Introduction
The core of epithelial-mesenchymal transition (EMT),
is to lose the epithelium-derived labelled protein E-cadherin and
acquire the labeled protein of the mesenchymal cells (such as
vimentin, N-cadherin) (1). E-cadherin
is one of the important components for adherent junction of the
epithelial cells. It is bound to the intracellular proteins to form
morphology, polarity, and functions of the epithelial cells thus
allowing for tight junctions between epithelial cells and
maintaining the integrity of the structure of the epithelial tissue
(2). In addition to the decreased
expression of the labeled E-cadherin, the EMT can also lead to
changes in other epithelium-derived characteristic phenotypes,
including tight junctions, gap junctions between epithelial cells,
and desmosome (3). Accordingly,
interstitial cytoskeletal reorganization occurs in the cells with
EMT.
Currently, a great deal of research shows that
several transcription factor families can directly act on the
E-cadherin promoter and directly inhibit its transcription. These
transcription factors include: zinc finger protein family Zeb (Zeb1
and Zeb2) (4), Snail family (Snail1,
Snail2, and Snail3) (5), Twist family
(Twist1 and Twist2) (6), fork-head
(7) transcription factor family FOXC2
(8) and other transcription factors
under investigation. The Zeb family is a generally-accepted EMT
activated transcription factor including Zeb1 and Zeb2. Zeb1 can
directly inhibit the expression of E-cadherin (9) and initiate the EMT process by means of
combination between zinc finger and E-box in the E-cadherin genetic
promoter region. Moreover, in addition to E-box, research showed
that the action of Zeb1 on E-cadherin may also have other
mechanisms. The compound formed by Zeb1 and the histone deacetylase
may participate in the transcription splicing of E-cadherin thus
inhibiting its genetic expression (10). Downregulation of the expression of
Zeb1 in mesenchymal tumor cells may lead to mesenchymal-epithelial
transition in tumor cells and epithelial cell phenotype in cells
(11). Research shows that Zeb1 can
significantly increase the invasiveness and metastasis rate of
tumors by inducing the EMT process of the tumor cells (12).
MicroRNA is a class of short non-coding RNA
comprising only 20–22 nucleotides. They silence the target gene or
inhibit their translation by specifically binding and cutting the
messenger RNA (13). A great deal of
research shows that the miR-23a family plays a role in inhibiting
the EMT of tumor cells thus leading to opposite
mesenchymal-epithelialization transition and promoting the
expression of E-cadherin (14).
Transfecting the mesenchymal pancreatic carcinoma and breast
carcinoma with the miR-23a precursor allows for transition from
mesenchyme to epithelium, increased expression of E-cadherin,
decreased expression of vimentin, and occurrence of epithelial
phenotype in cells (15). In
addition, research on the ovarian carcinoma cells indicates that
the overexpression of miR-23a can significantly inhibit the
migration ability of the cancer cells and strengthen their
sensitivity to the chemotherapeutic drugs. On the contrary,
overexpression of miR-23a can significantly decrease the
tumorigenic ability of pancreatic carcinoma cells (16).
miR-23a and Zeb1 have completely contrary functions
in regulation of EMT. Moreover, they also have diametrically
opposite functions in regulation of such characteristics as
stemness maintenance in the tumor stem cells, and drug resistance,
related to EMT. The former decreases the tumorigenic ability of the
tumor cells and increases their sensitivity to chemotherapy. The
latter increases the cellular proliferation ability and improves
drug resistance. Furthermore, direct mutual inhibition effect also
occurs. Therefore, Zeb1 and miR-23a form a bidirectional inhibitory
negative feedback loop. The roles of Zeb1-miR-23a in regulating EMT
of tumor cells, tumor deterioration, metastasis, drug resistance,
and recurrence are arousing wide attention to some human malignant
tumors (such as pancreatic, bladder, breast, and thyroid carcinoma)
(17). However, there is still lack
of research on ocular malignancies. Choroidal melanoma and
retinoblastoma are common intraocular tumors with a high morbidity,
a malignant grade, and a high susceptibility to metastasis.
Particularly, hematogenous metastasis has a high case fatality rate
as it occurs in the remote organ in an early stage of choroidal
melanoma. A probe into the mechanism of metastasis of these
malignant tumors makes it possible to identify new strategies to
better control and treat these tumors and decrease their case
fatality rates.
To further test the feedback regulation effect of
Zeb1 on miR-23a, we establish the Zeb1 overexpression lentiviral
vector to achieve stable transfection of the human choroidal
melanoma cell strain OCM-1. Then, we utilized the expression of the
puromycin resistance gene carried by the Zeb1 overexpression
lentiviral vector and used the puromycin to screen for the stable
transfected strains from the OCM-1 cells transfected with
lentivirus to obtain the stable strain cells with overexpression of
Zeb1 thus laying a foundation for subsequent establishment of the
animal models and the in vivo experiments of tumorigenicity
and metastasis ability of tumor cells. Based on the real
time-quantitative PCR (RT-qPCR) detection and western blot
analysis, we quantitatively detected the cells in the control and
Zeb1 overexpression group for Zeb1 and miR-23a and the role of
overexpression of Zeb1 in the cell strain in regulating miR-23a. We
also further detected the related factors and stemness factors of
the EMT to evaluate the role of Zeb1 in regulating EMT and stemness
tumorigenicity.
Materials and methods
Transfection of liposome-mediated
small RNA mimics and inhibitors
At 1 day before transfection, 4.5×104
cells were inoculated into a 6-well microplate and cultured in a 5%
CO2 incubator at 37°C after addition of 2 ml of basal
culture medium containing fetal bovine serum (Invitrogen: Thermo
Fisher Scientific, Inc., Carlsbad, CA, USA). The mixture of miR-23a
mimics or inhibitor and Lipofectamine™ 2000 (all from Invitrogen:
Thermo Fisher Scientific, Inc.) was added to each well containing
cells and culture medium. The culture plate was shaken gently back
and forth to mix well the solution and the cell culture medium. The
cells were placed in a CO2 incubator at 37°C. The
transfection efficiency was determined with fluorescence at 6 h
(Takara Bio, Inc., Tokyo, Japan). It was replaced by the standard
serum-containing culture medium at 12 h. The cells were collected
for further detection after they were incubated for 48 h in the 5%
CO2 incubator at 37°C. The study was approved by the
Ethics Committee of the Affiliated Hospital of Inner Mongolia
Medical University (Hohhot, Inner Mongolia, China).
