Introduction
In 2014, osteosarcoma was among the 10 most common
cancers in children and teenagers in the USA; about half of the 800
patients with osteosarcoma diagnosed annually are children and
teenagers (1). The long bones in the
legs and arms are the most common sites of osteosarcoma (2). Metastasis of osteosarcoma is determined
in >30% of patients who received only surgical treatment
(3). The lung is the most common
metastasis organ for osteosarcoma (3). According to a recent study, the
probability of successful treatment for non-metastatic osteosarcoma
is around 70%; however, following metastasis of osteosarcoma cells
to other organs, the treatment success rate may decrease by half,
down to as low as 20% (4). The
survival rate of patients with osteosarcoma treated with the
combined chemotherapy of cisplatin, doxorubicin and high-dose
methotrexate is ~70%, demonstrating the potential of this
chemotherapeutic regimen (5).
However, there is still room for improvement, and the side effects
of current chemotherapy should be reduced; therefore, research
focusing on safer and more effective therapeutic options is
urgently required.
Forkhead-box O3 (FOXO3) is a isomer of the FOX gene
family (6). FOXO3 is a
tumor-suppressive transcriptional factor that controls various
cellular processes, including cell cycle arrest, apoptosis and
tumor suppression, in various kinds of cancer (7,8). FOXO3 is
regulated via phosphorylation of specific serine/threonine residues
by oncogenic kinases in cancer cells (9). It has been reported that Akt
phosphorylates specific serine/threonine residues (Thr32, Ser253
and Ser315) on FOXO3, leading to the degradation of FOXO3 following
translocation from the nucleus to the cytoplasm (10). Translocation of FOXO3 into the
cytoplasm inhibits its tumor suppressive transcriptional activity,
which results in tumor development and progression (11,12).
Notably, numerous clinical studies using tissue microarrays (TMAs)
to characterize the association between the nuclear localization or
expression levels of FOXO3 and the survival rates of patients with
ovarian cancer have revealed that FOXO3 is a good prognostic
biomarker (13,14); thus, FOXO3 activation in cancer cells
may provide a promising strategy for developing anti-cancer
therapeutic drugs.
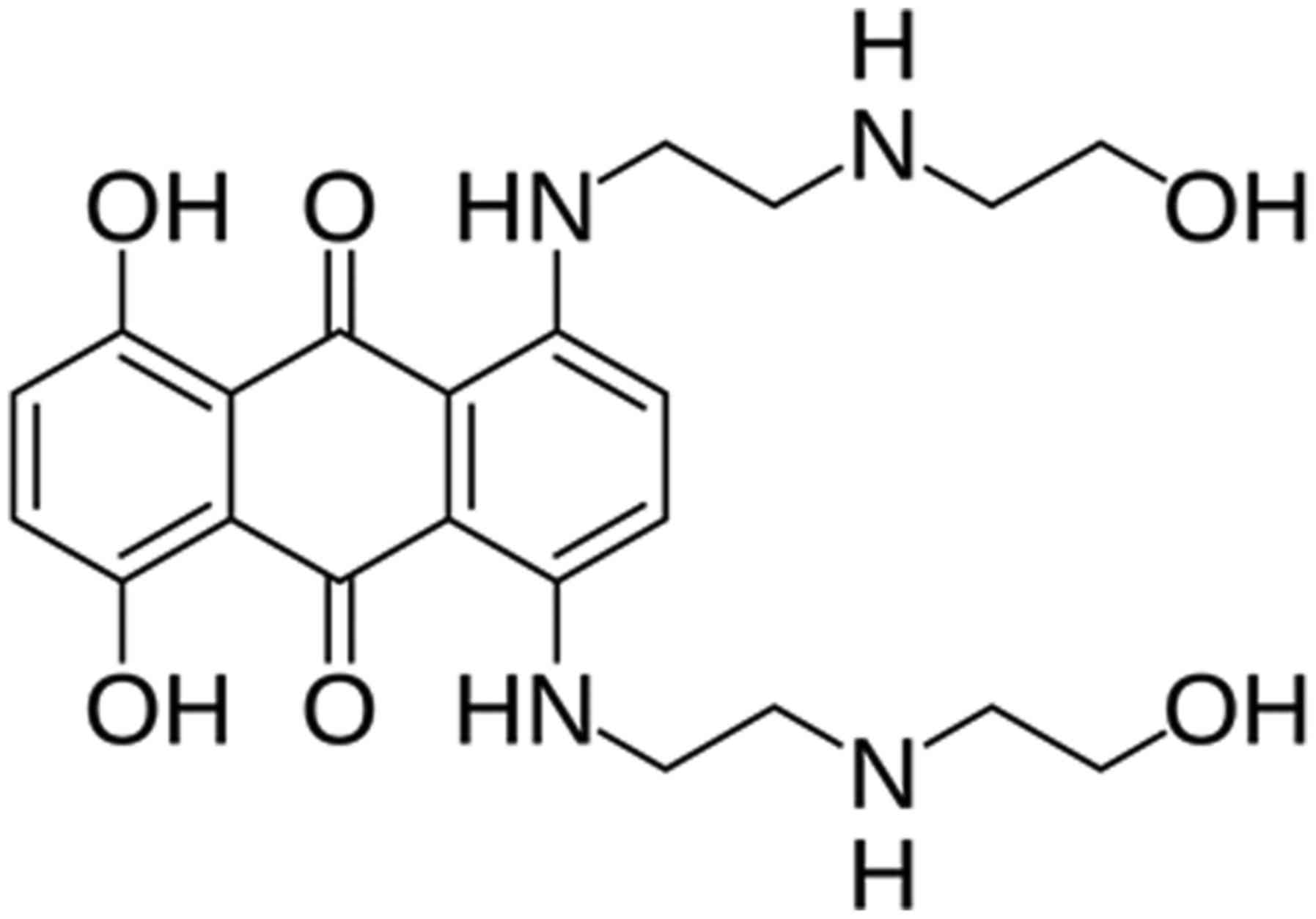
Mitoxantrone (MTZ; Fig.
1), a synthetic anti-tumor derivative of anthracycline
antibiotics, has been used largely for the treatment of tumor
types, including leukemia, lymphoma and prostate cancer, as well as
multiple sclerosis (15–18). MTZ is known to inhibit topoisomerase
II, which prevents the rejoining of DNA strands during the DNA
replication step, causing DNA damage by DNA double-strand breakage
(19). Consequently, MTZ affects the
cell cycle and induces apoptosis in cancer cells (20). In the present study, the potential of
MTZ as an anti-cancer therapeutic option for treating osteosarcoma
was demonstrated by indicating the underlying mechanism.
To develop a novel therapy against osteosarcoma, MTZ
was selected as a FOXO3 activator in osteosarcoma cells by applying
drug repositioning and investigating the anti-cancer activity and
underlying mechanisms of MTZ. Drug repositioning refers to the
application of previously clinically-used drugs against a specific
disease, then used for another disease by evaluating novel drug
activity, which saves time and cost for drug development (21). Based on the data, it was indicated
that MTZ treatment in osteosarcoma cells induces apoptosis through
FOXO3 upregulation, which increases pro-apoptotic genes but
decreases cell survival genes. FOXO3 is a promising candidate for
the development of osteosarcoma therapy, as these therapies may
sensitize osteosarcoma cells to FOXO3-mediated apoptosis and
suppress tumorigenesis.
Materials and methods
Cell culture
U2OS (p53 wild) and MG63 (p53 null) cells (American
Type Culture Collection, Manassas, VA, USA) were maintained in
Dulbecco's modified Eagle's media with 10% fetal bovine serum (FBS)
and 1% streptomycin/penicillin (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) at 37°C in a humidified incubator
containing 5% CO2 in air. Both cell lines were used at
passages 4–10 for all experiments.
Chemical reagents and antibodies
Mouse anti-β-actin antibody (cat. no. A5441), MTZ
(cat. no. M6545) and the following chemicals and solvents, dimethyl
sulfoxide (DMSO), glycerol, glycine, sodium chloride, Trizma base
and Tween20, were purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). Mouse anti-poly(ADP-ribose) polymerase(PARP)1
(cat. no. sc-8007), rabbit anti-FOXO3 (cat. no. sc-11351), mouse
anti-Lamin B1 (cat. no. sc-56145) and mouse anti-GAPDH antibodies
(cat. no. sc-32233) were purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). Rabbit anti-pAkt (cat. no. 9271), rabbit
anti-Akt (cat. no. 4691), rabbit anti-cleaved PARP1 (cat. no.
