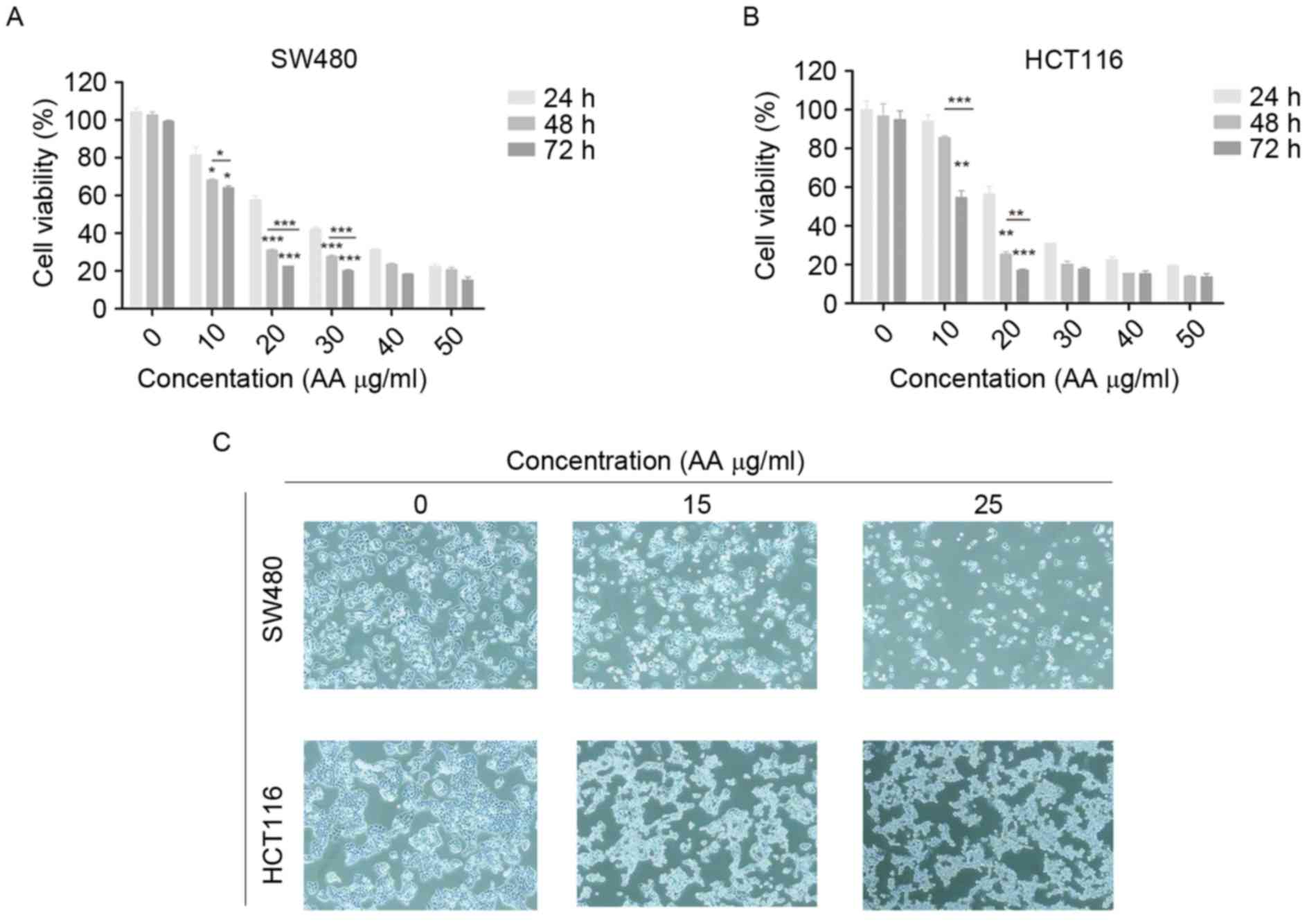
Inhibition of viability of colon
carcinoma cells by AA
MTT assay was used to determine the effects of AA on
the viability of SW480 and HCT116 cells. The cells were treated
with various concentrations of AA (0–50 µg/ml) for 24, 48, and 72
h. AA significantly inhibited the growth of SW480 and HCT116 cells
in a dose- and time-dependent manner (Fig. 1A and B; P<0.05, P<0.01,
P<0.001). Therefore, various concentrations (15, 20, and 25
µg/ml) of AA were selected for subsequent studies.
