Introduction
Lung cancer is the leading cause of death for both
men and women in many countries (1).
Νon-small cell lung cancer (NSCLC) accounts for approximately 85%
of lung cancer cases (2). Surgery is
the most effective treatment for NSCLC if the cancer is diagnosed
early. However, up to 75% of patients are diagnosed with NSCLC at
an advanced disease (3,4). Although there has been some progress in
methods for diagnosis and treatment, the prognosis of the patients
with NSCLC has not improved much, and the 5-year survival rate is
currently less than 15% (5).
Therefore, there is a need to develop new and safe therapeutic
agents against NSCLC.
Conventional chemotherapies kill normal cells in
addition to cancer cells by inducing cell cycle checkpoint arrest
or apoptosis. The initiation of apoptosis is tightly regulated by
intrinsic caspase-dependent pathways (including one that is
mitochondrion-dependent) and extrinsic caspase-dependent pathways
(such as the death receptor-induced caspase activation pathway)
(6,7).
A shared a terminal execution pathway begins with caspase-3
cleavage and ultimately leads to phagocytosis (8). The best characterized regulators of
apoptosis are the Bcl-2 family of proteins, which promote or
repress apoptosis by directly regulating a mitochondrial
apoptosis-induced channel. The multidomain pro-apoptotic protein
Bax/Bak is responsible for forming this channel, while Bcl-2,
Bcl-xL, or Mcl-1 inhibit its formation (9,10).
Additionally, the intrinsic ER stress pathway is triggered by
conditions that disturb protein folding in the endoplasmic
reticulum (ER). The induction of the ER stress pathway may
contribute to cell death by activating the IRE1 kinase via
activation of APOPTOTIC-SIGNALING KINASE (ASK) and p38 MAPK, which
itself activates CHOP by phosphorylating the CHOP transactivation
domain (11). Additionally, both p38
MAPK and the Jun-N-terminal kinase (JNK) were reported to promote
phosphorylation and activation of Bak (12). In addition, Bax and Bak induce cell
death by binding and activating INOSITOL-REQUIRING PROTEIN-1α
(IRE1α) (13). Intriguingly,
accumulating evidence indicates that activated p38 can cause
mitotic arrest in the somatic cell cycle at the spindle assembly
checkpoint (14).
Traditional herbal medicines with anti-cancer
properties that trigger apoptosis in various types of tumor cells
with minimal adverse effects have been valuable sources for
therapeutic agents. Polygonatum odoratum is widely used as
an herbal medicine with procoagulant (15), anti-hyperglycemic (16), anti-herpes simplex virus-II, and
apoptosis-inducing (17) activities,
that can also improve glucose tolerance (18). However, the active components in P.
odoratum for the anti-cancer effects and the underlying
mechanisms of these effects remain largely unknown.
Homoisoflavanone-1 is a type of phenolic compound
isolated from P. odoratum that has apparent antioxidant
activities (19,20). However, the effects of
homoisoflavanone-1 on human NSCLC cells, and therefore the
mechanism of this effect, have never been elucidated. In the
present study, we investigated the effect of homoisoflavanone-1 on
NSCLC A549 cell proliferation and cell cycle progression. Our
result shows that homoisoflavanone-1 has potential as a new natural
anti-tumor medicine for treatment of NSCLC.
Materials and methods
Cell culture and reagents
The human NSCLC cell line A549 was purchased from
Basic medical cell center of Peking Union Medical College (Peking,
China). A549 cells were cultured in DMEM containing 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), 1% penicillin, and 1% streptomycin in 5% CO2 at
37°C. Cells in the exponential phase were used in the
experiments.
Dimethyl sulfoxide (DMSO) and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
were purchased from Amresco, LLC, (Solon, OH, USA) and Propidium
iodide (PI)/Annexin V-FITC was obtained from Sigma-Aldrich; Merck
KGaA, (Darmstadt, Germany). Polyclonal antibodies against Caspase
3, Active caspase 3, Bak, Bcl-2, p-p38, p38, p53, Cdc2, p-Cdc2,
β-actin, and the horseradish peroxidase-conjugated secondary
antibody were bought from Cell Signaling Technology, Inc.,
(Danvers, MA, USA).
Isolation and purification of
homoisoflavanone-1
The homoisoflavanone-1 used in this study was
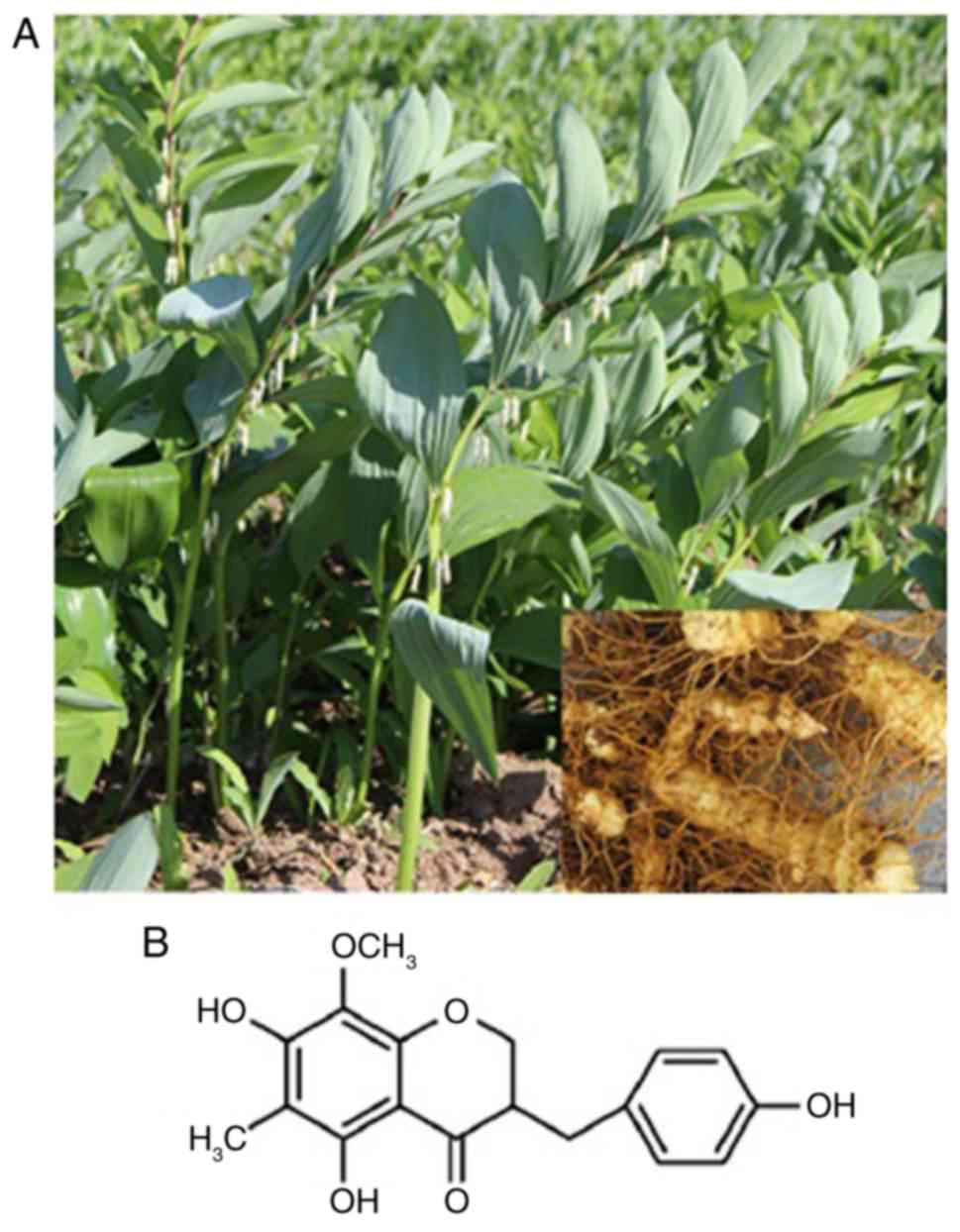
extracted and purified from P. odoratum roots (Fig. 1A) according to the methods described
previously with appropriate modification (20). Briefly, 10 kg of dried P. odoratum
roots was ground and subjected to two 95% ethanol extractions at
room temperature. The solvent was removed under reduced pressure,
and the concentrate was diluted in water, followed by filtering.