9541), rabbit anti-cleaved Caspase-3 (cat. no. 9664), rabbit
anti-Bax (cat. no. 5023), rabbit anti-Bim (cat. no. 2933), rabbit
anti-Bcl2 (cat. no. 2872) and rabbit anti-p27 antibodies (cat. no.
3686) were purchased from Cell Signaling Technology, Inc. (Danvers,
MA, USA). Goat anti-rabbit (cat. no. 111-035-003) and goat
anti-mouse (cat. no. 115-035-003) horseradish peroxidase-conjugated
IgG were obtained from Jackson ImmunoResearch Laboratories, Inc.
(West Grove, PA, USA). Enhanced chemiluminescence (ECL) reagents
were obtained from GenDEPOT (Barker, TX, USA).
WST-1 assay
Cells (1×103) were seeded in each well of
a 96-well plate and incubated for 18 h at 37°C in a humidified
incubator containing 5% CO2 in air. Following
incubation, cells were treated with DMSO (0.1%) as a control
vehicle and 0, 0.1, 0.2, 0.5, 0.8, and 1 µM indicated concentration
of MTZ for 72 h at 37°C. Following this, 20 µl WST-1 solution
(DoGenBio, Seoul, Korea) was added to each well for 4 h. The
visible absorbance at 460 nm for each well was then quantified
using a microplate reader. The assay was repeated 3 times.
Colony formation assay
Cells (0.5×103) were seeded in 6 cm
dishes and incubated for 18 h at 37°C in a humidified incubator
containing 5% CO2 in air. Following incubation, cells
were treated with DMSO (0.1%) as a control vehicle or the indicated
concentration of MTZ for 7 days. The colonies were washed twice
with PBS, fixed with 3.7% paraformaldehyde and stained with 1%
crystal violet solution in distilled water at room temperature for
15 min. Images were captured using the Chemi-doc detection system
(Bio-Rad Laboratoried, Hercules, CA, USA). The assay was repeated 3
times.
Western blot analysis
Cells were washed 3 times with PBS and lysed in
lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 2 mM EDTA, 1% Triton
X-100 and 0.1% SDS; pH 8.0) with protease and phosphatase
inhibitors (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Cell
lysates were centrifuged (10,000 × g at 4°C for 10 min). The
protein concentration was measured using a Bradford protein
determination assay (Bio-Rad Laboratories) and 20 µg protein was
loaded per the lane. Proteins were separated on 10% SDS-PAGE gels
and blotted onto nitrocellulose membranes (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). The membranes were blocked in 3% non-fat
dry milk for 1 h at room temperature and probed with primary
antibodies for FOXO3, pAkt, Akt, p27, LaminB1, GAPDH, cleaved
caspase3, Bim (EL), Bax, Bcl2, and β-actin. All antibodies were
diluted at 1:1,000 and incubated for 1 h at room temperature.
Membranes were then probed with HRP-tagged goat anti-mouse or
anti-rabbit IgG antibodies, diluted at 1:15,000 and 1:5,000
respectively, for 1 h at room temperature. Chemiluminescence was
detected using ECL.
Cell-based enzyme-linked immunosorbent
assay (ELISA)
U2OS cells [2×104/well in 100 µl DMEM
(Gibco; Thermo Fisher Scientific, Inc.)] were seeded into 96-well
plates and incubated at 37°C for 18 h in a CO2
incubator. Cells in each well were treated separately with DMSO
(negative control), LY294002 (Sigma-Aldrich; Merck KGaA) or
Wortmannin (positive control; Sigma-Aldrich; Merck KGaA), and
small-molecule compounds (20 µM/ml final conc.; http://www.enzolifesciences.com/BML-2843/screen-well-fda-approved-drug-library-v2/)
from the FDA-Approved Drug Library (Enzo Life Sciences, Inc.,
Farmingdale, NY, USA) (totaling 640 drugs) at 37°C and 5%
CO2 for 24 h. Then, cells were fixed and quenched by
adding 100 µl 4% formaldehyde (in PBS) at room temperature for 15
min and 100 µl 0.6% H2O2 (in PBS) at room
temperature for 15 min. To determine the level of the
phosphorylated-FOXO3-S318/321 in these cells, ELISA was performed
by treating cells with 100 µl blocking buffer [5% bovine serum
albumin (BSA) in Tris-buffered saline with 0.1% Tween-20 (TBST)] at
room temperature for 1 h. Subsequent to washing the cells with TBST
3 times, cells were treated with 100 µl primary antibody against
phospho-FOXO3-S318/321 (9465; 1:250 dilution in TBST containing 2%
BSA; Cell Signaling Technology, Inc.) at 4°C overnight. Subsequent
to washing, cells with TBST three times were treated with 100 µl
anti-rabbit IgG horseradish peroxidase-conjugated secondary
antibody (1:1,000 dilution in TBST containing 2% BSA; Jackson
ImmunoResearch Laboratories, Inc.) at room temperature for 2 h.
Following washing cells with TBST 3 times, cells were treated with
100 µl 3,3′,5,5′-tetramethylbenzidine peroxidase substrate
(Sigma-Aldrich; Merck KGaA) for 15 min at room temperature,
followed by adding 50 µl ELISA stop solution (2N
H2SO4; Sigma-Aldrich; Merck KGaA). The
optical density of each well was measured by reading the microplate
using a microplate reader (Molecular Devices, LLC, Sunnyvale, CA,
USA) at 450 nm.
Cytoplasmic and nuclear protein
fractionation
Cells were washed 3 times with PBS and lysed in
cytoplasmic fractional buffer [10 mM
4-(2-hydroxyethyl)-1-piperazineethanesulphonic acid (HEPES), pH
8.0, 50 mM NaCl, 500 mM sucrose, 1 mM EDTA, 0.5 mM spermidine, 0.15
mM spermine, 0.2% Triton X-100, 1 mM dithiothreitol, 2 µM
phenylmethylsulfonyl fluoride and 0.15 U/ml aprotinin] at 4°C for
30 min. Cell lysates were centrifuged (10,000 × g at 4°C for 30
min) and the supernatant was collected for the cytoplasmic
fraction. The pellet was washed twice with the washing buffer (10
mM HEPES pH 8.0, 50 mM NaCl, 25% glycerol, 0.1 mM EDTA, 0.5 mM
spermidine and 0.15 mM spermine) and lysed with nuclear fractional
buffer (10 mM HEPES pH 8, 350 mM NaCl, 25% glycerol, 0.1 mM EDTA,
0.5 mM spermidine and 0.15 mM spermine) at 4°C for 30 min. Lysates
were centrifuged (10,000 × g at 4°C for 30 min) and the supernatant
was collected for the nuclear fraction. All of buffers used in this
experiment were prepared in the Park laboratory (Sejong, Korea),
based on the previous report (12).
Immunofluorescence analysis
Cells (5×105) were seeded in 6 cm dishes
and incubated for 18 h at 37°C in a humidified incubator containing
5% CO2 in air. Following incubation, cells were treated
with DMSO (0.1%) as a control vehicle and 1 µM MTZ for 2 h sat
37°C. Cells were fixed with 4% paraformaldehyde solution for 15 min
at room temperature, permeabilized with Triton X-100 (0.2%),
blocked with bovine serum albumin and incubated with the primary
antibody against FOXO3 (diluted, 1,500) at room temperature for 1
h, followed by Alexa 594-conjugated anti-mouse secondary antibody
(cat. no. A-11037; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) a room temperature for 1 h. Following
counterstaining with DAPI at room temperature for 1 h, fluorescence
images were captured with a Zeiss LSM510 confocal microscope at
magnification, ×100.
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labeling
(TUNEL) assay
Cells (5×105) were seeded in 6 cm dishes
and incubated for 18 h at 37°C in a humidified incubator containing
5% CO2 in air. Following incubation, cells were treated
with DMSO (0.1%) as a control vehicle and 0.5 and 1 µM MTZ for 2 h.
