Introduction
Leukemia makes up about 1/3 of the case of cancer in
people younger than 15 years old and is not uncommon in adults
(1). Despite significant advances in
the chemotherapeutic management of leukemia, post-remission relapse
occur frequently in most patients (2). In this way, it is imperative to find
more efficient low-toxicity agents to use in chemo-preventive
therapies for leukemia.
Ailanthone, which is extracted from traditional
Chinese medicinal plant Ailanthus altissima (3), has been well-demonstrated to have
anti-tumor, anti-HIV, anti-inflammatory, anti-malarial,
anti-allergic, and anti-microbial activities (4,5). The
growth-inhibitory effect of ailanthone on varieties of tumor cells
(He La, Jurkat, Hep G2, Hep3B, R-Hep G2, MCF-7, MDA-MB-231, Huh7
and A549) in vitro has been reported (6–9). However,
the cytotoxicity of ailanthone on human HL-60 leukemia cells and
its underlying molecular mechanism are poorly understood.
Apoptosis, type I programmed cell death, is a series
of physiological changes that are mediated by genes and proteins,
and the cells depend on this mechanism to activate their own
destruction. If pathological interference take place during the
process of apoptosis, malignant tumors may form (10–12).
Autophagy, type II programmed cell death, is a conserved
decomposition process that allows the degradation and recycling of
cytoplasm, aggregated proteins, and excess or defective organelles
(13). Autophagy is mainly a response
to the stress of irradiation (14),
chemotherapeutic drugs (15), or
starvation (16). Despite its
contribution to cell survival, previous studies have demonstrated
that several anti-tumor agents induce cell death with autophagic
features in various cancer cells (17–19).
In the present study, we sought to investigate the
cytotoxicity of ailanthone in human HL-60 leukemia cells
in vitro and to elucidate the mechanisms that may underlie
its actions. We are searching for a new natural anti-tumor drug
that is efficient and has minimal toxicity.
Materials and methods
Materials
The pure ailanthone used in this study was extracted
from Ailanthus altissima. The ailanthone sample (purity
>98%) was provided by the Institute of Traditional Chinese
Medicine and Natural Products, Jinan University (Guangzhou, China).
Dimethyl sulfoxide (DMSO) was purchased from Shanghai Joey Chemical
Reagent Co., Ltd., (Shanghai, China).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazoliumbromide (MTT)
was purchased from Amresco, LLC, (Solon, OH, USA). Propidium iodide
(PI) and acridine orange (AO) was purchased from Nanjing Key Gen
Biotech Co., Ltd., (Nanjing, China) RPMI-1640 medium was purchased
from Gibco; Thermo Fisher Scientific, Inc., (Waltham, MA, USA).
Fetal bovine serum (FBS), penicillin/streptomycin solution (PS),
and antibodies for LC3I/II, beclin-1, p62, GAPDH were obtained from
Nanjing KeyGen Biotech Co., Ltd. Autophagy inhibitor Bafilomycin-A1
(BaF-A1) was purchased from Boster Biological Technology Co., Ltd.,
(Wuhan, China).
Cell line and culture
Human promyelocytic leukemia HL-60 cells were
obtained from Nanjing Key Gen Biotech Co., Ltd. HL-60 cells were
cultured in RPMI-1640 medium with 10% FBS and 1% PS solution in a
humidifying thermostat with 5% CO2 at 37°C. Stock
solutions of ailanthone were prepared in DMSO and stored at −20°C
and diluted to the required concentration with RPMI-1640 complete
medium.
MTT assay of cell viability
The cytotoxicity of ailanthone in human HL-60 cells
was assessed by using the MTT assays. Exponentially growing HL-60
cells (5×104 cells/well) were seeded and cultured in
96-well plates for 24 h and then treated with various
concentrations of ailanthone (1.25, 2.5, 5, 10, and 20 µM) or 0.1%
DMSO (control group) for 24, 48 and 72 h at 37°C. 20 µl MTT (5
mg/ml) was added to each well of the plate and then cultured for 4
h at 37°C. Subsequently, cells were washed in PBS followed by the
addition of 150 µl DMSO. Micro-plate spectrophotometer (RT-6000;
Rayto Life and Analytical Sciences Co., Ltd., Shenzhen, China) was
used to measure the optical densities at 490 nm spectral
wavelength.
Flow cytometric analysis of cell
apoptosis
HL-60 cells (5×105 cells/well) were
seeded into 6-well plates and cultured for 24 h at 37°C, and were
then treated with various concentrations of ailanthone (5, 10, and
20 µM) or 0.1% DMSO (control group) for 48 h at 37°C, respectively.
Cells were washed twice in cold PBS, and then the cell pellets were
mixed with annexin V APC/7-ADD (Nanjing Key Gen Biotech Co., Ltd.)
for 15 min at 37°C in the dark. Cell apoptosis was were evaluated
using a FACSCalibur Flow Cytometer (BD Biosciences, Franklin Lakes,
NJ, USA) and the rate of cell death was analyzed using a
FACSCalibur internal software system (BD Biosciences).
Flow cytometric analysis of cell
cycle
HL-60 cells (5×105 cells/well) were
seeded into 6-well plates and cultured for 24 h at 37°C, and then
treated with various concentrations of ailanthone (5, 10, and 20
µM) or 0.1% DMSO (control group) for 48 h at 37°C, respectively.
Cells were fixed and rendered permeable with 70% cold ethanol at
4°C. After this interval, cells were treated with 1% RNase and
stained with PI solution for 30 min at 4°C. Cell cycle phase
distribution of cells was established by FACSCalibur flow cytometer
(BD Biosciences) using the cell cycle analysis software (FlowJo
LLC, Ashland, OR, USA).
Detection of autophagy by AO
staining
AO staining was used to detect the presence of
acidic vesicular organelles (AVOs) after ailanthone treatment.
HL-60 cells (5×105 cells/well) were treated with
ailanthone at concentrations of 10 µM or 0.1% DMSO (control group)
for 48 h at 37°C, and then cells were washed twice in cold PBS and
fixed with 4% formaldehyde for 10 min. After the treatment, cells
were stained with 5 µg/ml AO solution for 15 min at 37°C in the
dark and observed using an inverted fluorescence microscope
(OlympusIX5; Olympus Corporation, Tokyo, Japan).