AA alters the morphology of colon
carcinoma cells
The cells were treated with AA at 15 and 25 µg/ml
for 24 h. The cells in the control groups grew markedly, while the
cells in the drug-treated groups became small and pyknotic, and the
cytoplasm was condensed. The cell organelles in the AA-treated
groups were swollen, which confirmed that AA had cytotoxic effects
on human colon carcinoma cells (Fig.
1C).
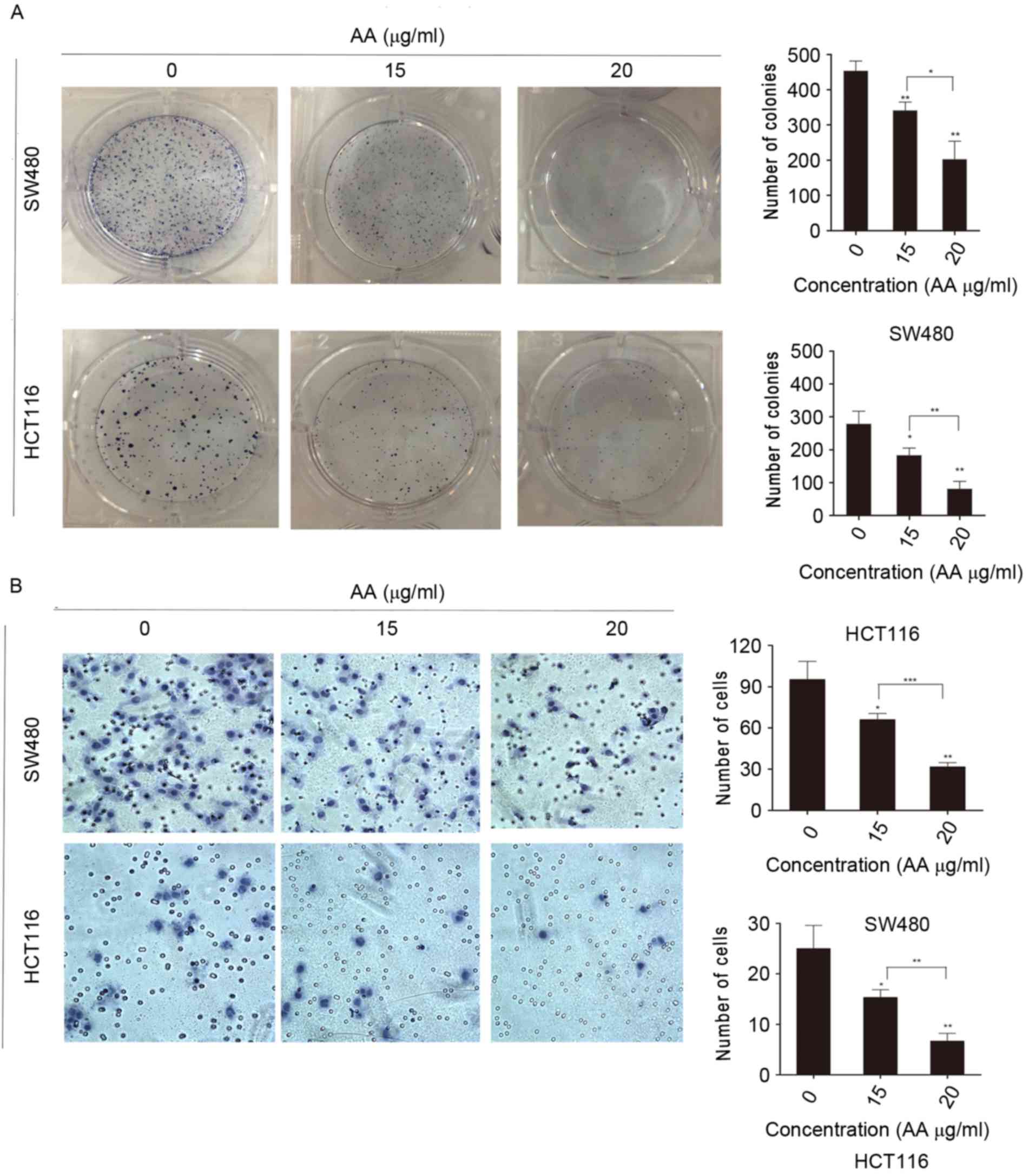
AA inhibits proliferation and colony
formation of colon carcinoma cells
Colony formation of SW480 and HCT116 cells was
inhibited by AA treatment (Fig. 2A).