Cells were fixed with 4% paraformaldehyde solution for 15 min at
room temperature and permeabilized with Triton X-100 (0.2%).
Apoptosis was determined by enzymatic labeling of DNA strand breaks
with a TUNEL assay kit (the DeadEnd Fluorometric TUNEL System;
Promega Corporation, Madison, WI, USA), according to the
manufacturer's procedures. Fluoroshield Mounting Medium with DAPI
(Abcam, Cambridge, UK) was used, and 3 fields of view were imaged
using the Olympus CKX53 light microscope (Olympus Corporation,
Tokyo, Japan; magnification, ×100).
Annexin V staining analysis
Cells (5×105) were seeded in 6-cm dishes
and incubated for 18 h at 37°C in a humidified incubator containing
5% CO2 in air. Following incubation, cells were treated
with DMSO (0.1%) as control vehicle and 0.5 and 1 µM MTZ for 2 h.
Cells were washed 3 times with PBS, trypsinized with Gibco™
Trypsin-EDTA (0.25%) with Phenol red (Gibco; Thermo Fisher
Scientific, Inc.) and resuspended in binding buffer from the
Annexin V Apoptosis Detection kit I (BD Pharmingen; BD
Biosciences). Cells were analyzed using FACSCalibur (BD
Biosciences, Franklin Lakes, NJ, USA) and the data were analyzed by
FlowJo (version 10; De Novo Software, Ashland, Oregon, USA). A
total of 10,000 events were collected in each run. The percentage
of cells that underwent apoptosis was determined using the
fluorescein isothiocyanate with propidium iodide from the Annexin V
Apoptosis Detection kit I, according to the manufacturer's
procedures.
siRNA transfection
Cells (5×105) were seeded in 6-cm dishes
and incubated for 18 h at 37ls that humidified incubator containing
5% CO2 in air. Following incubation, cells were
transfected with control (GE Healthcare Dharmacon, Inc., Lafayette,
CO, USA) or FOXO3 siRNA (GE Healthcare Dharmacon, Inc.) using the
ratio of 1 nM siRNA: 3 µl Dharmafect (GE Healthcare Dharmacon,
Inc.) in 300 µl serum free media for 6 h at 37°C in a humidified
incubator containing 5% CO2 in air. Following this, cell
culture media was replaced with the fresh media containing 10% FBS
and cells were incubated for 18 h.
Statistical analysis
Data are expressed as the mean ± standard error of
three independent experiments. Differences between groups were
analyzed with one-way analysis of variance with Tukey's post-hoc
test when the variances were equal with SPSS software (version
19.0; IBM Corp., Armonk, NY, USA). All statistical tests were
two-sided. P<0.05 was considered to indicate a statistically
significant difference.
Results
MTZ induces nuclear localization and
activation of FOXO3 in osteosarcoma cells
To identify small molecules that can activate FOXO3
in U2OS cells, a osteosarcoma cell line, a cell-based enzymatic
ELISA was performed to screen small molecules that may
significantly reduce the phosphorylation of Ser318/321 in FOXO3
(pS318/321 FOXO3), which is usually located in the cytoplasm of
cells (10). Various FOXO3
phosphorylation sites have been identified by a number of kinases,
and these serve a key role in regulating the cellular location and
transcriptional activity of FOXO3. Among these, Ser318 and Ser322
have been investigated by Rena et al (22). They indicated that the phosphorylation
of Ser318 by Akt induces the subsequent phosphorylation of Ser322
by casein kinase 1, which is critical for translocation of FOXO3
from the nucleus into the cytosol. In previous studies (11,12,23),
attempts were made to develop a novel cell-based ELISA assay system
using a phospho-FOXO3 antibody to screen small molecules causing
nuclear localization of FOXO3. Following testing various
commercially available phospho-FOXO3 antibodies including pThr32
and pSer253, which are possible Akt-phosphorylation sites
identified by Brunet et al (10), pSer318/321 FOXO3 antibody was selected
as the best antibody for the cell-based ELISA system, due to a
larger difference in the readout value being detected between the
positive controls (Wortmannin and LY294002) and the negative
control. Following screening small molecule drugs from a
commercially available food and drug administration (FDA)-approved
drug library (SCREEN-WELL® FDA approved drug library V2:
BML-2843-0100; Enzo Life Sciences, Inc.), MTZ was focused on
(Fig. 1), which indicated a decreased
level (>50%) of pS318/321 FOXO3, compared with the DMSO vehicle
control in U2OS cells (data not shown).
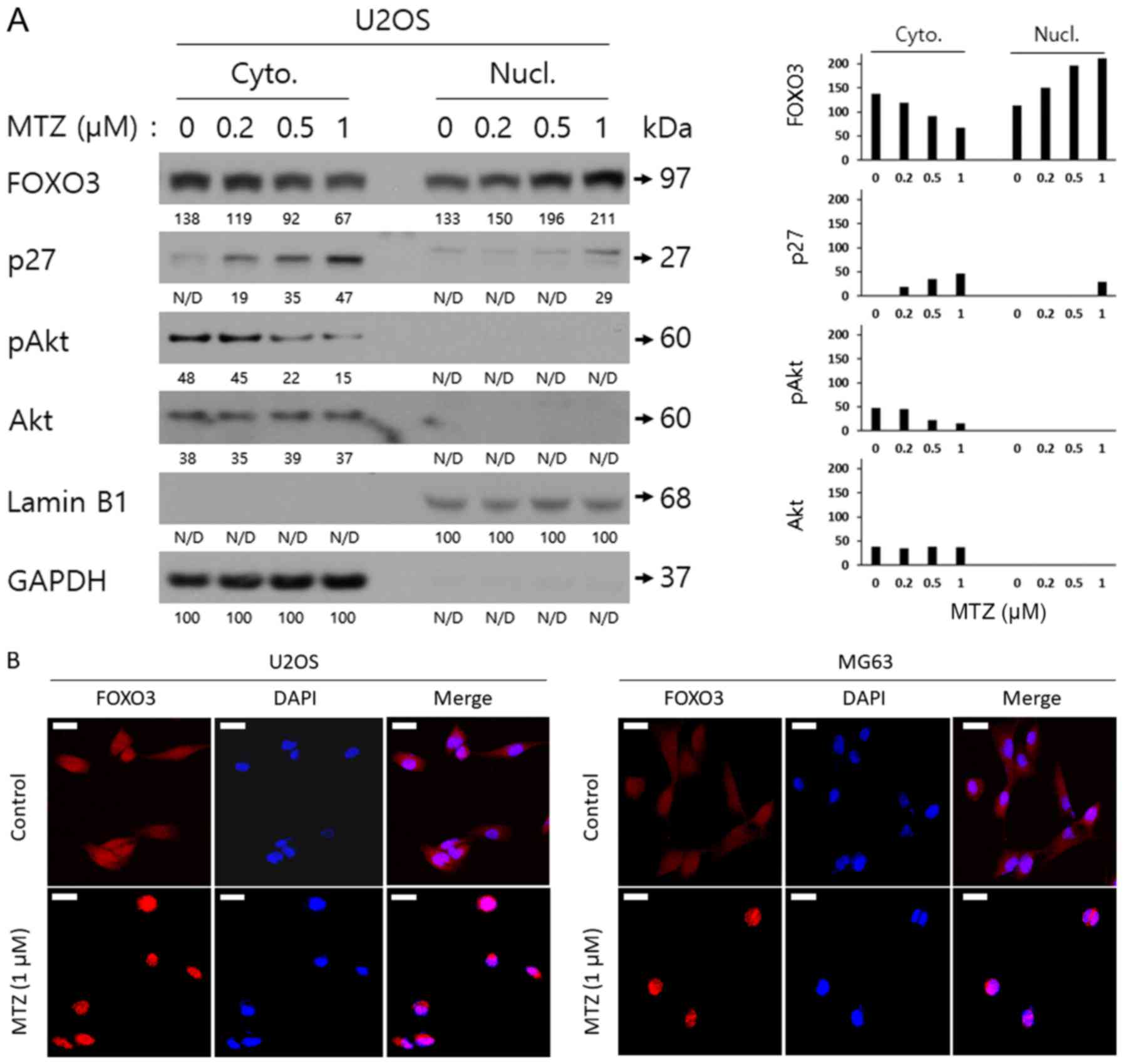
To demonstrate the mechanism by which MTZ treatment
decreases pS318/321 FOXO3 in U2OS cells, nuclear fractional western
blot analyses with lysates from U2OS cells previously treated with
various doses of MTZ were performed. As depicted in Fig. 2A, MTZ treatment reduced the expression
of Akt-pS473 (pS473 Akt) in the cytoplasm, in a dose-dependent
manner. In addition, the expression of FOXO3 in the cytoplasm was
decreased by MTZ treatment, whilst the expression of FOXO3 in the
nucleus was significantly increased by MTZ treatment, in a
dose-dependent manner. The expression of the cytoplasmic and
nuclear p27 protein was significantly increased by MTZ treatment,
in a dose-dependent manner. Additionally, it was confirmed that
FOXO3 translocates into the nucleus following MTZ treatment, by
carrying out confocal microscopy analysis (Fig. 2B). These results indicated that MTZ
may downregulate pS473 Akt, leading to the translocation and
activation of the FOXO3 protein from the cytoplasm to the nucleus
in osteosarcoma cells.
