Introduction
Cervical cancer, a malignant tumor, is the fourth
most common cancer in women. The main cause of cervical cancer is
continuous infection with human papilloma virus (HPV) (1–4). Cervical
cancer cells rapidly evade the immune system and promote tumor
progression by inhibiting antitumor immunity (5–8). Many
reports have demonstrated the expansion of various
immunosuppressive cells such as regulatory T cells,
tumor-associated macrophages, myeloid-derived suppressor cells
(MDSCs), and N2 neutrophils in cervical tumors (7–10).
Therefore, understanding changes in immunosuppressive cells is
important for tumor diagnosis and treatment.
In cancer patients and animal tumor models, there is
a significant accumulation of MDSCs, a heterogeneous and diverse
population, in the blood, lymph nodes, bone marrow, and cancer
tissues, and they can inhibit innate and adaptive immune responses
(11,12); this represents an important mechanism
of immune evasion for tumor cells. MDSCs have different phenotypes,
based on factors secreted during bone marrow differentiation and by
tumor cells, which affect cell differentiation (13). In mice, CD11b and Gr-1 are used as
specific markers of MDSCs. Further studies divided mouse MDSCs into
two major subsets, namely monocytic
(Gr-1+Ly6C+) and granulocytic
(Gr-1+Ly6G+) (14). In contrast, human MDSCs are not
associated with widely recognized specific markers (15,16).
However, similarly, they can also be classified as granulocytic
(CD15, CD66b, and CD33-expressing) and monocytic (CD14-expressing).
In kidney cancer patients,
CD14−CD15+CD11b+CD66+
granulocytic MDSCs are immunosuppressive (17). Various MDSC phenotypes in non-small
cell lung cancer include
Lin−/lowHLA-DR−CD11b+CD33+,
Lin−/lowHLA-DR−CD33+,
CD14+S100A9+,
CD14−CD15+CD11b+CD33+,
and CD14+HLA-DR− have been confirmed
(18–22). In addition, the immunosuppressive
effects of
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs have been reported in human malignant gliomas, breast
cancers, colon cancers, and kidney cancers (13,23–25), and
increased MDSC levels are associated with tumor burden and
prognosis in breast and colon cancer patients (19).
In the current study, the level of MDSCs in the
peripheral blood of 105 patients with different clinical stages and
in tumor tissue and corresponding adjacent tissue of 22 clinical
specimens were assessed. Cellular subsets and phenotypic
characteristics and function of these cells were analyzed. The
accumulated evidence can contribute to understanding the clinical
characteristics of peripheral blood and local tumor-infiltrating
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs in cervical cancer.
Materials and methods
Ethics statement
All cervical carcinoma patients and healthy donors
provided written informed consent prior to blood sampling and/or
tumor tissue harvesting. The research protocol was approved by the
Medical Ethics Committee of Chinese PLA General Hospital (Beijing,
China) and the 307th Hospital of Chinese PLA (Beijing, China).
Patients
Control samples from healthy volunteers (n=50) and
cervical cancer patient samples (n=105) were obtained at the
gynecology departments of the Chinese PLA general hospital and the
307th Hospital of Chinese PLA. All patients were newly diagnosed
and treatment-naive. Table I shows
the clinical characteristics of patients included in this
study.
 | Table I.Patient characteristics. |
Table I.
Patient characteristics.
| Variables | Total n number
(%) |
|---|
| Number of patients
(n) | 105 |
| Age [years; mean
(SD)] | 44.3 (7.8) |
| FIGO stage |
|
| Stage
I | 23 (21.9) |
| Stage
II | 28 (26.7) |
| Stage
III | 22 (21.0) |
| Stage
IV | 32 (30.4) |
| HPV type |
|
| 16 | 76 (72.4) |
| 18 | 24 (22.8) |
|
Other | 5 (4.8) |
| Histopathology |
|
|
Squamous | 64 (61.0) |
| Adeno
(squamous) | 41 (39.0) |
| Lymph node
metastasis |
|
| Lymph
nodes (+) | 61 (58.1) |
| Lymph
nodes (−) | 44 (41.9) |
| Vasoinvasion |
|
| No | 31 (29.5) |
|
Yes | 74 (70.5) |
| Parametrial
involvement |
|
| No | 26 (24.8) |
|
Yes | 79 (75.2) |
Flow cytometry
Blood samples for the detection of peripheral
circulating MDSCs were collected using EDTA anticoagulant tubes (BD
Biosciences, Franklin Lakes, NJ, USA). Monoclonal fluorescent
antibodies, CD11b-PECY7 (cat. no. A54822), HLA-DR-ECD (cat. no.
IM3636), and CD33-PECY5 (cat. no. IM26 47 U), were from Beckman
Coulter, Inc., (Brea, CA, USA). Lineage (CD3, CD14, CD16, CD19,
CD20, and CD56-FITC) (cat. no. 340546) antibodies were all from BD
Biosciences. Four-color analysis was used to confirm MDSCs.
Analysis of tumor-infiltrating MDSCs was performed using anti-human
CD45-FITC (cat. no. 304006), CD11b-PECY7 (cat. no. A54822), and
CD33-PECY5 (cat. no. IM2647 U; all from BioLegend, Inc., San Diego,
CA, USA). For phenotypic characterization of MDSCs, the MDSC
population was gated for the analysis of PE expression. Antibodies
involved in phenotype analysis included CD13-PE (cat. no. 301703),
CD39-PE (cat. no. 328208), CD34-PE (cat. no. 343505), CD73-PE (cat.
no. 344004), CD66b-PE (cat. no. 305105), CD115 (CSF-1R)-PE (cat.