As expected, these cell lines treated with AA presented markedly
reduced cell growth and colony formation compared with the control
cell lines. The number of colonies treated with AA in groups
significantly was reduced, which indicated that proliferation of
single cells was also decreased.
AA reduces migration of colon
carcinoma cells
To gain an insight into the role of AA in migration
and tumorigenesis of SW480 and HCT116 cells, the Transwell
migration assay was performed (Fig.
2B). It was demonstrated that the number of cells migrated in
AA-treated groups was significantly reduced compared with the
control. Together, the findings suggest that AA may reduce the
migration of SW480 and HCT116 cells. Additionally, the number of
SW480 cells migrated in the control group was higher compared to
the number in the HCT116 control group. This result indicated that
the migratory abilities of SW480 and HCT116 cells were
different.
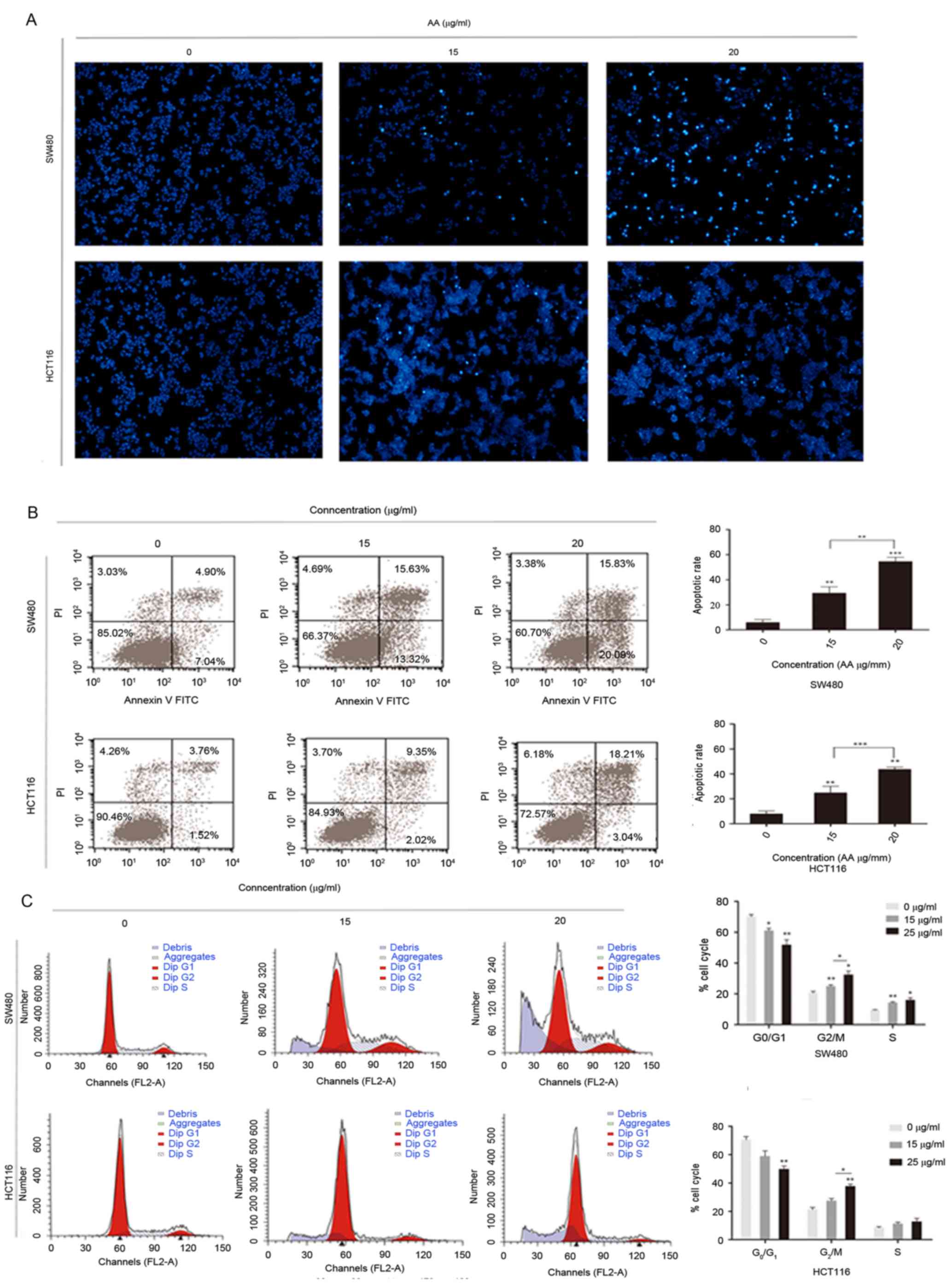
AA induces apoptosis of colon
carcinoma cells
To elucidate the underlying mechanism of apoptosis
in colorectal carcinoma cells, whether AA is able to induce
apoptosis in colon carcinoma cells was investigated by nuclear
staining with Hoechst 33342.
