Introduction
Hepatocellular carcinoma (HCC), a major subtype of
primary liver cancer, represents the sixth most prevalent
malignancy and the third most common cause of cancer-associated
mortalities globally (1,2). According to statistics, in 2015, 35,660
new HCC cases and 24,550 mortalities due to HCC were predicted in
the USA (3). HCC initiation and
progression involves a variety of risk factors, including chronic
infection with hepatitis C virus or hepatitis B virus (HBV),
alcohol or drug abuse and a high-fat or high-sugar diet (4). Among these risk factors, infection with
HBV is the most prevalent cause of the disease worldwide, and is
responsible for the increasing incidence of HCC (1,4).
Developments have been made in improving diagnosis and therapeutic
treatments for HCC, but due to difficulties in diagnosis, a high
frequency of recurrence and resistance to common chemotherapy and
radiotherapy agents, the prognosis for patients with HCC remains
poor (5–7). The 5-year survival rate is <5% for
patients with HCC and intra-hepatic or extra-hepatic metastasis
(8). Therefore, it is urgent to
elucidate the molecular mechanisms underlying HCC carcinogenesis
and progression, and to develop effective strategies for the
diagnosis, treatment and prognosis of HCC.
Previously, numerous studies have suggested that the
abnormal expression of microRNAs (miRNAs/miRs) is associated with
the carcinogenesis, progression, metastasis and recurrence of HCC
(9,10). miRNAs are an emerging class of
non-coding, endogenous, small RNA molecules, ~19–25 nucleotides in
length, that negatively regulate the protein production of target
mRNAs at the post-transcriptional level by interacting with the
3′untranslated regions (3′UTR) of target mRNAs (11,12). In
this manner, miRNAs were involved in various biological processes,
including differentiation, proliferation, angiogenesis, metabolism,
apoptosis, cell cycle and metastasis (13,14).
Accumulated evidence has suggested that miRNAs may act as a novel
group of oncogenes or tumor suppressors, and the deregulation of
miRNA and target mRNA expression levels may contribute to
carcinogenesis and cancer development in a substantial number of
human malignancies, including HCC (15–17).
Therefore, further investigation concerning the expression levels,
functions and molecular mechanisms underlying the effects of miRNAs
in HCC will provide insight into the carcinogenesis and progression
of HCC.
The aim of the present study was to investigate the
expression levels, functions and molecular mechanisms underlying
the effect of miR-205 in HCC. Initially, the expression levels of
miR-205 in HCC tissues and cell lines were evaluated. Secondly, the
effects of miR-205 on the proliferation and metastasis of HCC cells
were determined by MTT assay, migration and invasion assays.
Finally, vascular endothelial growth factor A (VEGFA) was
identified as a functional downstream target of miR-205 in HCC.
These results indicated that miR-205 may function as a tumor
suppressor in HCC by directly targeting VEGFA, suggesting a
potential targeted therapy for patients with HCC.
Materials and methods
Tissue specimens, cell culture and
cell transfection
The present study was approved by the Ethics
Committee of Guizhou Cancer Hospital (Guiyang, China). Written
informed consent was obtained from all patients with HCC prior to
enrollment in the present study. None of the patients had received
antitumor treatment prior to surgery. HCC tissues and matched
adjacent non-cancerous liver tissues were obtained from 32 patients
(19 male; 13 female; age range, 46–74 years; mean age, 62 years)
who underwent surgery at Guizhou Cancer Hospital between August
2013 and January 2015. All tissue specimens were immediately frozen
and stored in liquid nitrogen.
HepG2, HuH-7, SMMC-7721 and BEL-7402 human HCC cell
lines and L02 and HEK293T normal hepatic cell lines were purchased
from the American Type Culture Collection (Manassas, VA, USA). All
cell lines were maintained in Dulbecco's modified Eagle's medium
supplemented with 10% fetal bovine serum (FBS), 100 IU/ml
penicillin and 0.1 mg/ml streptomycin (all Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C with 5%
CO2.
An miR-205 mimic and a negative control (NC) were
purchased from Genepharm, Inc. (Sunnyvale, CA, USA); the miR-205
mimic sequence was 5′-UCCUUCAUUCCACCGGAGUCUG-3′ and the NC sequence
was 5′-UUCUCCGAACGUGUCACGUTT-3′. VEGFA small interfering (si)RNA
and NC siRNA were also purchased from Genepharm, Inc. The sequence
for VEGFA siRNA and NC siRNA were as follows: VEGFA siRNA sense,
5′-GGCAGAAUCAUCACGAAGUTT-3′ and antisense,
5′-ACUUCGUGAUGAUUCUGCCTT-3′; NC siRNA sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT−3′. miRNA and siRNA transfection was
performed using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from tissues (1 g) or cells
(2×106) was extracted using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. A TaqMan miRNA assay (Applied Biosystems; Thermo Fisher
Scientific, Inc.) was used for quantification of miR-205 expression
levels, according to the manufacturer's protocol. For
quantification of VEGFA mRNA expression levels, reverse
transcription was performed using the M-MLV Reverse Transcription
system (Promega Corporation, Madison, WI, USA). The temperature
protocol for reverse transcription was as follows: 95°C for 2 min;
20 cycles of 94°C for 1 min, 55°C for 1 min and 72°C for 2 min;
then 72°C for 5 min. qPCR was performed using the SYBR-Green Master
Mix (Takara Biotechnology Co., Ltd., Dalian, China). The
thermocycling conditions for qPCR were as follows: 95°C for 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. U6 and
β-actin were used as internal controls for miR-205 and VEGFA
expression levels, respectively. The primers were as follows:
miR-205, 5′-GCTCCTTCATTCCACCGG-3′ (forward) and
5′-CAGTGCAGGGTCCGAGGT-3′ (reverse); U6,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ (forward) and
5′-CGCTTCACGAATTTGCGTGTCAT-3′ (reverse); VEGFA,
5′-AACTTTCTGCTGTCTTGGGT-3′ (forward) and 5′-TCTCGATTGGATGGCAGTA-3′
(reverse); and β-actin, 5′-GGGACCTGACTGACTACCTC-3′ (forward) and
5′-TCATACTCCTGCTTGCTGAT-3′ (reverse). Each sample was analyzed in
triplicate. Relative expression levels were calculated using the
2−∆∆Cq method (18).