Western blotting
HL-60 cells (5×106 cells/well) were
washed twice in cold PBS and suspended in radio
immune-precipitation assay lysis buffer (Nanjing Key Gen Biotech
Co., Ltd.) on ice for 30 min, then treated with various
concentrations of ailanthone (5, 10, and 20 µM) or 0.1% DMSO
(control group) for 48 h at 37°C, respectively. The lysates were
then cleared by centrifugation at 14,000 × g for 15 min at 4°C.
Subsequently, a bicinchoninic acid protein assay kit (Nanjing Key
Gen Biotech Co., Ltd.) was used to measure the total protein
concentration of each sample. The Protein samples (30 µg) were
separated by 15% SDS-PAGE in each group and then transferred onto
polyvinylidene difluoride membranes (Pall Life Sciences, Port
Washington, NY, USA). Membranes were stuck with 5% (w/v) non-fat
dry milk dissolved in TBS containing 0.05% Tween-20 (TBST) at room
temperature for 1 h, and then washed three times with TBST.
Subsequently, membranes were incubated with primary antibodies
against beclin-1 (1:200), p62 (1:200), LC3I/II (1:200), and GAPDH
(1:400) for 12 h at 4°C. The members were washed three times with
TBST, and then incubated with HRP-conjugated goat anti-rabbit
(1:2,000) IgG (Nanjing Key Gen Biotech Co., Ltd.) secondary
antibodies at room temperature for 1 h. The immune-reactive bands
were visualized with enhanced chemiluminescent substrates (Thermo
Fisher Scientific, Inc.) using an X-ray film processor (Kodak,
Rochester, NY, USA). GAPDH served as a loading control. The
experiment was independently repeated three times, and Quantity One
software (v.4.6.2; Bio-Rad Laboratories, Inc., Hercules, CA, USA)
was used to analyze the densitometry of each band.
Statistical analysis
Data are here expressed as means ± SD. With SPSS
software for windows v.17.0 (SPSS, Inc., Chicago, IL, USA), one way
ANOVA followed by Newman-Keuls multiple comparison test was used
for analysis of variance and the student's t-test was used for
pair-wise comparison. P<0.05 was considered to indicate a
statistically significant difference.
Results
Ailanthone inhibits cell proliferation
in HL-60 cells
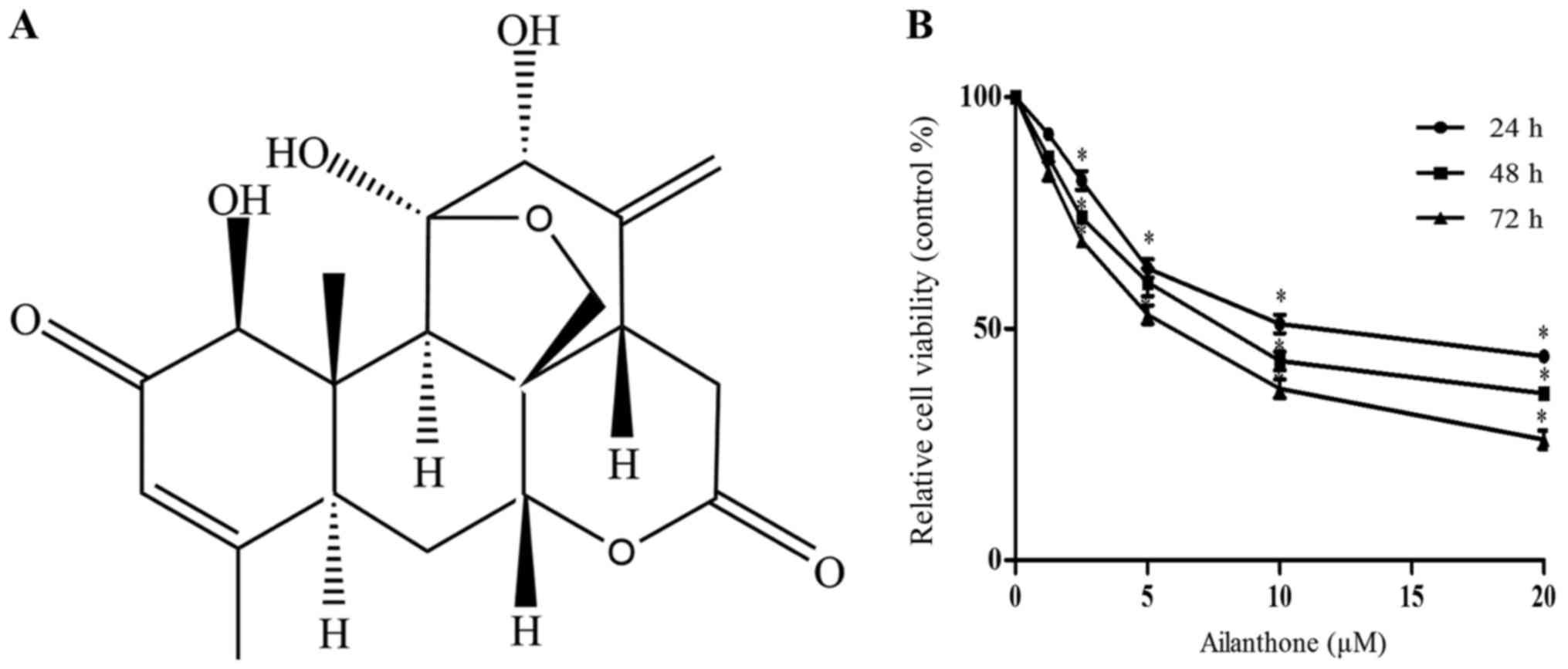
Ailanthone share its chemical structure with the
triterpenoids. Triterpenoids have many physiological activities,
including anti-tumor activities (Fig.
1A). The MTT assay was used to assess the cytotoxicity of
ailanthone in HL-60 cells. Treatment with ailanthone reduced the
viability of HL-60 cells in a dose- and time-dependent manner
(Fig. 1B), with half maximal
inhibitory concentration (IC50) values of 12.18, 8.497,
and 5.986 µM after 24, 48, and 72 h of treatment, respectively.