MTZ suppresses cell survival and
promotes apoptosis in osteosarcoma cells
To investigate the anti-cancer activity of MTZ
against osteosarcoma cells, U2OS and MG63 osteosarcoma cells were
treated with MTZ and growth rate of the cells was measured using
the WST-1 and colony formation assays. Cell viability was
determined using WST-1 assay, which involved incubating the U2OS
and MG63 cells with MTZ (0, 0.1, 0.2, 0.5, 0.8 and 1.0 for 72 h. As
depicted in Fig. 3A, MTZ had an
inhibitory effect on osteosarcoma cell growth, in a dose-dependent
manner. Notably, treatment with 0.5, 0.8 and 1 µM MTZ resulted in a
statistically significant (P<0.05) inhibition of cell viability;
therefore, based on the data on cell viability/cytotoxicity, it may
be concluded that ≥0.5 µM MTZ is the optimal dose for the
subsequent experiments. In addition, clonogenic assay results
demonstrated that MTZ treatment markedly suppressed the
colony-forming ability of osteosarcoma cells (Fig. 3B). Taken together, these results
indicated that MTZ displays a potent inhibitory effect on the cell
survival/proliferation of osteosarcoma cells. To examine the effect
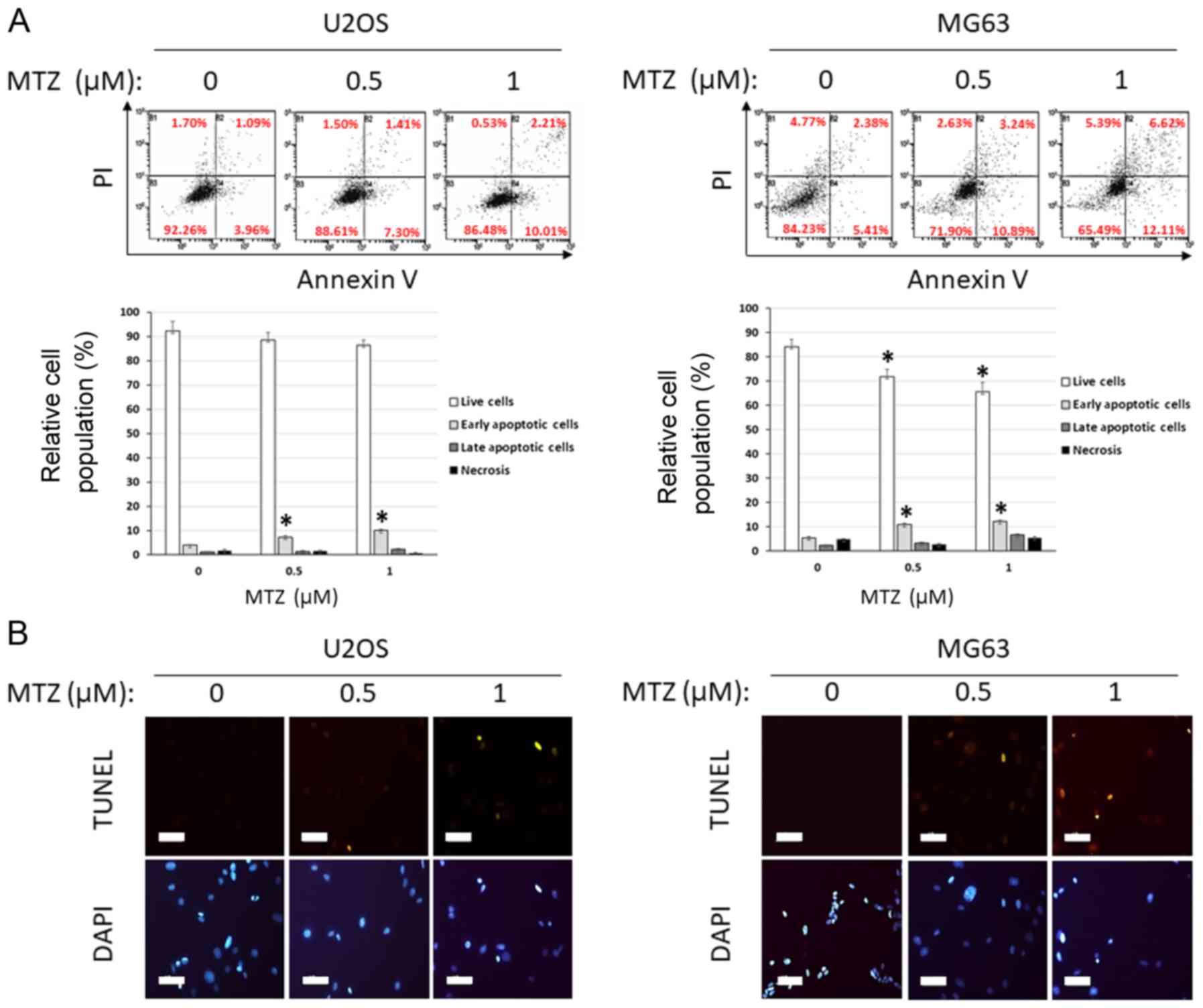
of MTZ on apoptosis in osteosarcoma cells, western blot analyses,
TUNEL assays and Annexin V staining analysis were performed. MTZ
treatment increased the cleavage of Caspase-3 and PARP1. MTZ
treatment also increased the expression of Bax and Bim extra-large
(EL) in osteosarcoma cells; however, MTZ treatment decreased the
expression of Bcl-2 (Fig. 3C). These
results indicated that the mechanism underlying MTZ and its
apoptotic activity may be through a Caspase-3-mediated mechanism in
osteosarcoma cells, upregulation of the pro-apoptotic Bax and Bim
and downregulation of the anti-apoptotic Bcl2. In addition,
compared with the DMSO control, MTZ treatment of osteosarcoma cells
resulted in an increase in the amount of Annexin V stained cells as
well as TUNEL-positive cells, both of which are typical markers of
apoptosis (Fig. 4A and B). Thus, the
present results indicated that MTZ treatment may induce cellular
apoptosis in osteosarcoma cells.