MTT assay
The cell proliferation rate was evaluated using the
MTT assay (5 mg/ml; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
HepG2 and HuH-7 cells were seeded into 96-well plates at a density
of 3,000 cells per well. Following overnight incubation, miRNA or
siRNA transfection was performed as aforementioned. MTT assays were
performed following transfection at room temperature for 24, 48, 72
and 96 h. In brief, 20 µl MTT solution was added to each well.
Following incubation with MTT solution at 37°C for an additional 4
h, 200 µl dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) was added
to dissolve the formazan at 37°C for 10 min. Finally, the
absorbance at 490 nm was detected using an ELISA reader (BioTek
Instruments, Inc., Winooski, VT, USA). Each sample was analyzed in
triplicate.
Cell migration and invasion
assays
Cell migration and invasion were evaluated using
Transwell chambers (Corning Incorporated, Corning, NY, USA) with 8
µm pore-size polycarbonate membranes. For the invasion assay,
Transwell chambers were pre-coated with Matrigel (BD Biosciences,
Franklin Lakes, NJ, USA). For the migration and invasion assays,
3×104 transfected HepG2 and HuH-7 cells suspended in 300
µl serum-free DMEM were added to the upper chamber. DMEM (500 µl)
supplemented with 20% FBS was added to the low chamber as a
chemoattractant. Following incubation at 37°C for 48 h, cells that
had not migrated or invaded to the bottom surface of the Transwell
chambers were carefully removed with cotton wool. The migrated and
invaded cells were fixed with 100% methanol at room temperature for
10 min, stained with 0.5% crystal violet for 10 min and imaged with
a light microscope (magnification, ×200).
Western blotting
Total protein was isolated from HepG2 and HuH-7
cells (2×106) using RIPA lysis buffer (Bioteke
Corporation, Beijing, China) supplemented with 0.1 mg/ml
phenylmethylsulfonyl fluoride, 1 mM sodium orthovanadate and 1
mg/ml aprotinin. Protein concentration was determined using a
bicinchoninic acid assay kit (Bioteke Instuments, Inc.). Total
protein (20 µg) was fractionated by 10% SDS-PAGE (Beyotime
Institute of Biotechnology, Haimen, China), followed by
transference to polyvinylidene (PVDF) membranes (Millipore,
Bedford, MA). Non-specific binding sites of PVDF membranes were
blocked using 10% non-fat milk in Tris-buffered saline with 0.1%
Tween (TBST) solution at room temperature for 1 h. Subsequently,
the membranes were probed with mouse anti-human VEGFA monoclonal
primary antibody (dilution, 1:1,000; cat. no., ab155944;) and mouse
anti-human GADPH monoclonal primary antibody (dilution, 1:1,000;
cat. no., ab9484; both Abcam, Cambridge, UK). Following incubation
overnight at 4°C, membranes were washed with TBST three times and
incubated with goat anti mouse IgG horseradish
peroxidase-conjugated secondary antibody (1:3,000 dilution; ab6789;
Abcam) for 2 h at room temperature. Following washing three times
with TBST, the membranes were visualized using enhanced
chemiluminescence solution (Pierce; Thermo Fisher Scientific,
Inc.).
Dual-luciferase reporter assay
The target genes of miR-205 were assessed using the
miRNA target prediction tool TargetScan (version 6.0; http://www.targetscan.org/vert_60/) (19). In order to explore whether VEGFA was a
direct target gene of miR-205, a dual-luciferase reporter assay was
performed. HEK293T cells were co-transfected with VEGFA-3′UTR
wild-type (Wt) or VEGFA-3′UTR mutant (Mut), miR-205 mimics or NC
using Lipofectamine 2000 according to the manufacturer's protocol.
Luciferase activities were evaluated 48 h following transfection
using a Dual-Luciferase Reporter System (Promega Corporation,
Madison, WI, USA). Renilla luciferase activity was detected
as an internal control for firefly luciferase activity. Each sample
was analyzed in triplicate.