RT-qPCR
A total of 106 intraocular tumor cells
were directly added to 1 ml TRIzol, mixed well on a vortex mixer,
and allowed to stand for 5 min at room temperature. After rotation,
the supernatant was transferred to a new tube and the same volume
of isopropanol was added (Beijing Chemical Reagents Co. Ltd.,
Beijing, China), the solution was mixed well for 1 min by turning
upside down, and allowed to stand for 5 min at room temperature and
centrifuged for 15 min at 8,000 × g at 4°C. After addition of
isopropanol of the same volume (Beijing Chemical Reagents Co.,
Ltd.), the cells were mixed well by gently turning upside down, and
allowed to stand for 10 min at room temperature and centrifuged for
10 min at 8,000 × g at 4°C. The supernatant was discarded and 1 ml
of 75% ethanol was added. An appropriate volume of DEPC water was
added (EMD Millipore, Billerica, MA, USA) to sufficiently dissolve
the sediment. The reaction system (Takara Bio, Inc.) was 25 µl:
fluorescence RT-qPCR reaction solution 20 µl, DNA polymerase 1 µl,
reverse transcriptase 0.35 µl, template RNA 5 µl, mixed well and
centrifuged for 10 sec at 3,000 × g. RT-qPCR amplification
procedures: reversely transcribed for 30 min at 50°C; pre-denatured
for 3 min at 95°C; denatured at 95°C; 15 sec, annealed for 30 sec
at 50°C, extended for 30 min at 72°C, 5 cycles in total; denatured
for 10 sec at 95°C, annealed for 40 sec at 55°C, 40 cycles in
total. The primer sequence was obtained from previous studies
(18,19).
Western blot analysis of protein
expression
OCM-1, WERI-RB1, and Y79 cells in the logarithmic
growth phase were diluted and inoculated into a 6-well microplate
for culture. The cells in the control and treatment group were
collected, respectively, and subjected to RIPA lysis (Promega
Corp., Madison, WI, USA), protein quantification, and western blot
analysis. The cells were first subjected to SDS-polyacrylamide gel
electrophoresis. The protein specimen was transferred to the
nitrocellulose (NC) membrane (Polysciences, Inc., Warrington, PA,
USA), and blocked. The Zeb1 antibody (1:1,000), E-cadherin (both
from Cell Signaling Technology, Inc., Danvers, MA, USA),
N-cadherin, vimentin (both from Abcam, Cambridge, UK) antibodies
were incubated (dilution rates, 1:1,000). The internal reference
was β-actin (1:2,000; Abcam). The cells were incubated for 2 h at
room temperature and incubated with the secondary antibody for 1 h.
The chemiluminescence method was used.
Immumofluorescence method
The OCM-1, WERI-RB1, and Y79 cells were rinsed 3
times for 5 min each with PBS after growing on the glass slide. The
cells were blocked for 20 min at 37°C after the goat serum working
solution was added dropwise. The cells were allowed to stand
overnight at 4°C and rinsed 3 times for 5 min each with PBS next
day after the goat anti-Zeb1 (1:500): rabbit anti-E-cadherin
(1:500) (both from Cell Signaling Technology, Inc.), mouse
anti-vimentin (1:500; Abcam) were added dropwise. The cells were
incubated for 2 h at 37°C after the specific secondary antibody
Alexa Fluor 594 (1:1,000) and Alexa Fluor 488 (1:1,000) (both from
Invitrogen: Thermo Fisher Scientific, Inc.) were added dropwise.
The cells were rinsed 3 times for 5 min each. The nuclei were
stained with DAPI. The cells were air dried, mounted with
anti-fluorescence quenching mounting solution, and preserved in the
dark at 4°C for microscopic examination.
Cell migration experiment
(Transwell)
Trypsin was added to the culture flask to digest the
cells. After the digestion was terminated, the cell suspension was
pipetted, added to a centrifugal tube, and centrifuged for 5 min at
3,000 × g. The cells were washed once or twice with PBS and
resuspended with the serum-free medium containing BSA until the
cell density reached 1×104/ml after the culture solution
was discarded. The cells and the culture medium were inoculated
into a Transwell 6-well microplate (Corning, Inc., Corning, NY,
USA). A total of 1 ml of the culture medium containing FBS was
added to the lower chamber. Then, 2 ml of the serum-free tumor cell
suspension prepared in advance was added to the upper chamber. The
cells were routinely cultured for 24 h. The culture medium in the
upper chamber was discarded. The cells on the upper chamber side of
the semipermeable membrane at the bottom were removed with a cotton
swab. The cells were fixed for 15 min with 95% ethanol, stained for
10 min with hematoxylin (Takara Biotechnology Co., Ltd., Dalian,
China), and observed and photographed under an inverted microscope
using four fields of view (×200). The cells passed through the
semipermeable membrane and entered the lower chamber. The cell
migration ability was detected.
Lentivirus transfection
One day before transfection, the cell suspension
with a density of 5×104 was inoculated into 3 wells of
the 6-well microplate and allowed to stand overnight in an
incubator. The old culture medium was discarded the following day.
The cells were washed once with the D-Hank's solution. Then, 2 ml
of the freshly-prepared 7% non-resistant culture medium Opti-MEM I
(Invitrogen: Thermo Fisher Scientific, Inc.) was added. The first
well served as the normal group and the second well served as the
experimental group. A total of 30 µl Zeb1 virus solution and 2 µl
transfection solution polybrene (Takara Bio, Inc.) were added in
the experimental group. The solution was mixed well and placed in
the incubator. The cellular growth was observed every other day and
the solution was replaced every third day. The OCM-1 cells were
collected and transferred to a 24-well microplate with a density of
5×104 cells after being transfected for 72 h with
lentivirus. The common DMEM culture medium containing serum was
added. The cells were incubated overnight. The screening culture
medium was prepared. The fresh culture medium contained 1.25 µg/ml
puromycin (Biosino Bio-Technology and Science Inc., Beijing,
China). The prepared puromycin screening culture medium was used to
replace the old culture medium. The old culture medium was replaced
with the freshly-prepared screening culture medium at ~12–24 h
after cell death. Apoptotic cells were removed. To minimize the
changes in cellular phenotype arising from puromycin, the puromycin
culture medium was removed and replaced with the normal culture
medium at 72 h. The survived cells were considered as stably
transfected cells.