no. 347303), PD-1 (CD279)-PE (cat. no. 329906), CD124 (IL-4Ra)-PE
(cat. no. 355003), PD-L1 (CD274)-PE (cat. no. 329706), and PD-L2
(CD273)-PE (cat. no. 329606). Isotype control antibodies (Mouse
IgG1-PE, cat. no. 400114; Mouse IgM-PE, cat. no. 401611; Mouse
IgG2a-PE, cat. no. 400214; Mouse IgG2b-PE, cat. no. 401208; Rat
IgG1-PE, cat. no. 400408) were used as controls. Beforementioned
antibodies were from BioLegend, Inc. For the detection of
peripheral blood and tumor-infiltrating cell phenotypes, a standard
amount of corresponding antibody was added. Subsequently, 500 µl of
OptiLyse C Lysing Solution (cat. no. A11895; Beckman Coulter, Inc.)
was added to each blood sample and incubated for 15 min; 500 µl PBS
was then added before 500 µl of FACS buffer was added; analysis was
performed by flow cytometry. For intracellular cytokine staining,
purified MDSCs were added at a ratio of 1:1 to the control group
(lymphocytes alone) or to the experimental group. Cell Stimulation
cocktail plus protein transport inhibitors (eBioscience; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) was added to each group
for 4 h of stimulation. Cell surface marker staining was performed
using CD3-ECD (cat. no. A07748), CD4-PC5 (cat. no. IM2636 U), and
CD8-PECY7 (cat. no. 6607102) (all from Beckman Coulter, Inc.). The
BD Cytofix/Cytoperm™ Plus Fixation/Permeabilization kit (cat. no.
555028) reagent box was used to process cells before intracellular
cytokine staining using IL-2-PE (cat. no. 506709) and IFNg-FITC
(cat. no. 552887) antibodies and corresponding isotype control
antibodies (Rat IgG2a-PE, cat. no. 559317; Mouse IgG1-FITC, cat.
no. 556649; all from BD Biosciences). After treatment, cells were
analyzed by flow cytometry for the production of T-cell cytokines.
Samples were obtained using a flow cytometer FC500-MPL (Beckmam
Coulter, Inc.), and data analysis was performed using FlowJo
software (Tree Star, Inc., Ashland, OR, USA). Absolute MDSC counts
were calculated using the following formula: [total white blood
cell count (cells/ml) percent MDSCs]/100 or [total
tumor-infiltrating immune cell count (cells/100 mg tumor) percent
MDSCs]/100.
Cell separation
Separation of PBMCs was performed using density
gradient centrifugation. Briefly, blood samples with EDTA
anticoagulant were carefully separated by Ficoll-Hypaque (GE
Healthcare, Chicago, IL, USA) separation media. The PBMC obtained
after centrifugation staining was used to determine cell viability
by trypan blue before flow cytometry.
To separate tumor-infiltrating immune cells, newly
resected tumor tissue (100 mg) and matching surrounding tissue from
22 Stage III or IV cervical cancer patients were cut into pieces
and digested using 500 mg/ml Liberase (collagenase) and 200 mg/ml
DNase (Roche Applied Science, Penzberg, Germany) for 45 min. The
cell suspension was then passed through a 70-µm cell strainer (BD
Biosciences). Centrifugation using a density gradient was then
performed as described, and the corresponding cell layer was
aspirated using a pipette.
In vitro inhibition analysis
experiment
Fresh blood samples (20 ml) from three stage IV
cervical cancer patients were used for PBMC extraction.
CD11b-PECY7, HLA-DR-ECD, CD33-PECY5, Lineage-FITC, and CD3
monoclonal fluorescent antibodies were added to PBMCs before being
sorted by the MoFloTM XDP cell sorting system (Beckman Coulter,
Inc.). Sorted cells had a purity >95%. For MDSC functional
analysis, purified CD3 T-cells were stained with 2 mM CFSE
(Invitrogen; Thermo Fisher Scientific, Inc.) and CFSE-stained
T-cells were cultured with Lin-/lowHLA-DR-CD11b+ CD33+ MDSCs at
ratios of 1:0, 1:0.25, 1:0.5, and 1:1. Soluble anti-CD3 (2 mg/ml)
and anti-CD28 (0.5 mg/ml) antibodies were added and cells were
incubated for 24 h before being measuring proliferation through
flow cytometry.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 5.0 software (GraphPad Software, United States); unpaired
Student's t tests (Mann-Whitney test) and unparametric Spearman
tests were used to assess differences and correlations between
study groups, respectively. P<0.05 was considered to indicate a
statistically significant difference.
Results
Increase in the proportion and numbers
of peripheral
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs in cervical cancer patients
The proportion of
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs in the peripheral blood of cervical cancer patients of
different clinical stages was measured by flow cytometry (Table I shows clinical patients data). The
ratio of MDSCs to total leukocytes in healthy volunteers, clinical
stage I–II, and Stage III–IV patients was calculated using flow
cytometry (as depicted in Fig. 1A).
The proportion of MDSCs in cervical cancer patients was
significantly higher compared to that in controls (P<0.0001;
Fig. 1B). MDSC levels were also
significantly increased in the peripheral blood of cervical cancer
patients compared to that in controls (P<0.0001; Fig. 1C).