The precipitate including insoluble metabolites was dissolved in
90% methanol. The methanol-soluble fraction was collected and
subjected to chromatography on a silica gel column, with a gradient
of petroleum ether and petroleum ether-ethyl acetate as the eluting
solvent, followed by thin layer chromatography to collect cytotoxic
fractions. Based on HPLC analysis of the collected components, an
elution with a single component was collected. Homoisoflavanone-1
(17.84 mg) was isolated by reverse-phase preparative HPLC, using
methanol:H2O (60:40) as the mobile phase, and was
identified by comparing ESI-MS/MS and spectroscopic
(1H-NMR and 13C-NMR) data.
The structures of homoisoflavanone-1 were in good
agreement with a previous report (20) of it being
5,7-dihydroxyl-6-methyl-8-methoxyl-3-(4′-hydroxylbenzyl)-chroman-4-one
(Fig. 1B).
Cell viability assay
The effect of homoisoflavanone-1 on A549 cell
viability was determined using the MTT assay. Briefly, cells were
incubated in 96-well plates at a density of 10×104 per
well for 24 h before treatment with homoisoflavanone-1 or DMSO, as
the control. Homoisoflavanone-1 was dissolved in DMSO at serial
concentrations (12.5, 25, 50, and 100 µg/ml), and 8 µl of these
solutions were added to each well and the samples were incubated
for 6, 12, and 24 h. The results were read using a microplate
spectrophotometer (BioTek Instruments, Inc., Winooski, VT, USA) at
570 nm. The rate of cell growth inhibition (%) was calculated as:
[1-A549 experiment group OD/A549 control OD] ×100%. The half
maximal inhibitory concentration (IC50) values were
determined by plotting a linear regression curve.
Colony formation assay
A549 cells were seeded in triplicate into 12-well
tissue culture plates and cultured for 6 days after treatment with
homoisoflavanone-1 at serial concentrations (12.5, 25, 50, and 100
µg/ml). Then, crystal violet was used for staining, and the rate of
colony formation was calculated.
Cell cycle analysis
A549 cells were seeded into 6-well plates, incubated
for 12 h, and then treated with either homoisoflavanone-1 (12.5,
25, 50, or 100 µg/ml) or DMSO for 24 h. Cells were collected, fixed
in 70% ethanol, centrifuged (3,000 g, 5 min) and washed twice with
ice-cold PBS. The cells were stained with PI for 20 min at 4°C in
the dark. Cell cycle analysis was carried out with a
fluorescence-activated cell sorting (FACS) Calibur flow cytometer
(BD FACSCalibur; BD Biosciences, Franklin Lakes, NJ, USA USA).
Cell apoptosis analysis
Apoptosis analyses were performed with an Annexin
Vfluorescein isothiocyanate (FITC) apoptosis detection kit
(BioVision, Inc., Milpitas, CA, USA). A549 cells were seeded in
96-well plates, incubated for 12 h, and then treated with
homoisoflavanone-1 (12.5, 25, 50, or 100 µg/ml) for 24 h. Cells
were collected and washed twice with ice-cold PBS, followed by
incubation with Annexin-V for 10 min in the dark. Next, the cells
were incubated with PI for 5 min and evaluated for apoptosis using
a FACS Calibur flow cytometer (BD FACSCalibur; BD Biosciences).
Immunoblot analysis
A549 cells were seeded in 6-well plates for 12 h,
and then treated with either homoisoflavanone-1 (12.5, 25, 50, or
100 µg/ml) or DMSO for 24 h. Cellular proteins were harvested and
immunoblotting was carried out as previously described (21) with appropriate modifications. Cells
were resuspended in lysis buffer (1% Triton-X100, 50 mM Hepes pH
7.4, 2 mM sodium orthovanadate, 1 mM edetic acid, 100 mM sodium
fluoride, 1 mM PMSF, 10 µg/ml of leupeptin and 10 µg/ml of
aprotinin), and the supernatants of the lysates were collected
after centrifugation at 14,000 g for 10 min at 4°C. The protein
concentration was determined using a protein concentration kit.
Proteins were separated with 12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to
polyvinyldene fluoride membranes. The membranes were blocked in 5%
skimmed milk for 2 h at room temperature. Protein was detected with
antibodies against active caspase 3, caspase 3, PARP, Bcl-2, Bak,
p-Cdc2, Cdc2, p-P38, P38, P53 or β-actin at 4°C. The membranes were
washed three times with TBST before incubation with the horseradish
peroxidase-conjugated secondary antibody at room temperature for 1
h. Immunoreactive bands were visualized using an ECL kit
(Sigma-Aldrich; Merck KGaA). Protein levels were quantified
relative to the control group.
Statistical analysis
All experiments were performed at least three
independent times (n=3), and data are presented as the mean ±
standard deviation (SD). The comparisons of multiple groups were
performed using the one-way analysis of variance and the group
differences were analyzed using Dunnett's post hoc tests.
Statistical analyses were performed in SPSS v.13.0 (SPSS, Inc.,
Chicago, IL, USA) and P<0.05 was considered to indicate a
statistically significant difference.