Statistical analysis
The statistical software SPSS 19.0 (IBM Corp.,
Armonk, NY, USA) was used for statistical analysis. Excel was used
for statistical charting. ANOVA was used for comparison of multiple
groups and the post hoc test was LSD. The differences were
considered statistically significant at P<0.05.
Results
Role of miR-23a transfection in
regulating the expression of the target gene Zeb1
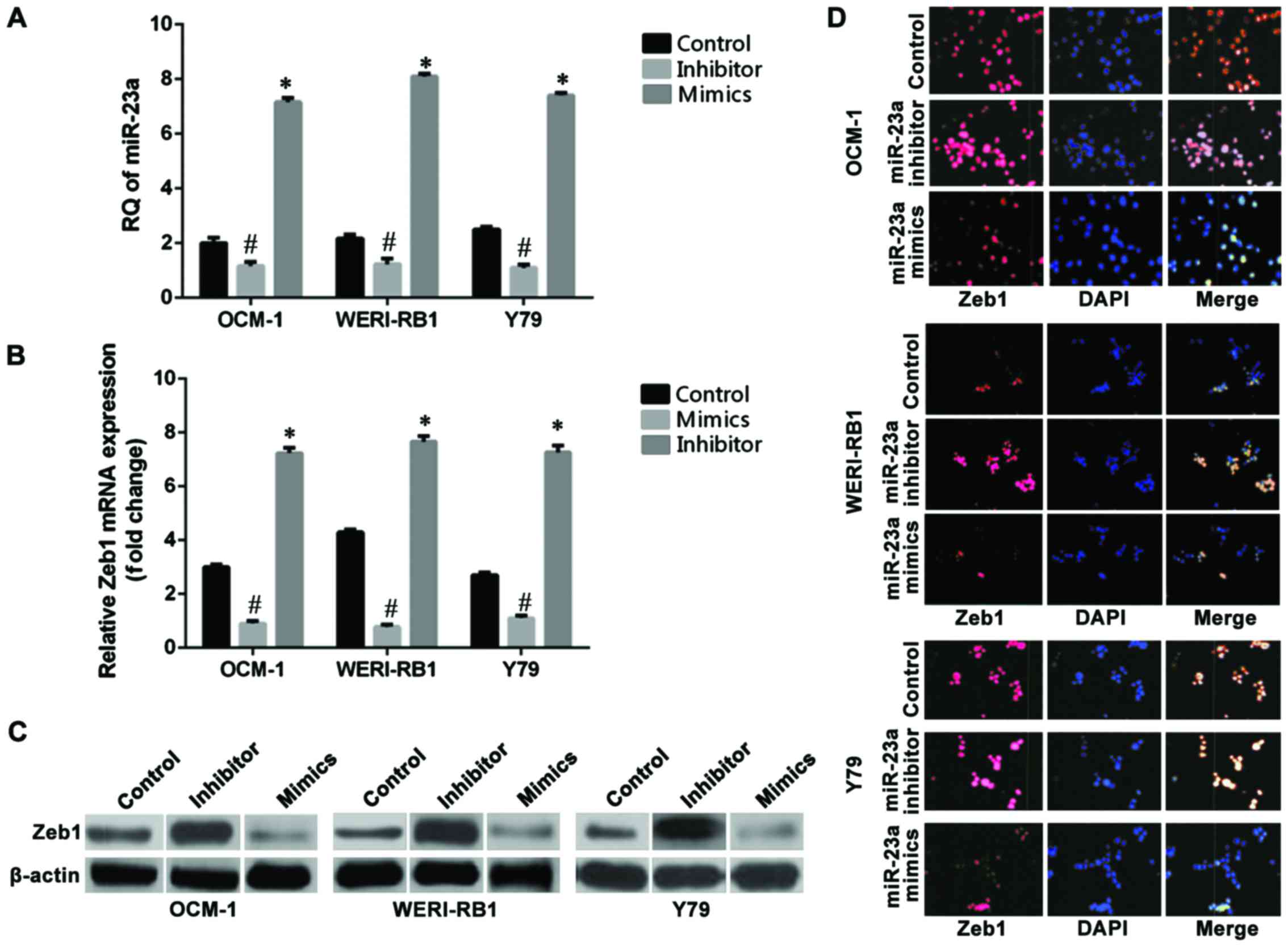
RT-qPCR was used to detect the expression levels of
miR-23a before and after transfection of cells. The results showed
(Fig. 1A) that the relative
quantitative (RQ) values of the non-transfection control group of
the OCM-1, WERI-RB1, and Y79 cell strains were 2.83, 3.28, and 2.3,
respectively. The relative expression levels of miR-23a in the
three groups of cells were 6.89, 7.74, and 5.86, respectively,
after the cells were transfected with the hsa-miR-23a mimic, which
increased significantly compared with those in the non-transfected
control group (p<0.05). The relative expression levels of
miR-23a in the three intraocular tumor cell strains were 0.19,
0.18, and 0.33, respectively, after the cells were transfected with
the hsa-miR-23a inhibitor, which decreased significantly compared
with those in the control group (p<0.05). RT-qPCR results showed
that the mRNA RQ values of Zeb1 in the non-transfected control
group of the OCM-1, WERI-RB1, and Y79 cell strains were OCM-1:
3.30, WERI-RB1: 4.45, and Y79: 2.74. The mRNA RQ values of Zeb1 of
the three cell strains were OCM-1: 1.07, WERI-RB1: 1.17, and Y79:
1.14 after they were transfected with the hsa-miR-23a mimic, which
decreased significantly compared with those in the non-transfected
control group (p<0.05) (Fig. 1B).