Statistical analysis
Data were presented as the mean ± standard
deviation. Statistical analyses were performed using SPSS version
17.0 (SPSS Inc., Chicago, IL, USA). All statistical analyses were
two-tailed. Data were analyzed using Student's t-tests and one-way
analysis of variance, with Student-Newman-Keuls tests used to
compare two groups in analyses with multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-205 expression level was decreased
in HCC tissues and cell lines
In order to determine miR-205 expression levels and
its function in HCC, expression levels of miR-205 were detected in
HCC tissues and matched adjacent non-cancerous liver tissues using
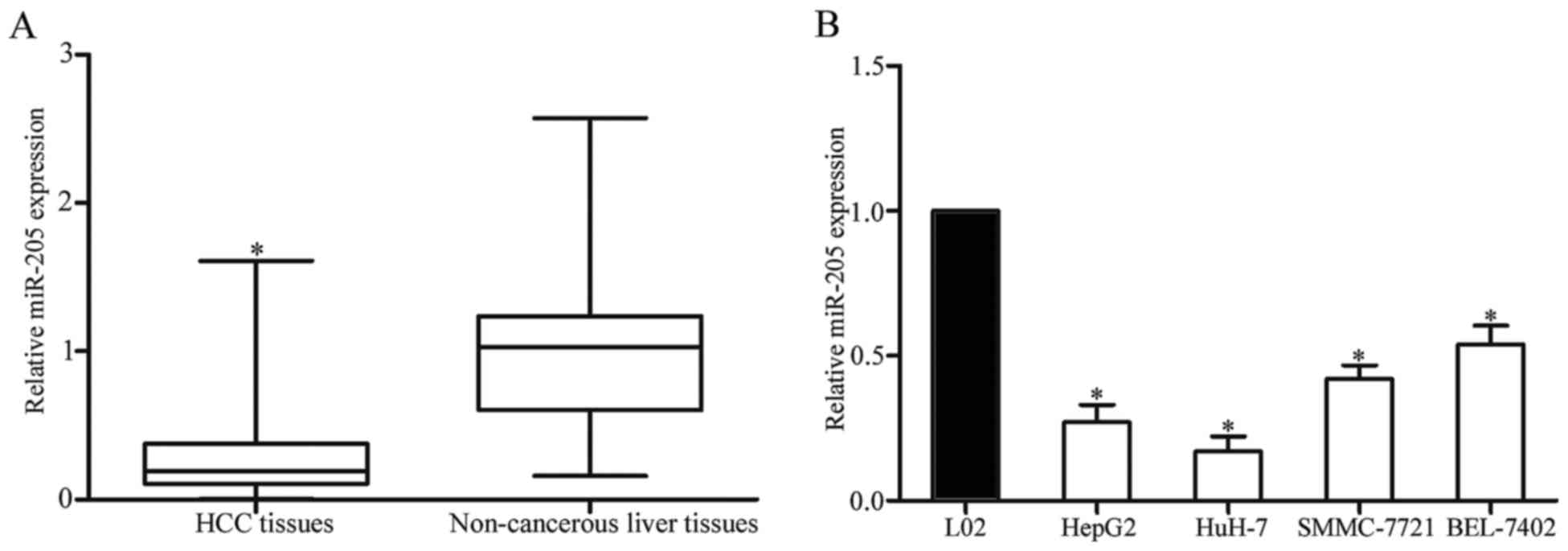
RT-qPCR. As presented in Fig. 1A,
miR-205 was significantly downregulated in HCC tissues compared
with in matched adjacent non-cancerous liver tissues. In addition,
miR-205 expression levels in HCC cell lines and a normal hepatic
cell line were also determined. As presented in Fig. 1B, the expression levels of miR-205
were decreased in the four HCC cell lines compared with the L02
normal hepatic cell line. These results indicated that the
downregulation of miR-205 may serve an important function in HCC
carcinogenesis and progression.
miR-205 inhibited the proliferation of
HCC cells
In order to investigate the functions of miR-205 in
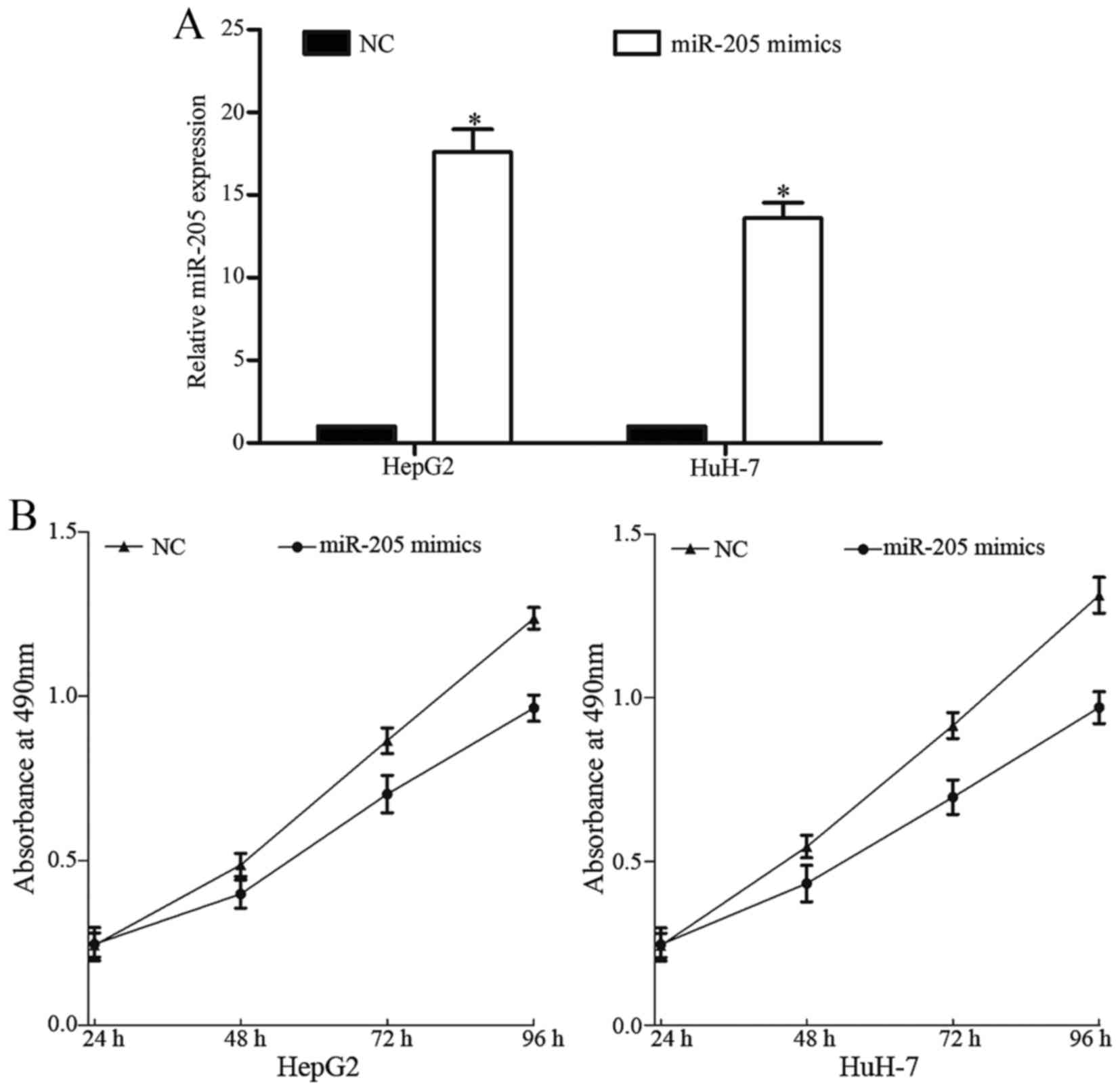
HCC, miR-205 mimics or NC were transfected into HepG2 and HuH-7
cells. As presented in Fig. 2A,
miR-205 was upregulated in HepG2 and HuH-7 cells following
transfection with miR-205 mimics compared with NC, suggesting that
HepG2 and HuH-7 cells were effective and adjustable models for the
functional studies of miR-205.