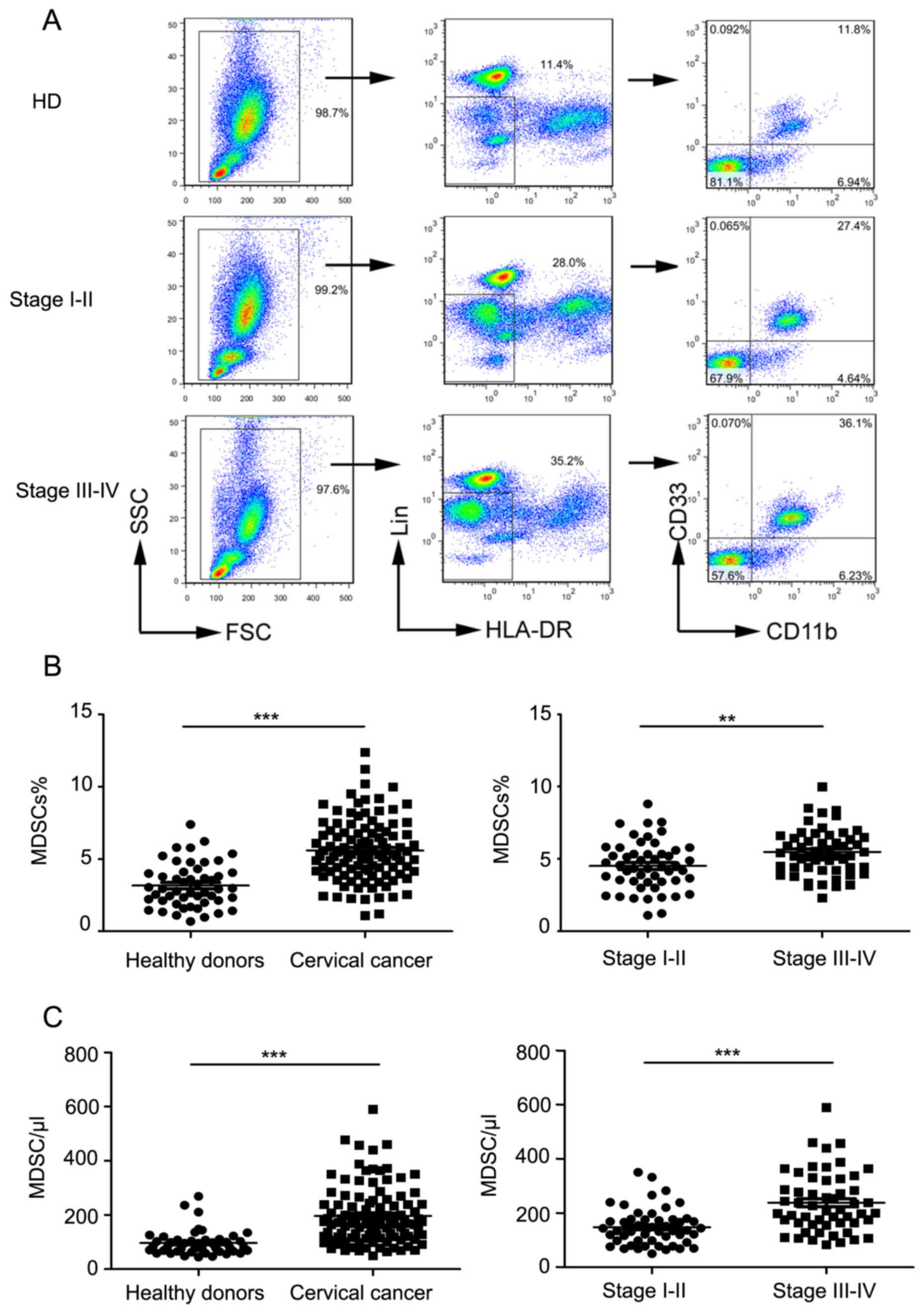 | Figure 1.Levels of peripheral blood
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs in normal subjects and cervical cancer patients. (A)
Representative flow cytometry and analysis strategy. (B) Comparison
of peripheral circulating MDSC proportions in normal controls
(n=50) and cervical cancer patients (n=105; ***P<0.0001, as
indicated), and comparison of patients in late stage with early
stage (**P=0.0014, as indicated). (C) Comparison of absolute MDSC
counts in normal controls (n=50) and cervical cancer patients
(n=105; ***P<0.0001, as indicated), and comparison of patients
in late stage with early stage (***P=0.0001, as indicated). MDSCs,
myeloid-derived suppressor cells; HLA-DR, human leukocyte
antigen-antigen D related; CD, cluster of differentiation; FSC,
forward scatter; SSC, side scatter; HD, healthy donors. |
Further, the proportion of MDSCs in clinical stage
III–IV patients was significantly higher than that in clinical
stage I–II patients (P=0.0014; Fig.
1B). Next, we found that the absolute number of MDSCs in stage
III–IV patients was significantly higher than that in stage I–II
patients (P<0.0001; Fig. 1C).