Ailanthone induces apoptosis and cell
cycle arrest in HL-60 cells
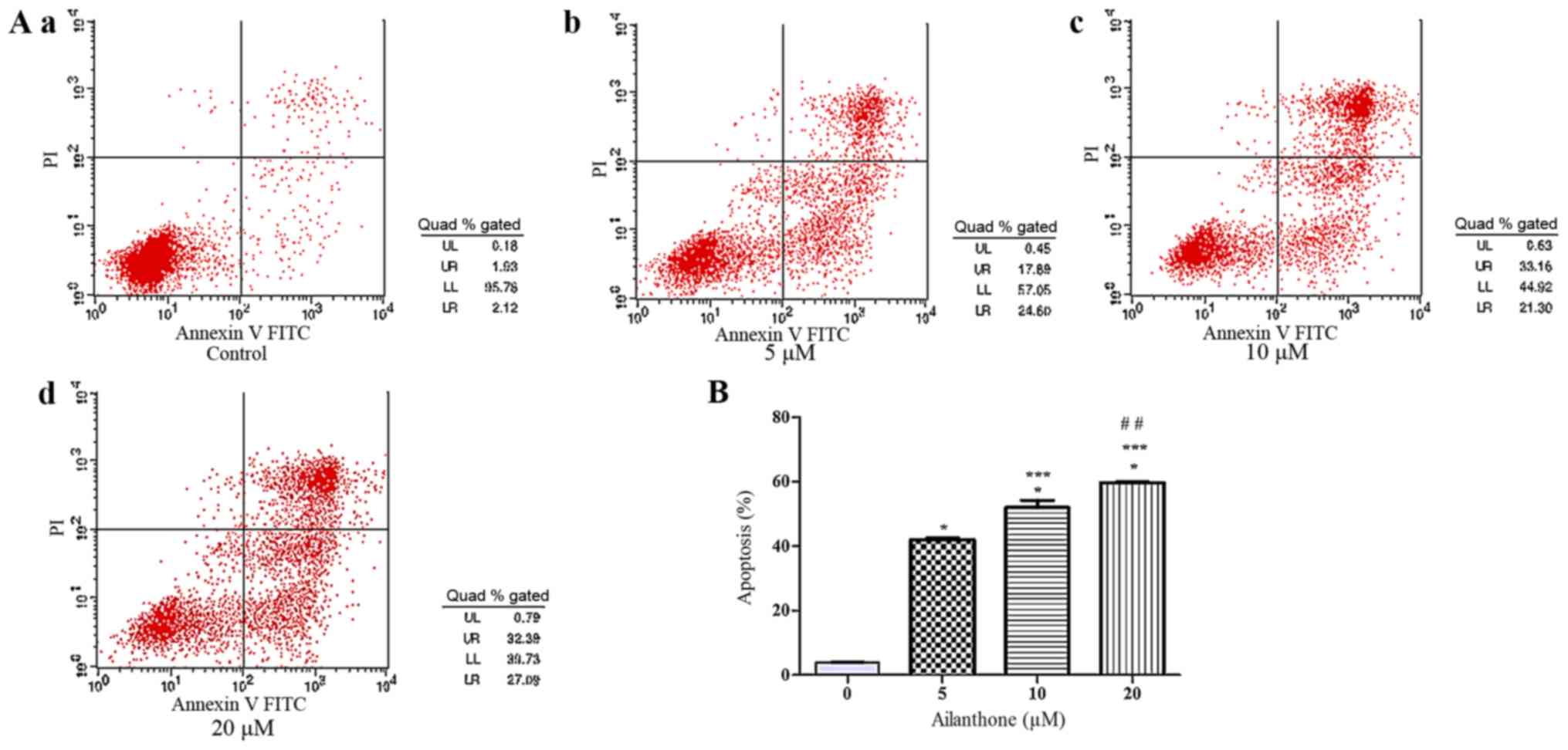
An annexin V-APC/7-ADD staining assay was used to
confirm that ailanthone induced apoptosis in HL-60 cells. In the
right quadrants of flow cytometry graphs, ailanthone significantly
increased the number of apoptotic cells (Fig. 2A). After treatment with 5, 10, and 20
µM ailanthone for 48 h at 37°C, the rate of apoptosis among the
cells was increased significantly from 42.02 to 59.68% and the
percentage of apoptotic cells was 42.02±0.54, 52.05±2.27 and
59.69±0.25%, which was significantly higher than in the control
group (3.92±0.14%; P<0.05; Fig.