Cytoplasmic agglutination, which is a characteristic
of active apoptosis, was observed in SW480 and HCT116 cells, which
have been treated for 24 h with 15 and 25 µg/ml AA. Cytoplasmic
agglutination was not observed in the control groups (Fig. 3A). Rates of apoptosis and cell cycle
distribution were determined by flow cytometry. Flow cytometric
analysis indicated the number of apoptotic cells was markedly
higher in the AA-treated groups compared with the control, and this
increase was concentration-dependent (Fig. 3B). Additionally, the rate of apoptosis
of the AA-treated groups were significantly higher compared with
the control. Notably, an increase in apoptosis was observed in the
sub-G1 population of SW480 and HCT116 cells treated with AA. There
were also marked changes in cell cycle distribution with a
decreased percentage of cells in the G1 phase and an increased
percentage of cells in the G2/M and S phases, compared with the
control (Fig. 3C) (P<0.05).
Effects of AA on
epithelial-mesenchymal transition (EMT) marker protein expression
in colon carcinoma cells
The expression of EMT-interrelated factors, as
initiators of the metastatic cascade in the tumor cells, including
vimentin, N-cadherin and E-cadherin were evaluated in the present
study (Fig. 4A). Consistent with the
findings of the Transwell migration assay, it was observed that the
level of E-cadherin expression was increased, while the expression
of vimentin and N-cadherin was reduced in the AA-treated groups.
These results indicated that AA-induced inhibition on migration in
colon carcinoma cells may be associated with EMT.
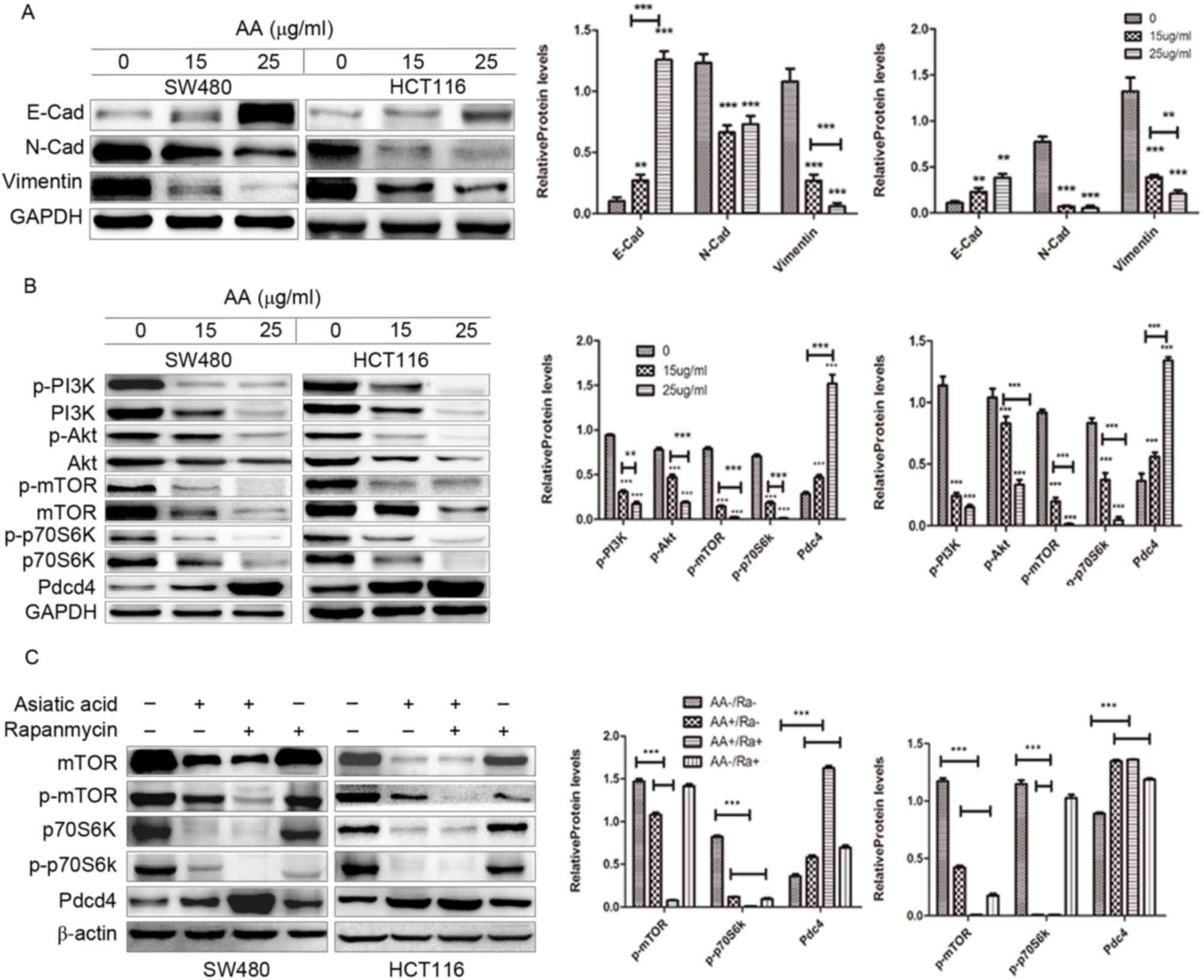 | Figure 4.Western blot analysis of the effects
of asiatic acid on the PI3K/Akt/mTOR/p70S6K/Pdcd4 signaling pathway
in SW480 and HCT116 cells, and densitometric quantification of the
blots compared with GAPDH/β-actin. (A) The expression level of
