Introduction
Osteosarcoma (OS) is one of the most malignant types
of bone tumor, which is most frequent in children and adolescents
and characterized by the formation of neoplastic bone tissue
(1). There is a high tendency to for
OS to undergo metastasis (2). The
treatment of OS typically includes surgery, radiation,
chemotherapy, or the combination of chemotherapy and radiotherapy;
however, these treatments are not successful in the long-term
(3). Numerous studies have indicated
the abilities of natural products to inhibit the development of
different types of cancer in multiple animal models, as previously
reviewed (4). These compounds may be
used as anti-proliferation or anti-metastasis agents.
Sennoside, an extract from senna, is widely used as
a stimulant laxative and its safety and efficiency have been
established (5); however, little is
known regarding its usefulness as an anti-tumor agent. Ellagic acid
(EA) is a polyphenol compound with strong antioxidant properties
that is found as ellagitannins in the fruits and nuts of several
plants. The oral administration of EA can protect the system from
alcohol toxicity by decreasing the expression of liver marker
enzymes and increasing the activity of the antioxidant cascade
(6,7);
it was previously demonstrated to induce apoptosis and cell cycle
arrest in various types of cancer cell (8–11).
However, its potential anti-tumor role in osteosarcoma and the
mechanisms for its effects remain elusive.
The c-Jun N-terminal kinase (JNK) signal
transduction pathway is associated with malignant cellular
transformation (12). Studies with
cultured cells have demonstrated that the JNK signal transduction
pathway participates in the proliferation, differentiation,
apoptosis and metastasis of osteoblasts (13). Phosphorylated JNKs activate the Jun
proto-oncogene, AP-1 transcription factor subunit (c-Jun), which
homodimerizes and/or heterodimerizes with Fos proto-oncogene, AP-1
transcription factor subunit to generate the activator protein-1
transcription complex (AP-1) (14),
which binds to specific DNA sequences at target promoters, and
regulates the expression of cognate genes that participate in the
differentiation and function of osteoblasts, as well as in the
pathogenesis of osteosarcoma (15–17).
It the present study, it is reported that EA and
Sennoside B can inhibit Saos-2 and MG63, two malignant osteosarcoma
cell lines, growth, migration and invasion, and induce cell cycle
arrest by the inhibition of c-Jun expression.
Materials and methods
Antibodies and reagents
Rabbit anti-poly(ADP-ribose) polymerase (PARP; cat
no. 88817), p-p38 (cat no. 4511), p-extracellular signal-regulated
kinase (ERK) 1/2 (cat no. 4370) and c-Jun (cat no. 9165) antibodies
were obtained from Cell Signaling Technology, Inc. (Danvers, MA,
USA); mouse anti-GAPDH (cat no. sc-32233) was purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). The EA, Sennoside B and
propidium iodide were obtained from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany).
Cell culture
Saos-2 and MG63 human osteosarcoma cells were
obtained from the American Type Culture Collection (Manassas, VA,
USA) and were maintained in Dulbecco's modified Eagle's medium
(DMEM; Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) with
10% fetal bovine serum (Hyclone; GE Healthcare Life Sciences), 100
U/ml penicillin and 100 µg/ml streptomycin at 37°C in a humidified
5% CO2 atmosphere.
Wound closure assay
Saos-2 and MG63 cells were plated in a 12-well cell
culture plate. When the cell density reached ≥90%, the monolayer
was scratched with a 200 µl pipette. Wound closure was monitored
with phase contrast microscopy and quantified as the relative wound
closure rate (wound distance at a specific time point/the original
wound distance).
Transwell assay
A total of 1×104 of Saos-2 or MG63 cells
were seeded in the upper chamber of Transwell plates (BD
Bioscience) with Matrigel, in serum-free conditions. DMEM
supplemented with 10% fetal calf serum (Hyclone; GE Healthcare Life
Sciences) and 50 ug/ml fibronectin (BD Biosciences, Franklin Lakes,
NJ, USA) was used as a chemoattractant in the lower chamber. At 48
h, cells remaining on the upper chamber were removed with a cotton
swab, while cells adhering to the lower membrane were stained with
0.1% crystal violet for 30 min at room temperature and photographed
with an inverted microscope (Zeiss AG, Oberkochen, Germany). The
area of positive staining was measured using image analysis
software (Image-Pro Plus 6.0; Media Cybernetics, Inc., Rockville,
MD, USA); the rate of invasion was calculated as the positive area
percentage. At least three independent experiments were performed
for each condition.