Cell proliferation was evaluated using the MTT
assay. miR-205 significantly reduced HepG2 and HuH-7 cell
proliferation (Fig. 2B). These
results indicated that overexpression of miR-205 inhibited the
proliferation of HepG2 and HuH-7 cells.
MiR-205 inhibited migration and
invasion of HCC cells
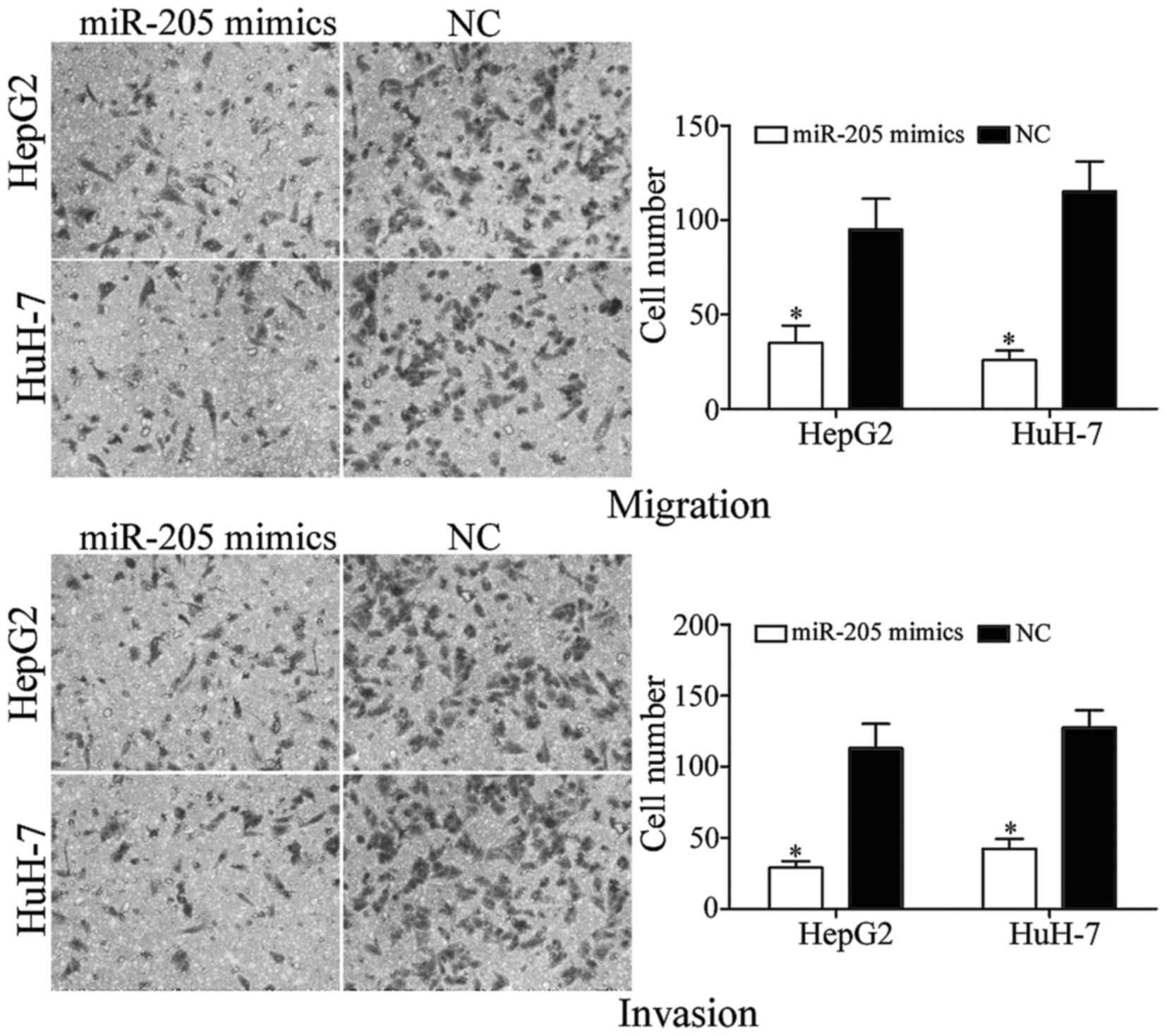
In order to investigate the function of miR-205 in
metastasis, Transwell migration and invasion assays were performed.