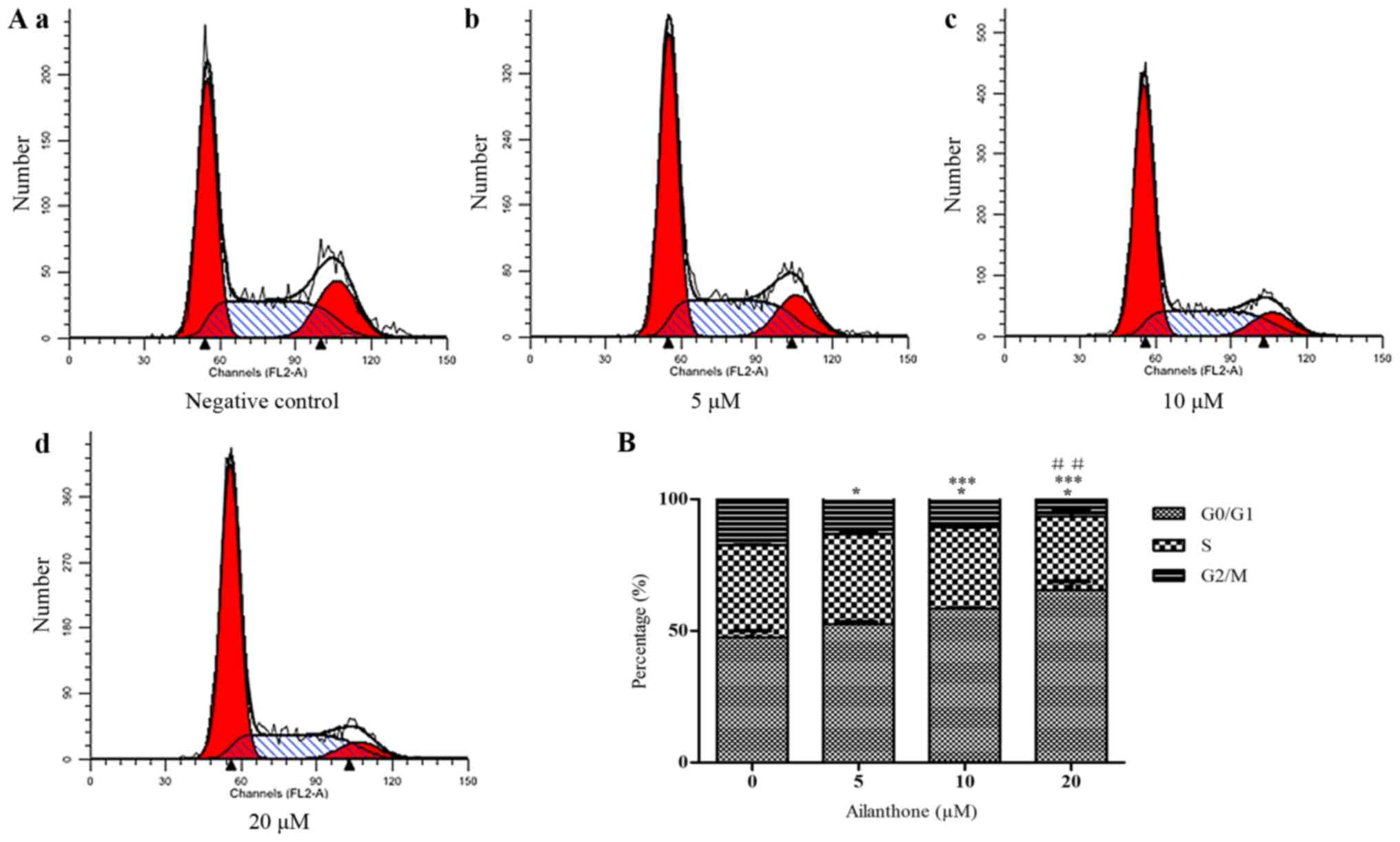
2B). In order to determine whether the anti-proliferative
effect of ailanthone on HL-60 cells was attributable to cell cycle
arrest, we measured DNA content as an indicator of the cell cycle
phase ratio by flow cytometry with PI staining (Fig. 3A). The percentage of G0/G1 cells
increased significantly in a dose-dependent manner and cells in
G0/G1 phase was 53.54±0.88, 58.42±0.31, and 65.57±3.16% in the 5,
10, and 20 µM groups, respectively. Ailanthone treatment induced
significant G0/G1-phase accumulation not observed in the control
group (47.5±2.54%; P<0.05; Fig.
3B).
Ailanthone induces autophagy in HL-60
cells
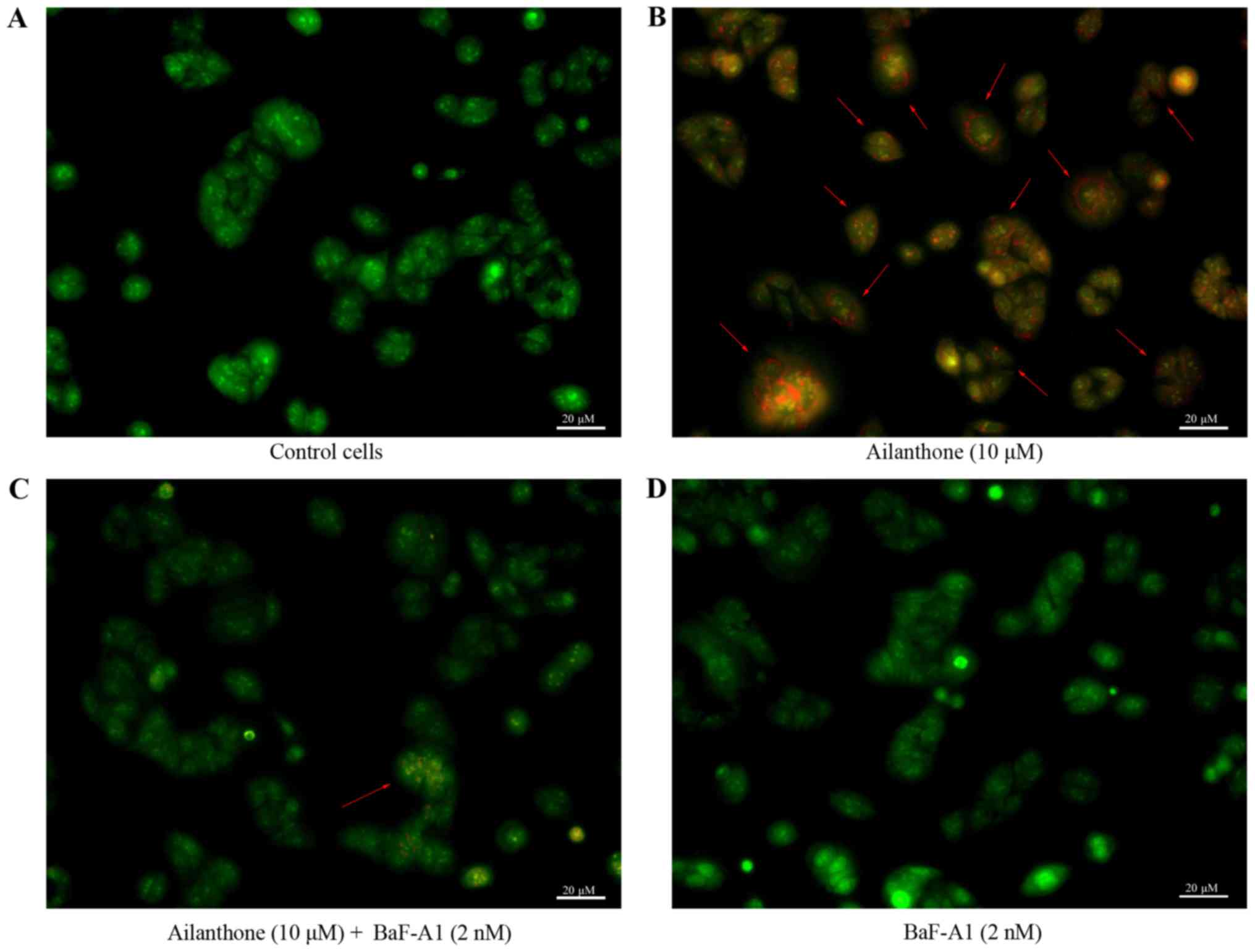
AO staining was used to observe the formation of
acidic vesicular organelles, the main characteristics of autophagy
(Fig. 4). Ailanthone induced
noticeable formation of AVOs, which displayed red fluorescence in
the lysosomal compartments of HL-60 cells (Fig. 4B). Control cells treated with 1% DMSO
displayed green fluorescence in the cytosolic and nuclear
compartments, indicating the lack of AVOs (Fig. 4A). Those results showed that AVOs to
be present in ailanthone-treated HL-60 cells, which suggest that
ailanthone may induce autophagy in HL-60 cells. In addition,
pretreatment with BaF-A1 displayed sporadic red fluorescence in the
cells suggests that ailanthone-induced formation of AVOs in HL-60
cells were attenuated (Fig. 4C).