MTT assays
Cell viability was assessed with an MTT assay in
replicates. Cells were seeded in 96-well plates at
2.5×103 cells per well (Saos-2 cells) or
1.0×103 cells per well (MG63 cells) and incubated for 0,
24, 48 or 72 h. Then the medium was replaced with 200 ml fresh DMEM
containing 0.5 mg/ml MTT, the cells were incubated for a further 4
h, then MTT formazan crystals were dissolved in DMSO and absorbance
at 490 nm was measured and analyzed.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
To assess mRNA levels, RNA was isolated from cells
using TRIzol reagent (Takara Biotechnology Co., Ltd., Dalian,
China) and cDNA was synthesized using the MLV reserve transcriptase
from Promega Corporation (Madison, WI, USA), according to the
manufacturers' protocols. The cDNA was subjected to qPCR (95.0°C
for 10 min for 1 cycle, then 95.0°C for 15 sec followed by 60.0°C
for 1 min for 40 cycles) using SYBR-green master mix (Toyobo Life
Science, Osaka, Japan) in the Mx3005P Real-Time PCR system
(Stratagene; Agilent Technologies, Inc., Santa Clara, CA, USA). The
comparative cycle quantification method (2∆∆Cq) was
employed to analysis the gene expression with housekeeping gene 18S
as an internal normalization control (18). Each experiment was performed in
duplicate and repeated three times. The primers for qPCR were as
follows: c-Jun forwards, TCCAAGTGCCGAAAAAGGAAG; reverse,
CGAGTTCTGAGCTTTCAAGGT; 18S forwards, GGACACGGACAGGATTGACA; reverse,
GACATCTAAGGGCATCACAG. I8S was used as an internal control.
Cell cycle analysis
Saos-2 cells or MG63 cells were treated with EA or
Sennoside B for 24 h. Cells were trypsinized, collected, fixed and
stained with propidium iodide solution following the manufacturer's
protocol of the Propidium Iodide Flow Cytometry kit (cat no.
ab139418, Abcam, Cambridge, UK). The cell cycle distribution was
then analyzed by the BD FACSCalibur (BD Biosciences) using
CellQuest Pro, version 5.1 (BD Biosciences).
Western blotting
Saos-2 or MG63 cells were treated with EA or
Sennoside B for 24 h, then scraped into ice-cold PBS and lysed with
lysis buffer containing 50 mM Tris-HCl (pH 7.5), 1 mM EDTA, 1%
Nonidet P-40, 150 mM NaCl, 10% glycerol and protease inhibitors.
The protein concentration was determined using the Pierce BCA
protein assay kit (cat no. 23225, Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Same quantities of protein (25 µg/lane) were
resolved using 12% PAGE-SDS gel, transferred to a nitrocellulose
membrane (Bio-Rad Laboratories, Inc., Hercules, CA, USA; cat no.
1620112), then followed by 5% non-fat milk blocking at room
temperature for 1 h. The proteins were analyzed with antibodies
against PARP (1:1,000 diluted in 5% BSA), p-ERK (1:2,000 diluted in
5% BSA), c-Jun (1:1,000 diluted in 5% BSA), p-p38 (1:1,000 diluted
in 5% BSA), and GAPDH (1:1,000 diluted in 5% BSA). The primary
antibodies were incubated with membrane at 4°C overnight, followed
by a 1 h secondary antibody incubation at room temperature.
HRP-conjugated goat anti-mouse (Jackson ImmunoResearch
Laboratories, Inc., West Grove, PA, USA; cat no. 115-035-146;
1:5,000 in 5% non-fat milk) and goat anti-rabbit (Jackson
ImmunoResearch Laboratories, Inc.; cat no. 115-035-045; 1:5,000 in
5% non-fat milk) secondary antibodies were used to visualize the
protein of interest using Amersham ECL Prime Western Blotting
Detection Reagent (cat no. RPN2236; GE Healthcare, Chicago, IL,
USA). All data were the results from 3 replicates.
Statistics
Statistical analysis was performed with GraphPad
Prism 6.0 (GraphPad Software, Inc., La Jolla, CA, USA).
Quantitative data are expressed as the means ± standard deviation.
Statistical analysis was performed with a one-way analysis of
variance followed by Dunnett's multiple comparisons test. P<0.05
was considered to indicate a statistically significant
difference.