E-cadherin was increased while vimentin and N-cadherin expression
was reduced. (B) The results showed downregulation of p-PI3K,
p-Akt, p-mTOR, p-p70S6K and upregulation of Pdcd4. (C) Pretreatment
with asiatic acid and rapamycin further decreased the expression of
mTOR and p70S6K and increased the expression of Pdcd4. **P<0.01,
***P<0.001. Asterisks alone indicate comparison of the control
group with 15 µg/ml, 25 µg/ml AA, respectively. The underlined
asterisks indicate a comparison between 15 µg/ml AA and 25 µg/ml
AA. PI3K, phosphoinositide 3-kinase; mTOR, mammalian target of
rapamycin; p70S6K, ribosomal protein S6 kinase β-1; Pdcd4,
programmed cell death protein 4; p, phosphorylated; AA, asiatic
acid; Ra, rapamycin. |
AA meditates anticancer effects by
regulating Pdcd4 via the PI3K/Akt/mTOR/p70S6K signaling
pathway
The PI3K/Akt/mTOR/p70S6K signaling pathway is one of
the major pathways, which regulate proliferation, apoptosis and
migration of cancer cells (13).
Therefore, whether AA affects this pathway in colon carcinoma cells
was investigated. As shown in Fig.
4B, treatment with AA markedly decreased the level of p-PI3K,
p-Akt (Ser473), p-mTOR and p-p70S6K in a concentration dependent
manner.
Additionally, the expression of Pdcd4 protein,
downstream factor of p70S6K, was increased following treatment with
AA. The mTOR inhibitor rapamycin was subsequently used to
investigate whether the PI3K/Akt/mTOR/p70S6K signaling pathway is
involved in the proliferation, migration and apoptosis of colon
cancer cells. It was observed that pre-treating cells with AA and
rapamycin further decreased the expression of mTOR and p70S6K, and
increased the expression of Pdcd4, compared with untreated cells
(Fig. 4C). This finding suggests that
AA partially exerts anticancer effect by regulating Pdcd4 via the
PI3K/Akt/mTOR/p70S6K signaling pathway in colon cancer cells.
