Introduction
Hereditary non-polyposis colorectal cancer (HNPCC),
also known as Lynch syndrome, is inherited as an autosomal dominant
disease and is the most common hereditary colorectal cancer,
accounting for ~50% of familial colorectal cancer and 3% of all
colorectal cancer cases (1). Unlike
with sporadic colorectal cancer, HNPCC is associated with specific
genetic factors and significant clinicopathological features. These
features are often associated with synchronous and metachronous
colorectal cancer and cause a high incidence of extraintestinal
malignant tumors, including endometrial, gastric, renal, pancreatic
and ovarian cancer types (2).
Inactivation of DNA mismatch repair (MMR) genes, including MLH1,
MSH2, MSH6 and PMS2, is the molecular genetic basis of
HNPCC pathogenesis. Mutation of MMR genes can result in loss of DNA
MMR function, leading to aberrant DNA replication, increased
spontaneous mutation frequency and microsatellite instability. This
ultimately leads to the transformation of normal cells into
malignant cells (3–5).
However, a previous study observed that certain
HNPCC patients, diagnosed by the presence of MMR gene mutations,
did not meet some of the clinical diagnostic criteria for HNPCC
(6). Furthermore, in certain patients
meeting the clinical diagnostic criteria for HNPCC, MMR gene
mutations could not be detected (7,8). Bashyam
et al (8) demonstrated that,
among 48 patients with Lynch syndrome, only 58% had MMR gene
expression defects, which indicated that other, as yet
unidentified, causative genes may be involved in the pathogenesis
of HNPCC.
Based upon this assumption, in the present study,
whole exome sequencing was performed in 3 HNPCC patients from 1
family and unreported mutations were observed in 15 gene loci.
Subsequently, peripheral blood was collected from control subjects,
sporadic colorectal cancer patients and the aforementioned 3 HNPCC
patients. Single nucleotide polymorphism (SNP) genotyping assays
were also performed on the aforementioned 15 genes using the DNA
MassARRAY Genetic Analysis system to further verify whether these
genes were associated with HNPCC pathogenesis.
Materials and methods
Blood sample collection
All procedures in studies involving human
participants were performed in accordance with the ethical
standards of the institutional and/or national research committee
and the 1964 Declaration of Helsinki and its later amendments or
comparable ethical standards. All patients signed informed consent
forms prior to participation in the study and the study was
approved by the Third Xiangya Hospital Ethics Committee (Changsha,
China).
Blood samples were collected from 96 subjects,
including 12 control subjects, 81 sporadic colorectal cancer
patients who were diagnosed by histopathology from January 2014 to
December 2016 at the Third Xiangya Hospital of Central South
University and 3 HNPCC patients from the aforementioned hospital
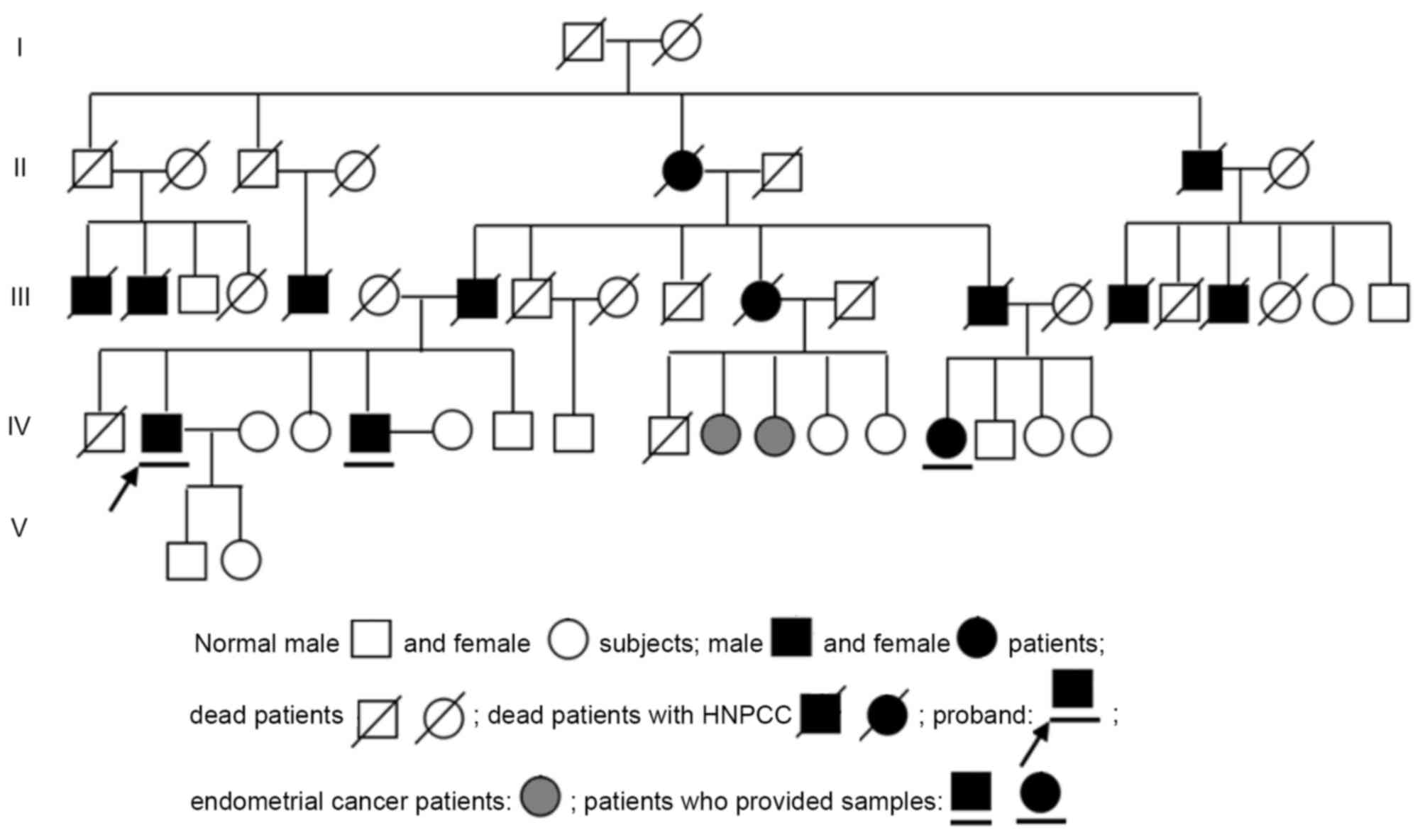
who met the Amsterdam Criteria (9),
which is outlined as follows: i) ≥3 colorectal cancer cases in the
same family diagnosed by histopathology, one case being a
first-degree relative (parent or sibling) of the other two cases;
ii) ≥2 successive generations affected; iii) ≥1 case with onset
prior to the age of 50 years; and iv) familial adenomatous
polyposis in HNPCC patients should be excluded. In the HNPCC family
investigated in the present study, the proband's father had
colorectal cancer that was diagnosed by histopathology and the
other 2 cases who provided samples were a sibling and a cousin of
the proband. The 3 patients experienced changes in their stools and
abdominal bloating prior to being hospitalized. Colorectal cancer
was diagnosed by histopathology (all pathology diagnoses were