Results
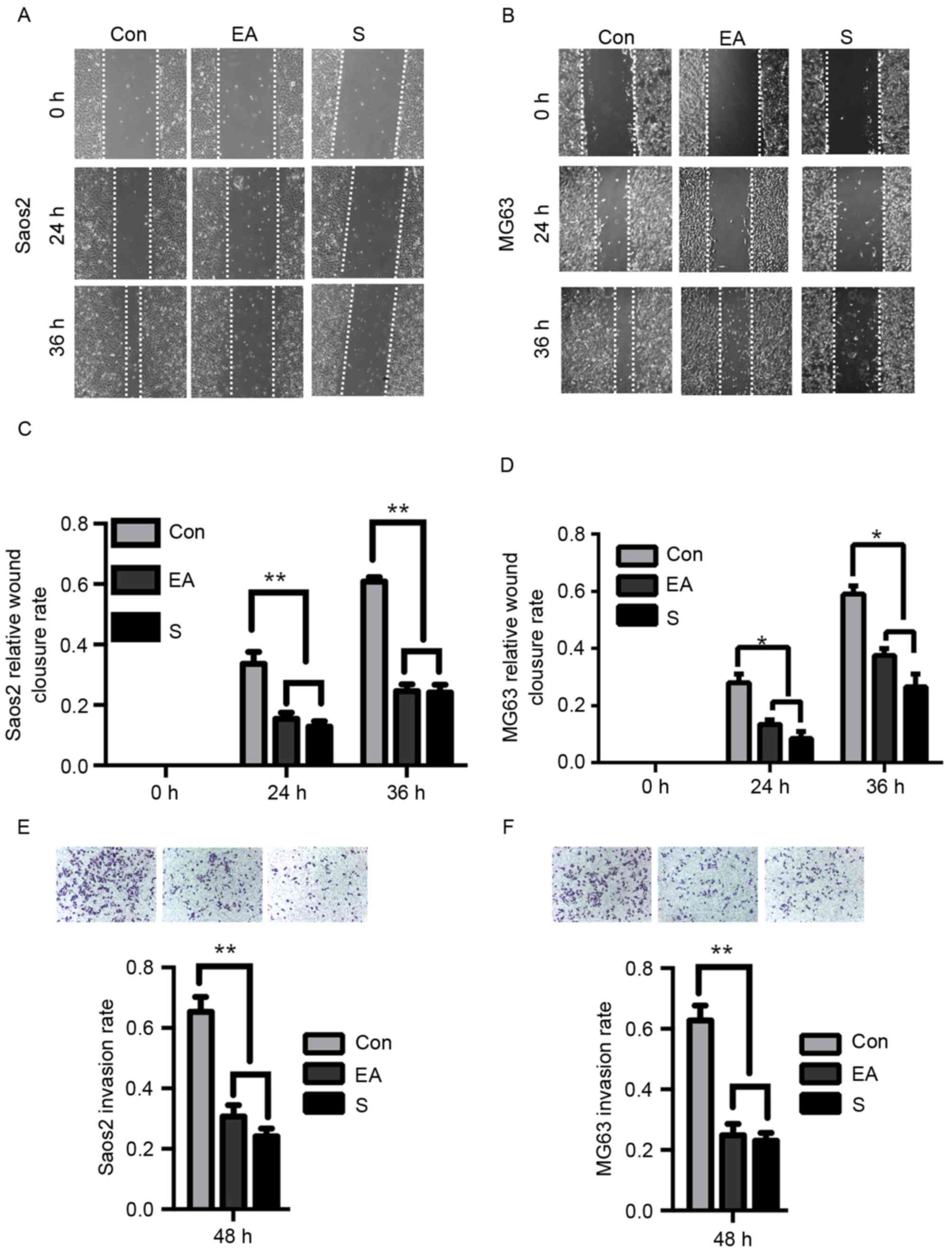
EA and Sennoside B inhibit the
migration and invasion of osteosarcoma cells
Previous reports have indicated the anti-tumor
potential of EA and Sennoside B (8).
In order to investigate their effect on osteosarcoma, Saos-2 and
MG63 cells were analyzed. A series of concentrations of each agent
were used in a wound-healing assay to verify their influence on the
migration of OS cells. It was indicated that the 20 µM EA or
Sennoside B was the minimal requirement for the inhibition of the
migration ability of Saos-2 cells (data not shown and Fig. 1A), which was consistent with the
result for MG63 cells (data not shown and Fig. 1B). The analysis of the wound-healing
assay demonstrated that when the cells were exposed to 20 µM EA or
Sennoside B for 24 or 36 h, the inhibition rate was 40–70%
(Fig. 1C and D; Saos-2, P<0.01;
MG63, P<0.05). Then, the invasion abilities of these cells in
the presence or absence of EA or Sennoside B were examined by
transwell assays. The EA- or Sennoside B-treated cells invaded
significantly less frequently than the control cells (Fig. 1E and F; P<0.01). Thus, it was
determined that EA or Sennoside B treatment can suppress the
migration and invasion of osteosarcoma cells.