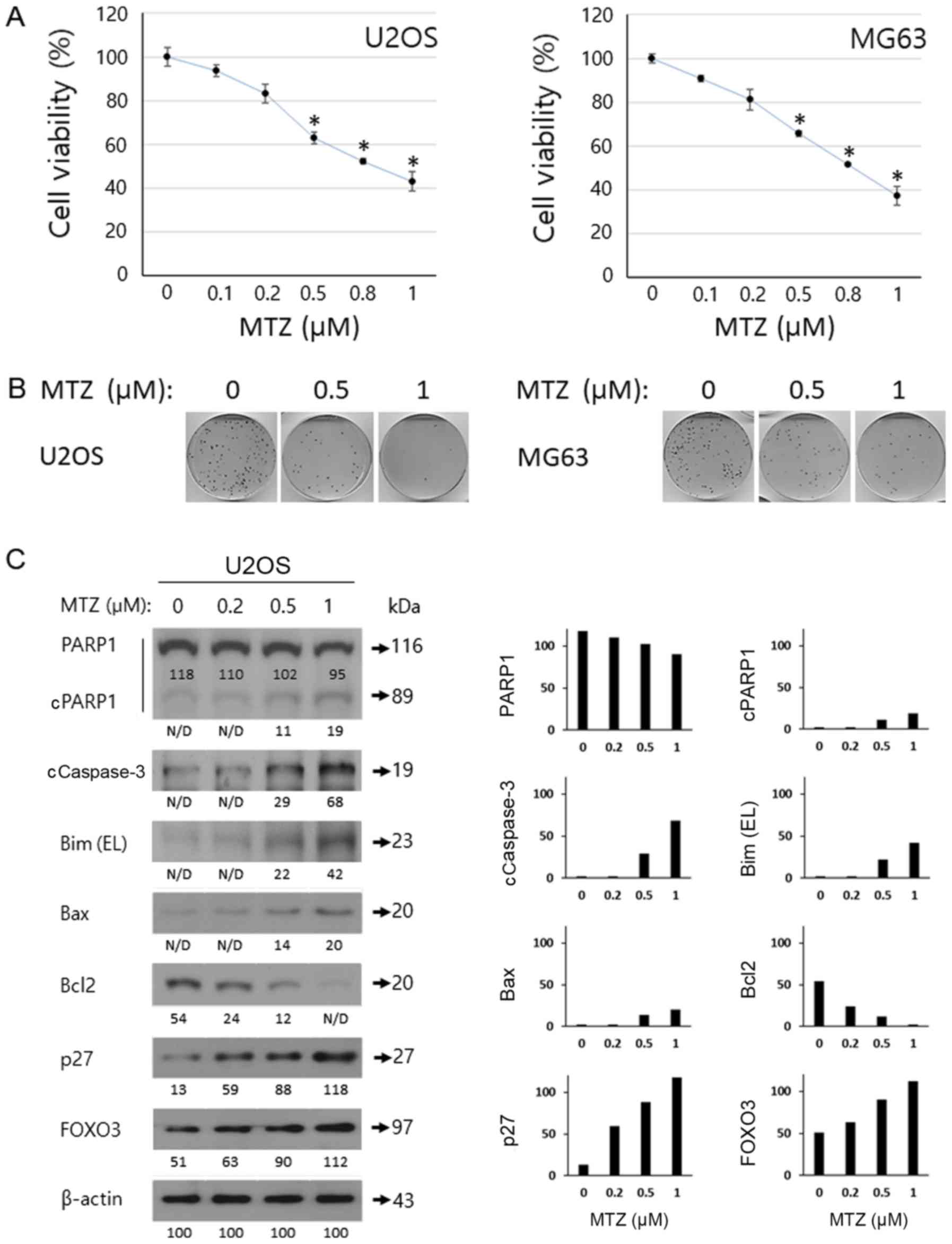 | Figure 3.Anti-cancer effect of MTZ against
osteosarcoma cells. (A) Dose-dependent effects of MTZ (0, 0.1, 0.2,
0.5, 0.8 and 1 cancer effect of MTZ against ost following 72 h
incubation. The cell viability was determined by WST-1 assay and
the relative cell survival rate was calculated by dividing the
optical density of each treatment condition by the optical density
of the control (DMSO) treatment. The mean of 3 biological
replicates is presented and the error bars represent standard error
of the mean. *P<0.05. (B) Dose-dependent effect of MTZ (0.5 and
1 µM) on U2OS and MG63 cells following 7 days. Represented images
of 3 scanned images are provided. (C) Apoptosis analysis of U2OS
cells treated with MTZ and western blotting results of U2OS cells
treated with MTZ (0, 0.2, 0.5, and 1 µM) for 48 h. β-actin was used
for a gel-loading control. Quantitative data are based on the
relative ratio of the indicated protein to β-actin. MTZ,
mitoxantrone; EL, extra-large; FOXO3, forkhead-box O3; PARP1,
poly(ADP-ribose) polymerase; c, cleaved. |
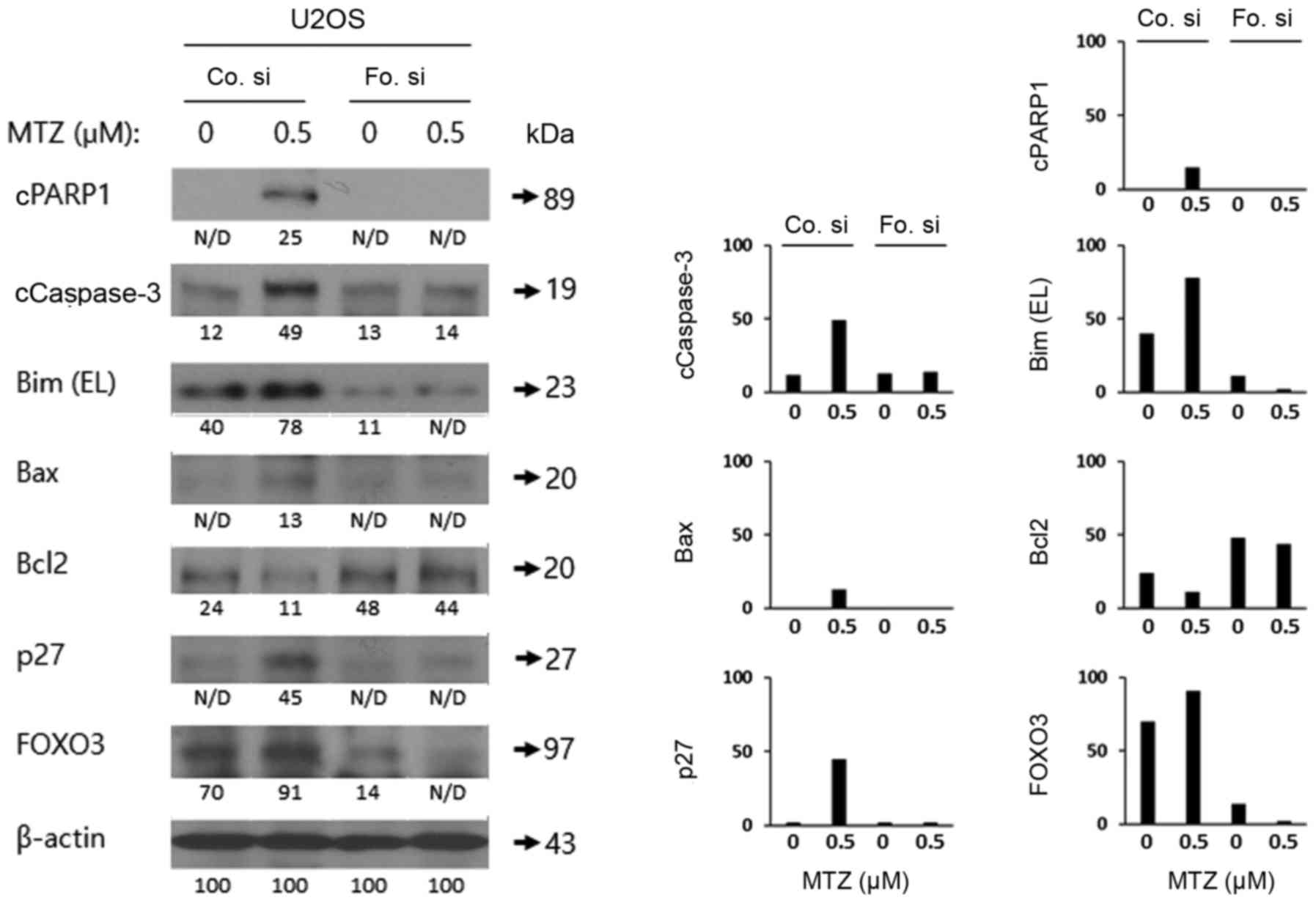
Knockdown of FOXO3 decreases
MTZ-mediated apoptosis in osteosarcoma cells
To examine the role of FOXO3 in MTZ-induced
apoptosis in osteosarcoma cells, FOXO3 expression was knocked down
in U2OS cells by transfecting siRNA against FOXO3 or control siRNA.