Discussion
In the present study, a novel role for AA on
antitumor activity of colon cancer cells was revealed. It was
demonstrated that AA was able to induce apoptosis and cell cycle
arrest of colon carcinoma cells. Additionally, AA was able to
modulate the expression of EMT marker proteins colon carcinoma
cells. The expression of PI3K, Akt, mTOR, p70S6K total protein and
phosphorylated proteins in AA-treated colon carcinoma cells was
significantly reduced compared with the control group. This is in
contrast to the upregulation of Pdcd4 expression observed in the
AA-treated cells compared to the control group. This suggests that
AA induced apoptosis by partially regulating Pdcd4 through the
PI3K/Akt/mTOR/p70S6K signaling pathway.
AA has been reported to have an essential role in
apoptosis, survival and proliferation in various types of
carcinoma, including human hepatoma, breast cancer, non-small cell
lung cancer cells, prostate cancer, glioblastoma, human melanoma,
multiple myeloma, B-cell lymphoma and neuroblastoma cells (14–18). The
mechanisms of apoptosis primarily involve two signaling pathways:
The mitochondrial death pathway and the cell death receptor
pathway. Tang et al (19)
previously demonstrated that AA is able to induce inhibition of
growth of colon cancer cells and apoptosis through the
mitochondrial death cascade. In the present study, it was found
that AA was able to induce colon cancer cell growth and apoptosis
by regulating the inhibition of the PI3K/Akt/mTOR/p70S6K signaling
pathway, therefore suggesting a new potential mechanism for the
effect of AA on apoptosis of colon carcinoma cells. Furthermore, it
was indicated that AA may inhibit proliferation and migration of
colon cancer cells.
As a critical event in the development of carcinoma
cells, EMT, is a process whereby epithelial cells first lose
cell-cell adhesion and cell polarity, followed by an increase in
migratory and invasive properties to become mesenchymal stem cells
(20,21). E-cadherin, N-cadherin and vimentin are
three important promoters, which mediate migration and invasion of
cancer cells through a number of signaling pathways (22,23).
Therefore, the present study investigated the expression of
EMT-associated factors. As expected, AA markedly increased the
level of E-cadherin and decreased the levels of N-cadherin and
vimentin in SW480 and HCT116, indicating the involvement of EMT in
AA-induced inhibition of colon carcinoma cell migration.
PI3K/Akt/mTOR/P70S6K is an important signaling
pathway, which regulates progression in numerous tumors (24). This pathway controls cell
proliferation, growth, translation, migration and survival, and
over-activation of the signaling pathway is associated with poor
prognosis (24). There are a number
of anticancer drugs, which mediate anticancer activity via the
PI3K/Akt/mTOR/p70S6K axis. For example, by targeting the
insulin-like growth factor 1 receptor, growth and metastasis can be
significantly inhibited through suppression of the
PI3K/Akt/mTOR/p70S6K signaling pathway (25).
In addition, bone morphogenetic protein 9 has been
shown to inhibit the proliferation and migration of breast cancer
cells, and it was demonstrated that the PI3K/Akt/mTOR/p70S6K
signaling pathway was involved in this process (25). Furthermore, activation of the
PI3K/Akt/mTOR/p70S6K signaling pathway has been shown to be
involved in S100A4-induced viability and migration in CRC cells
(26).
Pdcd4 is a novel tumor suppressor protein that
inhibits protein synthesis by suppressing translation (27,28). The
PI3K/Akt/mTOR signaling pathway constitutively represses Pdcd4
expression in acute myelocytic leukemia. All-trans retinoic acid
induces Pdcd4 through inhibition of the PI3K/Akt/mTOR signaling
pathway (29). Pdcd4 has also been
shown to inhibit invasion by activating activator protein-1
(30). Pdcd4 and PI3K/Akt/mTOR/p70S6K
constitute an important pathway, which modulates multiple
biological processes in carcinoma cells (31). A limited number of studies have
focused on the association between the PI3K/Akt signaling pathway
and cancer cells that exhibit increased expression of Pdc4,
including colon carcinoma cells. Notably, the present study found
that AA was able to inhibit the PI3K/Akt/mTOR signaling pathway,
which leads to apoptosis, an increase in the level of Pdcd4
expression and inhibition of cell invasion.