Pretreatment with BaF-A1 without ailanthone also did not indicate
the formation of AVOs (Fig. 4D). To
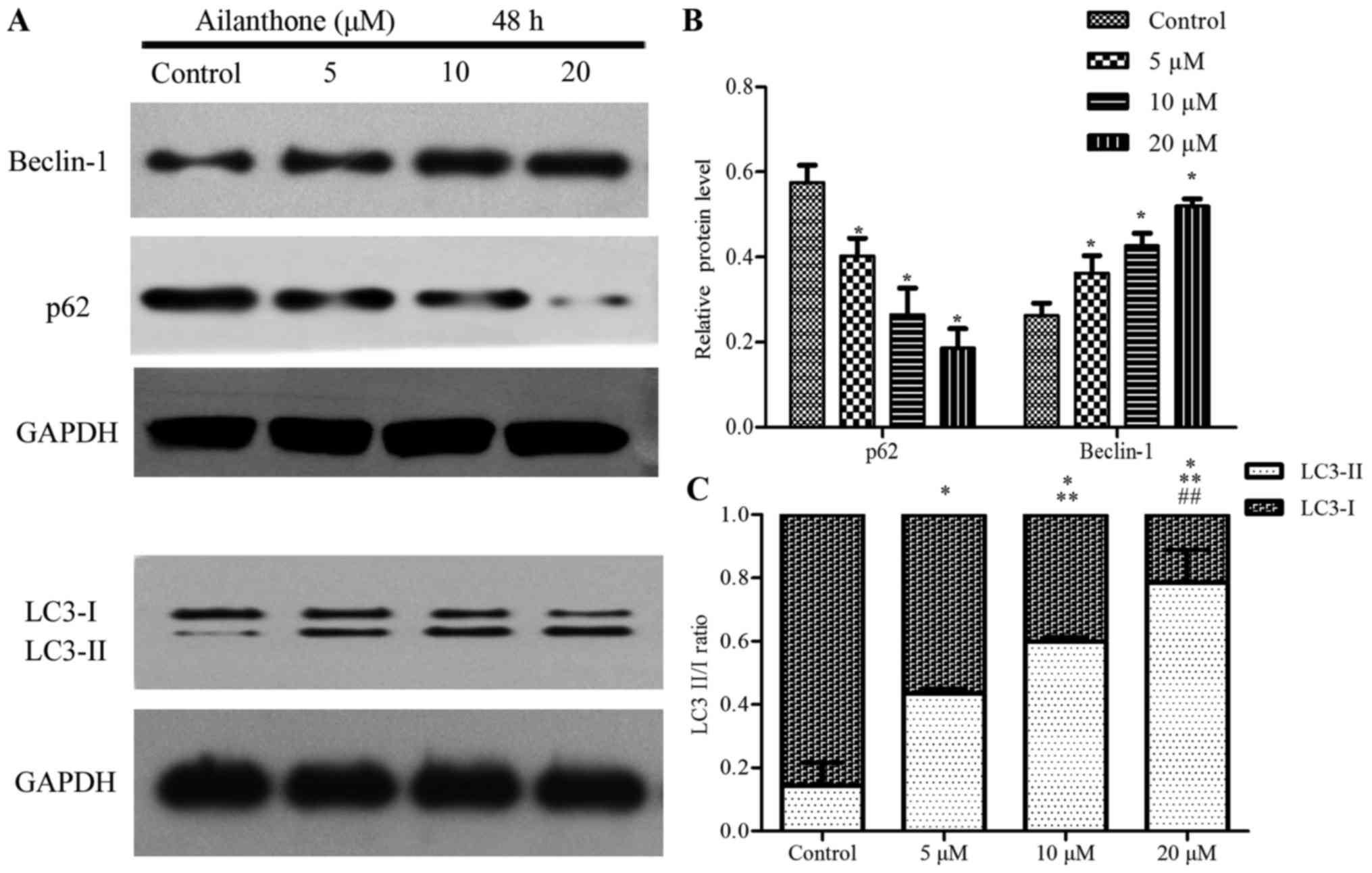
investigate the effect of ailanthone on the protein expression
levels in HL-60 cells that were associated with autophagy, we
performed Western blot analysis to measure the levels of beclin-1,
p62, and LC3-I/II (Fig. 5A).
Treatment with ailanthone markedly increased beclin-1 levels but
decreased and p62 levels (Fig. 5B).
The ratio of LC3-I/II showed that treatment with ailanthone
markedly increased LC3-II levels but decreased and LC3-I levels
(Fig. 5C).
Autophagy is associated with
ailanthone-mediated apoptosis of HL-60 cells
To investigate the role of autophagy in
ailanthone-induced apoptosis, autophagy inhibitor BaF-A1 (1 or 2
nM) was used. Treatment with ailanthone significantly suppressed
the viability of HL-60 cells, and the suppression was significantly
attenuated by BaF-A1 (Fig. 6A). The
rate of apoptotic in HL-60 cells was conspicuously reduced by
BaF-A1 pretreatment (Fig. 6B and C).