Additionally, western blot analysis of PARP1, Caspase-3, Bax, Bim
EL, Bcl2 and FOXO3 was performed. As depicted in Fig. 5, FOXO3 knockdown markedly attenuated
Caspase-3 and PARP1 cleavage and reduced the expression levels of
Bax and the Bim EL isoform, compared with the control siRNA.
However, MTZ treatment decreased the reduced the expression levels
of Bcl-2. Thus, the present results indicated that FOXO3 may serve
a key role in MTZ-mediated apoptosis in U2OS cells.
Discussion
Osteosarcoma was demonstrated in 2014 to be the most
common malignant tumor type of childhood (1) The incidence of osteosarcoma in young
children is >10% (24). There are
three types of osteosarcoma, low, middle and high grade
osteosarcoma (25). In cases of
low-grade osteosarcoma, such as parosteal osteosarcoma, patients
have a good prognosis, with a 91% five-year survival rate following
surgery alone (26). In middle-grade
osteosarcoma, although surgery has success occasionally, the
effectiveness of additional chemotherapy has not been clearly
demonstrated (27,28). High-grade osteosarcoma is the most
common subtype and is responsible for ~80% of osteosarcomas. The
use of combined chemotherapy before or following surgery may
improve the survival rate of patients by >60%, compared with
surgery alone (29). Thus, although
combined aggressive chemotherapy using usually DNA damaging agent
has been developed, >30% of patients with osteosarcoma still
have metastatic cancer in other tissues and the survival rate with
metastases is <1 year (3);
therefore, more effective therapeutic options are required for the
treatment of osteosarcomas.
There are standard combined chemotherapy treatment
protocols for osteosarcoma depending on the stage (first and
second-line treatment). First line treatment includes the
combination of: i) Cisplatin and doxorubicin; ii) cisplatin,
doxorubicin and methotrexate; iii) etoposide and ifosfamide; or iv)
cisplatin, epirubicin and ifosfamide (30–32).
Second line treatment involves the combination or sole use of: i)
gemcitabine and docetaxel; ii) etoposide and cyclophosphamide; iii)
topotecan and cyclophosphamide; iv) gemcitabine; v) etoposide and
ifosfamide; vi) carboplatin, etoposide and ifosfamide; vii)
etoposide, ifosfamide and methotrexate; or viii)
Samarium-153-ethylene diamine tetramethylene phosphonate (33–40).
However, several recent reports have demonstrated that resistance
against these chemotherapies requires consideration (41–43). There
are several publications dealing with p53 mutations in patients
with osteosarcoma (44–46). p53 is a tumor-suppressive
transcription factor that mediates apoptosis following DNA damage
in cells (47,48); therefore, it is difficult to
successfully treat patients with osteosarcoma with p53 mutation by
using chemotherapy, which activates p53. However, there are no
studies regarding FOXO3 mutations in patients with osteosarcoma;
therefore, it is beneficial to consider novel combination
chemotherapies that may activate FOXO3 using MTZ and other
FDA-approved drugs (11). Future
studies will examine the synergic effect of MTZ with other
FOXO3-activating small molecules, with the goal of inhibiting
osteosarcoma cell development.
In the present study, the aim was to identify small
molecules that activate FOXO3, and as a result FOXO3-mediated
apoptosis, in osteosarcoma cell lines, which can be used as a novel
therapeutic target. Previously, it was demonstrated that
trifluoperazine and bepridil may promote the translocation of FOXO3
into the nucleus in triple-negative breast cancer (TNBC) cells,
leading to suppression of TNBC in vitro and in vivo
(11). In the present study, using
the same drug discovery platform (cell-based ELISA), based on the
phosphorylation status of FOXO3 in U2OS cells, a possible FOXO3
activator from an FDA-approved drug library was screened. The focus
was on MTZ, a type of DNA damaging agent, for the present study due
to MTZ displaying the strongest activity, decreasing pS318/321
FOXO3 by >50%, compared with the DMSO vehicle control in U2OS
cells (data not shown). Recently, Tarrado-Castellarnau et al
(49) reported that methylseleninic
acid was a selenium supplement that may have anti-cancer activity
through the translocation of FOXO3 from the cytoplasm into the
nucleus following the inhibition of Akt phosphorylation in U2OS
cells stably expressing FOXO3; however, the endogenous FOXO3
expression levels or subcellular localization by methylseleninic
acid in U2OS cells were not examined. In the present study, it was
confirmed that MTZ induces nuclear localization and activation of
FOXO3 in osteosarcoma cells. In addition, Guzmán-Pérez et al
(50) demonstrated that
benzylglucosinolate-derived isothiocyanate from tropaeolum majus
promoted the translocation of FOXO1 from the cytoplasm into the
nucleus following inhibition of Akt phosphorylation in U2OS cells
stably expressing FOXO1. FOXO1 and FOXO3 are members of the human
FOX gene family, which includes the distinct forkhead DNA-binding
domain (6). Thus, they are known to
have very similar cellular functions, including cell cycle arrest
and cell death, as tumor-suppressive transcriptional factors
(49). Indeed, there have been
several previous reports indicating that FOXO1 has anti-tumor
activity in osteosarcoma (50–52);
therefore, it would be beneficial to study FOXO1 expression levels
in osteosarcoma cells with MTZ.
Akt has numerous substrates that are phosphorylated,
leading to activation or inhibition in cancer cells (7,53).
Previous research has investigated the effect of targeted therapies
to treat various cancer types (53).
In the present study, it was demonstrated that MTZ significantly
inhibits pS473 Akt phosphorylation and causes nuclear localization
and activation of FOXO3 in osteosarcoma cells (Fig. 2A and B). MTZ treatment inhibits
proliferation of osteosarcoma cells in vitro, whereas
silencing FOXO3 potently attenuates MTZ-mediated apoptosis in
osteosarcoma cells. Similar to the present study, Yuan et al
(54) evaluated the anti-cancer
activity of MTZ analogs in Huh-7 human hepatoma cells and
demonstrated that this MTZ analog inhibited the phosphorylation of
Akt. Hu et al (55) reported
that combination administration of MTZ and prednisolone, a steroid
medication, may be one option for tumor therapy. They indicated
that this combination displayed synergistic anti-cancer activity
through inhibition of the Akt pathway, including phosphorylated
glycogen synthase kinase 3β, ps6 ribosomal protein and pAMPK
(55). Future studies will elucidate
the molecular mechanism that MTZ treatment in osteosarcoma cells
inhibits the phosphorylation of Akt, by investigating the upstream
signaling proteins of Akt.
The aim was to demonstrate that MTZ displays
anti-cancer activity through the regulation of the Akt/FOXO3
pathway independent of p53 status, using osteosarcoma cell lines.
Although p53 serves a key role in cell cycle arrest and apoptosis
in numerous cancer types (12,54,56,57),
induction of cell cycle arrest and apoptosis in cancer cells by
FOXO3 activation does not depend on p53 expression or status. For
example, Matsuda et al (56)
reported that treatment with OTS167, a maternal embryonic leucine
zipper kinase (MELK) inhibitor, induced the upregulation of p21
protein expression in HCT116-p53(−/-) p53-null cells. They
indicated that FOXO1 and FOXO3, two known tumor-suppressive
Forkhead type transcriptional factors, may regulate p21 expression
via activation following de-phosphorylation through treatment with
a MELK inhibitor, independent of p53 status. Furthermore, Zhang
et al (58) demonstrated that
a tetrandrine derivative may potentially exert anti-cancer activity
via induction of PUMA with a p53-independent mechanism. In that
study, anti-cancer activity of tetrandrine derivatives were
observed in wild-type and p53-null cancer cells, and inhibition of
Akt/FOXO3 signaling may enhance tetrandrine derivative-mediated
PUMA expression; therefore, consistent with previous reports, it
was demonstrated that MTZ induces apoptosis in U2OS (p53 wild) and
MG63 (p53 null) osteosarcoma cell lines and have identified a novel
mechanism that MTZ regulates Akt/FOXO3, leading to the upregulation
of p27, Bax and Bim and downregulation of Bcl-2. Further studies
should knockdown p53 expression in U2OS cells and perform the same
experiments, including confocal analysis, western blotting analysis
and cell viability assay, of the present study to determine if the
MTZ treatment has the same anti-cancer activity as p53 wild type
U2OS cells.