Results
Homoisoflavanone-1 inhibits cell
proliferation and colony formation of A549 Cells
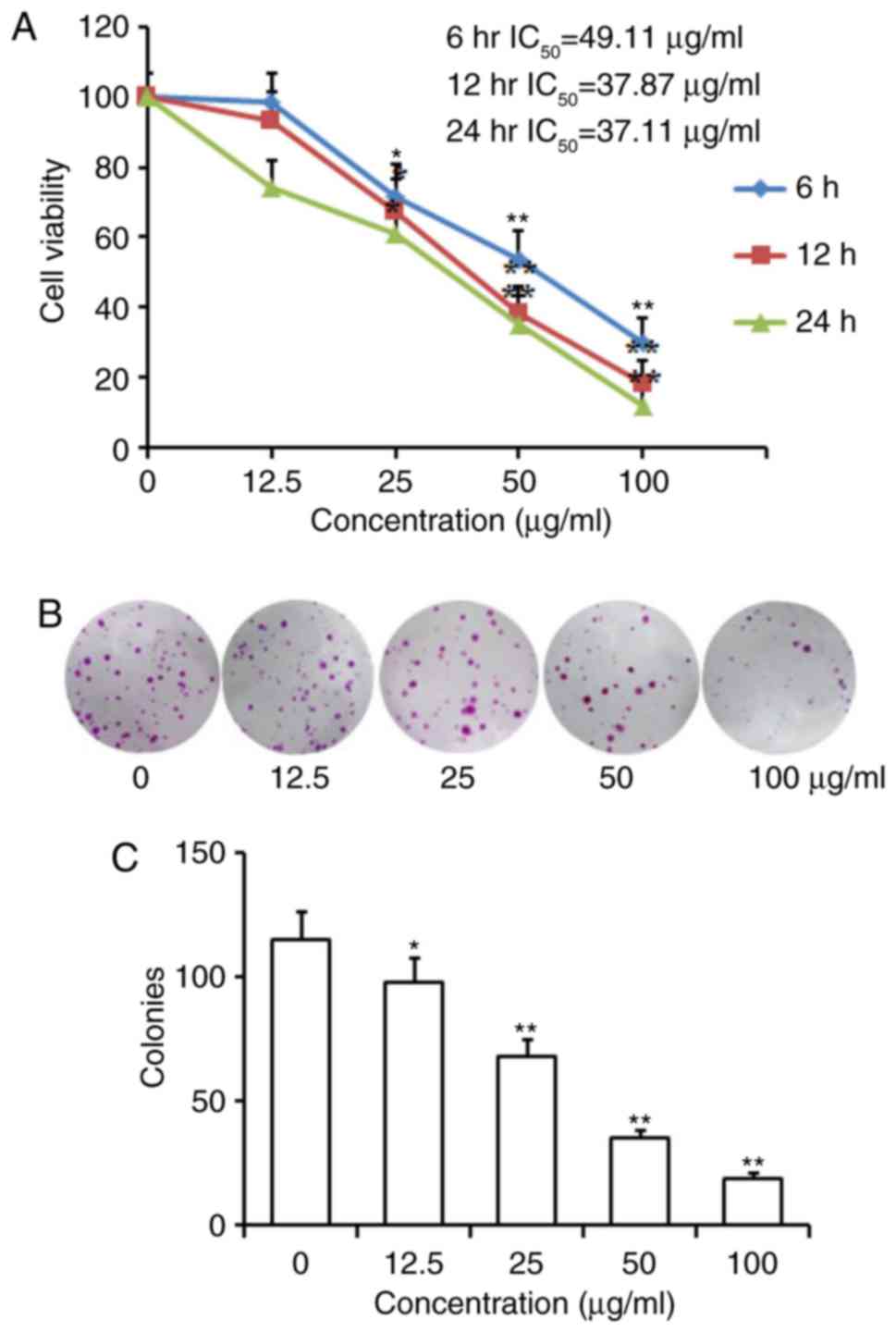
An MTT assay was performed to determine the effect
of homoisoflavanone-1 on the proliferation of A549 cells. As shown
in Table I and Fig. 2A, homoisoflavanone-1 inhibited cell
growth in a dose- and time-dependent manner. For example, a
remarkable inhibitory effect was observed in cells treated with
higher concentrations of homoisoflavanone-1 (50, 100 µg/ml) for 6,
12, and 24 h vs. controls at the same time points (P<0.01), and
a significant effect on the growth of A549 cells was still observed
at a lower concentration of homoisoflavanone-1 (25 µg/ml) for 12,
24 h (P<0.05). The IC50 values were 49.11, 37.87 and 37.11 µg/ml
for 6, 12 and 24 h incubation with homoisoflavanone-1,
respectively. In addition, the colony formation rate of A549 cells
in homoisoflavanone-1-treated groups was obviously lower than that
of the control group (Fig. 2B and
C).
 | Table I.Effects of homoisoflavanone-1 on cell
proliferation in lung cancer A549 cells. |
Table I.
Effects of homoisoflavanone-1 on cell
proliferation in lung cancer A549 cells.
| Concentration
(µg/ml) | 6 h | 12 h | 24 h |
|---|
| 0 | 99.04±4.47 | 123.15±9.50 | 114.18±15.21 |
| DMSO | 100.00±0.00 | 100.00±0.00 | 100.00±0.00 |
| 12.5 | 98.80±0.57 | 93.54±26.32 | 94.15±21.33 |
| 25 | 71.70±0.05 |
67.47±22.90a | 67.14±25.07
a |
| 50 |
53.66±4.94b |
38.11±5.94a | 39.27±27.99
b |
| 100 | 22.86±4.84
b |
11.78±5.61a |
7.51±1.22b |
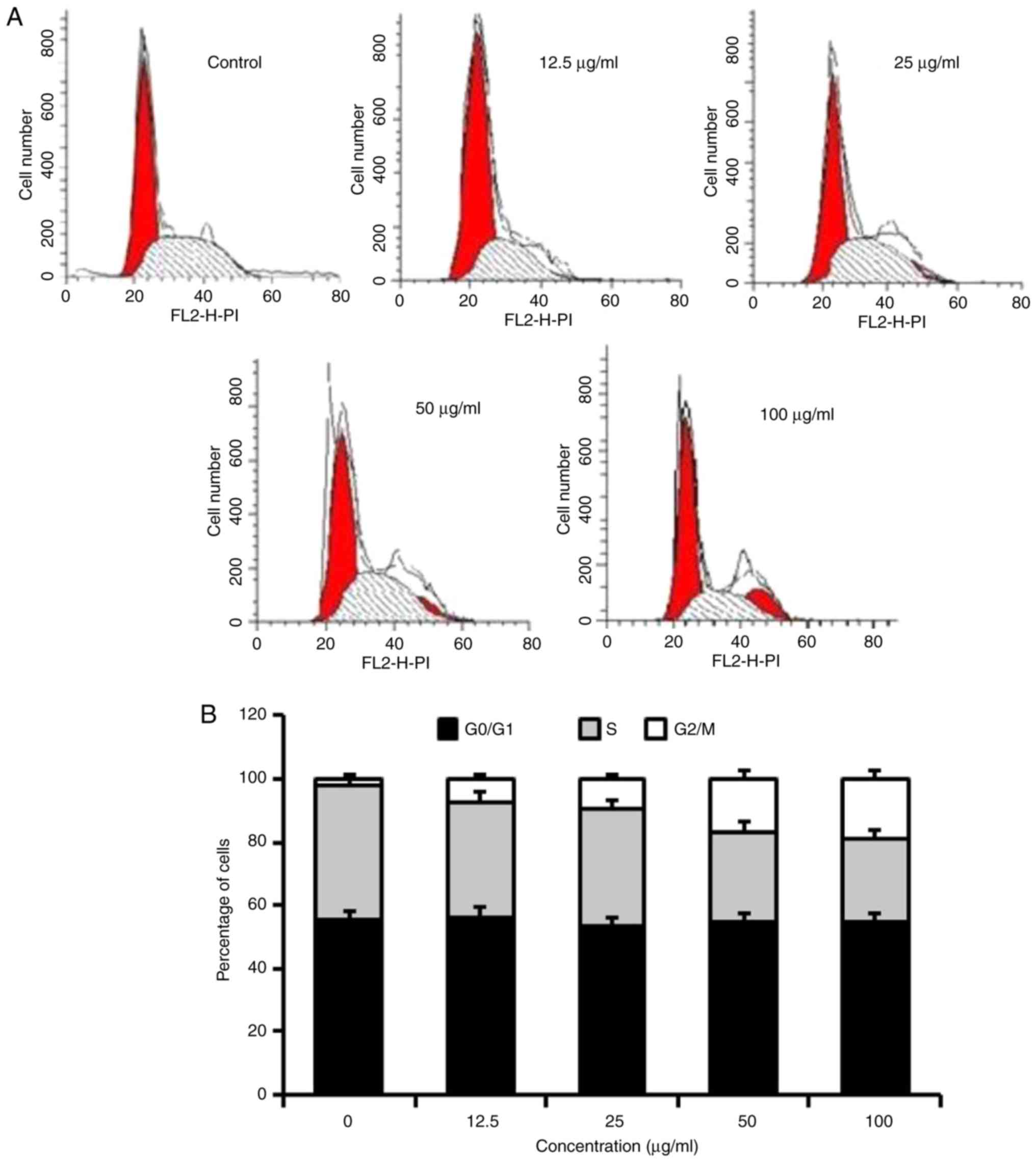
Homoisoflavanone-1 arrests the cell
cycle between the G2/M phases in A549 cells
To address the precise action responsible for the
anti-proliferative effect mediated by homoisoflavanone-1, the cell
cycle distribution profile was examined. As indicated in Table II, A549 cells were exposed to 0,
12.5, 25, 50, or 100 µg/ml homoisoflavanone-1 for 24 h. The ratio
of the G2/M phase was 1.07, 7.34, 10.13, 12.75 and 17.76 for 0,
12.5, 25, 50 and 100 µg/ml, respectively. There was no difference
in the proportions of cells in the G0/G1 phases, however, the
proportion of cells in the S phase was also reduced in response to
homoisoflavanone-1 treatment (Fig. 3A and
B).