Taken together, the findings suggest that AA
modulates the PI3K/AKT/mTOR signaling pathway, which affects cell
proliferation, growth, translation, migration and survival. To the
best of our knowledge, the present study is the first to describe
the induction of apoptosis by AA by regulating Pdcd4 through the
inhibition of the PI3K/Akt/mTOR/p70S6K signaling pathway Although
therapeutics for the treatment of colon carcinoma remains at
infancy, the present study suggests that AA may be a novel
drug.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81202430), Pearl
River S and T Nova Program of Guangzhou (grant no. 2014J2200092)
and the Science and Technology Planning Project of Guangdong (grant
no. 2014A020221012).
References
|
1
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Du C, Huang D, Peng Y, Yao Y, Zhao Y, Yang
Y, Wang H, Cao L, Zhu WG and Gu J: 5-Fluorouracil targets histone
acetyltransferases p300/CBP in the treatment of colorectal cancer.
Cancer Lett. 400:183–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Woo IS and Jung YH: Metronomic
chemotherapy in metastatic colorectal cancer. Cancer Lett.
400:319–324. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weidle UH, Birzele F and Krüger A:
Molecular targets and pathways involved in liver metastasis of
colorectal cancer. Clin Exp Metastasis. 32:623–635. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nasir MN, Habsah M, Zamzuri I, Rammes G,
Hasnan J and Abdullah J: Effects of asiatic acid on passive and
active avoidance task in male spraque-dawley rats. J
Ethnopharmacol. 134:203–209. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang X, Wu J, Dou Y, Xia B, Rong W,
Rimbach G and Lou Y: Asiatic acid protects primary neurons against
C2-ceramide-induced apoptosis. Eur J Pharmacol. 679:51–59. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee Soo Y, Jin DQ, Beak SM, Lee ES and Kim
JA: Inhibition of ultraviolet-A-modulated signaling pathways by
asiatic acid and ursolic acid in HaCaT human keratinocytes. Eur J
Pharmacol. 476:173–178. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee YS, Jin DQ, Kwon EJ, Park SH, Lee ES,
Jeong TC, Nam DH, Huh K and Kim JA: Asiatic acid, a triterpene,
induces apoptosis through intracellular Ca2+ release and
enhanced expression of p53 in HepG2 human hepatoma cells. Cancer
Lett. 186:83–91. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hsu YL, Kuo PL, Lin LT and Lin CC: Asiatic
acid, a triterpene, induces apoptosis and cell cycle arrest through
activation of extracellular signal-regulated kinase and p38
mitogen-activated protein kinase pathways in human breast cancer
cells. J Pharmacol Exp Ther. 313:333–344. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Adtani PN, Narasimhan M, Punnoose AM and
Kambalachenu HR: Antifibrotic effect of Centella asiatica Linn and
asiatic acid on arecoline-induced fibrosis in human buccal
fibroblasts. J Investig Clin Dent. 8:2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang L, Xu J, Zhao C, Zhao L and Feng B:
Antiproliferative, cell-cycle dysregulation effects of novel
asiatic acid derivatives on human non small cell lung cancer cells.
Chem Pharm Bull (Tokyo). 61:1015–1023. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gurfinkel DM, Chow S, Hurren R, Gronda M,
Henderson C, Berube C, Hedley DW and Schimmer AD: Disruption of the
endoplasmic reticulum and increases in cytoplasmic calcium are
early events in cell death induced by the natural triterpenoid
Asiatic acid. Apoptosis. 11:1463–1471. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thakor FK, Wan KW, Welsby PJ and Welsby G:
Pharmacological effects of asiatic acid in glioblastoma cells under
hypoxia. Mol Cell Biochem. 430:179–190. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kavitha CV, Jain AK, Agarwal C, Pierce A,
Keating A, Huber KM, Serkova NJ, Wempe MF, Agarwal R and Deep G:
Asiatic acid induces endoplasmic reticulum stress and apoptotic
death in glioblastoma multiforme cells both in vitro and in vivo.