The cells in the miR-23a mimic transfection group, the miR-23a
inhibitor transfection group, and the non-transfection group of the
human choroidal melanoma cell strains OCM-1, human retinoblastoma
cell strains WERI-RB1, and Y79 were collected and subjected to
protein immunoblotting for determination of the changes in
expression of Zeb1 at the level of protein (Fig. 1C). The immunofluorescence assay of
Zeb1 in the three intraocular tumor cell strains were OCM-1: 6.99,
WERI-RB1: 7.81, and Y79: 6.92 after the cell strains were
transfected with the hsa-miR-23a inhibitor, which increased
significantly compared with those in the control group (p<0.05)
(Fig. 1D).
Role of miR-23a transfection in
regulating the EMT in intraocular tumors
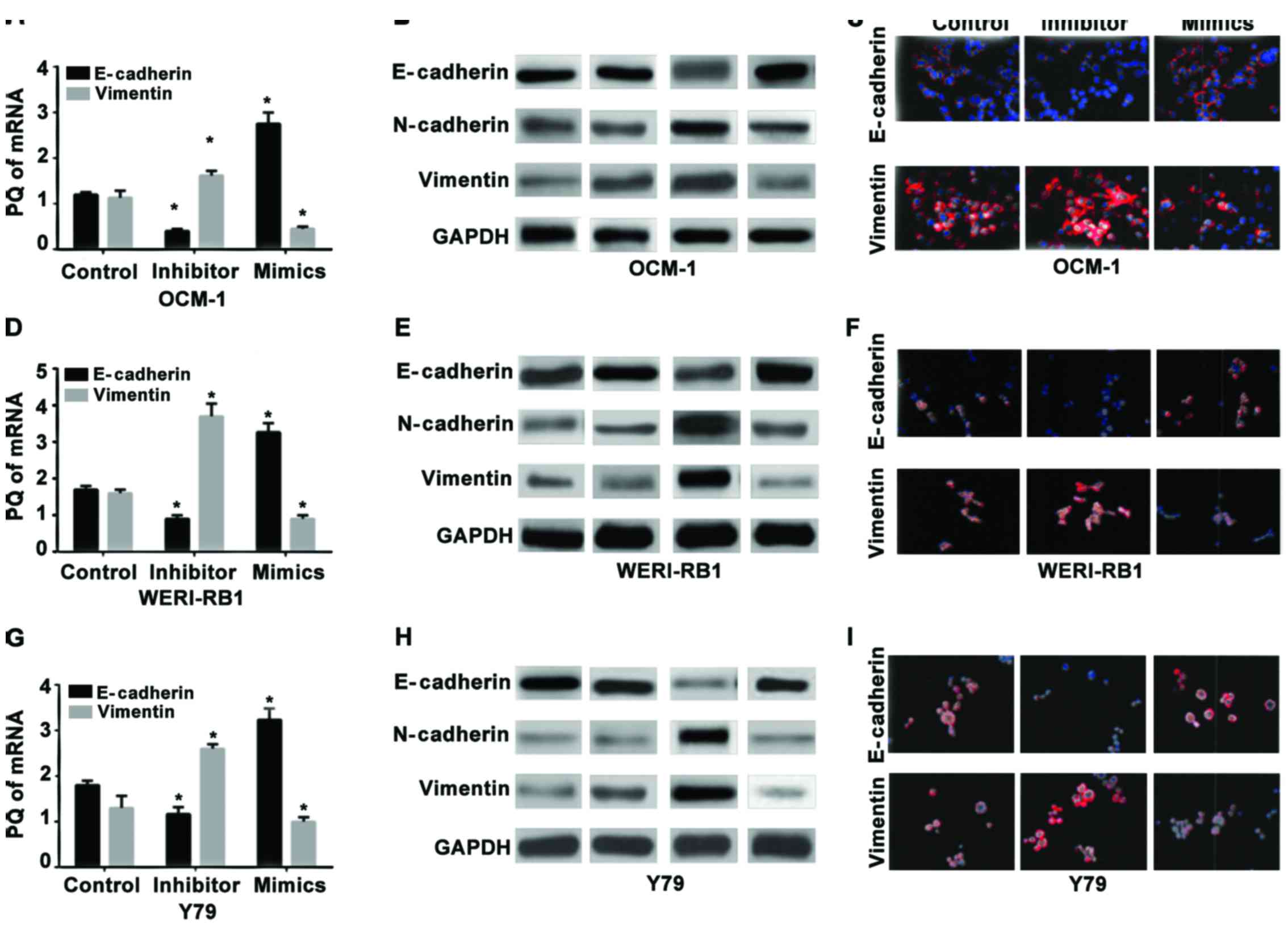
RT-qPCR was used to detect the expression levels of
mRNA in the epithelial cell-labeled gene E-cadherin and the
mesenchyme-labeled gene vimentin in the human choroidal melanoma
cell strains OCM-1 (Fig. 2A-C), human
retinoblastoma cell strains WERI-RB1 (Fig. 2D-F), and the Y79 cells (Fig. 2G-I) before and after miR-23a
transfection. The results showed that the relative expression
levels of mRNA of E-cadherin in the three strains of cells
following the hsa-miR-23a mimic transfection increased
significantly compared with those in the non-transfected control
group (p<0.05) while the expression levels of vimentin at the
level of RNA decreased significantly (p<0.05). Also, opposite
changes occurred in the miR-23a inhibitor groups of the three
strains of cells. The expression of E-cadherin decreased
significantly compared with that in the control group (p<0.05)
while the expression of vimentin increased significantly.