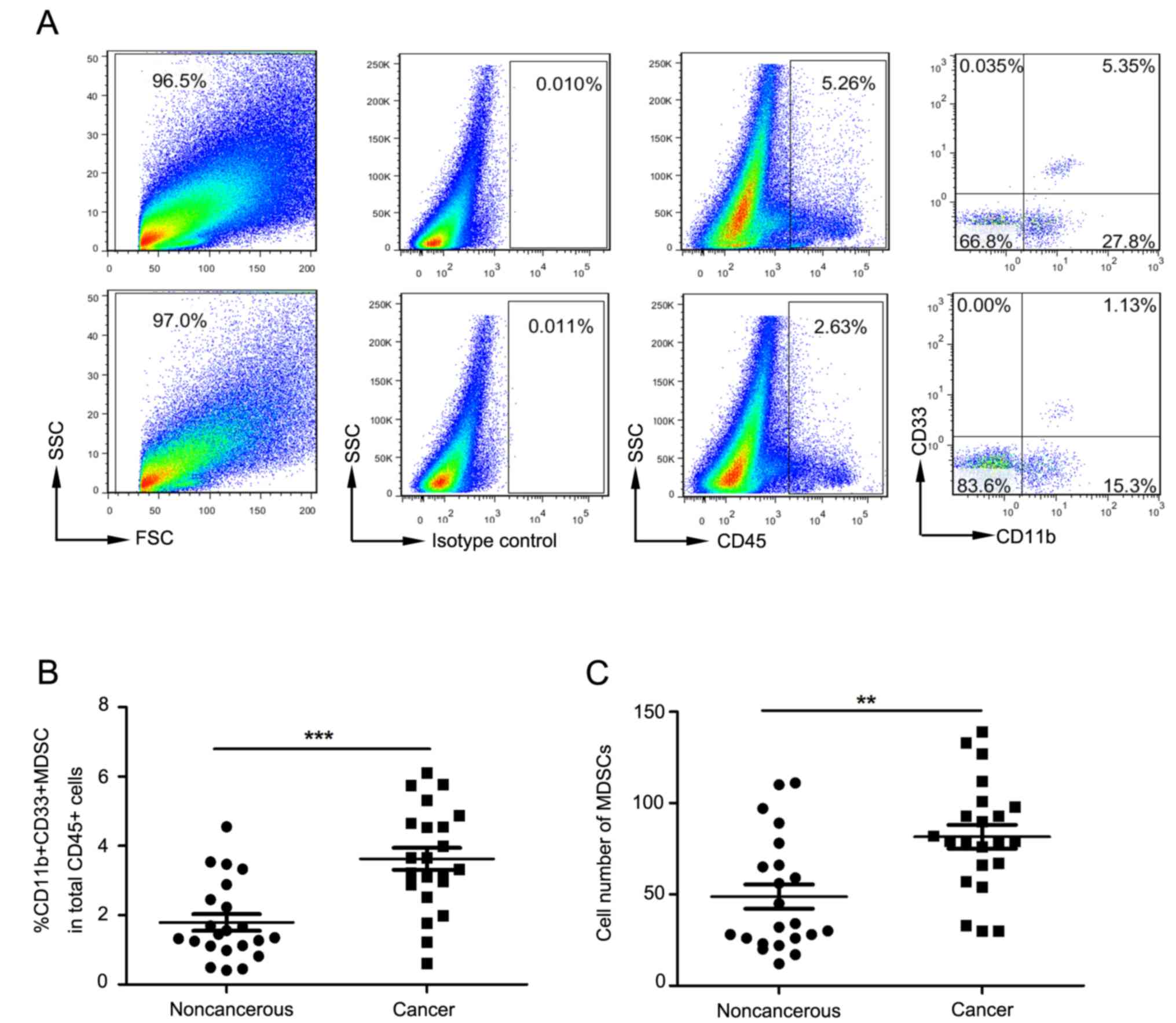
Elevation in the proportion and
numbers of tumor-infiltrating
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs in late stage cervical cancer patients
Fig. 2A depicts flow
cytometry used to examine tumor-infiltrating MDSCs in cervical
cancer patients. For all tumor-infiltrating CD45+
leukocytes, the proportion of MDSCs in cancer tissues was
significantly increased compared to that in surrounding
non-cancerous tissue (P<0.0001; Fig.
2B). The absolute count of MDSCs in cancer tissues was also
significantly increased compared to that in surrounding
non-cancerous tissue (P=0.001; Fig.
2C).
Functional characteristics of
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs
We used flow cytometry to analyze the phenotypic
characteristics of
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs, including myeloid and lymphoid markers (Fig. 3A). In the peripheral MDSCs of cervical
cancer patients, CD13 was highly expressed, CD124 (IL-4Ra), CD115,
and CD117 were lowly expressed, and CD66b, CD14, CD15, PDL1, PD1,
CD34 were not expressed. These cells expressed high levels of CD39
but did not express CD73. Intracellular staining also failed to
detect CD73 expression (results not shown); this was in accordance
with a previous study on colorectal cancer (26). Similar to that observed in mouse MDSCs
(14), PDL1 expression in
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs was low. No significant differences were observed with
respect to these markers between normal and cervical cancer
samples.
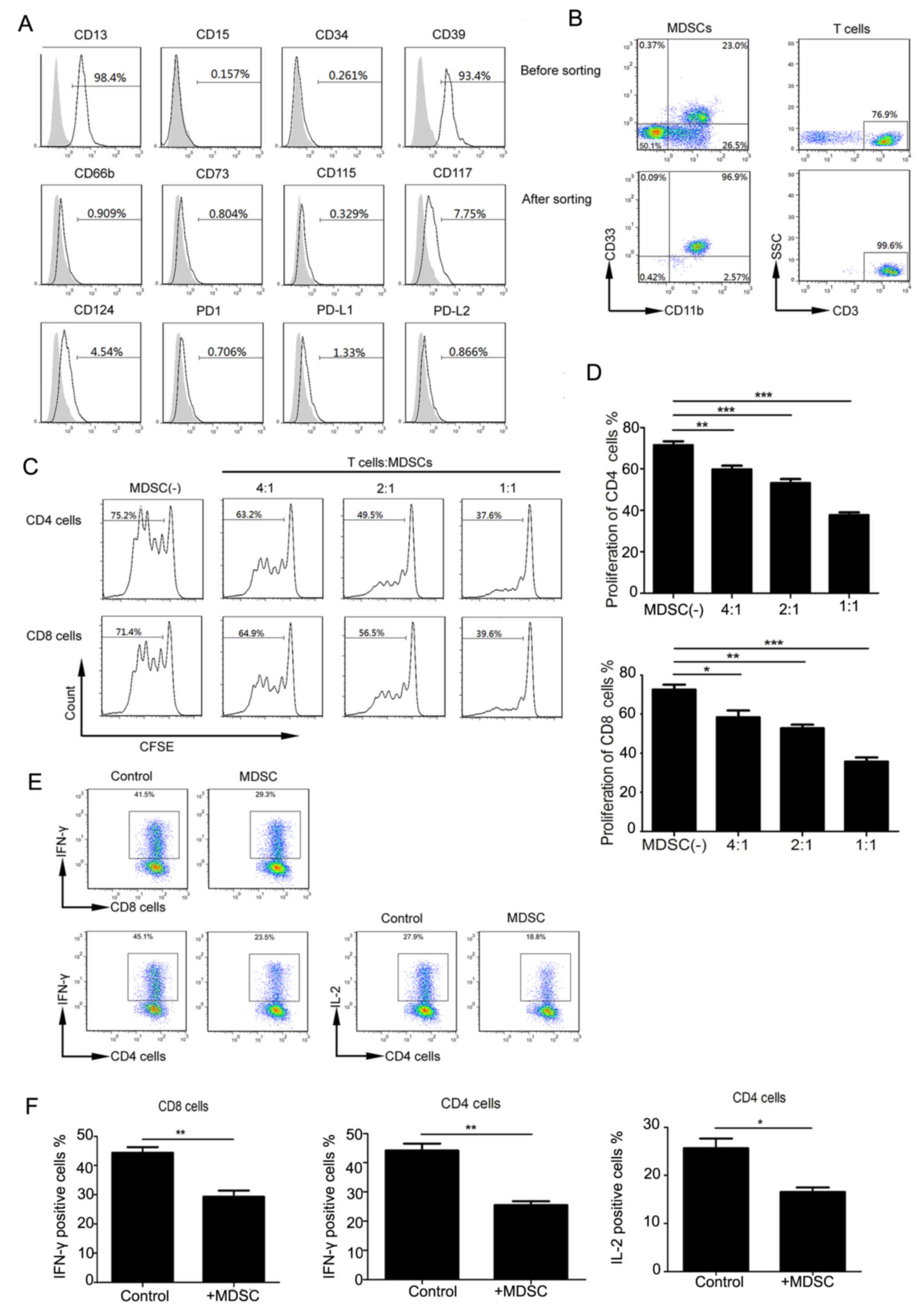 | Figure 3.Functional characteristics of
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs. (A) Expression of indicated molecules (gray histograms
represent isotype controls). (B) Flow cytometry was used to purify
MDSCs and T cells. Representative flow cytometry prior to and
following cell sorting. (C) Representative flow cytometry of the
inhibitory effect of MDSCs on CD4 and CD8 T cells. (D) Quantitation
of MDSC inhibition of CD8 and CD4 cells. *P<0.05, **P<0.01
and ***P<0.001, as indicated. (E) Representative flow cytometry
of IL-12 and IFN-γ secretion by CD4 and CD8 cells. (F) The positive
percentage of CD8 that secretes IFN-γ, and CD4 that secretes IFN-γ
and IL-2, respectively (**P=0.006, **P=0.0024 and *P=0.0372,
respectively). MDSCs, myeloid-derived suppressor cells; SSC, side
scatter; CFSE, carboxyfluorescein succinimidyl ester; CD, cluster
of differentiation; IL, interleukin; PD1, programmed cell death
protein 1; PD-L, programmed cell death ligand; IFN, interferon. |
Previous studies showed that