confirmed by two deputy or chief director pathologists) following
radical surgery (Table I). The
pedigree of the HNPCC family is presented in Fig. 1.
 | Table I.Clinical characteristics of 3
hereditary non-polyposis colorectal cancer patients. |
Table I.
Clinical characteristics of 3
hereditary non-polyposis colorectal cancer patients.
| Sample name | Sex | Age, years | Main symptoms | Pathological
types |
|---|
| Lah | Male | 47 | Stool changes for 1
year, abdominal bloating for 2 months |
Moderately-differentiated
adenocarcinoma |
| Lyh | Male | 45 | Blood in stools,
abdominal bloating, weight loss and stool changes for 2 years |
Moderately-differentiated
adenocarcinoma |
| Lyl | Female | 42 | Abdominal bloating
and pain, hypodynamia and stool changes for 3 months. | Well-differentiated
adenocarcinoma |
Whole exome sequencing
DNA was extracted from the peripheral blood of 3
HNPCC cases and purified using a DNeasy Blood and Tissue kit (cat.
no. 69506; Qiagen, Inc., Valencia, CA, USA) according to the
manufacturer's protocols. Exome sequences were subjected to DNA
sequencing on the Illumina platform using Illumina PE Flow Cell
v3-HS (Illumina, Inc., San Diego, CA, USA). In accordance with the
manufacturer's protocols, genomic DNA fragments were processed by
end repair, addition of adenosine (A) to 3′ ends, ligation, DNA
enrichment and hybridization. DNA libraries from samples were
constructed. The concentration, purity and size of the libraries
were measured using an Agilent 2100 Bioanalyzer (Agilent
Technologies Inc., Santa Clara, CA, USA) and a Qubit®
2.0 Fluorometer (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
The hybridization of sequencing primers and the generation of
clusters were performed using cBot (HiSeq 2500; Illumina, Inc.)
following the cBot User Guide (Part #15006165; Rev. F; lllumina,
Inc.). A paired-end sequencing was then performed on a
cluster-containing flow cell following the manufacturer's protocols
(HiSeq 2500; Illumina, Inc.). Data acquisition software (Illumina,
Inc.) was used for quality control and data analysis. The quality
control standards for sequencing results were as follows: The
average coverage for an exon region was ~100 times; if the average
coverage was <90 times, it was resequenced; and at 100 times
coverage, ≥85% of exon regions were covered by ≥1 sequence
(Table II). The Burrows-Wheeler
Alignment software package (version 0.5.9; Shanghai Biotechnology,
China) was used to map sequences using human hg19 as the reference
genome. Potential PCR duplicates were removed using rmdup of
Samtools-0.1.18 (Shanghai Biotechnology, China), and mapping
statistics were generated using Samtools flagstat (Shanghai
Biotechnology, China) (Table III).
Capture-enrichment methods were used to determine the amount of
fragment from the captured target region and the coverage and depth
of the target region.
| Table II.Sequence quality control results for
3 hereditary non-polyposis colorectal cancer patients. |
Table II.
Sequence quality control results for
3 hereditary non-polyposis colorectal cancer patients.
| Sample name | Orientation | Total reads, n | Total bases, n | Q20, % | Depth | 1×, % | Quality of
sequencing results |
|---|
| Lah-1 | Forward | 48,356,713 | 4,835,671,300 | 98 | 105.69 | 99.95 | Good |
| Lah-2 | Reverse | 48,356,713 | 4,835,671,300 | 97 |
|
| Good |
| Lyl-1 | Forward | 73,411,952 | 7,341,195,200 | 97 | 149.87 | 99.86 | Good |
| Lyl-2 | Reverse | 73,411,952 | 7,341,195,200 | 95 |
|
| Good |
| Lyh-1 | Forward | 57,778,031 | 5,777,803,100 | 97 | 120.25 | 99.96 | Good |
| Lyh-2 | Reverse | 57,778,031 | 5,777,803,100 | 95 |
|
| Good |
| Table III.Sequence mapping information for 3
hereditary non-polyposis colorectal cancer patients. |
Table III.