As presented in Fig. 3, upregulation
of miR-205 induced the suppression of tumor cell migration and
invasion in HepG2 and HuH-7 cells compared with NC controls. These
results indicated that miR-205 suppressed HCC cell metastasis in
vitro.
miR-205 directly targets the 3′UTR of
VEGFA
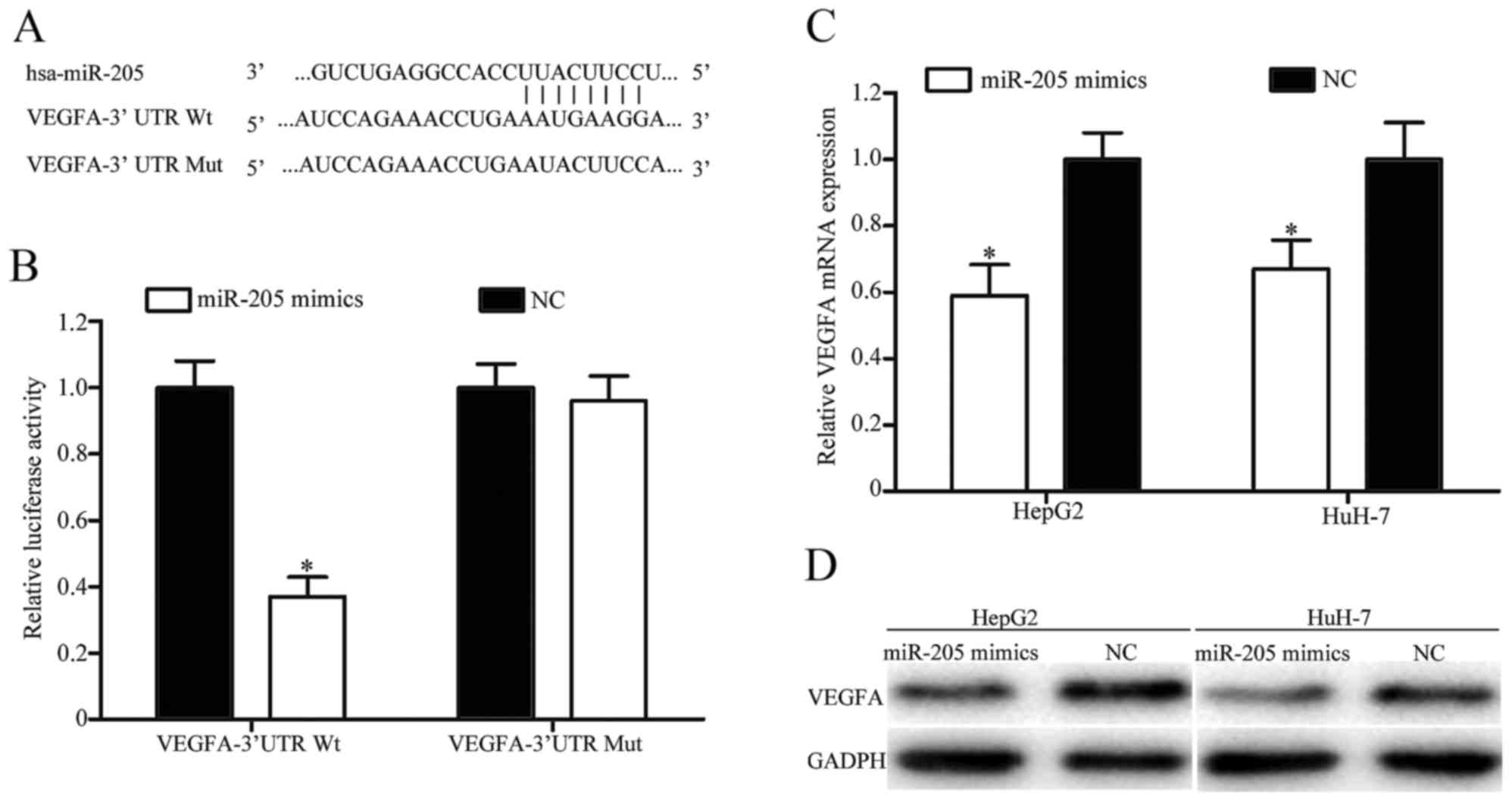
In order to explore the molecular mechanism
underlying the function of miR-205 in HCC, TargetScan was used to
predict the potential target genes of miR-205. As presented in
Fig. 4A, VEGFA was identified to
possess a putative miR-205 binding site in the 3′UTR. To verify
whether VEGFA was a direct target gene of miR-205, a
dual-luciferase reporter assay was performed. The results revealed
that overexpression of miR-205 decreased the luciferase activity of
VEGFA-3′UTR Wt, but did not decrease the luciferase activity of
VEGFA-3′UTR Mut (Fig. 4B).
Furthermore, RT-qPCR and western blotting was performed to
investigate the effect of miR-205 on the expression levels of VEGFA
mRNA and protein. As presented in Fig. 4C
and D, upregulation of miR-205 significantly inhibited the
expression level of VEGFA in HepG2 and HuH-7 cells at the mRNA and
protein levels, respectively. Taken together, these results
indicated that VEGFA was a direct target of miR-205 in HCC.