Effect of miR-23a transfection on the
migration capacity of the intraocular tumor cells
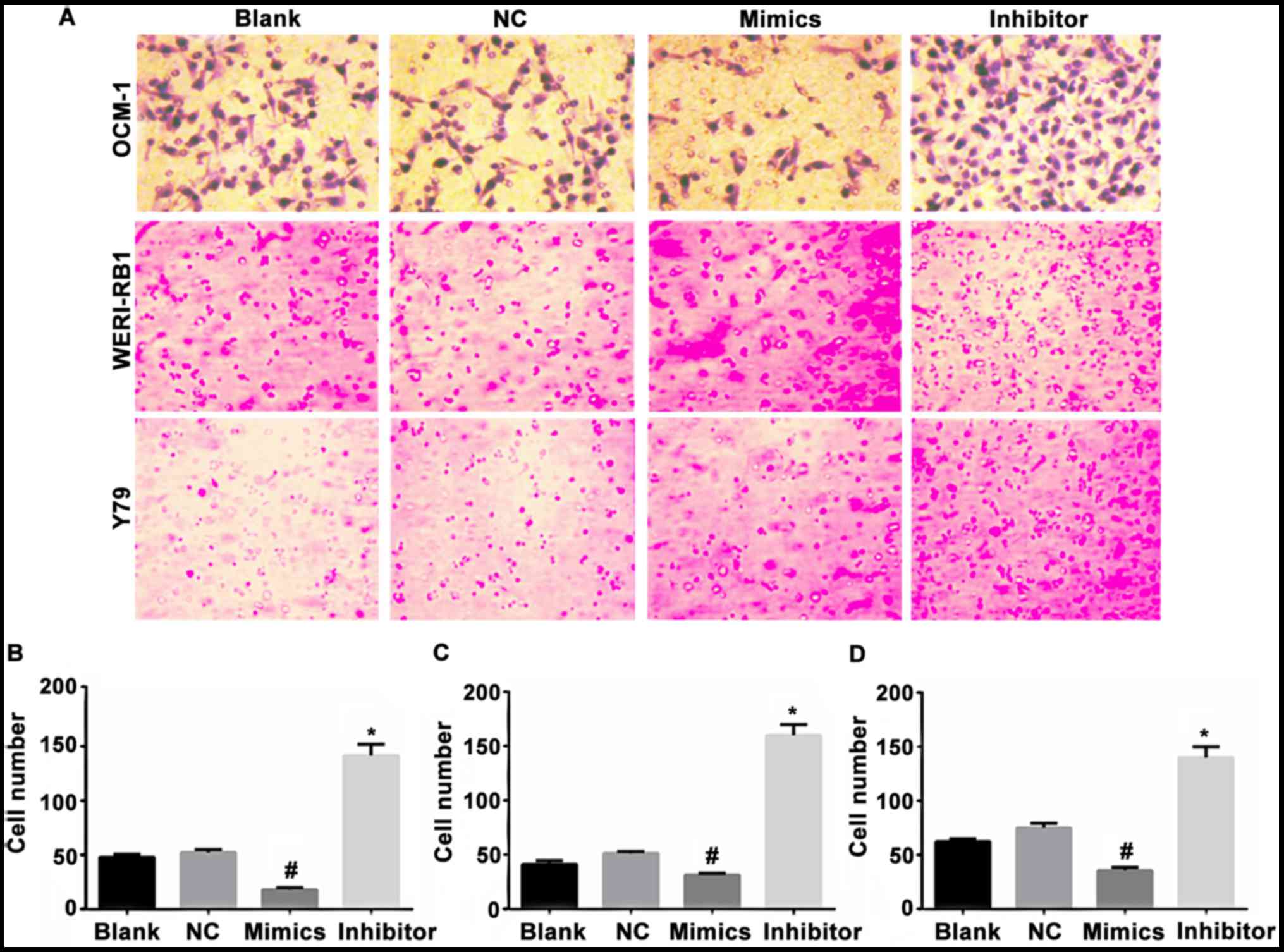
The cells in the non-transfected control group,
negative miR transfection control group, miR-23a mimic transfection
group, and the miR-23a inhibitor transfection group of the human
choroidal melanoma cell strains OCM-1, and human retinoblastoma
cell strains WERI-RB1 and Y79 were added to the Transwell 6-well
microplate. The cells that permeated the membrane and entered the
lower chamber were photographed and counted under high power lens.
The results showed that the average counts of the
membrane-permeating cells in the non-transfected control group
were: OCM-1: 54.75, WERI-RB1: 49.75, and Y79: 66.75. The average
counts of the cells in the negative transfection control group
were: OCM-1: 57.5, WERI-RB1: 55.25: Y79: 68.25. The differences
between the two groups were not statistically significant. The
average counts of the membrane-permeating cells in the miR-23a
mimic transfection group were: OCM-1: 25.75, WERI-RB1: 28.25, Y79:
32.5, which decreased when compared with those in the
non-transfection group (p<0.05) and negative control group
(p<0.05). The average counts of the cells in the miR-23a
inhibitor transfection group were: OCM-1: 153.5; WERI-RB1: 165.25;
Y79: 145, which increased significantly when compared with those in
the non-transfection group (p<0.05) and the negative control
group (p<0.05). Inhibition of miR-23a changed the shape of tumor
cells (Fig. 3).