Sequence mapping information for 3
hereditary non-polyposis colorectal cancer patients.
| Sample name | Filtered reads,
n | Mapped reads,
n | Map ratio, % | Unique mapped
reads, n | Unique mapped
ratio, % |
|---|
| Lyh | 107,719,930 | 105,656,069 | 98.08 | 95,653,932 | 88.80 |
| Lyl | 136,284,036 | 133,688,255 | 98.10 | 120,411,348 | 88.35 |
| Lah | 92,361,758 | 91,224,395 | 98.77 | 83,257,847 | 90.14 |
SNP genotyping
DNA extraction
DNA was extracted from peripheral blood lymphocytes
of the 96 samples using a DNeasy Blood and Tissue kit (Qiagen,
Inc.), according to the manufacturer's protocols. DNA was
quantified and assessed using a NanoDrop®ND-1000
spectrophotometer (NanoDrop Technologies, Inc., Thermo Fisher
Scientific, Inc., Wilmington, DE, USA) and 0.8% agarose gel
electrophoresis.
PCR amplification
PCR primer mixes were obtained from Invitrogen
(Invitrogen, Thermo Fisher Scientific, Inc.) (Table IV). The Q-PCR Detection Kit was
purchased from GeneCopoeia (Rockville, MD, USA). The First Strand
cDNA Synthesis kit was purchased from Fermentas (Thermo Fisher
Scientific, Inc.). Total PCR volume was 5 µl, including 1 µl
template DNA, 1.8 µl ddH2O, 0.5 µl 10X PCR Buffer, 0.1
µl 25 mmol/l dNTPs, 0.4 µl 25 mmol/l MgCl2, 1 µl PCR
Primer (0.5 mmol/l) and 0.2 µl Gold Tag PCR enzyme (Advanced
Biotechnologies Inc., Eldersburg, MD, USA). PCR conditions were
95°C for 2 min, then 45 cycles of 95°C for 30 sec, 50°C for 30 sec,
72°C for 1 min and 72°C for 5 min.
| Table IV.Names and sequences of polymerase
chain reaction primers. |
Table IV.
Names and sequences of polymerase
chain reaction primers.
| Name of primer | Sequence of
primer |
|---|
|
BB14228-3chr1_112298707-F |
ACGTTGGATGACCGCTCAGGATCTCAGCAG |
|
BB14228-3chr14_68264412-F |
ACGTTGGATGAGCAACCTTCCCGAAGATAC |
|
BB14228-3chr1_46509382-F |
ACGTTGGATGATCCTTGGTTCAGCACAACG |
|
BB14228-3chr6_35911730-F |
ACGTTGGATGCCCTCTGGTGAGTATGAATC |
|
BB14228-3chr2_145156750-F |
ACGTTGGATGAATTTTCAGCAGTTCATCGG |
|
BB14228-3chr8_95952304-F |
ACGTTGGATGCTGTTTACCGGCATCTCTTG |
|
BB14228-3chr2_219249005-F |
ACGTTGGATGCCTGAAGATCTGACTCGATG |
|
BB14228-3chr4_151827481-F |
ACGTTGGATGGAACTCAATTGCTATGCAGG |
|
BB14228-3chr2_37439069-F |
ACGTTGGATGACATCCATGAATGTTCCTCC |
|
BB14228-3chr2_67630823-F |
ACGTTGGATGGACAAATGCTTTGAAAGAGG |
|
BB14228-3chr3_52474994-F |
ACGTTGGATGTCACCTAGCTGGTCAAAGTG |
|
BB14228-3chr3_50325883-F |
ACGTTGGATGCAGAACAATGAGCTACTCCG |
|
BB14228-3chr12_88514827-F |
ACGTTGGATGTGAGAGAACAGCTGAACTGG |
|
BB14228-3chr4_187541196-F |
ACGTTGGATGAGTTATCGTTCCGATCACTG |
|
BB14228-3chr10_124221227-F |
ACGTTGGATGAGAGTCGCCATGCAGATCC |
|
BB14228-3chr1_112298707-R |
ACGTTGGATGAAAGCAGCAGTGACTCGAAG |
|