VEGFA is involved in miR-205-induced
suppression of HCC cell proliferation, migration and invasion
To explore whether VEGFA acted as a critical
mediator of miR-205 in HCC, VEGFA siRNA was used to knock down
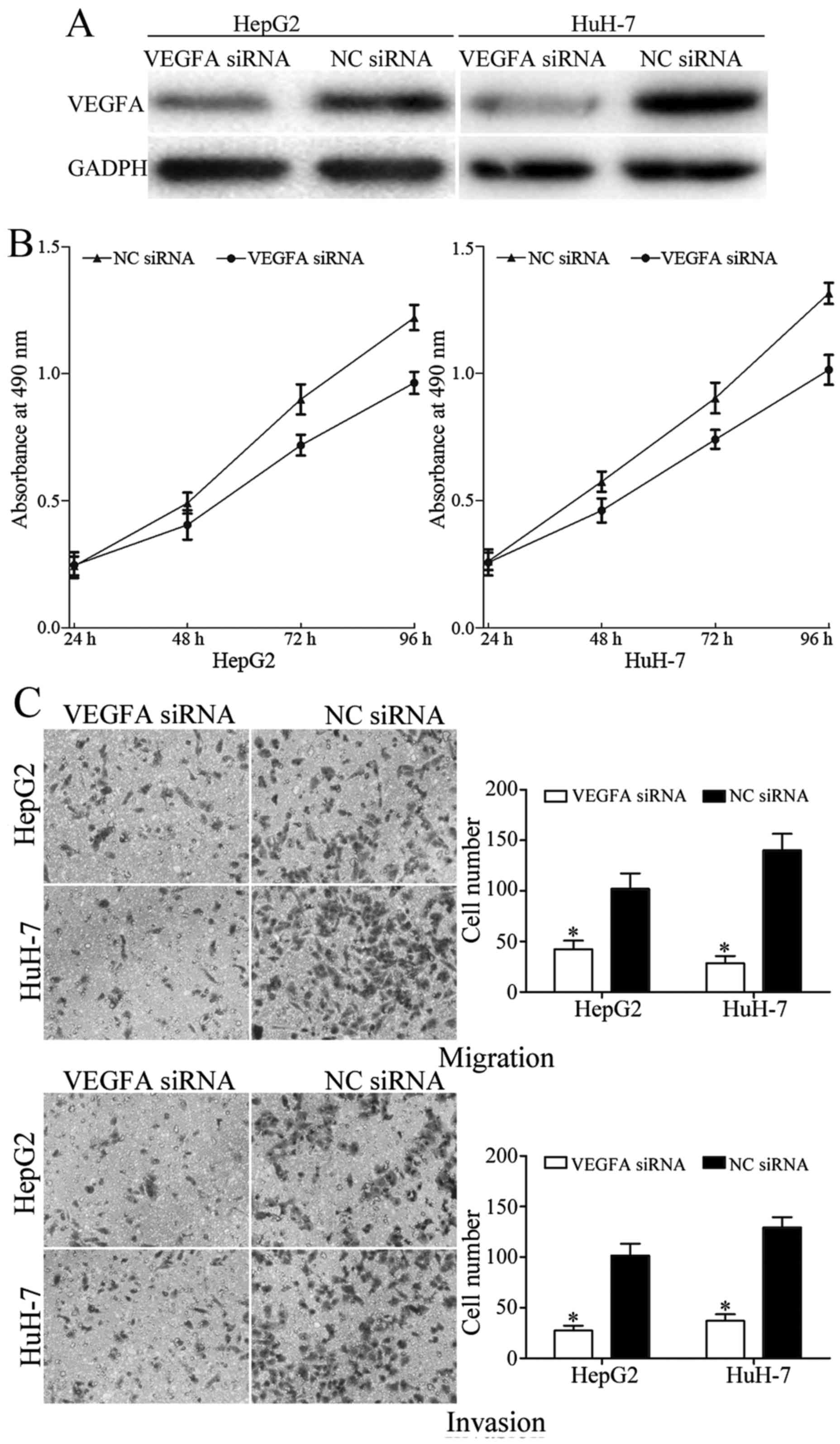
VEGFA expression. As presented in Fig.
5A, VEGFA was significantly downregulated in HepG2 and HuH-7
cells following transfection with VEGFA siRNA. Furthermore, MTT,
migration and invasion assays were performed to evaluated the
effect of VEGFA siRNA on the growth and metastasis of HCC. The
results demonstrated that knockdown of VEGFA suppressed the
proliferation, migration and invasion of HepG2 and HuH-7 cells
(Fig. 5B and C). Together, these
results suggested that miR-205 suppressed growth, migration and
invasion of HCC by downregulating VEGFA.
Discussion
It is now widely accepted that miRNAs modulate
various biological processes, including cancer initiation and
progression. In the present study, miR-205 expression levels in HCC
tissues and cell lines were determined, and the biological
functions and molecular mechanisms underlying miR-205 in HCC
carcinogenesis and progression were also investigated. It was
revealed that miR-205 was downregulated in HCC tissues and cell
lines. Overexpression of miR-205 significantly suppressed the
proliferation, migration and invasion of HCC cells by directly
targeting VEGFA. These findings suggested that miR-205 may be a
notable tumor suppressor in HCC.
Previous studies have demonstrated that miR-205
functions as an oncogene in numerous types of human cancer
(20–22). For example, in laryngeal squamous cell
carcinoma, miR-205 was upregulated and enhanced cell growth and
invasion via negative regulation of CDK2AP1 expression level
(20). Expression levels of miR-205
were reported to be increased in endometrial cancer tissues
(21). Kaplan-Meier survival analysis
revealed that high expression levels of miR-205 were significantly
associated with poor prognosis of patients with endometrial cancer
(21). In addition, miR-205 promoted
proliferation, metastasis and inhibited apoptosis of endometrial
cancer cells by targeting the protein kinase B signaling pathway,
phosphatase and tensin homolog (PTEN) and estrogen-related receptor
γ (23–25). In ovarian cancer, miR-205 was
upregulated and miR-205 expression levels were associated with high
pathological grade and advanced clinical stage of patients with
epithelial ovarian cancer. Ectopic miR-205 expression improved
migration and invasion abilities of ovarian cancer cells by
directly targeting zing-finger E-box binding homeobox 1 (22). Lei et al (24) and Bai et al (25) investigated miR-205 expression levels
in non-small cell lung cancer (NSCLC) tissues and cell lines.