After treatment with 0 and 10 µM ailanthone for 48 h at 37°C, the
rate of apoptosis among the cells pretreated with 2 nM BaF-A1 was
increased from 5.77±0.25 to 26.93±1.74%, which were significantly
lower than in the group without pretreated with BaF-A1 (from
3.92±0.14 to 52.05±2.27%; P<0.05′). Those results suggested that
the anti-proliferative effects of ailanthone on HL-60 cells may be
partially due to the induction of autophagy-mediate apoptosis.
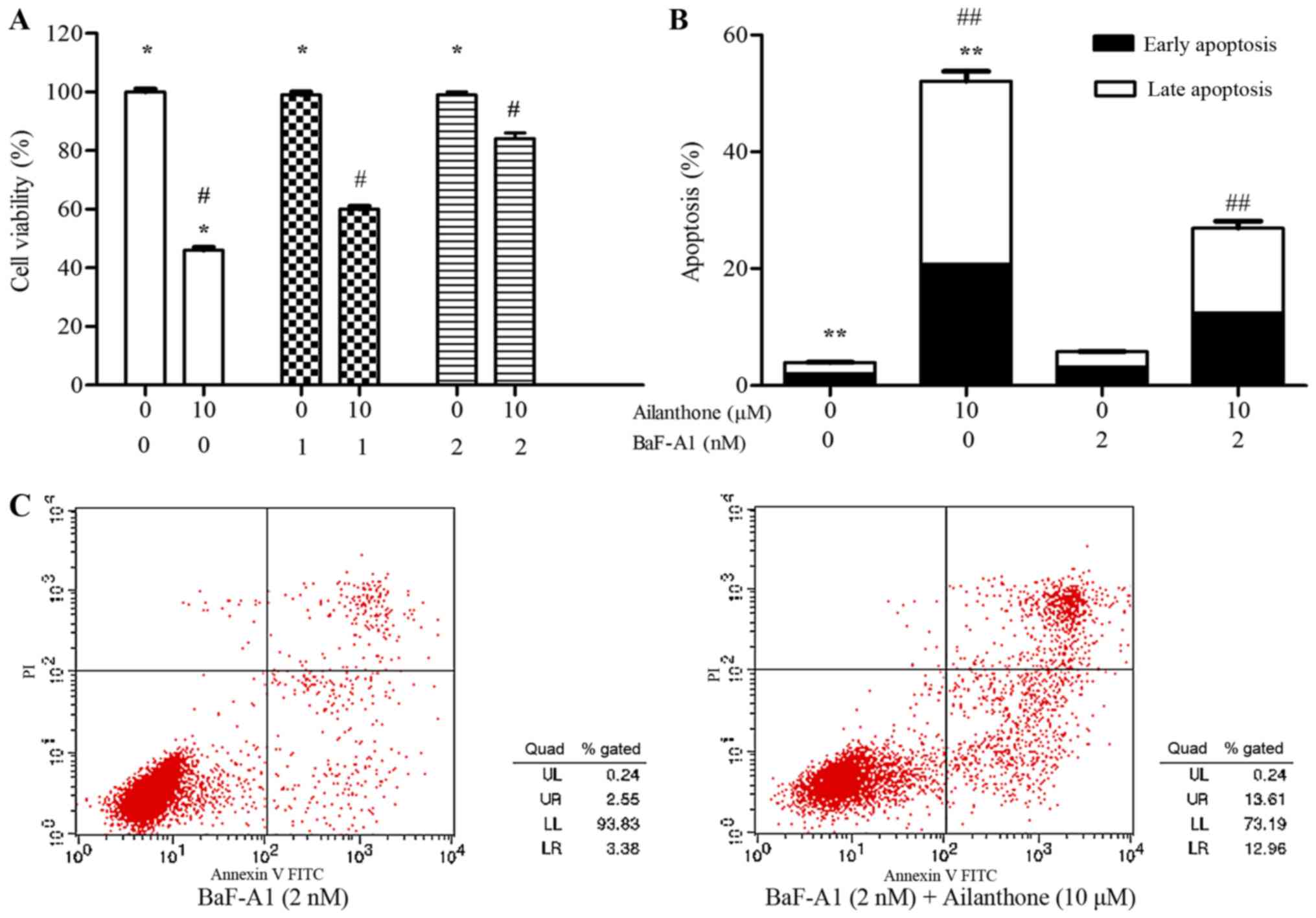 | Figure 6.Autophagy is associated with
ailanthone-mediated apoptosis of HL-60 cells. (A) Cells were
pretreated with and without BaF-A1 (1 or 2 nM) for 30 min,
following treatment with and without ailanthone (10 µM) for 48 h at
37°C. MTT assay was used to determine the cell viability. (B) Cells
were pretreated with and without BaF-A1 (2 nM) for 30 min,
following by treatment with and without ailanthone (10 µM) for 48 h
at 37°C, (C) annexin V-FITC staining assay was used to analyze the
cell apoptosis, ANOVA using Newman-Keuls multiple comparison test.
*P<0.05 vs. control, #P<0.05 vs. (0, 10 µM)
groups, **P<0.05 vs. control, ##P<0.05 vs. (0, 10
µM) groups. MTT,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazoliumbromide. |
Discussion
Ailanthone, which is extracted from the traditional
Chinese medicinal plant Ailanthus altissima (3), has been thoroughly demonstrated to have
anticancer activity in previous studies (4–9). However,
the anti-proliferative effects of ailanthone on HL-60 cells and
mechanisms that may underlie these effects have been poorly
understood. The present study is the first to demonstrate the
potent-cytotoxicity of ailanthone against HL-60 cells. The
mechanisms that underlie these effects may involve induction of
autophagic cell death in HL-60 cells.
In the present study, we found that the rate of
apoptosis in ailanthone-treated HL-60 cells to increase in a
dose-dependent manner and this effect may be associated with an
increase in the number of cells arrested at the G0/G1 phase. The
cell cycle is the overall process of the cell from the beginning of
a division to the end of the next division that allows the cell to
proliferate, and is divided into four consecutive phases, known as
G0/G1, S, and G2/M phases (20).