BB14228-3chr14_68264412-R |
ACGTTGGATGAGCTCTTCAGATTACCTGCC |
|
BB14228-3chr1_46509382-R |
ACGTTGGATGTATCTGCAAAGCGAGGGCAT |
|
BB14228-3chr6_35911730-R |
ACGTTGGATGATCATCCGTCTATGGCTTCC |
|
BB14228-3chr2_145156750-R |
ACGTTGGATGCCATCAACCCATACAAGGAC |
|
BB14228-3chr8_95952304-R |
ACGTTGGATGTCTCCTCCATTGGACATGAC |
|
BB14228-3chr2_219249005-R |
ACGTTGGATGTTCAGCCTGCGGAAGCTATG |
|
BB14228-3chr4_151827481-R |
ACGTTGGATGACCTTTTCAAGGCTATATCC |
|
BB14228-3chr2_37439069-R |
ACGTTGGATGACTTGGTAACCTGGATGACG |
|
BB14228-3chr2_67630823-R |
ACGTTGGATGGAGAAGAAAGTGATCGTGGG |
|
BB14228-3chr3_52474994-R |
ACGTTGGATGTGGAGCACTTTCCTCAAGGC |
|
BB14228-3chr3_50325883-R |
ACGTTGGATGTCTCAAAGCGTGGAACCTTG |
|
BB14228-3chr12_88514827-R |
ACGTTGGATGAACCTCTTCAGAGCCTCAAC |
|
BB14228-3chr4_187541196-R |
ACGTTGGATGTCTCTGCAATCTCTGCACTG |
|
BB14228-3chr10_124221227-R |
ACGTTGGATGAGCGGCCGGCCCGGGACAG |
|
BB14228-3chr1_112298707-EX |
CCGCCAACAGCACATCC |
|
BB14228-3chr14_68264412-EX |
GGCTCCTTCTTGTCCGA |
|
BB14228-3chr1_46509382-EX |
TCTGTGCATGAACAGGG |
|
BB14228-3chr6_35911730-EX |
TGGTCAGTGGAGAGGAGA |
|
BB14228-3chr2_145156750-EX |
ACTATGCTATGAACATGGA |
|
BB14228-3chr8_95952304-EX |
CCATTGGACATGACTCAAAC |
|
BB14228-3chr2_219249005-EX |
AAGCTATGGGCCTTCACGGGG |
|
BB14228-3chr4_151827481-EX |
CTATATCCTATTACCAAGAAGC |
|
BB14228-3chr2_37439069-EX |
GATGAAGTTTCTTTAGGAAGTA |
|
BB14228-3chr2_67630823-EX |
GAAAGTGATCGTGGGGTTTTAT |
|
BB14228-3chr3_52474994-EX |
TCCTCAAGGCCAGGCTGGTCTGCT |
|
BB14228-3chr3_50325883-EX |
TTGCAGGCCTTCAGGGCAGTGGCA |
|
BB14228-3chr12_88514827-EX |
AGCCTCAACTAATTCTTTATCCTTTT |
|
BB14228-3chr4_187541196-EX |
CTCTGCACTGTAGAAAGGTTTTTCAA |
|
BB14228-3chr10_124221227-EX |
GGCCGGCCCGGGACAGCTGCGCCGAG |
Shrimp alkaline phosphatase (SAP)
purification
The total volume for the SAP purification reaction
was 2 µl. This included 1.53 µl ddH2O, 0.17 µl SAP
Buffer and 0.3 µl SAP enzyme (Sequenom, San Diego, CA, USA).
Reaction conditions were 37°C for 40 min and 85°C for 5 min.
Extension reaction
The extension reaction was performed using a 9700
PCR instrument (Sequenom, Inc., San Diego, CA, USA) according to
the manufacturer's protocol. The Complete iPLEX® Gold
Genotyping Reagent Set was purchased from Sequenom. The total
volume of the extension reaction was 2 µl and included 0.619 µl
ddH2O, 0.2 µl iPLEX GOLD Buffer, 0.2 µl iPLEXTermination
mix, 0.94 µl iPLEX Extension Primer mix and 0.041 µl iPLEX Enzyme.
Extension reaction conditions were 40 cycles of 94°C for 30 sec and
94°C for 5 sec, and 5 cycles of 52°C for 5 sec and 80°C for 5 sec,
followed by 1 cycle of 72°C for 3 min. The PCR products were
purified using resin, were spotted onto a chip and were analyzed on