Role of the Zeb1 gene in regulating
the feedback of miR-23a
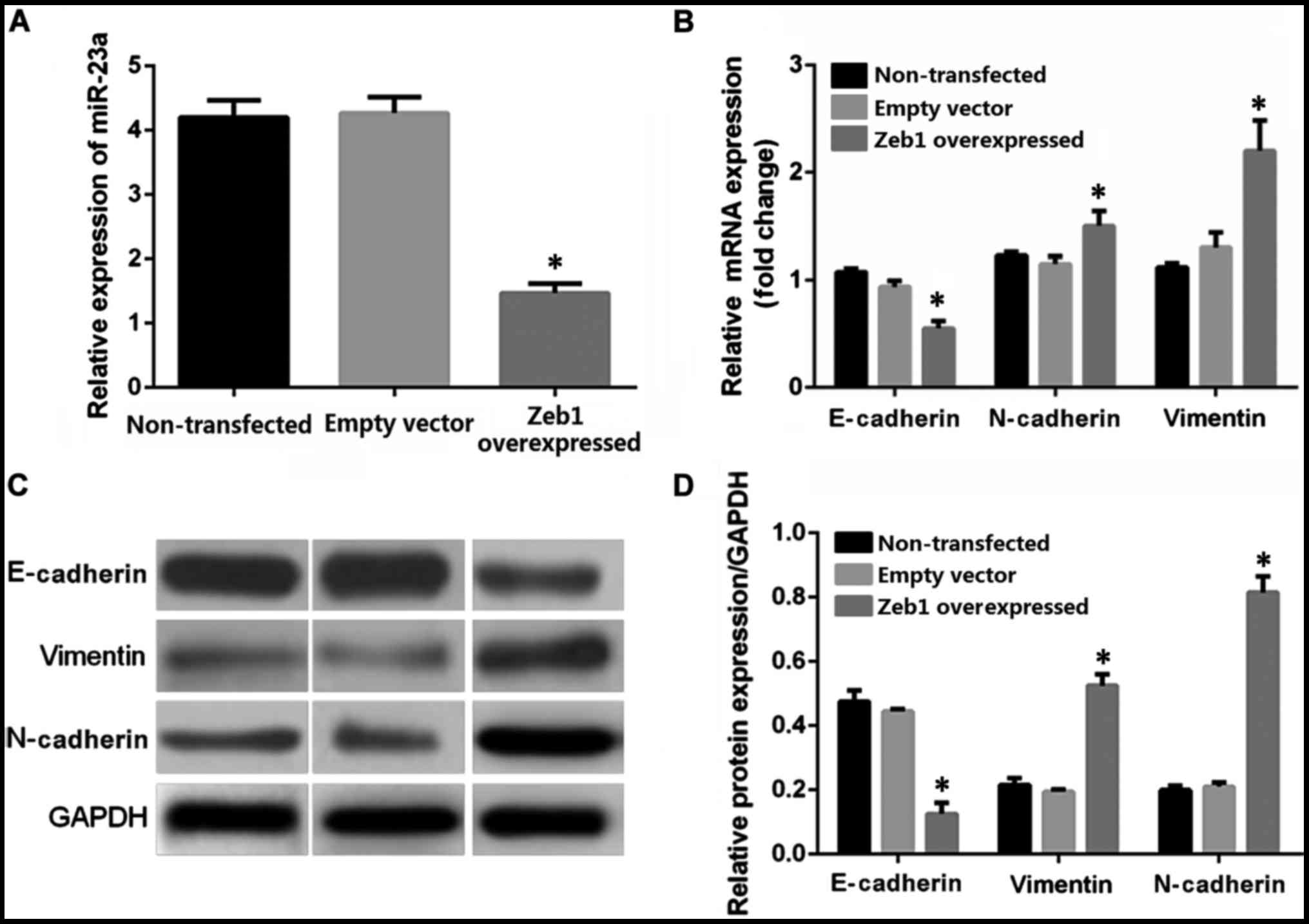
RT-qPCR was employed for real-time quantification
detection of miR-23a in the OCM-1 cells in the Zeb1 overexpressed
stably transfected group, the non-transfected control group, and
blank transfected control group. The results showed that the
average relative expression levels in the non-transfected control
group, empty vector control group, and the Zeb1 overexpressed
transfected group were 4.04, 4.13, and 1.34, respectively (Fig. 4A). The Zeb1 overexpressed transfection
decreased the expression level of miR-23a in the OCM-1 cells by
~67% relative to the control group, p<0.05. RT-qPCR was utilized
to detect the relative expression levels of mRNA in the
epithelium-derived marker E-cadherin, mesenchymal cell-labeled
genes vimentin and N-cadherin in the human choroidal melanoma OCM-1
cells of the Zeb1 gene overexpressed stable strains and the control
groups (non-transfected control group and empty vector transfection
control group). The average expression levels of E-cadherin in the
non-transfected control group, empty vector transfection control
group, and Zeb1 overexpressed transfected group were 1.09, 1, and
0.60, respectively. The expression level of E-cadherin in the
overexpressed transfection group decreased when compared with that
in the control group, p<0.05 (Fig.
4B). The average expression levels of vimentin in various
groups were 1.18, 1, and 1.69, respectively. The average expression
levels of N-cadherin were 1, 1.11, and 2.09. The average expression
level in the Zeb1 overexpressed transfected group increased
significantly when compared with that in the two control groups,
p<0.05. The differences between the blank control transfection
group and the non-transfection group were not significant. Western
blot analysis was used for quantitative analysis of the protein
levels in E-cadherin, vimentin, and N-cadherin in the OCM-1 cells
in the Zeb1 overexpression group and the two control groups. The
result was basically consistent with the RT-qPCR result. The Zeb1
overexpression transfection decreased the expression level of the
E-cadherin protein compared with that in the control group, whereas
the expression levels of vimentin and N-cadherin increased compared
with that in the control group (Fig. 4C
and D).
Discussion
Zeb1 is one of the important transcription factors
in EMT. It is bound to E-box in the genetic transcription promoter
sequence of the epithelial cell-labeled marker E-cadherin to
inhibit transcription (20) and
initiate EMT (21). Zeb1 is an
essential factor for cells to maintain their mesenchyme
characteristics. Normal epithelial tissue and low-malignant highly
differentiated cancers tend to not or seldom express the factor.
However, the Zeb1 gene is often highly expressed in poorly
differentiated malignant tumors. In other solid tumor cell strains,
miR-23a can directly act on and degrade Zeb1 thus eliminating its
inhibition over E-cadherin, upregulating the expression of
E-cadherin, reversing EMT, and producing mesenchymal epithelization
(22). Furthermore, research also
showed that the Zeb1 gene directly inhibits the E-cadherin gene via
E-box. A study suggested that microRNA-23a suppressed CDH1
expression and promoted EMT (23). In
this study, the results showed that the protein E-cadherin
increased while the expression of the mesenchyme-labeled proteins
of vimentin and N-cadherin decreased in the mimics group. This may
be caused by different cells used in different studies. Zeb1 has a
negative feedback effect on miR-23a. They can form a negative
feedback loop. The results showed that miR-23a and Zeb1 form a
bidirectional inhibitory negative feedback loop, which plays an
important role in regulating EMT. In addition, the compound formed
by the Zeb1 protein and the histone deacetylase may participate in
transcription and splicing of E-cadherin thus inhibiting its
expression. miR-23a or b can inhibit the formation of such a
compound thus promoting the expression of E-cadherin (24,25).