Although G0 is often referred to as the quiescent phase, G0-phase
cells are still quite with respect to cellular growth and are
strictly regulated to determine when the cell will enter other
stages of the cell cycle (21). The
mitogenic signaling mediated by the RAS/RAF/MAPK pathway promotes
this shift, whose endpoint is the stimulation of D-type cyclin
production (22). Our study suggested
that ailanthone induces cell cycle arrest of HL-60 cells at the
G0/G1 phase. In addition, a previous study found that ailanthone
significantly induced cell cycle arrest at the G1/S phase in Huh-7
hepatocellular carcinoma cells (9).
In eukaryotic cells, the process of G2/M cell cycle is regulated by
protein B (cyclinB)-p34cdc2, G2/M phase arrest takes place mainly
in a p53-dependent manner. Watson et al (23), investigated a p53 wild-type MCF-7 cell
line and p53 mutated MDA-MB-231 cell line. They found that the cell
cycle of p53 wild-type MCF-7 cell line was arrested at the G1 and
G2 phases under ionizing radiation, while p53 mutated MDA-MB-231
cells were arrested at the G2/M phase. Drug-induced cell cycle
modulation not only varies between the same cell line treated with
different drugs, but also in different cells after treatment with
the same drug. Lavhale et al (24), found ailanthus excelsa chloroform
extract-1 extracted from Ailanthus excels could induce
S/G2-M cell cycle arrest in MDA-MB-231, MCF-7, and PC3 cells and a
G1 arrest in B16F10 cells. Treatment with AECHL-1 results in a
significant decrease in the levels of c-Myc, CDK-4, and cyclin D1
in B16F10 cells, while the expression level of p21 was increased.
p21 forms a complex with CDK2/CDK4/CDK6 and inhibits the CDK-cyclin
kinase activity phase and arrests cells in the G1 phase (24). Therefore, we believe that the
induction of cell cycle arrest at different phase by compounds
extracted from ailanthus may be associated with a variety of
factors, such as mutation of p53 gene and expression level of p21
in tumor cells and so on. In conclusion, these results demonstrated
that the anti-proliferative effects of ailanthone on HL-60 cells
were partially due to the induction of apoptosis and G0/G1 phase
cell cycle arrest.
A previous study showed that some anti-cancer
chemotherapy drugs can induce autophagic apoptosis in malignant
tumor cells, thus inhibiting the proliferation of tumor cells
(25,26). In autophagy, targeted cytoplasm
constituents are isolated from other parts of the cell, which forms
a double membrane called autophagosome. Then, the autophagosome
enters the lysosome through the cytoplasm, and the two organelles
fuse. In the lysosome, the contents of the autophagosome are
degraded by acidic lysosome hydrolase (27,28). Our
experiment indicated the presence of acidic vesicular organelles in
HL-60 cells after treatment with ailanthone by AO staining, which
suggested that ailanthone may induce HL-60 cells autophagy.
Bafilomycin-A1 is a V-type ATPase inhibitor, which can prevent
acidification and alters the membrane potential of some specific
layers. Treatment with Baf-A1 will eventually lead to block in
fusion of autophagosomes with lysosomes, thus preventing the
maturity of autophagy (29,30). There are several autophagy inhibitory
reagents different from Baf-A1, such as E64d/pepstatin-A or
chloroquine. E-64d is a membrane-permeable cysteine protease
inhibitor, which can block the activity of a subset of lysosomal
hydrolases. Pepstatin-A is an aspartyl protease inhibitor to
inhibit lysosomal protein degradation. E-64d should be used in
combination with pepstatin-A to inhibit lysosomal protein
degradation (31). Chloroquine is
lysosomal compounds, and elevate/neutralize the lysosomal/vacuolar
pH. Chloroquine improve the lysosomal pH value and the ultimately
inhibition of autophagosomes and lysosome fusion, thereby
preventing autophagosome maturation of autophagosomes into
autoly-sosomes (32). Using
E64d/pepstatin-A or chloroquine may better interpret the autophagy
flux. However, our experiment refers whether or not autophagy is
activated. Our currently study using Baf-A1 could clear to see the
formations of autophagic vacuoles were suppressed under
microscope.
We then investigated ailanthone-induced autophagy in
HL-60 cells. The autophagy levels of the cells were found to be
very low in a physiological environment; the major function of
autophagy is degradation and recycling of senescent organelles and
longevity proteins, which allow the cells to live longer (14–16). The
role of autophagy in cells is paradoxical and when cells are faced
with physiological or pathological stresses, the autophagy levels
can increase significantly, which can lead to cell death (33,34).
Autophagy is associated with molecular regulation of autophagy
associated protein, such as LC3, beclin-1, and p62. LC3 is an
important protein in the autophagy process, during which
cytoplasmic pattern LC3 (LC3-I) is converted to autophagosomal
membrane LC3 (LC3-II), resulting in elevated LC3-II levels
(35). The beclin-1 gene is located
on chromosome 17q21, which is homologous with the yeast autophagy
gene Apg6/Vps30 and involves in the composition of the class III
PI3K complex (36). Beclin-1 is a
critical protein in the formation of autophagosome encoded by
beclin-1 gene. Beclin-1 may mediate cell apoptosis by binding to
the apoptosis-associated protein bcl-2 (37). p62 is a multifunctional ubiquitin
protein, participating in the ubiquitin-proteasome system and the
autophage system. With the occurrence of autophagy, p62 protein is
incorporated into autophagosomes and then degraded (38). We found that ailanthone increased
beclin-1 and LC3-II levels but decreased and p62 levels in a
dose-dependent manner. Those results indicated that beclin-1, p62,
and LC3 may participate in ailanthone inducted-autophagy in HL-60
cells. A recent paper found that ailanthone inhibits the
proliferation activity of vestibular schwannoma cells by blocking
the Ras/Ras/Raf/MEK/ERK and mTOR pathways through down-regulating
the expression of mir-21. They observed that ailanthone reduced
CyclinD1 protein levels, indicating that ailanthone may block
vestibular schwannom cells in the G1 phase of cell cycle.
Furthermore, their experiment also found that ailanthone enhanced
the protein expression of LC3-II and Beclin-1, reduced the
expression of p62, indicating that ailanthone may induce vestibular
schwannom cells autophagy (39).