| Table II.Effects of homoisoflavanone-1 on the
cell cycle of A549 cells. |
Table II.
Effects of homoisoflavanone-1 on the
cell cycle of A549 cells.
| Concentration
(µg/ml) | G0/G1 phase | S phase | G2/M phase |
|---|
| 0 | 55.39±2.37 | 43.99±1.50 | 1.07±1.11 |
| 12.5 | 57.89±2.57 | 36.39±2.09 |
7.34±1.31a |
| 25 | 53.48±3.05 | 36.92±1.98 |
10.13±2.07a |
| 50 | 48.54±2.97 | 27.73±1.71* |
12.75±1.89b |
| 100 | 54.52±2.86 | 26.21±1.42* |
17.76±1.43b |
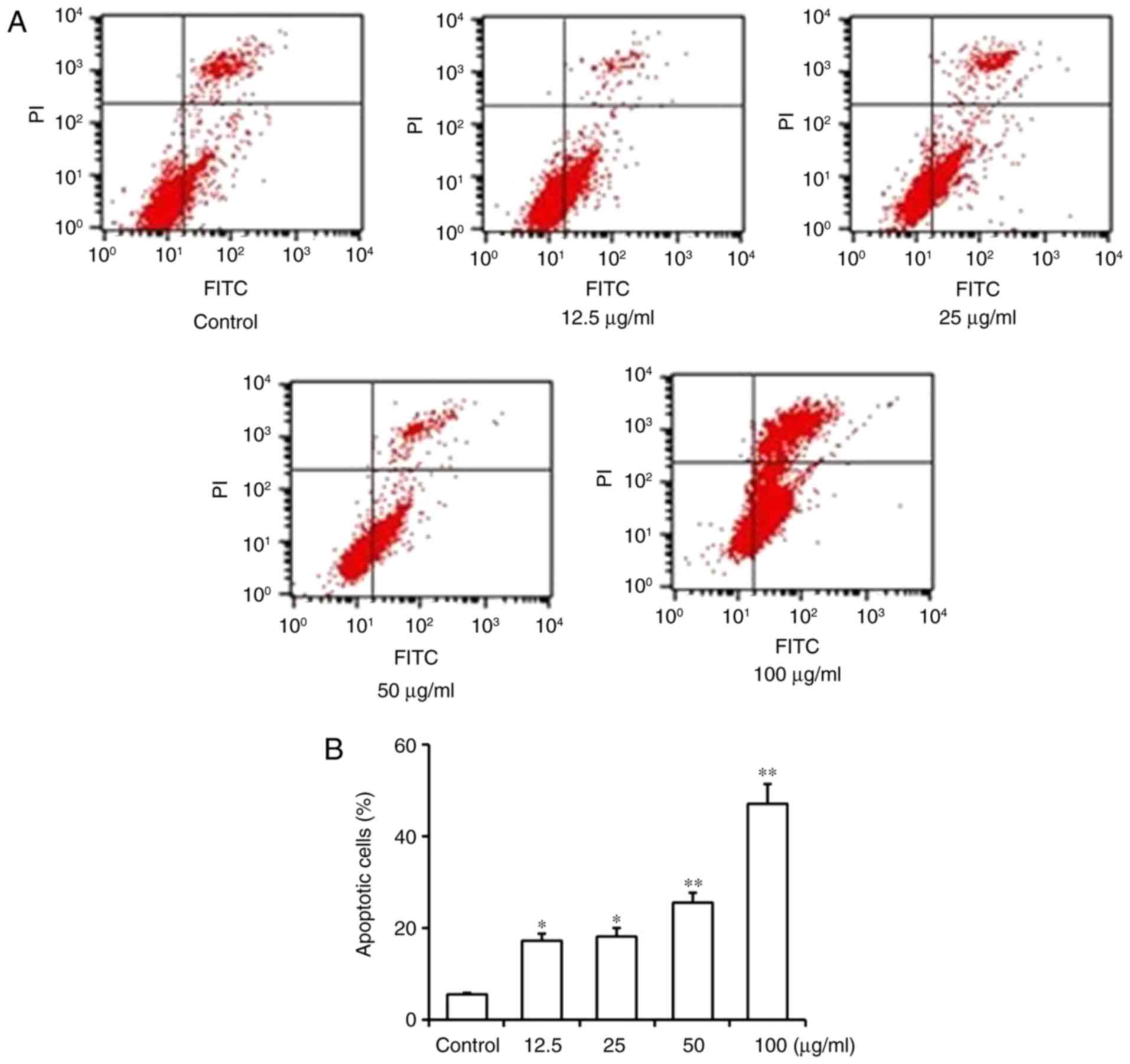
Homoisoflavanone-1 induces apoptosis
in A549 cells
We conducted flow cytometry to evaluate the effect
of homoisoflavanone-1 on A549 cell apoptosis. The results
demonstrate that A549 cells treated with increasing concentrations
(12.5, 25, 50 and 100 µg/ml) of homoisoflavanone-1 over 24 h showed
an increase in the proportion of apoptotic cells (17.22, 18.23,
25.53, and 47.12%, respectively) (Fig. 4A
and B). These data showed that homoisoflavanone-1 induced
dose-dependent apoptosis in the cells and significantly increased
the rate of apoptosis compared with untreated control cells
(5.44%).