The expression of Zeb1 gene at the levels of mRNA
and protein in the OCM-1, WERI-RB1, and Y79 cell strains
transfected with miR-23a decreased significantly compared with that
in the control group. Moreover, the expression of RNA and protein
of the E-cadherin gene with epithelial cell specificity increased
significantly. However, the expression of the mesenchyme-labeled
proteins vimentin and N-cadherin decreased due to inhibition.
Mesenchymal-epithelial transition occurred in cells. On the
contrary, among the three intraocular tumor cell strains in the
miR-23a inhibitor transfection group, the expression level of Zeb1
increased significantly relative to the control group. The
expression level of E-cadherin decreased. The expression levels of
vimentin and N-cadherin increased. EMT occurred in the cells. This
conclusion is consistent with the result of multiple researches on
tumor cells at other sites. We also detected evidence for
epithelium-mesenchyme mutual transition in the intraocular tumor
cell strains.
We analyzed the expression level of miR-23a, the
epithelial marker E-cadherin and mesenchyme marker vimentin in nine
groups of cells (groups of non-transfected and transfected
intraocular tumor cell strains) and their ratio. It indicated that
miR-23a and the ratio of E-cadherin RNA and protein expression,
E-cadherin, and vimentin protein are positively correlated.
However, miR-23a and the expression level of the vimentin protein
are negatively correlated. It further demonstrated the roles of
miR-23a in promoting mesenchyme epithelization and inhibiting EMT.
However, the sample size in this study is excessively small and
deviation is also introduced to the effect of different
transfection intervention on EMT. Therefore, the relevant
significance obtained in this chapter can only be considered as a
prediction of the trend of the function of EMT in the three
intraocular tumors based on miR-23a.
The Transwell experiment was used to detect the
effect of the changes in the expression level of miR-23a on the
migration capacity of the three strains of tumor cells. It was
found that miR-23a can significantly inhibit the migration capacity
of these tumor cells. Inhibition of miR-23a changed the shape of
tumor cells. The capacity of the cells to permeate the
semipermeable membrane in the mimic transfection group decreased
significantly. The cells in the inhibitor transfection group
exhibited stronger capacity of motion and migration. In combination
with the detection results of epithelial and mesenchymal markers of
the cells in various groups, the changes in motion and migration
capacity of the tumor cells can be associated with EMT. EMT can
significantly strengthen the migration capacity of the cells while
mesenchymal-epithelial transition significantly decreases the
cellular migration capacity. The mesenchyme transition process
enables the epithelium-derived tumor cells to lose the original
closely-linked proteins and produce cytoskeletal proteins of
mesenchymal cells. They are more likely to change their morphology
and have stronger motion and transmembrane migration capacity.
Therefore, miR-23a probably changes the motion capacity of the
cells regulating the EMT of the tumor cells.
The human choroidal melanoma cell strains OCM-1 were
subjected to puromycin resistance screening at 72 h after
transfected with the Zeb1 overexpressed lentivirus particles to
obtain the stable transfected OCM-1 cell strains. RT-qPCR detection
has found that the expression of miR-23a in the tumor cells is
significantly inhibited, indicating that Zeb1 plays a role in
inhibiting the feedback of miR-23a. Both RT-qPCR and western blot
analysis showed that the expression of the epithelial cell gene
E-cadherin in the OCM-1 cells following Zeb1 overexpressed stable
transfection is downregulated. The expression of the genes of
vimentin and N-cadherin related to mesenchymal cells is
upregulated. It demonstrates the role of Zeb1 in promoting EMT. It
suggests that a negative feedback regulation loop with miR-23a/Zeb1
mutual inhibition occurs in the intraocular malignant tumor cell
strains, which coincides with the results of the experiments on
other human tumors. It demonstrates that mutual transition between
epithelial and mesenchymal cells can occur in the intraocular
malignant tumor cells. miR-23a can induce mesenchymal-epithelial
transition in the tumor cells whereas Zeb1 can promote EMT. The
tumorigenic ability of these tumor cells may increase with the
increase of EMT.
The mechanism of miR-23a regulating Zeb1 is still
unknown. It may be related to binding to 3′UTR. However, the Zeb1
3′UTR reporter gene was not done because only locus analysis was
used and the results of interfering miR-23a and overexpressing Zeb1
should be supported by interfering Zeb1 and overexpressing miR-23a.
Such tests will be carried out in the future. The expression of
miR-23a and Zeb1 should be recorded at different time-points to
test the ‘feedback’. Due to the cost of the experiment, this
experiment does not take into account the process of multiple
time-points repeated verification, so this project only discusses
the problem of specific time.
In conclusion, EMT is detected in the human
intraocular malignant tumor cells (choroidal melanoma cell strains
OCM-1, retinoblastoma cell strains WERI-RB1 and Y79). The report
verifies the presence of the bidirectional negative feedback
regulation loop of miR-23a/Zeb1 and the role of the pathway in
regulating EMT in tumor cells and regulating the migration capacity
of tumor cells in vitro in the intraocular tumor system.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW and HZ conceived and designed the study, and
drafted this paper. YW, YL and WG collected, analyzed and
interpreted the experiment data, and revised the manuscript
critically for important intellectual content. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the Affiliated Hospital of Inner Mongolia Medical University
(Hohhot, Inner Mongolia, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cho MH, Park JH, Choi HJ, Park MK, Won HY,
Park YJ, Lee CH, Oh SH, Song YS, Kim HS, et al: DOT1L cooperates
with the c-Myc-p300 complex to epigenetically derepress CDH1
transcription factors in breast cancer progression. Nat Commun.
6:78212015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ichikawa K, Kubota Y, Nakamura T, Weng JS,
Tomida T, Saito H and Takekawa M: MCRIP1, an ERK substrate,
mediates ERK-induced gene silencing during epithelial-mesenchymal
transition by regulating the co-repressor CtBP. Mol Cell. 58:35–46.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang L, Yang H, Abel EV, Ney GM, Palmbos
PL, Bednar F, Zhang Y, Leflein J, Waghray M, Owens S, et al: ATDC
induces an invasive switch in KRAS-induced pancreatic
tumorigenesis. Genes Dev. 29:171–183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen L, Gibbons DL, Goswami S, Cortez MA,
Ahn YH, Byers LA, Zhang X, Yi X, Dwyer D, Lin W, et al: Metastasis
is regulated via microRNA-200/ZEB1 axis control of tumour cell
PD-L1 expression and intratumoral immunosuppression. Nat Commun.