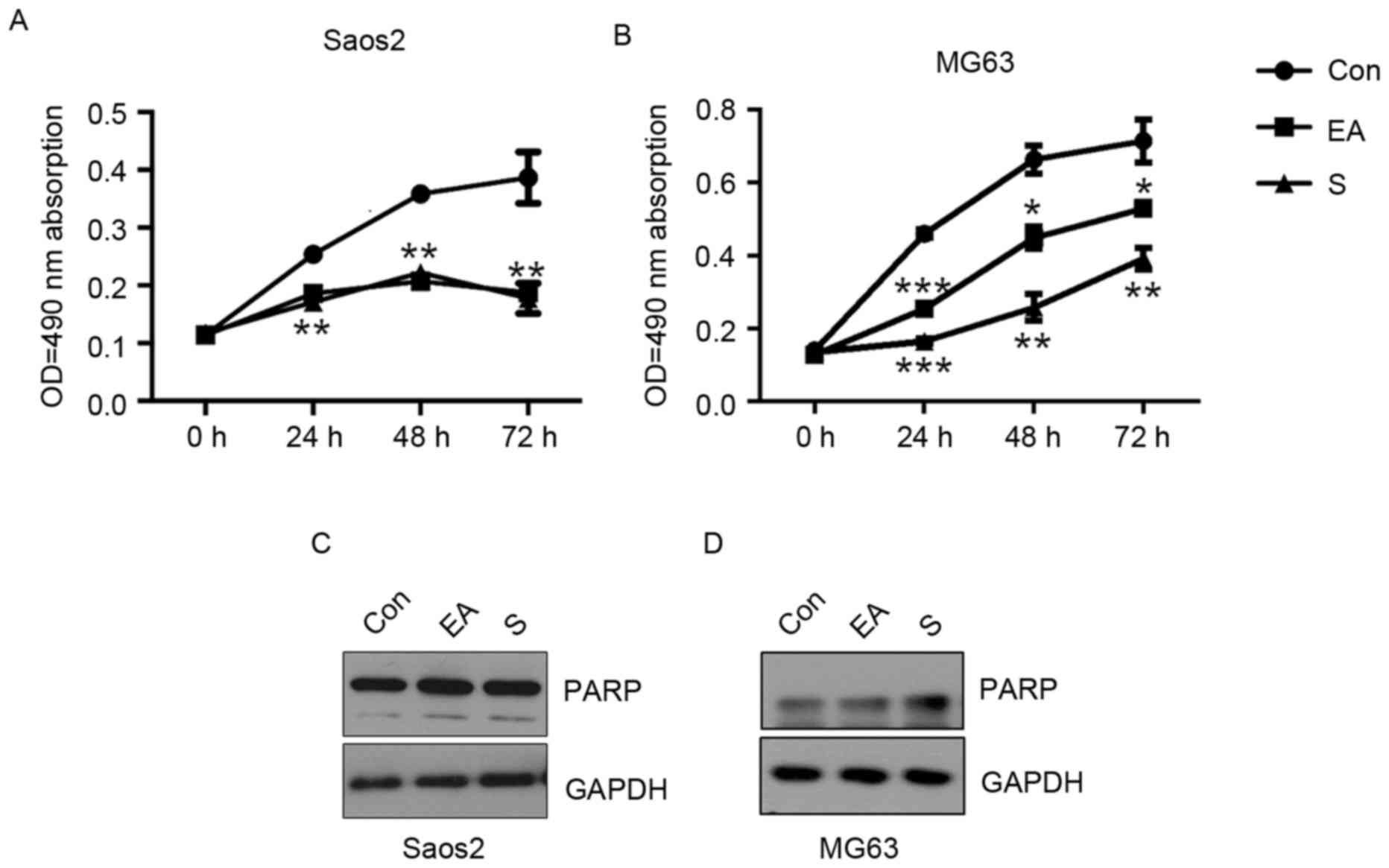
EA and Sennoside B inhibit the growth
of osteosarcoma cells without inducing apoptosis
Saos-2 and MG63 cells were treated with 20 µM EA or
Sennoside B, and viability was examined by an MTT assay. The
results suggested both EA and Sennoside B significantly inhibited
cell growth from 24 h at 20 µM, compared with the control groups
(Fig. 2A and B; Saos-2, P<0.01;
MG63, P<0.0001). To assess whether the induction of apoptosis
served a role in the repression of the proliferation of OS cells,
the cells were subjected to EA and Sennoside B treatment for 24 h
and were collected for western blotting. No significant difference
in cleaved PARP expression was observed between the control and
treated groups (Fig. 2C and D), even
though a previous study demonstrated that EA inhibited
proliferation by the induction of apoptosis in colon and prostate
cancer cells (19).
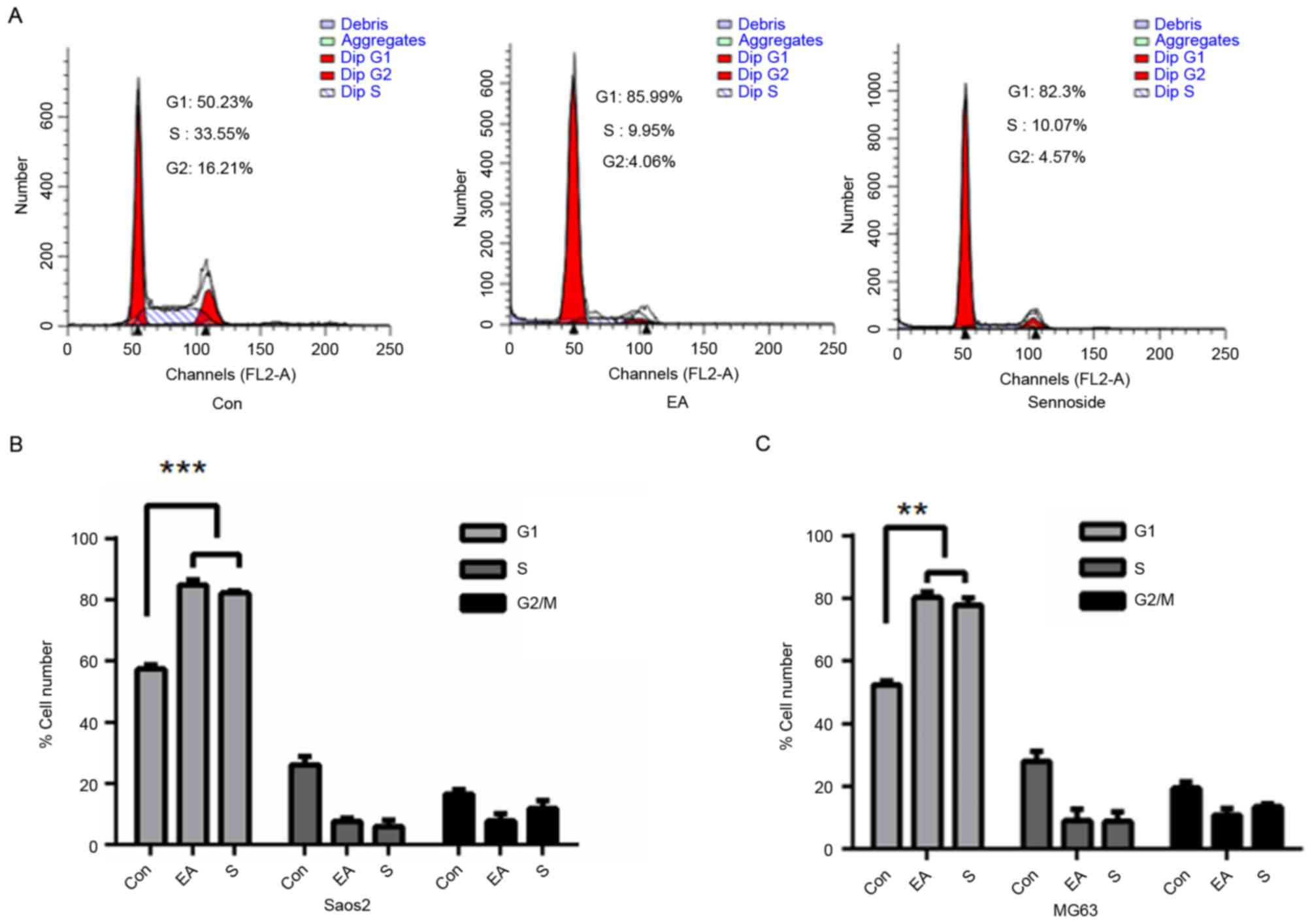
EA and Sennoside B induce cell cycle
arrest
As EA and Sennoside B inhibited cell growth without
inducing apoptosis, the cell cycle distribution of cells treated
with 20 µM EA or Sennoside B for 24 h was analyzed. It was
identified that >80% of Saos-2 cells in the treated groups were
in the G1 stage, compared with <60% in the control
groups (Fig. 3A and B; P<0.0001).
Similar results were detected for MG63 cells (Fig. 3C; P<0.01).