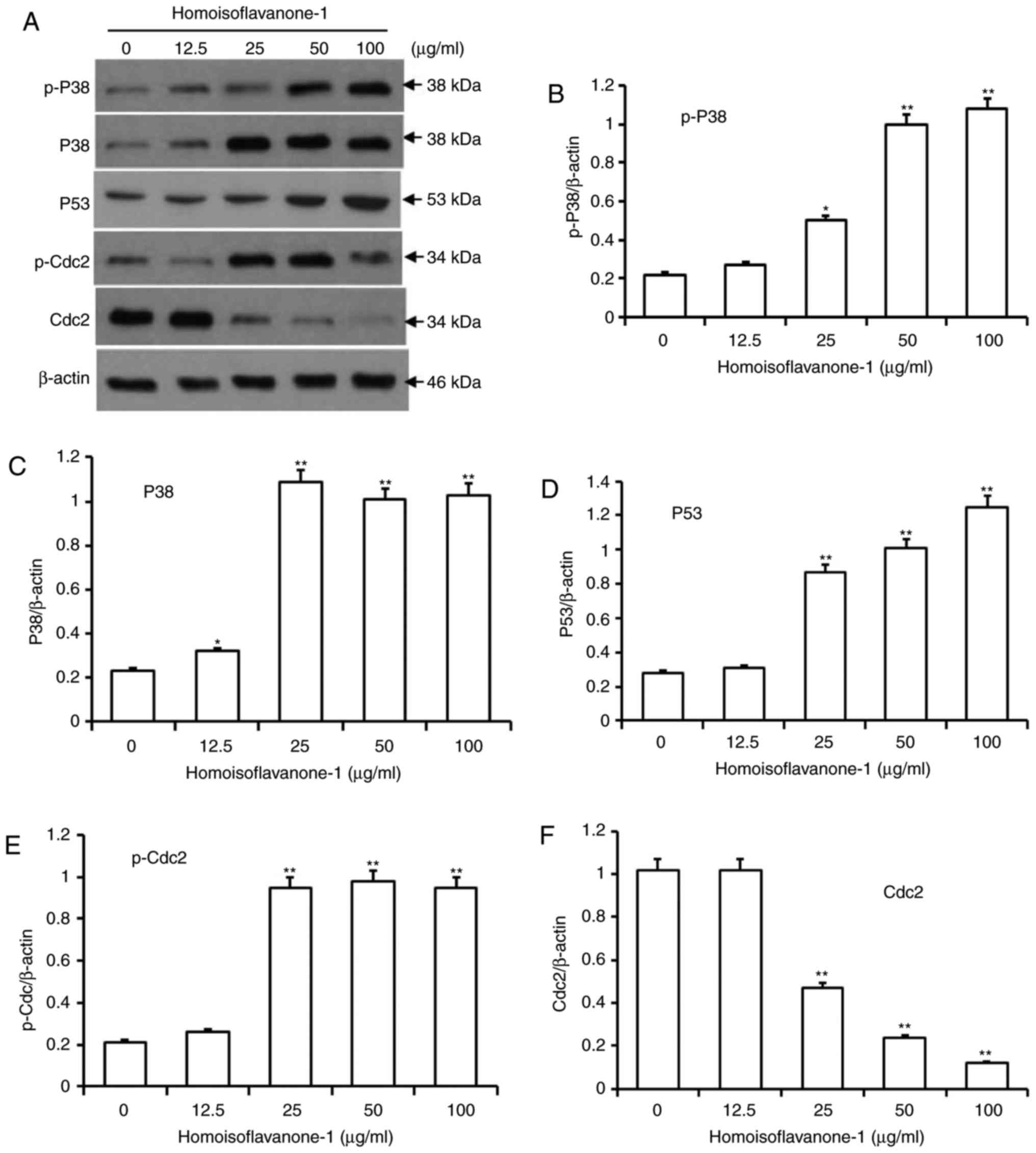
Homoisoflavanone-1 regulates cell
cycle-related proteins in A549 Cells
Activated p38, which is involved in various types
cell differentiation, can cause mitotic arrest at the spindle
assembly checkpoint in somatic cell cycle (14,22).
Additionally, p53 is a key regulator mediating cell cycle arrest
induced by a range of factors, including DNA damage and exposure to
chemotherapeutic compounds (23). To
investigate the molecular events governing
homoisoflavanone-1-mediated cell cycle retardation, we used
immunoblots to detect the protein levels of p-p38, p38 and p53 in
A549 cells after 24 h of various doses of homoisoflavanone-1 (0,
12.5, 25, 50 and 100 µg/ml) treatment. As shown in Fig. 5A-D, there was a dose-dependent
increase in the active forms of p-p38, p38 and p53 in response to
homoisoflavanone-1 treatment.
Cdc 2/cyclin B complexes initiate mitosis (M) by
phosphorylating both regulatory and structural proteins involved in
mitosis (24). CDK activity is itself
regulated by phosphorylation and dephosphorylation, as simply
binding to cyclins is not sufficient for activation of the
complexes (25). As shown in Fig. 5E and F, p-Cdc2 levels increased in a
dose-dependent manner in cells that were treated with
homoisoflavanone-1 (0, 12.5, 25, 50, and 100 µg/ml), while Cdc2
levels decreased.
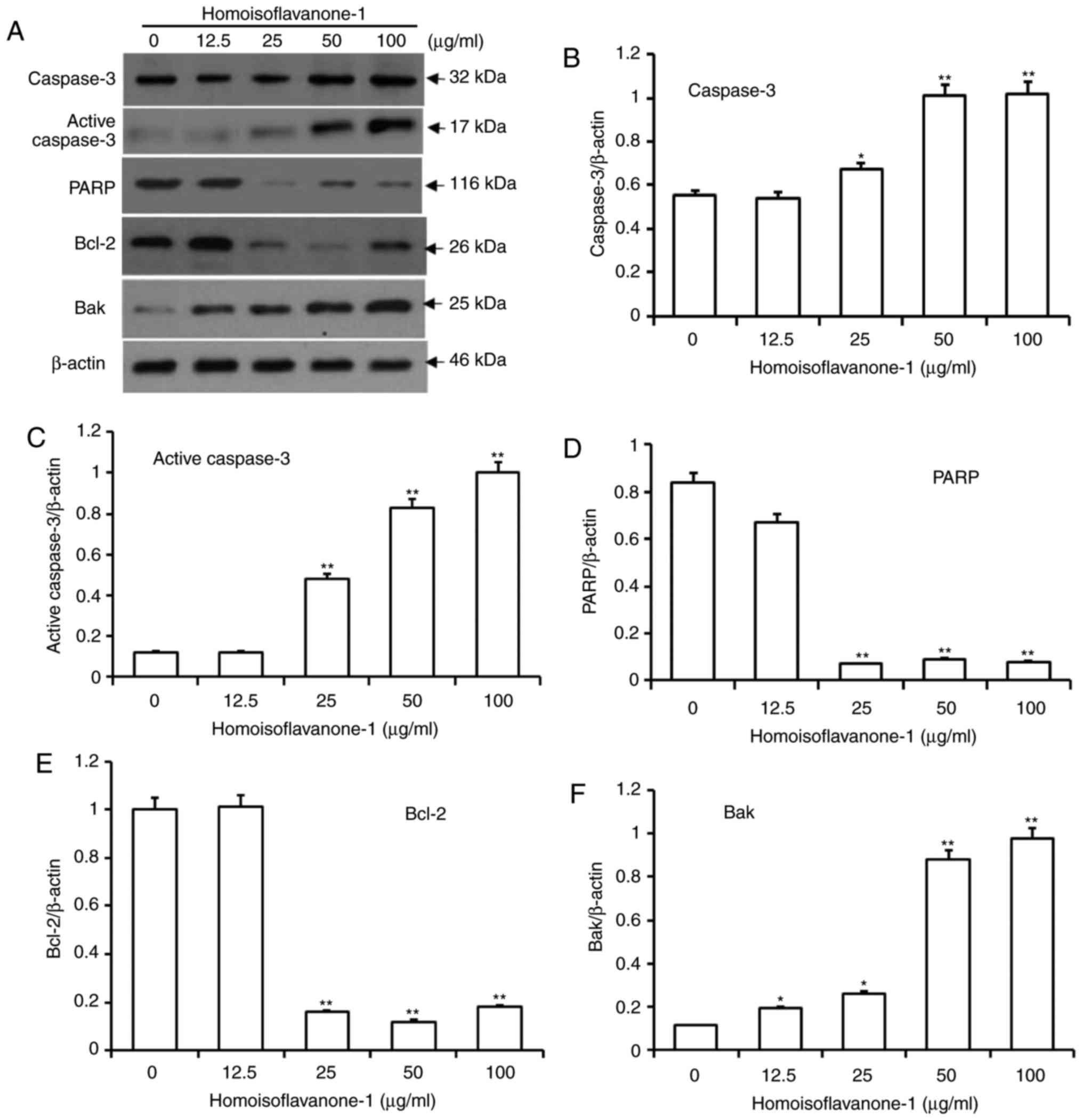
Homoisoflavanone-1 induces intrinsic
mitochondria-mediated apoptosis
As an effector caspase, caspase-3 remains inactive
until apoptotic signaling leads to its cleavage by an initiator
caspase (26). Poly ADP-ribose
polymerase (PARP) is cleaved between Asp214 and Gly215 by the
activated caspase-3, which leads to PARP inactivation and further
facilitates apoptotic cell death. As shown in Fig. 6A-D, the amount of caspase 3 and active
caspase-3 proteins increased dramatically in a dose-dependent
manner after cells were treated with high concentrations of
homoisoflavanone-1 (25, 50 and 100 µg/ml), while the amount of PARP
protein decreased.