the MassARRAY Platform SEQUENOM Analyzer 4 (Sequenom, Inc.).
Results
Whole exome sequencing generated 60.4 Gb of data
from the 3 HNPCC patients. These data were screened as follows: i)
Remove bases with low scores in accordance with the quality control
specifications that require the sample coverage to be <5 and
fraction variation in a single nucleotide to be <40; ii) filter
and eliminate bases that do not fall in exonic regions; iii) filter
and eliminate proven mutations in controls and common mutations
carried by controls that are archived in public genetic mutation
databases, namely HAPMAP (ftp://ftp.ncbi.nlm.nih.gov/hapmap/), 1,000 Genomes
(http://www.internationalgenome.org/1000-genomes-project-publications)
and dbSNP (https://www.ncbi.nlm.nih.gov/snp/); iv) reserve single
nucleotide loci changes in non-synonymous mutations and filter out
single nucleotide changes in synonymous mutations; and v) select
the non-synonymous mutations that are common to the 3 cases. From
this analysis the following mutations were identified in 15 genes
(Table V): DDX20
(rs112298707), ZFYVE26 (rs68264412),
PIK3R3 (rs46509382), SLC26A8
(rs35911730), ZEB2 (rs145156750),
TP53INP1 (rs95952304), SLC11A1
(rs219249005), LRBA (rs151827481),
CEBPZ (rs37439069), ETAA1 (rs67630823),
SEMA3G (rs52474994), IFRD2
(rs50325883), FAT1 (rs18754119), CEP290
(rs88514827) and HTRA1 (rs124221227). SNP
genotyping of these 15 genes was then performed in 96 subjects
using the DNA MassARRAY Genetic Analysis system (Sequenom)
(Table VI). Among the 96 samples,
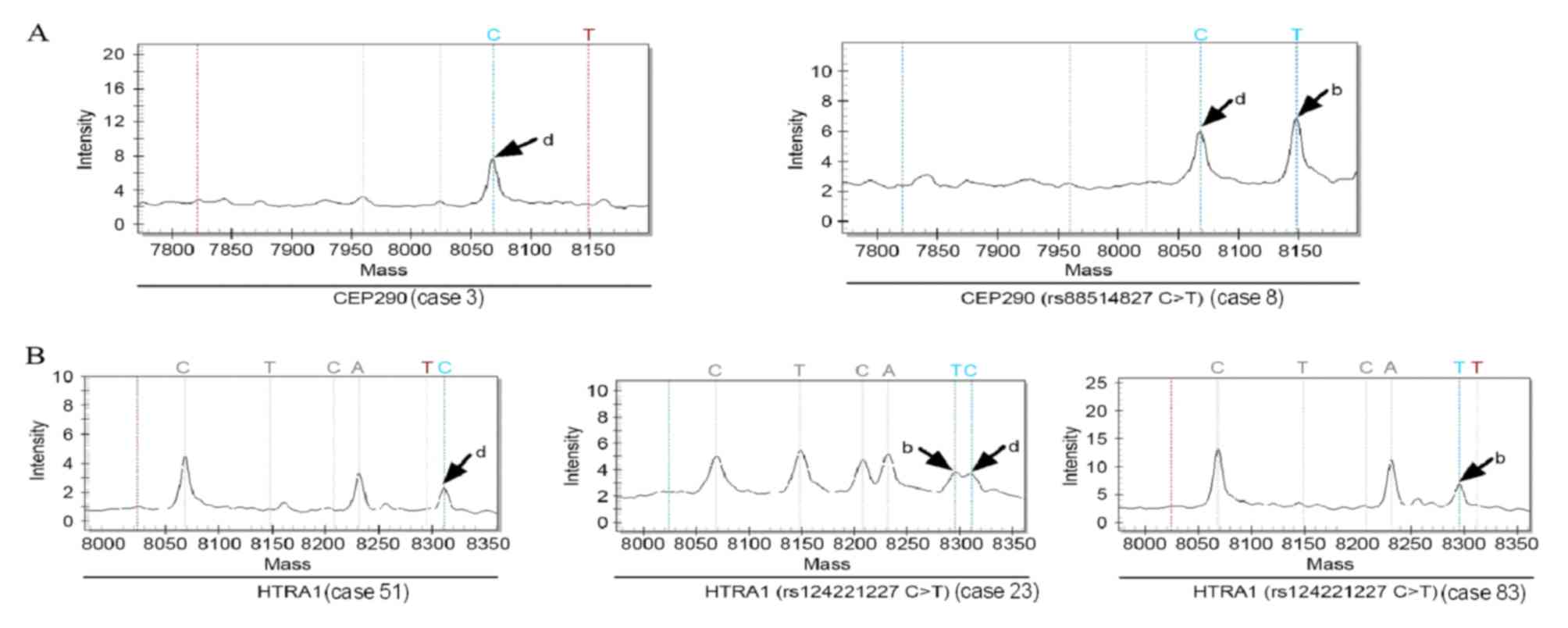
SNP genotyping was successful at all 15 loci in 92, but genotyping
of HTRA1 (rs124221227C>T) failed in 4 of the
sporadic colorectal cancer samples (Fig.
2). The genotype of CEP290 (rs88514827C>T) in
all 12 control subjects was wild-type, while 1 of the 81 patients
with sporadic colorectal cancer had a mutation in CEP290
(rs88514827C>T) (Fig. 2A).
A total of 5/12 control subjects and 30/81 sporadic colorectal
cancer patients had mutations in HTRA1
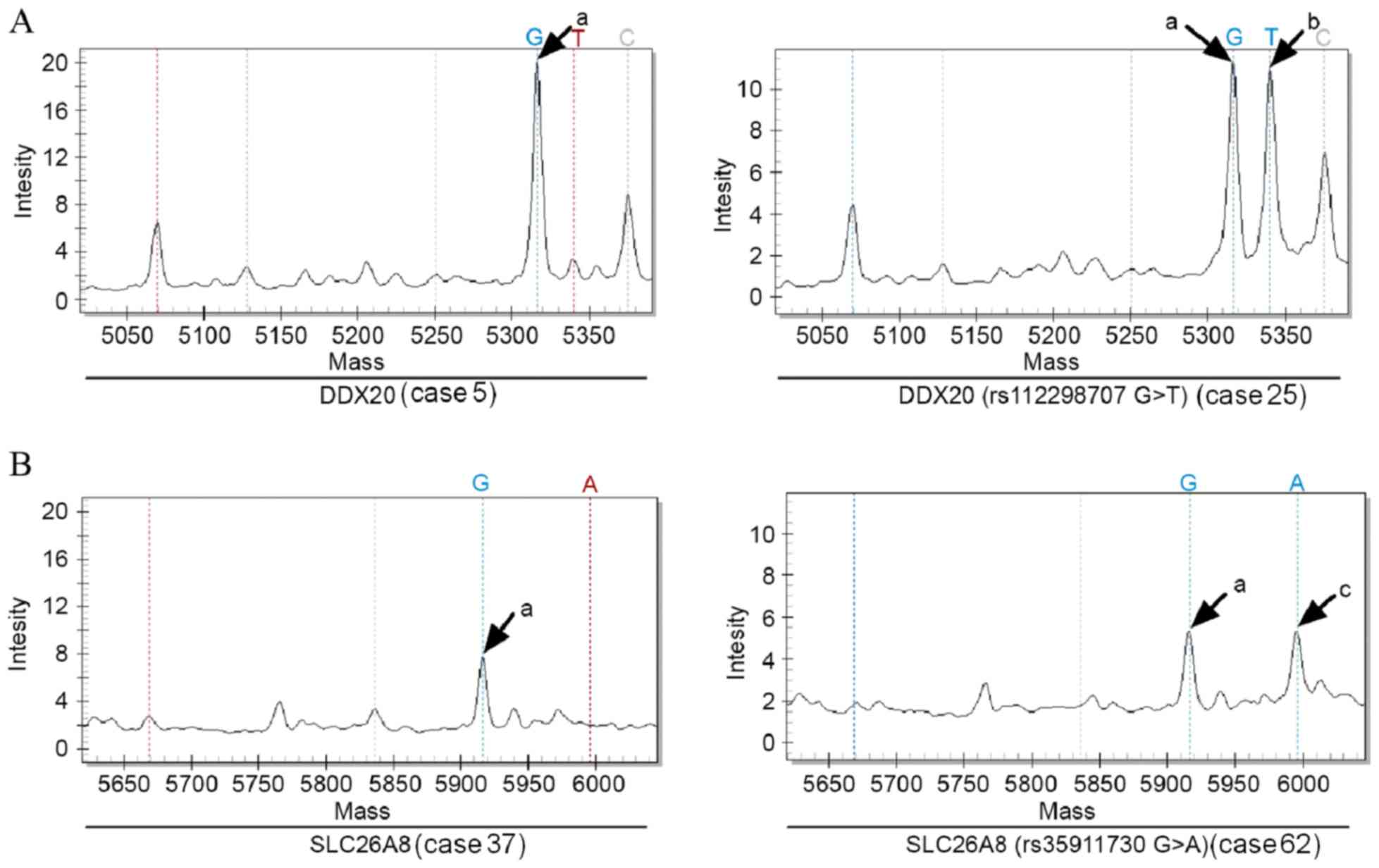
(rs124221227C>T). The genotypes of the other 13 genes in
the 12 control subjects and 81 sporadic colorectal cancer patients
were all wild-type, namely DDX20 (rs112298707G>T),
ZFYVE26 (rs68264412C>T), PIK3R3
(rs46509382T>C), SLC26A8
(rs35911730G>A), ZEB2 (rs145156750C>A),
TP53INP1 (rs95952304T>C), SLC11A1
(rs219249005C>G), LRBA (rs151827481C>T),
CEBPZ (rs37439069A>G), ETAA1
(rs67630823A>G), SEMA3G (rs52474994G>A),
IFRD2 (rs50325883 T>G) and FAT1
(rs187541196A>C) (two of these were selected as examples
and are presented in Fig. 3). In all
3 HNPCC patients, all 15 genes carried the same mutations.