Mol Carcinog. 54:1417–1429. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park BC, Bosire KO, Lee ES, Lee YS and Kim
JA: Asiatic acid induces apoptosis in SK-MEL-2 human melanoma
cells. Cancer Lett. 218:81–90. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu Q, Lv T, Chen Y, Wen L, Zhang J, Jiang
X and Liu F: Apoptosis of HL-60 human leukemia cells induced by
Asiatic acid through modulation of B-cell lymphoma 2 family
proteins and the mitogen-activated protein kinase signaling
pathway. Mol Med Rep. 12:1429–1434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu MF, Xiong YY, Liu JK, Qian JJ, Zhu L
and Gao J: Asiatic acid, a pentacyclic triterpene in Centella
asiatica, attenuates glutamate-induced cognitive deficits in mice
and apoptosis in SH-SY5Y cells. Acta Pharmacol Sin. 33:578–587.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tang XL, Yang XY, Jung HJ, Kim SY, Jung
SY, Choi DY, Park WC and Park H: Asiatic acid induces colon cancer
cell growth inhibition and apoptosis through mitochondrial death
cascade. Biol Pharm Bull. 32:1399–1405. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kiesslich T, Pichler M and Neureiter D:
Epigenetic control of epithelial-mesenchymal-transition in human
cancer. Mol Clin Oncol. 1:3–11. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gheldof A and Berx G: Cadherins and
epithelial-to-mesenchymal transition. Prog Mol Biol Transl Sci.
116:317–336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bao B, Wang Z, Ali S, Kong D, Li Y, Ahmad
A, Banerjee S, Azmi AS, Miele L and Sarkar FH: Notch-1 induces
epithelial-mesenchymal transition consistent with cancer stem cell
phenotype in pancreatic cancer cells. Cancer Lett. 307:26–36. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zavadil J, Narasimhan M, Blumenberg M and
Schneider RJ: Transforming growth factor-beta and microRNA: mRNA
regulatory networks in epithelial plasticity. Cells Tissues Organs.
185:157–161. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Syed DN, Afaq F, Sarfaraz S, Khan N,
Kedlaya R, Setaluri V and Mukhtar H: Delphinidin inhibits cell
proliferation and invasion via modulation of Met receptor
phosphorylation. Toxicol Appl Pharmacol. 231:52–60. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ren W, Liu Y, Wan S, Fei C, Wang W, Chen
Y, Zhang Z, Wang T, Wang J, Zhou L, et al: BMP9 inhibits
proliferation and metastasis of HER2-positive SK-BR-3 breast cancer
cells through ERK1/2 and PI3K/AKT pathways. PLoS One. 9:e968162014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang H, Duan L, Zou Z, Li H, Yuan S, Chen
X, Zhang Y, Li X, Sun H, Zha H, et al: Activation of the
PI3K/Akt/mTOR/p70S6K pathway is involved in S100A4-induced
viability and migration in colorectal cancer cells. Int J Med Sci.
11:841–849. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wen YH, Shi X, Chiriboga L, Matsahashi S,
Yee H and Afonja O: Alterations in the expression of PDCD4 in
ductal carcinoma of the breast. Oncol Rep. 18:1387–1393.
2007.PubMed/NCBI
|
|
28
|
Afonja O, Juste D, Das S, Matsuhashi S and
Samuels HH: Induction of PDCD4 tumor suppressor gene expression by
RAR agonists, antiestrogen and HER-2/neu antagonist in breast
cancer cells. Evidence for a role in apoptosis. Oncogene.
23:8135–8145. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Leupold JH, Yang HS, Colburn NH, Asangani
I, Post S and Allgayer H: Tumor suppressor Pdcd4 inhibits
invasion/intravasation and regulates urokinase receptor (u-PAR)
gene expression via Sp-transcription factors. Oncogene.
26:4550–4562. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang HS, Jansen AP, Nair R, Shibahara K,
Verma AK, Cmarik JL and Colburn NH: A novel transformation
suppressor, Pdcd4, inhibits AP-1 transactivation but not NF-kappaB
or ODC transactivation. Oncogene. 20:669–676. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vikhreva PN, Shepelev MV and Korobko IV:
mTOR-dependent transcriptional repression of Pdcd4 tumor suppressor
in lung cancer cells. Biochim Biophys Acta. 1839:43–49. 2014.
View Article : Google Scholar : PubMed/NCBI
|