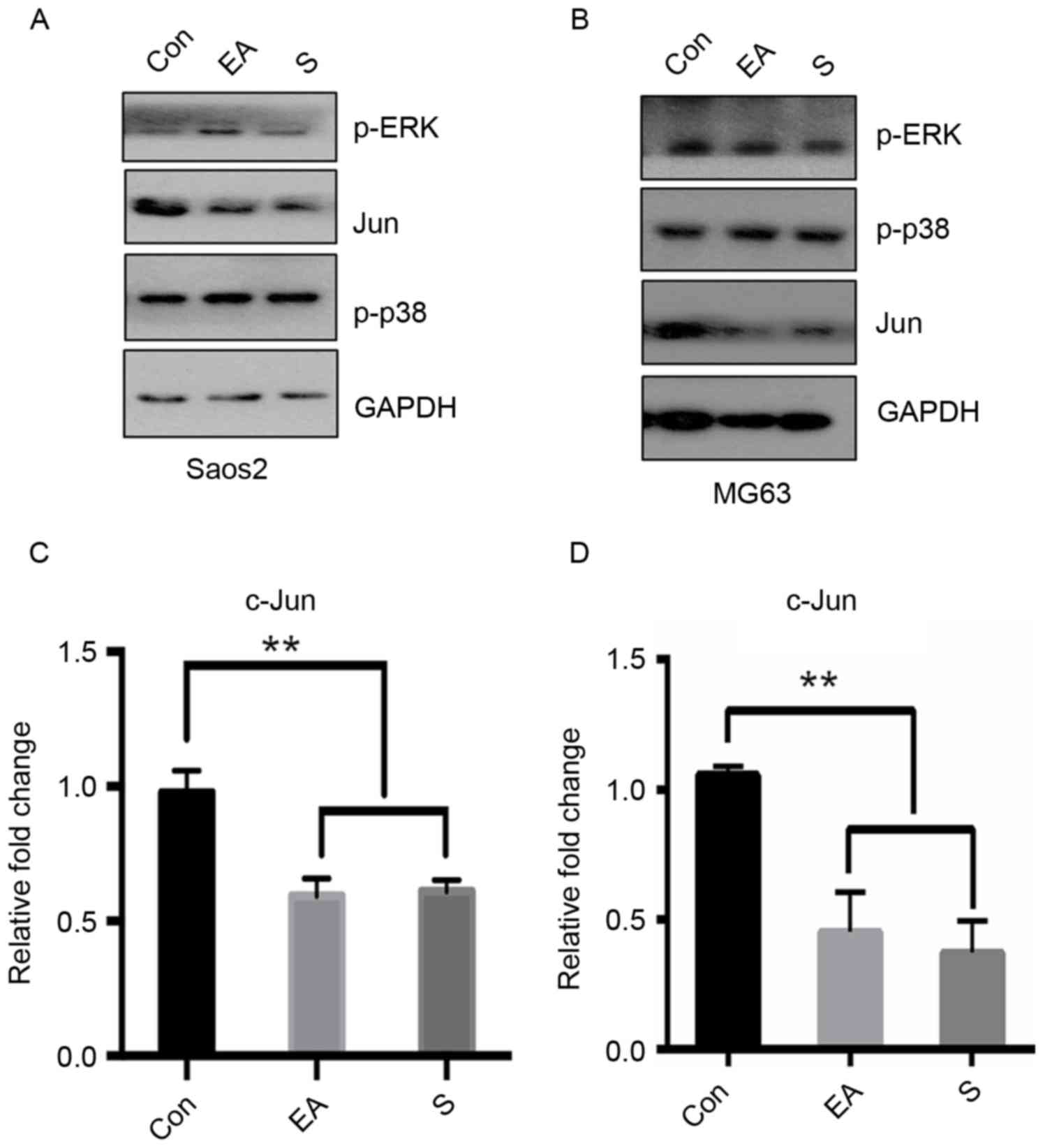
EA and Sennoside B repress c-Jun
expression
The accumulation of cells in the G1 phase
indicated the inhibition of the signals required for cell cycle
progression. The expression of factors from key signaling pathways
associated with G1 cell cycle progression was thus
examined following exposure to EA or Sennoside B. After 24 h of
treatment, the c-Jun protein level was observed to decrease to a
greater extent than the other assessed proteins (Fig. 4A and B). In order to further confirm
the effect of EA or Sennoside B on c-Jun, mRNA was extracted for
RT-qPCR following the same treatment. These results suggested that
EA or Sennoside B treatment repressed the transcription of c-Jun
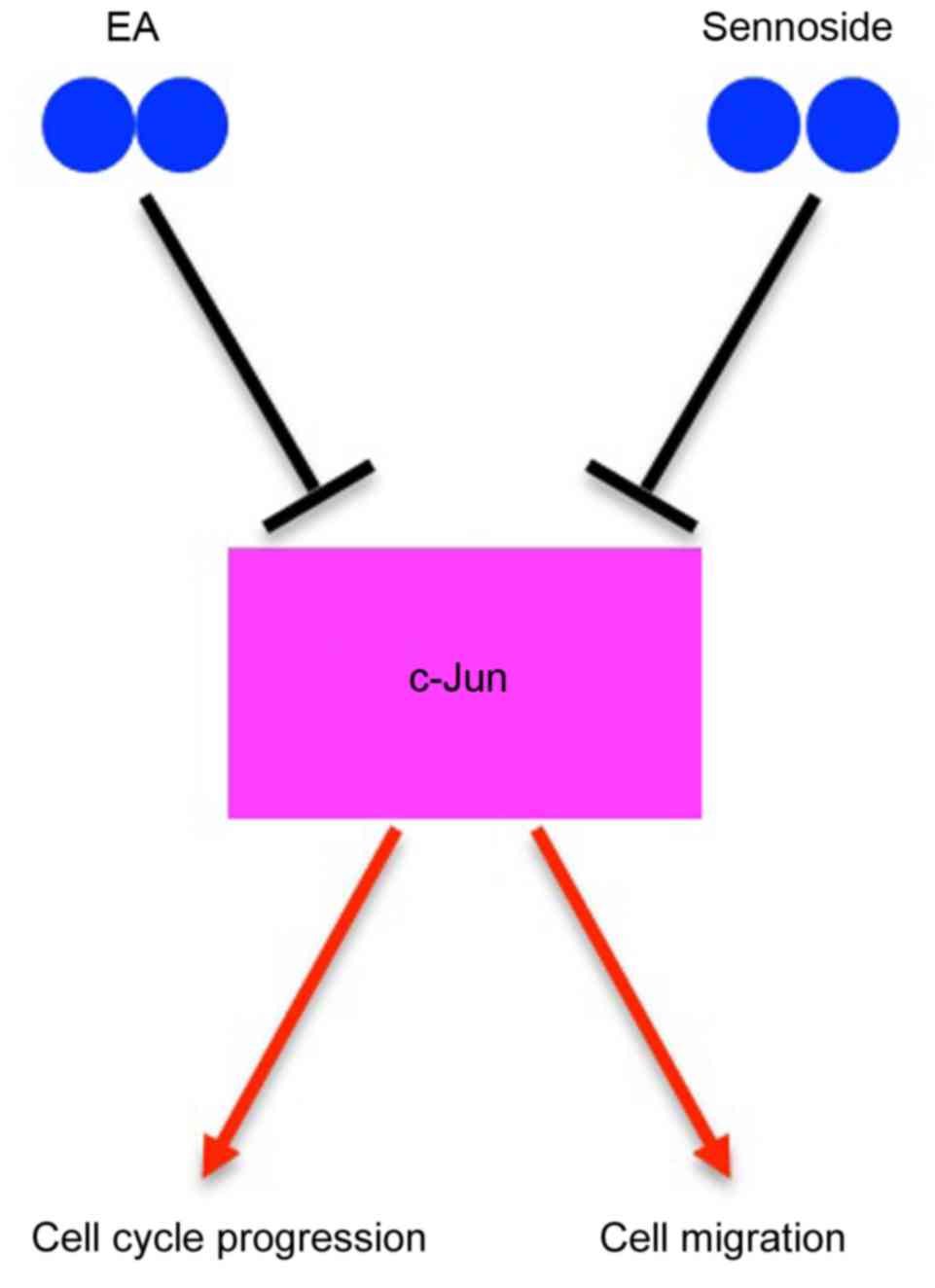
mRNA (Fig. 4C and D). A proposed
schematic was drawn to represent the potential mechanism for the EA
or Sennoside B inhibition of the growth, migration and invasion of
osteosarcoma cells (Fig. 5).
Discussion
In the present study, it was demonstrated that both
EA and Sennoside B treatment can inhibit cell migration, invasion
and proliferation. The inhibitory effect of EA or Sennoside B on
proliferation was attributed to the induction of cell cycle arrest.
The data regarding the effects of EA on osteosarcoma are consistent
with previous studies in other cancer types (7–11).
However, the inhibitory effects of Sennoside B were demonstrated