| Table V.15 single nucleotide mutations
detected following integration. |
Table V.
15 single nucleotide mutations
detected following integration.
| Gene name | Chromosome
number | Position | Reference base | Sequencing
base | Mutation type | Gene position |
|---|
| DDX20 | chr1 | 112298707 | G | T | Nonsense | Exon |
| ZFYVE26 | chr14 | 68264412 | C | T | Nonsense | Exon |
| PIK3R3 | chr1 | 46509382 | T | C | Nonsense | Exon |
| SLC26A8 | chr6 | 35911730 | G | A | Nonsense | Exon |
| ZEB2 | chr2 | 145156750 | C | A | Nonsense | Exon |
|
TP53INP1 | chr8 | 95952304 | T | C | Nonsense | Exon |
| SLC11A1 | chr2 | 219249005 | C | G | Nonsense | Exon |
| LRBA | chr4 | 151827481 | C | T | Nonsense | Exon |
| CEBPZ | chr2 | 37439069 | A | G | Nonsense | Exon |
| ETAA1 | chr2 | 67630823 | A | G | Nonsense | Exon |
| SEMA3G | chr3 | 52474994 | G | A | Nonsense | Exon |
| IFRD2 | chr3 | 50325883 | T | G | Nonsense | Exon |
| FAT1 | chr4 | 187541196 | A | C | Nonsense | Exon |
| CEP290 | chr12 | 88514827 | C | T | Nonsense | Exon |
| HTRA1 | chr10 | 124221227 | C | T | Nonsense | Exon |
| Table VI.Single nucleotide polymorphism
genotyping results at 15 gene loci. |
Table VI.
Single nucleotide polymorphism
genotyping results at 15 gene loci.
|
| SNP genotype |
|---|
|
|
|
|---|
| Gene locus | Control
subjects | Sporadic colorectal
cancer patients | HNPCC family |
|---|
| DDX20
(rs112298707) | GG | GG | GT |
| ZFYVE26
(rs68264412) | CC | CC | CT |
| PIK3R3
(rs46509382) | TT | TT | CT |
| SLC26A8
(rs35911730) | GG | GG | GA |
| ZEB2
(rs145156750) | CC | CC | CA |
| TP53INP1
(rs95952304) | TT | TT | CT |
| SLC11A1
(rs219249005) | CC | CC | CG |
| LRBA
(rs151827481) | CC | CC | CT |
| CEBPZ
(rs37439069) | AA | AA | GA |
| ETAA1
(rs67630823) | AA | AA | GA |
| SEMA3G
(rs52474994) | GG | GG | GA |
| IFRD2
(rs50325883) | TT | TT | GT |
| FAT1
(rs187541196) | AA | AA | CA |
| CEP290
(rs88514827) | CC | CC (80/81) CT
(1/81) | CT |
| HTRA1
(rs124221227) | CC (7/12) TC
(5/12) | CC (47/81) TC
(29/81) TT (1/81) not detected (4/81) | TC |
Discussion
HNPCC is the most common hereditary colorectal
cancer and exhibits familial aggregation; it is often accompanied
by synchronous and metachronous colorectal cancer. The incidence of
extraintestinal malignant tumors in HNPCC patients was previously
revealed to be significantly higher than that in normal subjects
(2). MMR gene defects are the
molecular genetic basis of HNPCC pathogenesis, and ~90% of MMR gene
mutations occur in the hMSH2 and hMLHl genes
(10). However, in certain patients
who meet the clinical diagnostic criteria for HNPCC, MMR gene
defects cannot be detected (11,12).