Upregulation of miR-205 increased the proliferation, migration,
invasion and chemoresistance of NSCLC cells via regulation of the
PTEN signaling pathway (26,27). These results suggested that miR-205
may serve important functions in these types of cancer, and should
be investigated as a potential therapeutic target for possible
therapeutic strategies.
miR-205 has been reported to be a tumor suppressor
in various types of cancer, including osteosarcoma (28), thyroid cancer (29), breast cancer (30,31), renal
cell carcinoma (32), oral carcinoma
(33) and prostate cancer (34). In accordance with these results, the
present study indicated that miR-205 was downregulated in HCC
tissues and overexpression of miR-205 inhibited HCC cell
proliferation, migration and invasion in vitro. These
conflicting results concerning miR-205 expression levels and
functions demonstrate that miR-205 acted as an oncogene in certain
types of cancer and as a tumor suppressor in others. This
contradiction may be explained by the ‘imperfect complementarity’
of the interactions between miRNAs and target genes (35).
In order to understand the molecular mechanisms
underlying miR-205-induced suppression of cell proliferation,
migration and invasion in HCC, TargetScan and dual-luciferase
reporter assays were performed. VEGFA was verified to be a direct
target gene of miR-205 in HCC. Furthermore, RT-qPCR and western
blotting were preformed to investigate whether miR-205 regulated
VEGFA mRNA and protein expression levels. The results revealed that
miR-205 significantly decreased VEGFA expression at the mRNA and
protein levels. Taken together, miR-205 negatively regulated VEGFA
expression by directly binding to the 3′UTR of VEGFA in HCC.
Identification of the target gene of miR-205 is essential for
understanding its functions in HCC carcinogenesis and progression.
It is also important for developing novel therapeutic targets for
HCC.
VEGFA, a 35–45 kD heparin-binding glycoprotein, is a
key regulator of angiogenesis which is known to be a fundamental
factor in the local growth of tumors and progression to metastasis
(28). High expression levels of
VEGFA have been reported in various types of human cancers,
including HCC (36,37). VEGFA serves an important function in
tumor proliferation, migration, invasion and angiogenesis (38–40).
Therefore, anti-VEGFA targeted therapy, including bevacizumab, has
been widely used to treat cancers in a clinical setting (41). Regarding its cancer-associated
functions in HCC, VEGFA is worth paying attention to as a potential
target to treat patients with HCC. The present study revealed that
miR-205 targeted VEGFA to suppress cell proliferation and
metastasis in HCC. Collectively, miR-205/VEGFA based targeted
therapy may be a novel therapeutic treatment for HCC.
In conclusion, the present study demonstrated that
miR-205 was significantly downregulated in HCC tissues and cell
lines, and overexpression of miR-205 inhibited the proliferation,
migration and invasion of HCC cells by directly targeting VEGFA.
These results help to further the understanding of the molecular
mechanisms underlying HCC carcinogenesis and progression, and may
potentially lead to novel targeted therapies for HCC in future
studies.
References
|
1
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
El-Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu Y, Cain-Hom C, Choy L, Hagenbeek TJ, de
Leon GP, Chen Y, Finkle D, Venook R, Wu X, Ridgway J, et al:
Therapeutic antibody targeting of individual Notch receptors.
Nature. 464:1052–1057. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang X, Qin J and Lu S: Up-regulation of
miR-877 induced by paclitaxel inhibits hepatocellular carcinoma
cell proliferation though targeting FOXM1. Int J Clin Exp Pathol.
8:1515–1524. 2015.PubMed/NCBI
|
|
7
|
Yang LY, Fang F, Ou DP, Wu W, Zeng ZJ and
Wu F: Solitary large hepatocellular carcinoma: A specific subtype
of hepatocellular carcinoma with good outcome after hepatic
resection. Ann Surg. 249:118–123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou YM, Zhang XF, Yu F, Liu XB, Wu LP, Li
B and Yang JM: Efficacy of surgical resection for pulmonary
metastases from hepatocellular carcinoma. Med Sci Monit.
20:1544–1549. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen X, Bo L, Zhao X and Chen Q:
MicroRNA-133a inhibits cell proliferation, colony formation
ability, migration and invasion by targeting matrix
metallopeptidase 9 in hepatocellular carcinoma. Mol Med Rep.