To the best of our knowledge, this is the first
report demonstrating that regulation of the Akt/FOXO3 signaling
pathway though treatment with MTZ is involved in controlling
osteosarcoma cell development. Future studies should perform TMA
analysis of patients with osteosarcoma, which will support the
results of the present study and provide the clinical basis for a
novel anti-osteosarcoma therapeutic strategy. Taken together, these
results indicated that the anti-cancer activity of MTZ treatment
may induce FOXO3-mediated apoptotic signaling pathways, and its
application may be a novel chemotherapeutic agent for
osteosarcoma.
Acknowledgements
Not applicable.
Funding
This research was supported by the Basic Science
Research Program through the National Research Foundation of Korea
NRF) funded by the Ministry of Education, Science and Technology
(NRF-2014R1A6A3A04054307 and 2015R1A2A2A01003762).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Author's contributions
SHP, JL, MAK, KYJ and JRK conceived and designed the
experiments, Performed the experiments, analyzed the data, wrote
the paper. KYJ and JRK take responsibility for the integrity of the
data analysis. All authors reviewed the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Moore DD and Luu HH: Osteosarcoma. Cancer
Treat Res. 162:65–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lamoureux F, Trichet V, Chipoy C,
Blanchard F, Gouin F and Redini F: Recent advances in the
management of osteosarcoma and forthcoming therapeutic strategies.
Expert Rev Anticancer Ther. 7:169–181. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Luetke A, Meyers PA, Lewis I and Juergens
H: Osteosarcoma treatment-where do we stand? A state of the art
review. Cancer Treat Rev. 40:523–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sakamoto A and Iwamoto Y: Current status
and perspectives regarding the treatment of osteo-sarcoma:
Chemotherapy. Rev Recent Clin Trials. 3:228–231. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chou AJ and Gorlick R: Chemotherapy
resistance in osteosarcoma: Current challenges and future
directions. Expert Rev Anticancer Ther. 6:1075–1085. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Katoh M and Katoh M: Human FOX gene family
(Review). Int J Oncol. 25:1495–1500. 2004.PubMed/NCBI
|
|
7
|
Alvarez B, Martinez-A C, Burgering BM and
Carrera AC: Forkhead transcription factors contribute to execution
of the mitotic programme in mammals. Nature. 413:744–747. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Furukawa-Hibi Y, Kobayashi Y, Chen C and
Motoyama N: FOXO transcription factors in cell-cycle regulation and
the response to oxidative stress. Antioxid Redox Signal. 7:752–760.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fu Z and Tindall DJ: FOXOs, cancer and
regulation of apoptosis. Oncogene. 27:2312–2319. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo
P, Hu LS, Anderson MJ, Arden KC, Blenis J and Greenberg ME: Akt
promotes cell survival by phosphorylating and inhibiting a forkhead
transcription factor. Cell. 96:857–868. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Park SH, Chung YM, Ma J, Yang Q, Berek JS
and Hu MC: Pharmacological activation of FOXO3 suppresses
triple-negative breast cancer in vitro and in vivo. Oncotarget.
7:42110–42125. 2016.PubMed/NCBI
|
|
12
|
Park SH, Lee JH, Berek JS and Hu MC:
Auranofin displays anticancer activity against ovarian cancer cells
through FOXO3 activation independent of p53. Int J Oncol.
45:1691–1698. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fei M, Zhao Y, Wang Y, Lu M, Cheng C,
Huang X, Zhang D, Lu J, He S and Shen A: Low expression of Foxo3a
is associated with poor prognosis in ovarian cancer patients.
Cancer Invest. 27:52–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu M, Zhao Y, Xu F, Wang Y, Xiang J and
Chen D: The expression and prognosis of FOXO3a and Skp2 in human
ovarian cancer. Med Oncol. 29:3409–3415. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jancic J, Nikolic B, Ivancevic N, Djuric
V, Zaletel I, Stevanovic D, Peric S, van den Anker JN and Samardzic
J: Multiple sclerosis in pediatrics: Current concepts and treatment
options. Neurol Ther. 5:131–143. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Atwal M, Lishman EL, Austin CA and Cowell
IG: Myeloperoxidase enhances etoposide and mitoxantrone-mediated
DNA damage: A target for myeloprotection in cancer chemotherapy.
Mol Pharmacol. 91:49–57. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Casadei B, Pellegrini C, Pulsoni A,
Annechini G, De Renzo A, Stefoni V, Broccoli A, Gandolfi L, Quirini
F, Tonialini L, et al: 90-yttrium-ibritumomab tiuxetan
consolidation of fludarabine, mitoxantrone, rituximab in
intermediate/high-risk follicular lymphoma: Updated long-term
results after a median follow-up of 7 years. Cancer Med.
5:1093–1097. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nussbaum N, George DJ, Abernethy AP, Dolan
CM, Oestreicher N, Flanders S and Dorff TB: Patient experience in
the treatment of metastatic castration-resistant prostate cancer:
State of the science. Prostate Cancer Prostatic Dis. 19:111–121.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hasinoff BB, Wu X, Patel D, Kanagasabai R,
Karmahapatra S and Yalowich JC: Mechanisms of action and reduced
cardiotoxicity of pixantrone; a topoisomerase II targeting agent
with cellular selectivity for the topoisomerase IIα isoform. J
Pharmacol Exp Ther. 356:397–409. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ferrer A, Marce S, Bellosillo B, Marcé S,
Bellosillo B, Villamor N, Bosch F, López-Guillermo A, Espinet B,
Solé F, Montserrat E, Campo E and Colomer D: Activation of
mitochondrial apoptotic pathway in mantle cell lymphoma: High
sensitivity to mitoxantrone in cases with functional DNA-damage
response genes. Oncogene. 23:8941–8949. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chong CR and Sullivan DJ Jr: New uses for
old drugs. Nature. 448:645–646. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rena G, Woods YL, Prescott AR, Peggie M,
Unterman TG, Williams MR and Cohen P: Two novel phosphorylation
sites on FKHR that are critical for its nuclear exclusion. EMBO J.
21:2263–2271. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bae JS, Lee J, Park Y, Park K, Kim JR, Cho
DH, Jang KY and Park SH: Attenuation of MUC4 potentiates the
anticancer activity of auranofin via regulation of the
Her2/Akt/FOXO3 pathway in ovarian cancer cells. Oncol Rep.
38:2417–2425. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
ESMO/European Sarcoma Network Working
Group: Bone sarcomas: ESMO clinical practice guidelines for
diagnosis, treatment and follow-up. Ann Oncol. 25:iii113–iii123.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zambo I and Veselý K: WHO classification
of tumours of soft tissue and bone 2013: The main changes compared
to the 3rd edition. Cesk Patol. 50:64–70. 2014.(In Czech).
PubMed/NCBI
|
|
26
|
Schajowicz F, McGuire MH, Araujo Santini
E, Muscolo DL and Gitelis S: Osteosarcomas arising on the surfaces
of long bones. J Bone Joint Surg Am. 70:555–564. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Grimer RJ, Bielack S, Flege S, Cannon SR,
Foleras G, Andreeff I, Sokolov T, Taminiau A, Dominkus M,
San-Julian M, et al: Periosteal osteosarcoma-a European review of
outcome. Eur J Cancer. 41:2806–2811. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cesari M, Alberghini M, Vanel D, Palmerini
E, Staals EL, Longhi A, Abate M, Ferrari C, Balladelli A and
Ferrari S: Periosteal osteosarcoma: A single-institution
experience. Cancer. 117:1731–1735. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bernthal NM, Federman N, Eilber FR, Nelson
SD, Eckardt JJ, Eilber FC and Tap WD: Long-term results (>25
years) of a randomized, prospective clinical trial evaluating
chemotherapy in patients with high-grade, operable osteosarcoma.