In the present study, 3 HNPCC cases underwent whole
exome sequencing. Mutations were newly identified at 15 gene loci.
These 15 genes were investigated using an SNP genotyping assay in
96 subjects, including HNPCC patients, sporadic colorectal cancer
patients and control subjects. The 15 loci carried the same
mutations in all 3 HNPCC patients. However, in the 12 control
subjects and 81 sporadic colorectal cancer patients, genotypes were
wild-type at 13 of the 15 gene loci, indicating that mutations in
these 13 genes may be associated with HNPCC pathogenesis. A number
of these 13 genes have been revealed to be associated with the
development and progression of malignant tumors (13–28),
autoimmune diseases, tuberculosis and other infectious diseases
(28), and sperm differentiation
(29). However, the consequences of
mutations in these 13 genes have not previously been reported in
the pathology of colorectal cancer.
The results of the present study revealed that
certain sporadic colorectal cancer patients and control subjects
carry mutations in the HTRA1 gene. The expression level of
the HTRA1 gene is associated with the prognosis of various
types of malignant cancer, including liver cancer, breast cancer
and mesothelioma (30,31). Additionally, 1 of the 81 sporadic
colorectal cancer patients in the present study carried a mutation
in the CEP290 gene that was also present in colorectal
cancer patients from the HNPCC family. However, there have been no
reports of a correlation between CEP290 mutations and the
pathogenesis of malignant tumors. Future studies will further
verify whether HTRA1 and CEP290 are susceptibility
genes for HNPCC by expanding sample sizes.
In the present study, 13 genes that may be
susceptibility genes for HNPCC were identified by whole exome
sequencing and SNP genotyping experiments. In the future, studies
will focus on large-scale genetic screening and in vivo and
in vitro experiments in order to investigate the mechanisms
of the confirmed mutations in the development and progression of
colorectal cancer. It is anticipated that more pathogenic genes
will be discovered and that our understanding of the molecular
genetic basis of HNPCC will be improved, thereby providing
theoretical guidance for the diagnosis and treatment of HNPCC.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81572965) and the
125 Talent Project/New Xiangya Project of the Third Xiangya
Hospital of Central South University.
References
|
1
|
Kastrinos F and Stoffel EM: History,
genetics, and strategies for cancer prevention in Lynch syndrome.
Clin Gastroenterol Hepatol. 12:715–727; quiz e41-43. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Watson P and Riley B: The tumor spectrum
in the Lynch syndrome. Fam Cancer. 4:245–248. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fishel R, Lescoe MK, Rao MR, Copeland NG,
Jenkins NA, Garber J, Kane M and Kolodner R: The human mutator gene
homolog MSH2 and its association with hereditary nonpolyposis colon
cancer. Cell. 75:1027–1038. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bronner CE, Baker SM, Morrison PT, Warren
G, Smith LG, Lescoe MK, Kane M, Earabino C, Lipford J, Lindblom A,
et al: Mutation in the DNA mismatch repair gene homologue hMLH1 is
associated with hereditary non-polyposis colon cancer. Nature.
368:258–261. 1994. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Whitehouse A, Meredith DM and Markham AF:
DNA mismatch repair genes and their association with colorectal
cancer (Review). Int J Mol Med. 1:469–474. 1998.PubMed/NCBI
|
|
6
|
Hampel H, Frankel WL, Martin E, Arnold M,
Khanduja K, Kuebler P, Nakagawa H, Sotamaa K, Prior TW, Westman J,
et al: Screening for the Lynch syndrome (hereditary nonpolyposis
colorectal cancer). N Engl J Med. 352:1851–1860. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lynch HT and de la Chapelle A: Hereditary
colorectal cancer. N Engl J Med. 348:919–932. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bashyam MD, Kotapalli V, Raman R,
Chaudhary AK, Yadav BK, Gowrishankar S, Uppin SG, Kongara R, Sastry
RA, Vamsy M, et al: Evidence for presence of mismatch repair gene
expression positive Lynch syndrome cases in India. Mol Carcinog.
54:1807–1814. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vasen HF, Mecklin JP, Khan PM and Lynch
HT: The International Collaborative Group on Hereditary
Non-Polyposis Colorectal Cancer (ICG-HNPCC). Dis Colon Rectum.
34:424–425. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Peltomäki P and Vasen H: Mutations
associated with HNPCC predisposition-Update of ICG-HNPCC/INSiGHT
mutation database. Dis Markers. 20:269–276. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lindor NM: Familial colorectal cancer type
X: The other half of hereditary nonpolyposis colon cancer syndrome.