for the first time, to the best of our knowledge.
The high incidence of mortality in osteosarcoma is
associated with tumor metastasis (1).
Metastasis is a complex cascade, including various physiological
alterations to induce extracellular matrix (ECM) degradation
(20). Matrix metallopeptidases
(MMPs), particularly MMP-2 and MMP-9, have been reported as the
most important factors in the degradation of type IV collagen, a
major component of the basement membrane, which induces tumor cell
growth and metastasis (21). It has
been reported that EA inhibits either MMP2 activity or the
secretion of collagenases and gelatinases (22). AP-1 may serve a dominant role in the
transcriptional activation of the MMP promoters, and increasing
evidence has indicated the key role of c-Jun in the regulation of
MMPs (23). In the present study,
both EA and Sennoside B treatment repressed the transcription of
c-Jun, ultimately leading to a decrease in its protein level. This
mechanism partially explains why two different compounds inhibited
Saos-2 and MG63 cells in a similar manner.
Previous studies indicated that EA could inhibit
cancer cell growth by inducing apoptosis and cell cycle arrest
(8–11,19), but
Sennoside B's effect on cell growth was previously unreported. The
results of the present study suggested that both EA and Sennoside B
could inhibit cell proliferation without inducting apoptosis. The
oncogenic protein c-Jun regulates a range of cell cycle
progression-associated genes, including cyclin D1 (24). Cell cycle arrest in the present study
was likely to be caused by the repression of c-Jun expression.
However, other factors can induce the cell cycle arrest of
osteosarcoma cells. For example, the knockdown of ROR2 in
osteosarcoma cells inhibited cell proliferation and colony
formation by inducing cell cycle arrest (25). In addition, it was previously reported
that the exogenous expression of miR-497 in human osteosarcoma MG63
cells suppressed cell proliferation, colony formation, migration
and invasion, and induced apoptosis and arrest at the
G0/G1 phase of the cell cycle (26).
In conclusion, it was identified that EA and
Sennoside B may share a common mechanism to inhibit the growth,
migration and invasion of Saos-2 and MG63 cells, specifically, the
repression of c-Jun expression at the mRNA level. Further studies
are required to understand the detailed molecular mechanisms of
action of EA and Sennoside B on the regulation of the expression of
c-Jun, which may provide useful information for their possible
application in osteosarcoma prevention and therapy.
Acknowledgements
The present study was supported by the Shanghai
Youth Science and Technology Talent Sailing Program (grant no.
14YF1405900).