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs can inhibit T-cell proliferation in other tumor types. We
extracted highly pure (> 95%) MDSCs and CD3+ T-cells (Fig. 3B) from peripheral blood. CFSE-labeled
CD3+ T cells were co-cultured with MDSCs at different
ratios and stimulated with CD3 and CD28 antibodies. Upon analyzing
CFSE fluorescence in CD4 and CD8 T cell subsets, we found that with
an increasing proportion of MDSCs, CD4 and CD8 T cells were
significantly inhibited (Fig. 3C and
D).
MDSCs can function through multiple mechanisms,
including inhibiting cytokine production in T cells. We tried to
verify the effects of MDSCs from cervical cancer patients on CD4
and CD8 cells by analyzing IL-2 and IFN-γ in CD4 T-cells and IFN-γ
production in CD8 T-cells. The cytokine production in CD4 and CD8
cells was decreased in the experimental MDSC group compared to that
in the control group (no MDSCs) (Fig.
3E). IFN-γ production in CD8, and IFN-γ and IL-2 production in
CD4 T cells, respectively, in the control and experimental groups
(P=0.006, P=0.0024 and P=0.0372 respectively) (Fig. 3F). These data suggest that cervical
cancer-associated MDSCs can inhibit cytokine production in T cells,
resulting in decreased proliferation cytotoxicity.
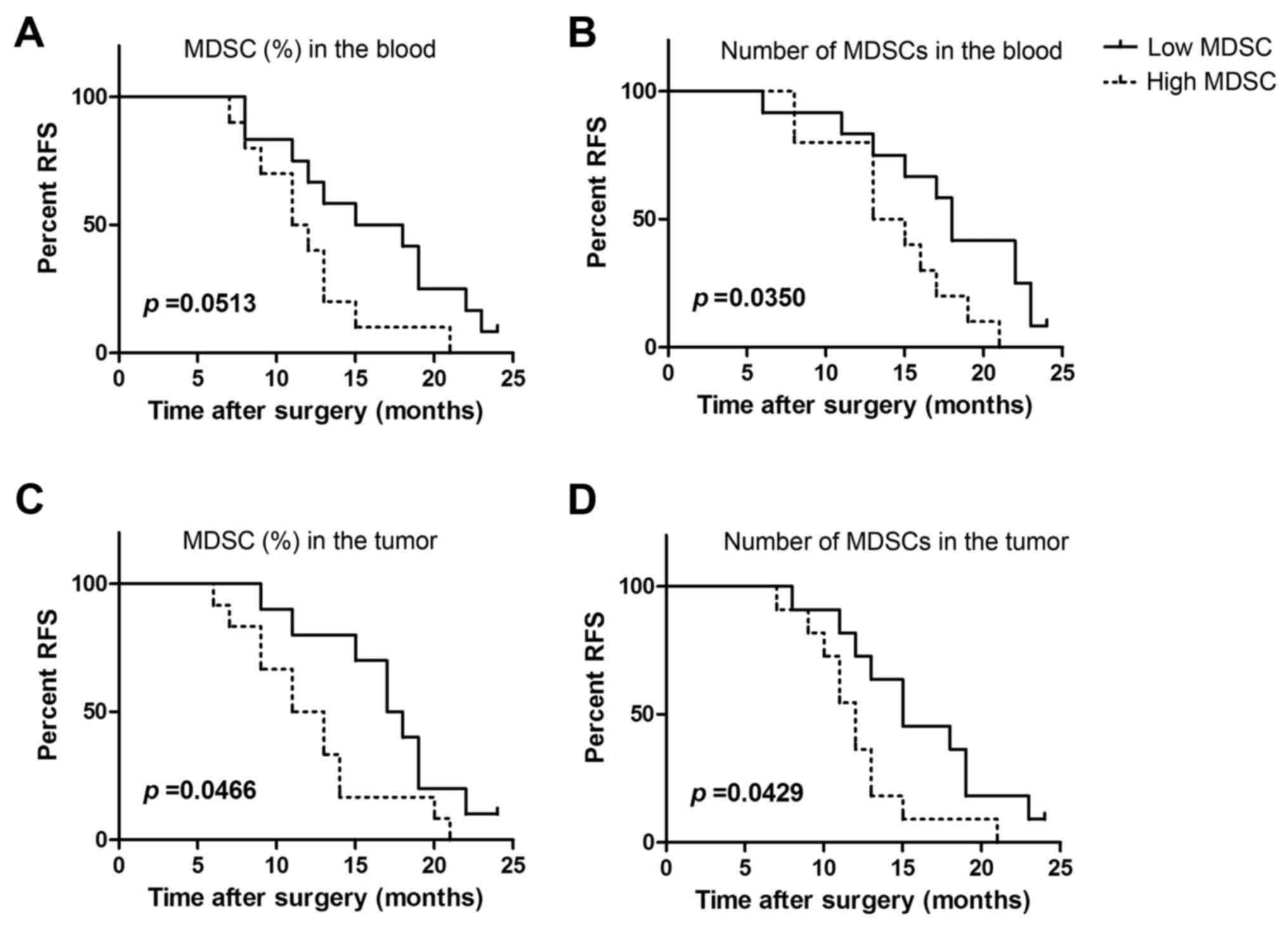
Peripheral and tumor-infiltrating
MDSCs is associated with metastasis in late stage cervical
cancer
We further analyzed the proportions and absolute
numbers of peripheral circulating MDSCs in late stage cervical
cancer patients with tumor relapse and metastasis. Peripheral blood
MDSCs from 22 patients were divided based on MDSC proportions into
the high group (>mean value) and the low group (<mean value).
Relapse was found to occur more readily in the high group (Fig. 4A). Although our results did not reach
statistical significance (P=0.0531), some correlation was observed.
Similarly, levels of peripheral MDSCs in clinical stage IV cervical
cancer patients were found to significantly correlate with RFS
(P=0.035; Fig. 4B).
We next analyzed the relationship between MDSC
proportion and absolute counts and tumor recurrence and metastasis
in 22 tumors from clinical stage IV cervical cancer patients. We
observed that a higher proportion of MDSCs was significantly
associated with recurrence (P=0.0466; Fig. 4C). MDSC levels in tumors were also
found to be significantly associated with tumor recurrence and
metastasis (P=0.0429; Fig. 4D). Thus,
the proportion of peripheral and tumor-infiltrating MDSCs are
related to tumor progression in cervical cancer patients.
Discussion
MDSCs play an important role in tumor immune evasion
and tolerance. We examined changes in