Bcl-2 proteins regulate caspase activity, and
function in intrinsic mitochondrion-mediated apoptosis (27). Within the Bcl-2 family, Bcl-2 is
anti-apoptotic, whereas Bak is pro-apoptotic (28). After treatment with various
concentrations of homoisoflavanone-1, the amount of Bcl-2 protein
decreased significantly, whereas the amount of Bak protein
increased in a dose-dependent manner (Fig. 6E and F).
Homoisoflavanone-1 treatment increased
the abundance of er stress-related proteins
To further examine homoisoflavanone-1 induced
apoptosis in A549 cells, we assessed the expressions of the ER
stress-related proteins PERK, ATF4, and GADD34 in
homoisoflavanone-1 treated cells. As shown in Fig. 7A and B, phomoisoflavanone-1 (100
µg/ml) significantly increased the abundance of ER stress-related
proteins in a time-dependent manner. PERK levels peaked 12 h after
homoisoflavanone-1 treatment, while ATF4 and GADD34 reached a peak
at 24 h with the same treatment. Homoisoflavanone-1 also increased
the expression of these three proteins in a dose-dependent manner
at both 12 and 24 h (Fig. 7C-F).
These results suggest that the ER stress pathway functions in
homoisoflavanone-1-induced lung cancer cell apoptosis.
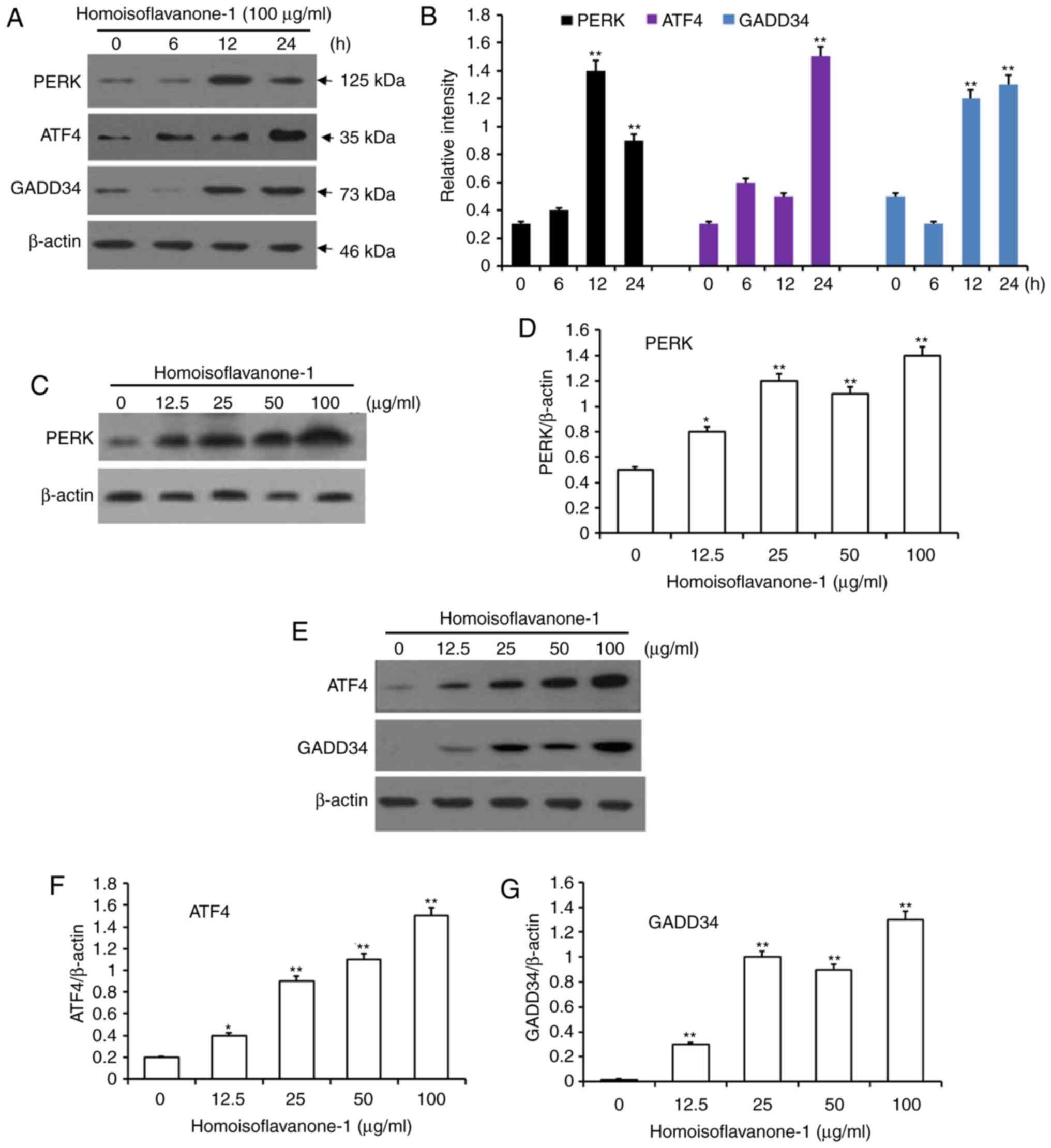 | Figure 7.Accumulation of proteins associated
with the ER stress pathway for apoptosis in response to
homoisoflavanone-1 treatment. A549 cells were treated with
homoisoflavanone-1 (100 µg/ml) for 0, 6, 12 and 24 h. (A) Western
blot analysis was performed to detect the protein expression of (B)
PERK, ATF4 and GADD34. A549 cells were treated with 0, 12.5, 25, 50
or 100 µg/ml of homoisoflavanone-1 for 12 h and (C) western blot
analysis was performed to detect the protein expression of (D)
PERK. A549 cells were treated with 0, 12.5, 25, 50 or 100 µg/ml of
homoisoflavanone-1 for 24 h and (E) western blot analysis was
performed to detect the protein expression of (F) ATF4 and (G)
GADD34. β-actin was the loading control. Data are presented as the
mean ± standard deviation from three independent experiments.