11:3900–3907. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang ZQ, Meng H, Wang N, Liang LN, Liu
LN, Lu SM and Luan Y: Serum microRNA 143 and microRNA 215 as
potential biomarkers for the diagnosis of chronic hepatitis and
hepatocellular carcinoma. Diagn Pathol. 9:1352014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu J: Control of protein synthesis and
mRNA degradation by microRNAs. Curr Opin Cell Biol. 20:214–221.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yates LA, Norbury CJ and Gilbert RJ: The
long and short of microRNA. Cell. 153:516–519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer. 96
Suppl:R40–R44. 2007.PubMed/NCBI
|
|
15
|
Li B, Liu L, Li X and Wu L: miR-503
suppresses metastasis of hepatocellular carcinoma cell by targeting
PRMT1. Biochem Biophys Res Commun. 464:982–987. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Fang L, Yu W and Wang Y:
MicroRNA-125b suppresses the migration and invasion of
hepatocellular carcinoma cells by targeting transcriptional
coactivator with PDZ-binding motif. Oncol Lett. 9:1971–1975. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zheng C, Li J, Wang Q, Liu W, Zhou J, Liu
R, Zeng Q, Peng X, Huang C, Cao P and Cao K: MicroRNA-195 functions
as a tumor suppressor by inhibiting CBX4 in hepatocellular
carcinoma. Oncol Rep. 33:1115–1122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhong G and Xiong X: miR-205 promotes
proliferation and invasion of laryngeal squamous cell carcinoma by
suppressing CDK2AP1 expression. Biol Res. 48:602015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Karaayvaz M, Zhang C, Liang S, Shroyer KR
and Ju J: Prognostic significance of miR-205 in endometrial cancer.
PLoS One. 7:e351582012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Niu K, Shen W, Zhang Y, Zhao Y and Lu Y:
MiR-205 promotes motility of ovarian cancer cells via targeting
ZEB1. Gene. 574:330–336. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jin C and Liang R: miR-205 promotes
epithelial-mesenchymal transition by targeting AKT signaling in
endometrial cancer cells. J Obstet Gynaecol Res. 41:1653–1660.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Su N, Qiu H, Chen Y, Yang T, Yan Q and Wan
X: miR-205 promotes tumor proliferation and invasion through
targeting ESRRG in endometrial carcinoma. Oncol Rep. 29:2297–2302.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang G, Hou X, Li Y and Zhao M: MiR-205
inhibits cell apoptosis by targeting phosphatase and tensin homolog
deleted on chromosome ten in endometrial cancer Ishikawa cells. BMC
Cancer. 14:4402014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lei L, Huang Y and Gong W: miR-205
promotes the growth, metastasis and chemoresistance of NSCLC cells
by targeting PTEN. Oncol Rep. 30:2897–2902. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bai J, Zhu X, Ma J and Wang W: miR-205
regulates A549 cells proliferation by targeting PTEN. Int J Clin
Exp Pathol. 8:1175–1183. 2015.PubMed/NCBI
|
|
28
|
Wang L, Shan M, Liu Y, Yang F, Qi H, Zhou
L, Qiu L and Li Y: miR-205 suppresses the proliferative and
migratory capacity of human osteosarcoma Mg-63 cells by targeting
VEGFA. Onco Targets Ther. 8:2635–2642. 2015.PubMed/NCBI
|
|
29
|
Salajegheh A, Vosgha H, Rahman Md A, Amin
M, Smith RA and Lam AK: Modulatory role of miR-205 in angiogenesis
and progression of thyroid cancer. J Mol Endocrinol. 55:183–196.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang H and Fan Q: MicroRNA-205 inhibits
the proliferation and invasion of breast cancer by regulating AMOT
expression. Oncol Rep. 34:2163–2170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang H, Li B, Zhao H and Chang J: The
expression and clinical significance of serum miR-205 for breast
cancer and its role in detection of human cancers. Int J Clin Exp
Med. 8:3034–3043. 2015.PubMed/NCBI
|
|
32
|
Chen Z, Tang ZY, He Y, Liu LF, Li DJ and
Chen X: miRNA-205 is a candidate tumor suppressor that targets ZEB2
in renal cell carcinoma. Oncol Res Treat. 37:658–664. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim JS, Park SY, Lee SA, Park MG, Yu SK,
Lee MH, Park MR, Kim SG, Oh JS, Lee SY, et al: MicroRNA-205
suppresses the oral carcinoma oncogenic activity via
down-regulation of Axin-2 in KB human oral cancer cell. Mol Cell
Biochem. 387:71–79. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang N, Li Q, Feng NH, Cheng G, Guan ZL,
Wang Y, Qin C, Yin CJ and Hua LX: miR-205 is frequently
downregulated in prostate cancer and acts as a tumor suppressor by
inhibiting tumor growth. Asian J Androl. 15:735–741. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu Z, Ni L, Chen D, Zhang Q, Su Z, Wang Y,
Yu W, Wu X, Ye J, Yang S, et al: Identification of miR-7 as an
oncogene in renal cell carcinoma. J Mol Histol. 44:669–677. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yamaguchi R, Yano H, Iemura A, Ogasawara
S, Haramaki M and Kojiro M: Expression of vascular endothelial
growth factor in human hepatocellular carcinoma. Hepatology.
28:68–77. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Miura H, Miyazaki T, Kuroda M, Oka T,
Machinami R, Kodama T, Shibuya M, Makuuchi M, Yazaki Y and Ohnishi
S: Increased expression of vascular endothelial growth factor in
human hepatocellular carcinoma. J Hepatol. 27:854–861. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhuang Y and Wei M: Impact of vascular
endothelial growth factor expression on overall survival in
patients with osteosarcoma: A meta-analysis. Tumour Biol.
35:1745–1749. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu Y, Zheng Q, Wu H, Guo X, Li J and Hao
S: Rapamycin increases pCREB, Bcl-2, and VEGF-A through ERK under
normoxia. Acta Biochim Biophys Sin (Shanghai). 45:259–267. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wiszniak S, Mackenzie FE, Anderson P,
Kabbara S, Ruhrberg C and Schwarz Q: Neural crest cell-derived VEGF
promotes embryonic jaw extension. Proc Natl Acad Sci USA.
112:6086–6091. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hicklin DJ and Ellis LM: Role of the
vascular endothelial growth factor pathway in tumor growth and
angiogenesis. J Clin Oncol. 23:1011–1027. 2005. View Article : Google Scholar : PubMed/NCBI
|