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs in cervical cancer patients based on clinical stage and
obtained results that were consistent with other tumor types.
The samples were all fresh, and conventional sample
preparation protocols were used. In addition to examining MDSC
phenotypes, we also assessed. In mice, MDSC populations have been
verified (14), whereas in humans,
three MDSC populations are recognized. A recent review introduced
the phenotypes and characteristics of mouse and human MDSCs. Mouse
MDSCs are classified as mixed MDSCs
(Gr-1+CD11b+), which was further classified,
based on Ly6C and Ly6 G expression, as PMN-MDSCs
(CD11b+Ly6CloLy6G+) and M-MDSCs
(CD11b+Ly6ChiLy6G−). There are three
recognized human MDSC subsets, namely PMN-MDSCs
(CD14−CD11b+CD15+/CD66b+),
M-MDSCs
(CD11b+CD14+HLA-DRlow/−CD15−),
and E-MDSC
(Lin−/lowHLA-DR−CD11b+CD33+).
This review also summarized the biological functions of these
MDSCs, including inhibiting T lymphocyte proliferation, IL-2 and
IFN-γ production, and function (27).
Our results were consistent with those previously reported;
specifically, we found that MDSCs inhibit T-cell proliferation,
IL-2 and IFN-γ production in CD4 T cells, and IFN-γ production in
CD8 T cells.
In these other studies, changes in MDSCs were shown
to correlate linearly with tumor burden (19,28,29). With
increasing clinical stage, circulating MDSCs increase. Late stage
cancer patients with higher levels of MDSCs are more prone to
recurrence, which in turn affects prognosis (13). Similarly, we showed that peripheral
circulating and tumor-infiltrating MDSC levels are associated with
RFS. Possible reasons for these are as follows: Circulating tumor
cells or tumor cells in the tumor microenvironment could secrete
cytokines resulting in the expansion of MDSCs and produce
pro-inflammatory and angiogenic cytokines to recruit MDSCs,
promoting tumor proliferation and invasion (30).
Previous studies have shown that the proportion of
MDSCs is higher in the microenvironment of different tumor types
(21,31,32). We
also showed that infiltrating MDSC numbers were increased in
cervical cancer patients.
We found that
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs have a specific phenotypic profile. These cells highly
express the myeloid marker CD13, exhibit low expression of CD115,
CD124, and CD117, and do not express the monocytic marker CD14 or
the granulocytic markers CD15 and CD66b. These phenotypes are
generally similar to those previously reported (13). Interestingly, this population of cells
highly expresses CD39 but not CD73, which both synergistically
promote immunosuppression (33).