5:52412014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Y, Bu F, Royer C, Serres S, Larkin
JR, Soto MS, Sibson NR, Salter V, Fritzsche F, Turnquist C, et al:
ASPP2 controls epithelial plasticity and inhibits metastasis
through β-catenin-dependent regulation of ZEB1. Nat Cell Biol.
16:1092–1104. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang P, Wei Y, Wang L, Debeb BG, Yuan Y,
Zhang J, Yuan J, Wang M, Chen D, Sun Y, et al: ATM-mediated
stabilization of ZEB1 promotes DNA damage response and
radioresistance through CHK1. Nat Cell Biol. 16:864–875. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chao CH, Chang CC, Wu MJ, Ko HW, Wang D,
Hung MC, Yang JY and Chang CJ: MicroRNA-205 signaling regulates
mammary stem cell fate and tumorigenesis. J Clin Invest.
124:3093–3106. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Park KS, Raffeld M, Moon YW, Xi L, Bianco
C, Pham T, Lee LC, Mitsudomi T, Yatabe Y, Okamoto I, et al: CRIPTO1
expression in EGFR-mutant NSCLC elicits intrinsic EGFR-inhibitor
resistance. J Clin Invest. 124:3003–3015. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ,
Tao QF, Liu F, Pan W, Wang TT, Zhou CC, et al: A long noncoding RNA
activated by TGF-β promotes the invasion-metastasis cascade in
hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lehmann W, Mossmann D, Kleemann J, Mock K,
Meisinger C, Brummer T, Herr R, Brabletz S, Stemmler MP and
Brabletz T: ZEB1 turns into a transcriptional activator by
interacting with YAP1 in aggressive cancer types. Nat Commun.
7:104982016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zheng X, Carstens JL, Kim J, Scheible M,
Kaye J, Sugimoto H, Wu CC, LeBleu VS and Kalluri R:
Epithelial-to-mesenchymal transition is dispensable for metastasis
but induces chemoresistance in pancreatic cancer. Nature.
527:525–530. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang Y, Ahn YH, Chen Y, Tan X, Guo L,
Gibbons DL, Ungewiss C, Peng DH, Liu X, Lin SH, et al: ZEB1
sensitizes lung adenocarcinoma to metastasis suppression by PI3K
antagonism. J Clin Invest. 124:2696–2708. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tian K, Di R and Wang L: MicroRNA-23a
enhances migration and invasion through PTEN in osteosarcoma.
Cancer Gene Ther. 22:351–359. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wen YC, Lee WJ, Tan P, Yang SF, Hsiao M,
Lee LM and Chien MH: By inhibiting snail signaling and miR-23a-3p,
osthole suppresses the EMT-mediated metastatic ability in prostate
cancer. Oncotarget. 6:21120–21136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tang J, Zhao H, Cai H and Wu H: Diagnostic
and prognostic potentials of microRNA-27a in osteosarcoma. Biomed
Pharmacother. 71:222–226. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang XW, Liu N, Chen S, Wang Y, Zhang ZX,
Sun YY, Qiu GB and Fu WN: High microRNA-23a expression in laryngeal
squamous cell carcinoma is associated with poor patient prognosis.
Diagn Pathol. 10:222015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma G, Dai W, Sang A, Yang X and Gao C:
Upregulation of microRNA-23a/b promotes tumor progression and
confers poor prognosis in patients with gastric cancer. Int J Clin
Exp Pathol. 7:8833–8840. 2014.PubMed/NCBI
|
|
18
|
He Y, Meng C, Shao Z, Wang H and Yang S:
MiR-23a functions as a tumor suppressor in osteosarcoma. Cell
Physiol Biochem. 34:1485–1496. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang L, Yang X, Ji W, Deng J, Qiu F, Yang
R, Fang W, Zhang L, Huang D, Xie C, et al: Effects of a functional
variant c.353T>C in snai1 on risk of two contextual diseases.
Chronic obstructive pulmonary disease and lung cancer. Am J Respir
Crit Care Med. 189:139–148. 2014.PubMed/NCBI
|
|
20
|
Li C, Ma H, Wang Y, Cao Z, Graves-Deal R,
Powell AE, Starchenko A, Ayers GD, Washington MK, Kamath V, et al:
Excess PLAC8 promotes an unconventional ERK2-dependent EMT in colon
cancer. J Clin Invest. 124:2172–2187. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Caramel J, Papadogeorgakis E, Hill L,
Browne GJ, Richard G, Wierinckx A, Saldanha G, Osborne J,
Hutchinson P, Tse G, et al: A switch in the expression of embryonic
EMT-inducers drives the development of malignant melanoma. Cancer
Cell. 24:466–480. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hur K, Toiyama Y, Takahashi M, Balaguer F,
Nagasaka T, Koike J, Hemmi H, Koi M, Boland CR and Goel A:
MicroRNA-200c modulates epithelial-to-mesenchymal transition (EMT)
in human colorectal cancer metastasis. Gut. 62:1315–1326. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng L, Yang T, Kuang Y, Kong B, Yu S,
Shu H, Zhou H and Gu J: MicroRNA-23a promotes neuroblastoma cell
metastasis by targeting CDH1. Oncol Lett. 7:839–845. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Celià-Terrassa T, Meca-Cortés O, Mateo F,
de Paz Martínez A, Rubio N, Arnal-Estapé A, Ell BJ, Bermudo R, Díaz
A, Guerra-Rebollo M, et al: Epithelial-mesenchymal transition can
suppress major attributes of human epithelial tumor-initiating
cells. J Clin Invest. 122:1849–1868. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mizuguchi Y, Isse K, Specht S, Lunz JG
III, Corbitt N, Takizawa T and Demetris AJ: Small proline rich
protein 2a in benign and malignant liver disease. Hepatology.
59:1130–1143. 2014. View Article : Google Scholar : PubMed/NCBI
|