*P<0.05 and **P<0.01 vs. the control group. ER, endoplasmic
reticulum. |
Discussion
Homoisoflavanone-1 is a natural product that can be
purified from P. odoratum, a traditional herbal medicine due
to its anti-hyperglycemic effects (16), procoagulant activity (15), and apoptosis inducing activity
(17,29). In the present study, we demonstrated
that homoisoflavanone-1 exerted anti-cancer activity by reducing
the viability of A549 cells, and that this activity was associated
with cell cycle arrest and apoptosis. Moreover, homoisoflavanone-1
activated the mitochondrion and ER stress apoptotic pathways in
A549 cells, concomitant with changes in the cell cycle and the
expression of apoptosis-related proteins.
Cell cycle progression from G2 to M is regulated by
cyclin B/Cdc2, which is a component of the mitosis-promoting factor
(MPF) (30). To move past the M
phase, the cyclin B/Cdc2 must be inactivated by the cyclin-CDK
complex bound to p21 (31).
Interestingly, p53 is upstream of p21 in this cascade, and
regulates it at the transcriptional level (32). Then, active p38 can induce p53 and p21
activation, at least in part by increasing ROS levels. Upregulated
p38 expression can also promote Cdc2 phosphorylation, which
inhibits the transcription of cyclin B1 and Cdc2 and
therefore reduces the levels of the Cdc2/cyclin B1 complex required
for progression from G2 to M (33).
Our findings of increased levels of P-P38, p38, p53 and p-Cdc2, and
decreased levels of Cdc2, suggest that these proteins are involved
in the homoisoflavanone-1-induced G2/M arrest by activating the
p38-p53 signaling pathway.
As a process in programmed cell death, apoptosis is
necessary for cell growth, development, and homeostasis in
metazoans associated with G2/M arrest (34,35). Three
well-studied pathways initiate apoptosis: the
mitochondrion-mediated intrinsic pathway, the ER stress-induced
pathway, and the death receptor-induced extrinsic pathway (36). In the mitochondria-mediated intrinsic
pathway, apoptosis is mediated primarily by Bcl-2 family proteins
including anti-apoptotic proteins such as Bcl-2 and pro-apoptotic
proteins such as Bak. Altering the balance between Bcl-2 and Bak
can increase permeability of the mitochondrial outer membrane,
leading to cytochrome c release, and ultimately activate caspase
cascades (37,38). In addition, ligands in the extrinsic
pathway induce the caspase-8 initiator protease, which activates
effector proteases such as caspase-3 (39,40).
Active caspase-3 cleaves PARP, a nuclear DNA repair enzyme involved
maintaining genomic integrity, which further facilitates apoptotic
cell death (41,42). In the present study, flow cytometric
analysis showed homoisoflavanone-1 induced apoptosis in A549 cells.
This was further supported by a reduced Bcl-2/Bak ratio, reflecting
by increase in Bak and a decrease in Bcl-2. Together, these data
indicate that homoisoflavanone-1 affects the response to
mitochondrial-mediated apoptosis. Moreover, homoisoflavanone-1
treatment resulted in an increase in active caspase-3 levels and a
decrease in PARP levels, which suggests that homoisoflavanone-1
also induced caspase-associated cell apoptosis.
ER stress-induced cancer cell apoptosis is a
high-profile signaling target for the development of cancer therapy
drugs. When the cell is under reactive oxygen and calcium ion
stress, the ER stress pathway is induced by unfolded or misfolded
protein accumulation in the ER lumen (43). Activated PERK is an important sensor
for ER stress. When ER stress occurs, the downstream signaling
pathway is induced and inhibits protein translation, consequently
restoring ER homeostasis (44). As ER
stress increases and/or time passes, the PERK signaling pathway
activates ATF4 (45), which in turn
drives growth arrest and transcription of DNA damage-inducible
34 (GADD34). Continued expression of GADD34 will then
induce cell death (46). Consistent
with the present study, the protein levels of PERK peaked 12 h
after homoisoflavanone-1 (100 mg/l) treatment, while ATF4 and
GADD34 peaked 24 h after the same treatment. In addition, we
observed an increase in abundance of the ER stress-related proteins
PERK, ATF4, and GADD34 in a dose-dependent manner, indicating that
ER stress was activated in response to homoisoflavanone-1 treatment
in A549 cells.
In conclusion, our study presents the first evidence
of a role for homoisoflavanone-1 in inducing cell cycle arrest and
in promoting apoptosis in NSCLC A549 cells and reveals that
homoisoflavanone-1 accomplishes this by regulating the
mitochondrion-caspase-dependent and ER stress pathways. These
findings suggest that homoisoflavanone-1 extracted from P.
odoratum may function as a tumor suppressor and has potential
as a therapeutic agent to treat lung cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by Scientific
Research Fund of Heilongjiang Provincial Education Department
(grant no. 2016-KYYWF-0859).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DN participated in the study design and all
experimental procedures as well as drafting the manuscript. MJ and
TX and JS performed the experiments. ML performed protein
identification. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kohler BA, Ward E, McCarthy BJ, Schymura
MJ, Ries LA, Eheman C, Jemal A, Anderson RN, Ajani UA and Edwards
BK: Annual report to the nation on the status of cancer, 1975–2007,
featuring tumors of the brain and other nervous system. J Natl
Cancer Inst. 103:714–736. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wallace WA: The challenge of classifying
poorly differentiated tumours in the lung. Histopathology.
54:28–42. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Smith SL, Palma D, Parhar T, Alexander CS
and Wai ES: Inoperable early stage non-small cell lung cancer:
Comorbidity, patterns of care and survival. Lung Cancer. 72:39–44.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao D, Yang G, Meng Q, Liu J and Yang S:
Linobiflavonoid inhibits human lung adenocarcinoma A549 cells:
Effect on tubulin protein. Mol Biol Rep. 40:6019–6025. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fang C, Zhang J, Qi D, Fan X, Luo J, Liu L
and Tan Q: Evodiamine induces G2/M arrest and apoptosis via
mitochondrial and endoplasmic reticulum pathways in H446 and H1688
human small-cell lung cancer cells. PLoS One. 9:e1152042014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Martinvalet D, Zhu P and Lieberman J:
Granzyme A induces caspase-independent mitochondrial damage, a
required first step for apoptosis. Immunity. 22:355–370. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dejean LM, Martinez-Caballero S, Manon S
and Kinnally KW: Regulation of the mitochondrial apoptosis-induced
channel, MAC, by BCL-2 family proteins. Biochim Biophys Acta.
1762:191–201. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scorrano L, Oakes SA, Opferman JT, Cheng
EH, Sorcinelli MD, Pozzan T and Korsmeyer SJ: BAX and BAK
regulation of endoplasmic reticulum Ca2+: A control point for
apoptosis. Science. 300:135–139. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang XZ and Ron D: Stress-induced
phosphorylation and activation of the transcription factor CHOP
(GADD153) by p38 MAP Kinase. Science. 272:1347–1349. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim BJ, Ryu SW and Song BJ: JNK- and p38
kinase-mediated phosphorylation of Bax leads to its activation and
mitochondrial translocation and to apoptosis of human hepatoma
HepG2 cells. J Biol Chem. 281:21256–21265. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hetz C, Bernasconi P, Fisher J, Lee AH,
Bassik MC, Antonsson B, Brandt GS, Iwakoshi NN, Schinzel A,
Glimcher LH and Korsmeyer SJ: Proapoptotic BAX and BAK modulate the
unfolded protein response by a direct interaction with IRE1alpha.