These two molecules are expressed on human regulatory T cells and
mediate an immunosuppressive effect (34), inhibiting the function of Th1, Th2,
CTL, and NK cells (33,35). In a mouse study, the expression of
CD39 and CD73 increased the immunosuppressive activity of MDSCs
(36). We also found that this cell
population expresses low levels of PD-L1 and does not express PD-L2
or its receptor PD-1 and B7 family members, but can still regulate
the immune response and induce immune tolerance (37).
Consistent with the results of previous studies,
circulating MDSCs were shown to inhibit T-cell proliferation. Some
reports have confirmed that BM-MDSCs inhibit T-cell proliferation
by decreasing their expression of CD3ε and CD3ξ (13). Our in vitro experiments also
confirmed that
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs can inhibit IL-12 and IFN-γ production in T-cells and reduce
T cell performance. BM-MDSCs also express arginase I, which
depletes extracellular L-arginine, resulting in downregulation of
CD3ε chain and diminished T-cell proliferation (17). Lastly, we confirmed that
Lin−/lowHLA-DR−CD11b+CD33+
MDSCs are associated with tumor burden in cervical cancer. Abnormal
accumulation of peripheral blood or local MDSCs is an important
immunological mechanism of T cell anergy. Our studies could provide
the foundation for immunotherapy to treat cervical cancer, and
particularly immunotherapy targeting MDSCs.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LW performed flow cytometry and was a major
contributor in writing the manuscript. HL and HG analyzed and
interpreted the patient data for the cervical carcinoma patients,
and were major contributors in writing the manuscript. QW and SY
performed the separation of PBMC. YQ and LW performed the
separation of tumor-infiltrating immune cells. GW and QW performed
the in vitro inhibition analysis experiments. LZ and CL
performed the collection of patient samples. RZ developed the
methodology, performed data analysis and was a major contributor in
revising the manuscript. TL and SJ made substantial contributions
to the conception and design, analysis and interpretation of data,
and fund support.
Ethics approval and consent to
participate
The research protocol was approved by the Medical
Ethics Committee of Chinese PLA General Hospital (Beijing, China)
and the 307th Hospital of Chinese PLA (Beijing, China). All
patients provided written informed consent for participation in the
present study.
Consent for publication
Written informed consent was obtained from all
participants.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ho GY, Bierman R, Beardsley L, Chang CJ
and Burk RD: Natural history of cervicovaginal papillomavirus
infection in young women. N Engl J Med. 338:423–428. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Trottier H and Franco EL: The epidemiology
of genital human papillomavirus infection. Vaccine. 24 Suppl
1:S1–S15. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Woodman CB, Collins SI and Young LS: The
natural history of cervical HPV infection: Unresolved issues. Nat
Rev Cancer. 7:11–22. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Grabowska AK and Riemer AB: The invisible
enemy-how human papillomaviruses avoid recognition and clearance by
the host immune system. Open Virol J. 6:249–256. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kanodia S, Fahey LM and Kast WM:
Mechanisms used by human papillomaviruses to escape the host immune
response. Curr Cancer Drug Targets. 7:79–89. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kobayashi A, Weinberg V, Darragh T and
Smith-McCune K: Evolving immunosuppressive microenvironment during
human cervical carcinogenesis. Mucosal Immunol. 1:412–420. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Piersma SJ: Immunosuppressive tumor
microenvironment in cervical cancer patients. Cancer Microenviron.
4:361–375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jordanova ES, Gorter A, Ayachi O, Prins F,
Durrant LG, Kenter GG, van der Burg SH and Fleuren GJ: Human
leukocyte antigen class I, MHC class I chain-related molecule A and
CD8+/regulatory T-cell ratio: Which variable determines survival of
cervical cancer patients? Clin Cancer Res. 14:2028–2035. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
De Vos van Steenwijk PJ, Ramwadhdoebe TH,
Goedemans R, Doorduijn EM, van Ham JJ, Gorter A, van Hall T,
Kuijjer ML, van Poelgeest MI, van der Burg SH and Jordanova ES:
Tumor-infiltrating CD14-positive myeloid cells and CD8-positive
T-cells prolong survival in patients with cervical carcinoma. Int J
Cancer. 133:2884–2894. 2013.PubMed/NCBI
|
|
11
|
Gabrilovich DI, Ostrand-Rosenberg S and
Bronte V: Coordinated regulation of myeloid cells by tumours. Nat
Rev Immunol. 12:253–268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gabrilovich DI and Nagaraj S:
Myeloid-derived suppressor cells as regulators of the immune
system. Nat Rev Immunol. 9:162–174. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Solito S, Falisi E, Diaz-Montero CM, Doni
A, Pinton L, Rosato A, Francescato S, Basso G, Zanovello P,
Onicescu G, et al: A human promyelocytic-like population is
responsible for the immune suppression mediated by myeloid-derived
suppressor cells. Blood. 118:2254–2265. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Youn JI, Nagaraj S, Collazo M and
Gabrilovich DI: Subsets of myeloid-derived suppressor cells in
tumor-bearing mice. J Immunol. 181:5791–5802. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Montero AJ, Diaz-Montero CM, Kyriakopoulos
CE, Bronte V and Mandruzzato S: Myeloid-derived suppressor cells in
cancer patients: A clinical perspective. J Immunother. 35:107–115.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Greten TF, Manns MP and Korangy F: Myeloid
derived suppressor cells in human diseases. Int Immunopharmacol.