References
|
1
|
Wermers RA, Tiegs RD, Atkinson EJ,
Achenbach SJ and Melton LJ III: Morbidity and mortality associated
with Paget's disease of bone: A population-based study. J Bone
Miner Res. 23:819–825. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the surveillance, epidemiology and end results program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liao CL, Lai KC, Huang AC, Yang JS, Lin
JJ, Wu SH, Wood Gibson W, Lin JG and Chung JG: Gallic acid inhibits
migration and invasion in human osteosarcoma U-2 OS cells through
suppressing the matrix metalloproteinase-2/-9, protein kinase B
(PKB) and PKC signaling pathways. Food Chem Toxicol. 50:1734–1740.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hong WK and Sporn MB: Recent advances in
chemoprevention of cancer. Science. 278:1073–1077. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ishibashi K, Kumamoto K, Kuwabara K,
Hokama N, Ishiguro T, Ohsawa T, Okada N, Miyazaki T, Yokoyama M,
Tsuji Y, et al: Usefulness of sennoside as an agent for mechanical
bowel preparation prior to elective colon cancer surgery. Asian J
Surg. 35:81–87. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hussein RH and Khalifa FK: The protective
role of ellagitannins flavonoids pretreatment against
N-nitrosodiethylamine induced-hepatocellular carcinoma. Saudi J
Biol Sci. 21:589–596. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sohn EH, Koo HJ, Hang DTT, Jang SA,
Namkoong S, Lim JD and Kang SC: Protective effects of ellagic acid
on ethanol-induced toxicity in hepatic HepG2 cells. Mol Cell
Toxicol. 9:249–256. 2013. View Article : Google Scholar
|
|
8
|
Li TM, Chen GW, Su CC, Lin JG, Yeh CC,
Cheng KC and Chung JG: Ellagic acid induced p53/p21 expression, G1
arrest and apoptosis in human bladder cancer T24 cells. Anticancer
Res. 25:971–979. 2005.PubMed/NCBI
|
|
9
|
Mertens-Talcott SU and Percival SS:
Ellagic acid and quercetin interact synergistically with
resveratrol in the induction of apoptosis and cause transient cell
cycle arrest in human leukemia cells. Cancer Lett. 218:141–151.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Narayanan BA, Geoffroy O, Willingham MC,
Re GG and Nixon DW: p53/p21(WAF1/CIP1) expression and its possible
role in G1 arrest and apoptosis in ellagic acid treated cancer
cells. Cancer Lett. 136:215–221. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Päivärinta E, Pajari AM, Törrönen R and
Mutanen M: Ellagic acid and natural sources of ellagitannins as
possible chemopreventive agents against intestinal tumorigenesis in
the Min mouse. Nutr Cancer. 54:79–83. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Whitmarsh AJ and Davis RJ: Transcription
factor AP-1 regulation by mitogen-activated protein kinase signal
transduction pathways. J Mol Med (Berl). 74:589–607. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Papachristou DJ, Batistatou A, Sykiotis
GP, Varakis I and Papavassiliou AG: Activation of the JNK-AP-1
signal transduction pathway is associated with pathogenesis and
progression of human osteosarcomas. Bone. 32:364–371. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Angel P and Karin M: The role of Jun, Fos
and the AP-1 complex in cell-proliferation and transformation.
Biochim Biophys Acta. 1072:129–157. 1991.PubMed/NCBI
|
|
15
|
Hipskind RA and Bilbe G: MAP kinase
signaling cascades and gene expression in osteoblasts. Front
Biosci. 3:d804–d816. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu B and Shuai K: Induction of apoptosis
by protein inhibitor of activated Stat1 through c-Jun NH2-terminal
kinase activation. J Biol Chem. 276:36624–36631. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Suzuki A, Guicheux J, Palmer G, Miura Y,
Oiso Y, Bonjour JP and Caverzasio J: Evidence for a role of p38 MAP
kinase in expression of alkaline phosphatase during osteoblastic
cell differentiation. Bone. 30:91–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Umesalma S, Nagendraprabhu P and
Sudhandiran G: Ellagic acid inhibits proliferation and induced
apoptosis via the Akt signaling pathway in HCT-15 colon
adenocarcinoma cells. Mol Cell Biochem. 399:303–313. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chambers AF and Matrisian LM: Changing
views of the role of matrix metalloproteinases in metastasis. J
Natl Cancer Inst. 89:1260–1270. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pitchakarn P, Chewonarin T, Ogawa K,
Suzuki S, Asamoto M, Takahashi S, Shirai T and Limtrakul P: Ellagic
acid inhibits migration and invasion by prostate cancer cell lines.
Asian Pac J Cancer Prev. 14:2859–2863. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chakraborti S, Mandal M, Das S, Mandal A
and Chakraborti T: Regulation of matrix metalloproteinases: An
overview. Mol Cell Biochem. 253:269–285. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bakiri L, Lallemand D, Bossy-Wetzel E and
Yaniv M: Cell cycle-dependent variations in c-Jun and JunB
phosphorylation: A role in the control of cyclin D1 expression.
EMBO J. 19:2056–2068. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang J, Shi Y, Li H, Tan D, Yang M and Wu
X: Knockdown of receptor tyrosine kinase-like orphan receptor 2
inhibits cell proliferation and colony formation in osteosarcoma
cells by inducing arrest in cell cycle progression. Oncol Lett.
10:3705–3711. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ge L, Zheng B, Li M, Niu L and Li Z:
MicroRNA-497 suppresses osteosarcoma tumor growth in vitro and in
vivo. Oncol Lett. 11:2207–2212. 2016. View Article : Google Scholar : PubMed/NCBI
|