Science. 312:572–576. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takenaka K, Moriguchi T and Nishida E:
Activation of the protein kinase p38 in the spindle assembly
checkpoint and mitotic arrest. Science. 280:599–602. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang H, Chen L, Kou JP, Zhu DN, Qi J and
Yu BY: Steroidal sapogenins and glycosides from the fibrous roots
of Polygonatum odoratum with inhibitory effect on tissue factor
(TF) procoagulant activity. Steroids. 89:1–10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Deng Y, He K, Ye X, Chen X, Huang J, Li X,
Yuan L, Jin Y, Jin Q and Li P: Saponin rich fractions from
Polygonatum odoratum (Mill.) Druce with more potential hypoglycemic
effects. J Ethnopharmacol. 141:228–233. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang Y, Xu HL, Zhang ZT, Liu JJ, Li WW,
Ming H and Bao JK: Characterization, molecular cloning, and in
silico analysis of a novel mannose-binding lectin from Polygonatum
odoratum (Mill.) with anti-HSV-II and apoptosis-inducing
activities. Phytomedicine. 18:748–755. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park S, Hong SM, Ahn IS, Kim YJ and Lee
JB: Huang-Lian-Jie-Du-Tang supplemented with Schisandra chinensis
Baill. and Polygonatum odoratum Druce improved glucose tolerance by
potentiating insulinotropic actions in islets in 90%
pancreatectomized diabetic rats. Biosci Biotechnol Biochem.
73:2384–2392. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang D, Zeng L, Li D and Pu W: Antioxidant
activities of different extracts and homoisoflavanones isolated
from the Polygonatum odoratum. Nat Prod Res. 27:1111–1114. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo H, Zhao H, Kanno Y, Li W, Mu Y, Kuang
X, Inouye Y, Koike K, Jiang H and Bai H: A dihydrochalcone and
several homoisoflavonoids from Polygonatum odoratum are activators
of adenosine monophosphate-activated protein kinase. Bioorg Med
Chem Lett. 23:3137–3139. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bloom J and Cross FR: Multiple levels of
cyclin specificity in cell-cycle control. Nat Rev Mol Cell Biol.
8:149–160. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nebreda AR and Porras A: p38 MAP kinases:
Beyond the stress response. Trends Biochem Sci. 25:257–260. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
El-Deiry WS: The role of p53 in
chemosensitivity and radiosensitivity. Oncogene. 22:7486–7495.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Malumbres M: Physiological relevance of
cell cycle kinases. Physiol Rev. 91:973–1007. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Morgan DO: Cyclin-dependent kinases:
Engines, clocks, and microprocessors. Annu Rev Cell Dev Biol.
13:261–291. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Walters J, Pop C, Scott FL, Drag M, Swartz
P, Mattos C, Salvesen GS and Clark AC: A constitutively active and
uninhibitable caspase-3 zymogen efficiently induces apoptosis.
Biochem J. 424:335–345. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Youle RJ and Strasser A: The BCL-2 protein
family: Opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Harris MH and Thompson CB: The role of the
Bcl-2 family in the regulation of outer mitochondrial membrane
permeability. Cell Death Differ. 7:1182–1191. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ouyang L, Chen Y, Wang XY, Lu RF, Zhang
SY, Tian M, Xie T, Liu B and He G: Polygonatum odoratum lectin
induces apoptosis and autophagy via targeting EGFR-mediated
Ras-Raf-MEK-ERK pathway in human MCF-7 breast cancer cells.
Phytomedicine. 21:1658–1665. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Arion D, Meijer L, Brizuela L and Beach D:
cdc2 is a component of the M phase-specific histone H1 kinase:
Evidence for identity with MPF. Cell. 55:371–378. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bloom J and Pagano M: To be or not to be
ubiquitinated? Cell Cycle. 3:138–140. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bates S, Ryan KM, Phillips AC and Vousden
KH: Cell cycle arrest and DNA endoreduplication following
p21Waf1/Cip1 expression. Oncogene. 17:1691–1703. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kang N, Jian JF, Cao SJ, Zhang Q, Mao YW,
Huang YY, Peng YF, Qiu F and Gao XM: Physalin A induces G2/M phase
cell cycle arrest in human non-small cell lung cancer cells:
Involvement of the p38 MAPK/ROS pathway. Mol Cell Biochem.
415:145–155. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fuchs Y and Steller H: Programmed cell
death in animal development and disease. Cell. 147:742–758. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Green DR, Galluzzi L and Kroemer G: Cell
biology. Metabolic control of cell death. Science. 345:12502562014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Beere HM: Death versus survival:
Functional interaction between the apoptotic and stress-inducible
heat shock protein pathways. J Clin Invest. 115:2633–2639. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang C and Youle RJ: The role of
mitochondria in apoptosis*. Annu Rev Genet. 43:95–118. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang F, Kong DS, Zhang ZL, Lei N, Zhu XJ,
Zhang XP, Chen L, Lu Y and Zheng SZ: Tetramethylpyrazine induces
G0/G1 cell cycle arrest and stimulates mitochondrial-mediated and
caspase-dependent apoptosis through modulating ERK/p53 signaling in
hepatic stellate cells in vitro. Apoptosis. 18:135–149.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cardile V, Musumeci G, Sicurezza E, Caggia
S, Rusu MC, Leonardi R and Loreto C: Expression of TRAIL and its
receptors DR5 and DcR2 in orthodontic tooth movement. Histol
Histopathol. 28:933–940. 2013.PubMed/NCBI
|
|
40
|
Musumeci G, Loreto C, Leonardi R,
Castorina S, Giunta S, Carnazza ML, Trovato FM, Pichler K and
Weinberg AM: The effects of physical activity on apoptosis and
lubricin expression in articular cartilage in rats with
glucocorticoid-induced osteoporosis. J Bone Miner Metab.
31:274–284. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shakibaei M, Csaki C, Nebrich S and
Mobasheri A: Resveratrol suppresses interleukin-1beta-induced
inflammatory signaling and apoptosis in human articular
chondrocytes: Potential for use as a novel nutraceutical for the
treatment of osteoarthritis. Biochem Pharmacol. 76:1426–1439. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Virág L and Szabó C: The therapeutic
potential of poly(ADP-ribose) polymerase inhibitors. Pharmacol Rev.
54:375–429. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Boyce M and Yuan J: Cellular response to
endoplasmic reticulum stress: A matter of life or death. Cell Death
Differ. 13:363–373. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang J, Hu X and Jiang H: ERS-PERK
signaling pathwaymediated Nrf2/ARE-HO-1 axis: A novel therapeutic
target for attenuating myocardial ischemia and reperfusion injury.
Int J Cardiol. 203:779–780. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Harding HP, Novoa I, Zhang Y, Zeng H, Wek
R, Schapira M and Ron D: Regulated translation initiation controls
stress-induced gene expression in mammalian cells. Mol Cell.
6:1099–1108. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sano R and Reed JC: ER stress-induced cell
death mechanisms. Biochim Biophys Acta. 1833:3460–3470. 2013.
View Article : Google Scholar : PubMed/NCBI
|