Surg Oncol Clin N Am. 18:637–645. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nieminen TT, O'Donohue MF, Wu Y, Lohi H,
Scherer SW, Paterson AD, Ellonen P, Abdel-Rahman WM, Valo S,
Mecklin JP, et al: Germline mutation of RPS20, encoding a ribosomal
protein, causes predisposition to hereditary nonpolyposis
colorectal carcinoma without DNA mismatch repair deficiency.
Gastroenterology. 147:595–598.e5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang H, Dinney CP, Ye Y, Zhu Y, Grossman
HB and Wu X: Evaluation of genetic variants in microRNA-related
genes and risk of bladder cancer. Cancer Res. 68:2530–2537. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shin EM, Hay HS, Lee MH, Goh JN, Tan TZ,
Sen YP, Lim SW, Yousef EM, Ong HT, Thike AA, et al: DEAD-box
helicase DP103 defines metastatic potential of human breast
cancers. J Clin Invest. 124:3807–3824. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sagona AP, Nezis IP, Bache KG, Haglund K,
Bakken AC, Skotheim RI and Stenmark H: A tumor-associated mutation
of FYVE-CENT prevents its interaction with Beclin 1 and interferes
with cytokinesis. PLoS One. 6:e170862011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wen JF BB, Zhang XB and Jiang-Ping XU:
Expression and significance of ZFYVE26 in hepatocellular carcinoma.
J Prac Med. 28:1939–1942. 2012.
|
|
17
|
Cao G, Dong W, Meng X, Liu H, Liao H and
Liu S: MiR-511 inhibits growth and metastasis of human
hepatocellular carcinoma cells by targeting PIK3R3. Tumour Biol.
36:4453–4459. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang G, Yang X, Li C, Cao X, Luo X and Hu
J: PIK3R3 induces epithelial-to-mesenchymal transition and promotes
metastasis in colorectal cancer. Mol Cancer Ther. 13:1837–1847.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wong TS, Gao W and Chan JY: Transcription
regulation of E-cadherin by zinc finger E-box binding homeobox
proteins in solid tumors. Biomed Res Int. 2014:9215642014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shahbazi J, Lock R and Liu T: Tumor
protein 53-induced nuclear protein 1 enhances p53 function and
represses tumorigenesis. Front Genet. 4:802013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou X, Ma L, Li J, Gu J, Shi Q and Yu R:
Effects of SEMA3G on migration and invasion of glioma cells. Oncol
Rep. 28:269–275. 2012.PubMed/NCBI
|
|
22
|
Valletta D, Czech B, Spruss T, Ikenberg K,
Wild P, Hartmann A, Weiss TS, Oefner PJ, Müller M, Bosserhoff AK
and Hellerbrand C: Regulation and function of the atypical cadherin
FAT1 in hepatocellular carcinoma. Carcinogenesis. 35:1407–1415.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Morris LG, Kaufman AM, Gong Y, Ramaswami
D, Walsh LA, Turcan Ş, Eng S, Kannan K, Zou Y, Peng L, et al:
Recurrent somatic mutation of FAT1 in multiple human cancers leads
to aberrant Wnt activation. Nat Genet. 45:253–261. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang JW, Gamsby JJ, Highfill SL, Mora LB,
Bloom GC, Yeatman TJ, Pan TC, Ramne AL, Chodosh LA, Cress WD, et
al: Deregulated expression of LRBA facilitates cancer cell growth.
Oncogene. 23:4089–4097. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Herold T, Metzeler KH, Vosberg S, Hartmann
L, Röllig C, Stölzel F, Schneider S, Hubmann M, Zellmeier E,
Ksienzyk B, et al: Isolated trisomy 13 defines a homogeneous AML
subgroup with high frequency of mutations in spliceosome genes and
poor prognosis. Blood. 124:1304–1311. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu DI, Liu L, Ren C, Kong D, Zhang P, Jin
X, Wang T and Zhang G: Epithelial-mesenchymal interconversions and
the regulatory function of the ZEB family during the development
and progression of ovarian cancer. Oncol Lett. 11:1463–1468. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Childs EJ, Mocci E, Campa D, Bracci PM,
Gallinger S, Goggins M, Li D, Neale RE, Olson SH, Scelo G, et al:
Common variation at 2p13.3, 3q29, 7p13 and 17q25.1 associated with
susceptibility to pancreatic cancer. Nat Genet. 47:911–916. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Archer NS, Nassif NT and O'Brien BA:
Genetic variants of SLC11A1 are associated with both autoimmune and
infectious diseases: Systematic review and meta-analysis. Genes
Immun. 16:275–283. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lohi H, Kujala M, Makela S, Lehtonen E,
Kestila M, Saarialho-Kere U, Markovich D and Kere J: Functional
characterization of three novel tissue-specific anion exchangers
SLC26A7, -A8 and -A9. J Biol Chem. 277:14246–14254. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu F, Jin L, Luo TP, Luo GH, Tan Y and
Qin XH: Serine protease HtrA1 expression in human hepatocellular
carcinoma. Hepatobiliary Pancreat Dis Int. 9:508–512.
2010.PubMed/NCBI
|
|
31
|
Lehner A, Magdolen V, Schuster T, Kotzsch
M, Kiechle M, Meindl A, Sweep FC, Span PN and Gross E:
Downregulation of serine protease HTRA1 is associated with poor
survival in breast cancer. PLoS One. 8:e603592013. View Article : Google Scholar : PubMed/NCBI
|