11:802–807. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rodriguez PC, Ernstoff MS, Hernandez C,
Atkins M, Zabaleta J, Sierra R and Ochoa AC: Arginase I-producing
myeloid-derived suppressor cells in renal cell carcinoma are a
subpopulation of activated granulocytes. Cancer Res. 69:1553–1560.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Almand B, Clark JI, Nikitina E, van Beynen
J, English NR, Knight SC, Carbone DP and Gabrilovich DI: Increased
production of immature myeloid cells in cancer patients: A
mechanism of immunosuppression in cancer. J Immunol. 166:678–689.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Diaz-Montero CM, Salem ML, Nishimura MI,
Garrett-Mayer E, Cole DJ and Montero AJ: Increased circulating
myeloid-derived suppressor cells correlate with clinical cancer
stage, metastatic tumor burden and doxorubicin-cyclophosphamide
chemotherapy. Cancer Immunol Immunother. 58:49–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Srivastava MK, Bosch JJ, Thompson JA,
Ksander BR, Edelman MJ and Ostrand-Rosenberg S: Lung cancer
patients' CD4(+) T cells are activated in vitro by MHC II
cell-based vaccines despite the presence of myeloid-derived
suppressor cells. Cancer Immunol Immunother. 57:1493–1504. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng PH, Lee KY, Chang YL, Chan YF, Kuo
LW, Lin TY, Chung FT, Kuo CS, Yu CT, Lin SM, et al:
CD14(+)S100A9(+) monocytic myeloid-derived suppressor cells and
their clinical relevance in non-small cell lung cancer. Am J Respir
Crit Care Med. 186:1025–1036. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang A, Zhang B, Wang B, Zhang F, Fan KX
and Guo YJ: Increased CD14(+)HLA-DR(−/low) myeloid-derived
suppressor cells correlate with extrathoracic metastasis and poor
response to chemotherapy in non-small cell lung cancer patients.
Cancer Immunol Immunother. 62:1439–1451. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kusmartsev S, Su Z, Heiser A, Dannull J,
Eruslanov E, Kübler H, Yancey D, Dahm P and Vieweg J: Reversal of
myeloid cell-mediated immunosuppression in patients with metastatic
renal cell carcinoma. Clin Cancer Res. 14:8270–8278. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Montero AJ, Diaz-Montero CM, Deutsch YE,
Hurley J, Koniaris LG, Rumboldt T, Yasir S, Jorda M, Garret-Mayer
E, Avisar E, et al: Phase 2 study of neoadjuvant treatment with
NOV-002 in combination with doxorubicin and cyclophosphamide
followed by docetaxel in patients with HER-2 negative clinical
stage II–IIIc breast cancer. Breast Cancer Res Treat. 132:215–223.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Raychaudhuri B, Rayman P, Ireland J, Ko J,
Rini B, Borden EC, Garcia J, Vogelbaum MA and Finke J:
Myeloid-derived suppressor cell accumulation and function in
patients with newly diagnosed glioblastoma. Neuro Oncol.
13:591–599. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang B, Wang Z, Wu L, Zhang M, Li W, Ding
J, Zhu J, Wei H and Zhao K: Circulating and tumor-infiltrating
myeloid-derived suppressor cells in patients with colorectal
carcinoma. PLoS One. 8:e571142013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bronte V, Brandau S, Chen SH, Colombo MP,
Frey AB, Greten TF, Mandruzzato S, Murray PJ, Ochoa A,
Ostrand-Rosenberg S, et al: Recommendations for myeloid-derived
suppressor cell nomenclature and characterization standards. Nat
Commun. 7:121502016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Melani C, Chiodoni C, Forni G and Colombo
MP: Myeloid cell expansion elicited by the progression of
spontaneous mammary carcinomas in c-erbB-2 transgenic BALB/c mice
suppresses immune reactivity. Blood. 102:2138–2145. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bronte V, Apolloni E, Cabrelle A, Ronca R,
Serafini P, Zamboni P, Restifo NP and Zanovello P: Identification
of a CD11b(+)/Gr-1(+)/CD31(+) myeloid progenitor capable of
activating or suppressing CD8(+) T cells. Blood. 96:3838–3846.
2000.PubMed/NCBI
|
|
30
|
Ostrand-Rosenberg S and Sinha P:
Myeloid-derived suppressor cells: Linking inflammation and cancer.
J Immunol. 182:4499–4506. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Eruslanov E, Neuberger M, Daurkin I,
Perrin GQ, Algood C, Dahm P, Rosser C, Vieweg J, Gilbert SM and
Kusmartsev S: Circulating and tumor-infiltrating myeloid cell
subsets in patients with bladder cancer. Int J Cancer.
130:1109–1119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Corzo CA, Condamine T, Lu L, Cotter MJ,
Youn JI, Cheng P, Cho HI, Celis E, Quiceno DG, Padhya T, et al:
HIF-1α regulates function and differentiation of myeloid-derived
suppressor cells in the tumor microenvironment. J Exp Med.
207:2439–2453. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ghiringhelli F, Bruchard M, Chalmin F and
Rébé C: Production of adenosine by ectonucleotidases: A key factor
in tumor immunoescape. J Biomed Biotechnol. 2012:4737122012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Deaglio S, Dwyer KM, Gao W, Friedman D,
Usheva A, Erat A, Chen JF, Enjyoji K, Linden J, Oukka M, et al:
Adenosine generation catalyzed by CD39 and CD73 expressed on
regulatory T cells mediates immune suppression. J Exp Med.
204:1257–1265. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bergamin LS, Braganhol E, Zanin RF,
Edelweiss MI and Battastini AM: Ectonucleotidases in tumor cells
and tumor-associated immune cells: An overview. J Biomed
Biotechnol. 2012:9598482012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ryzhov S, Novitskiy SV, Goldstein AE,
Biktasova A, Blackburn MR, Biaggioni I, Dikov MM and Feoktistov I:
Adenosinergic regulation of the expansion and immunosuppressive
activity of CD11b+Gr1+ cells. J Immunol. 187:6120–6129. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zou W and Chen L: Inhibitory B7-family
molecules in the tumour microenvironment. Nat Rev Immunol.
8:467–477. 2008. View Article : Google Scholar : PubMed/NCBI
|