Cancer. 118:5888–5893. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bielack SS, Smeland S, Whelan JS, Marina
N, Jovic G, Hook JM, Krailo MD, Gebhardt M, Pápai Z, Meyer J, et
al: Methotrexate, doxorubicin, and cisplatin (MAP) plus maintenance
pegylated interferon Alfa-2b versus MAP alone in patients with
resectable high-grade osteosarcoma and good histologic response to
preoperative MAP: First results of the EURAMOS-1 good response
randomized controlled trial. J Clin Oncol. 33:2279–2287. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Benjamin RS, Wagner MJ, Livingston JA,
Ravi V and Patel SR: Chemotherapy for bone sarcomas in adults: The
MD anderson experience. Am Soc Clin Oncol Educ Book. e656–e660.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Su W, Lai Z, Wu F, Lin Y, Mo Y, Yang Z and
Wu J: Clinical efficacy of preoperative chemotherapy with or
without ifosfamide in patients with osteosarcoma of the extremity:
Meta-analysis of randomized controlled trials. Med Oncol.
32:4812015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Navid F, Willert JR, McCarville MB, Furman
W, Watkins A, Roberts W and Daw NC: Combination of gemcitabine and
docetaxel in the treatment of children and young adults with
refractory bone sarcoma. Cancer. 113:419–425. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rodriguez-Galindo C, Daw NC, Kaste SC,
Meyer WH, Dome JS, Pappo AS, Rao BN and Pratt CB: Treatment of
refractory osteosarcoma with fractionated cyclophosphamide and
etoposide. J Pediatr Hematol Oncol. 24:250–255. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Saylors RL 3rd, Stine KC, Sullivan J,
Kepner JL, Wall DA, Bernstein ML, Harris MB, Hayashi R and Vietti
TJ: Pediatric Oncology Group: Cyclophosphamide plus topotecan in
children with recurrent or refractory solid tumors: A pediatric
oncology group phase II study. J Clin Oncol. 19:3463–3469. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Merimsky O, Meller I, Kollender Y, Issakov
J, Flusser G and Inbar M: Gemcitabine in bone sarcoma resistant to
doxorubicin-based chemotherapy. Sarcoma. 4:7–10. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Goorin AM, Harris MB, Bernstein M,
Ferguson W, Devidas M, Siegal GP, Gebhardt MC, Schwartz CL, Link M
and Grier HE: Phase II/III trial of etoposide and high-dose
ifosfamide in newly diagnosed metastatic osteosarcoma: A pediatric
oncology group trial. J Clin Oncol. 20:426–433. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Van Winkle P, Angiolillo A, Krailo M,
Cheung YK, Anderson B, Davenport V, Reaman G and Cairo MS:
Ifosfamide, carboplatin, and etoposide (ICE) reinduction
chemotherapy in a large cohort of children and adolescents with
recurrent/refractory sarcoma: The children's cancer group (CCG)
experience. Pediatr Blood Cancer. 44:338–347. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Michelagnoli MP, Lewis IJ, Gattamaneni HR,
Bailey CC and Lashford LS: Ifosfamide/etoposide alternating with
high-dose methotrexate: Evaluation of a chemotherapy regimen for
poor-risk osteosarcoma. Br J Cancer. 79:1174–1178. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Anderson PM, Wiseman GA, Dispenzieri A,
Arndt CA, Hartmann LC, Smithson WA, Mullan BP and Bruland OS:
High-dose samarium-153 ethylene diamine tetramethylene phosphonate:
Low toxicity of skeletal irradiation in patients with osteosarcoma
and bone metastases. J Clin Oncol. 20:189–196. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hattinger CM, Vella S, Tavanti E, Fanelli
M, Picci P and Serra M: Pharmacogenomics of second-line drugs used
for treatment of unresponsive or relapsed osteosarcoma patients.
Pharmacogenomics. 17:2097–2114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu T, Li Z, Zhang Q, De Amorim Bernstein
K, Lozano-Calderon S, Choy E, Hornicek FJ and Duan Z: Targeting
ABCB1 (MDR1) in multi-drug resistant osteosarcoma cells using the
CRISPR-Cas9 system to reverse drug resistance. Oncotarget.
7:83502–83513. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Buondonno I, Gazzano E, Jean SR, Audrito
V, Kopecka J, Fanelli M, Salaroglio IC, Costamagna C, Roato I,
Mungo E, et al: Mitochondria-targeted doxorubicin: A new
therapeutic strategy against doxorubicin-resistant osteosarcoma.
Mol Cancer Ther. 15:2640–2652. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen Z, Guo J, Zhang K and Guo Y: TP53
mutations and survival in osteosarcoma patients: A meta-analysis of
published data. Dis Markers. 2016:46395752016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ru JY, Cong Y, Kang WB, Yu L, Guo T and
Zhao JN: Polymorphisms in TP53 are associated with risk and
survival of osteosarcoma in a Chinese population. Int J Clin Exp
Pathol. 8:3198–3203. 2015.PubMed/NCBI
|
|
46
|
Yao D, Cai GH, Chen J, Ling R, Wu SX and
Li YP: Prognostic value of p53 alterations in human osteosarcoma: A
meta analysis. Int J Clin Exp Pathol. 7:6725–6733. 2014.PubMed/NCBI
|
|
47
|
Pakos EE, Kyzas PA and Ioannidis JP:
Prognostic significance of TP53 tumor suppressor gene expression
and mutations in human osteosarcoma: A meta-analysis. Clin Cancer
Res. 10:6208–6214. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ozaki T, Wu D, Sugimoto H, Nagase H and
Nakagawara A: Runt-related transcription factor 2 (RUNX2) inhibits
p53-dependent apoptosis through the collaboration with HDAC6 in
response to DNA damage. Cell Death Dis. 4:e6102013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tarrado-Castellarnau M, Cortes R, Zanuy M,
Tarragó-Celada J, Polat IH, Hill R, Fan TW, Link W and Cascante M:
Methylseleninic acid promotes antitumour effects via nuclear FOXO3a
translocation through Akt inhibition. Pharmacol Res. 102:218–234.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Guzman-Perez V, Bumke-Vogt C, Schreiner M,
Mewis I, Borchert A and Pfeiffer AF: Benzylglucosinolate Derived
Isothiocyanate from Tropaeolum majus Reduces Gluconeogenic Gene and
Protein Expression in Human Cells. PLoS One. 11:e01623972016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jin S, Pang RP, Shen JN, Huang G, Wang J
and Zhou JG: Grifolin induces apoptosis via inhibition of PI3K/AKT
signalling pathway in human osteosarcoma cells. Apoptosis.
12:1317–1326. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pei H, Jin Z, Chen S, Sun X, Yu J and Guo
W: MiR-135b promotes proliferation and invasion of osteosarcoma
cells via targeting FOXO1. Mol Cell Biochem. 400:245–252. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Toker A and Marmiroli S: Signaling
specificity in the Akt pathway in biology and disease. Adv Biol
Regul. 55:28–38. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yuan CL, Lin SW and Cheng MH: Inhibition
of Molecular Signaling in Huh-7 Cells by AM3: A Novel
Chemotherapeutic Agent for Hepatocellular Carcinoma. Med Chem.
49–56. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hu T, Cao H, Yang C, Zhang L, Jiang X, Gao
X, Yang F, He G, Song X, Tong A, et al: LHD-modified
mechanism-based liposome coencapsulation of Mitoxantrone and
Prednisolone using novel lipid bilayer fusion for tissue-specific
colocalization and synergistic antitumor effects. ACS App Mater
Interfaces. 8:6586–6601. 2016. View Article : Google Scholar
|
|
56
|
Matsuda T, Kato T, Kiyotani K, Tarhan YE,
Saloura V, Chung S, Ueda K, Nakamura Y and Park JH: p53-independent
p21 induction by MELK inhibition. Oncotarget. 8:57938–57947. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liu H, Yin J, Wang C, Gu Y, Deng M and He
Z: FOXO3a mediates the cytotoxic effects of cisplatin in lung
cancer cells. Anticancer Drugs. 25:898–907. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang YX, Liu XM, Wang J, Li J, Liu Y,
Zhang H, Yu XW and Wei N: Inhibition of AKT/FoxO3a signaling
induced PUMA expression in response to p53-independent cytotoxic
effects of H1: A derivative of tetrandrine. Cancer Biol Ther.
16:965–975. 2015. View Article : Google Scholar : PubMed/NCBI
|