Unlike this, our study is the first to demonstrate the
potent-cytotoxicity of ailanthone against HL-60 cells. We observed
the formation of acidic vesicular organelles by AO staining,
visually see the characteristic performance of autophagy. In
addition, we investigated whether ailanthone-induced autophagy is
associated with apoptotic cell death in HL-60 cells. The MTT assay
and annexin V-FITC staining assay suggested that the
anti-proliferative and pro-apoptotic effects of ailanthone on HL-60
cells were significantly reversed following pretreatment of
autophagy inhibitor BaF-A1. These results suggested that
ailanthone-induced autophagy may play a pivotal role in apoptotic
cell death in human promyelocytic leukemia cells.
In summary, the present study is the first to
demonstrate that ailanthone extracted from Ailanthus
altissima has anti-proliferative effects on HL-60 cells in
vitro. We further found that these effects were partially due
to the induction of apoptosis and G0/G1 phase cell cycle arrest. We
found that ailanthone induces HL-60 cell autophagy possibly through
the modulation of beclin-1, p62, and LC3 proteins expression. In
addition, we found that the anti-proliferative and pro-apoptotic
effects of ailanthone on HL-60 cells were significantly reversed by
inhibiting autophagy. These results suggest that ailanthone-induced
autophagy in the HL-60 cells likely involves autophagic cell death.
However, considering the potential fluctuation of p62 and LC3-II
levels during the process, the absence of data at other time points
besides 48 h would be a limitation of the present study. Further
studies are need to determine the details of the mechanism
underlying autophagic and apoptotic cell death induced by
ailanthone in leukemia cells, and to evaluate the
anti-proliferative effects of ailanthone on leukemia cells in
vivo. Our results revealed the pharmacological activity of
ailanthone on HL-60 cells and suggest that ailanthone could be a
suitable therapeutic agent for the treatment of leukemia.
Acknowledgements
The authors would like to thank the Institute of
Traditional Chinese Medicine and Natural Products, Jinan University
for providing the pure sample of ailanthone.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81241102).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZJ and CW designed the study. CW was a major
contributor in writing the manuscript. CC and YC performed cell
culture and MTT assay experiments. LZ and YW performed the
experiments and analyzed the data regarding ailanthone-induced
apoptosis and cell cycle arrest in HL-60 cells. CL performed AO
staining. YH and ZY performed western blotting. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vardiman JW, Thiele J, Arber DA, Brunning
RD, Borowitz MJ, Porwit A, Harris NL, Le Beau MM,
Hellström-Lindberg E, Tefferi A and Bloomfield CD: The 2008
revision of the World Health Organization (WHO) classification of
myeloid neoplasms and acute leukemia: Rationale and important
changes. Blood. 114:937–951. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Routledge DJ and Bloor AJ: Recent advances
in therapy of chronic lymphocytic leukaemia. Br J Haematol.
174:351–367. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Y, Wang WJ, Su C, Zhang DM, Xu LP, He
RR, Wang L, Zhang J, Zhang XQ and Ye WC: Cytotoxic quassinoids from
Ailanthus altissima. Bioorg Med Chem Lett. 23:654–657. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
kundu P and Laskar S: A brief resume on
the genus Ailanthus: Chemical and pharmacological aspects.
Phytochem Rev. 9:pp379–412. 2010. View Article : Google Scholar
|
|
5
|
Okunade AL, Bikoff RE, Casper SJ, Oksman
A, Goldberg DE and Lewis WH: Antiplasmodial activity of extracts
and quassinoids isolated from seedlings of Ailanthus altissima
(Simaroubaceae). Phytother Res. 17:675–677. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fukamiya N, Lee KH, Muhammad I, Murakami
C, Okano M, Harvey I and Pelletier J: Structure-activity
relationships of quassinoids for eukaryotic protein synthesis.
Cancer Lett. 220:37–48. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rosati A, Quaranta E, Ammirante M, Turco
MC, Leone A and De Feo V: Quassinoids can induce mitochondrial
membrane depolarisation and caspase 3 activation in human cells.
Cell Death Differ. 11 Suppl 2:S216–S218. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang XL, Yuan YL, Zhang DM, Li F and Ye
WC: Shinjulactone O, a new quassinoid from the root bark of
Ailanthus altissima. Nat Prod Res. 28:1432–1437. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhuo Z, Hu J, Yang X, Chen M, Lei X, Deng
L, Yao N, Peng Q, Chen Z, Ye W and Zhang D: Ailanthone inhibits
Huh7 cancer cell growth via cell cycle arrest and apoptosis in
vitro and in vivo. Sci Rep. 5:161852015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kondo S, Tanaka Y, Kondo Y, Hitomi M,
Barnett GH, Ishizaka Y, Liu J, Haqqi T, Nishiyama A, Villeponteau
B, et al: Antisense telomerase treatment: Induction of two distinct
pathways, apoptosis and differentiation. FASEB J. 12:801–811. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Portt L, Norman G, Clapp C, Greenwood M
and Greenwood MT: Anti-apoptosis and cell survival: A review.
Biochim Biophys Acta. 1813:238–259. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Arozena Acevedo A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Paglin S, Hollister T, Delohery T, Hackett
N, McMahill M, Sphicas E, Domingo D and Yahalom J: A novel response
of cancer cells to radiation involves autophagy and formation of
acidic vesicles. Cancer Res. 61:439–444. 2001.PubMed/NCBI
|
|
15
|
Kondo Y, Kanzawa T, Sawaya R and Kondo S:
The role of autophagy in cancer development and response to
therapy. Nat Rev Cancer. 5:726–734. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Klionsky DJ and Emr SD: Autophagy as a
regulated pathway of cellular degradation. Science. 290:1717–1721.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kanzawa T, Kondo Y, Ito H, Kondo S and
Germano I: Induction of autophagic cell death in malignant glioma
cells by arsenic trioxide. Cancer Res. 63:2103–2108.
2003.PubMed/NCBI
|
|
18
|
Tsang CK, Qi H, Liu LF and Zheng XF:
Targeting mammalian target of rapamycin (mTOR) for health and
diseases. Drug Discov Today. 12:112–124. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xia Z, Zhang H, Xu D, Lao Y, Fu W, Tan H,
Cao P, Yang L and Xu H: Xanthones from the leaves of garcinia cowa
induce cell cycle arrest, apoptosis, and autophagy in cancer cells.
Molecules. 20:11387–11399. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schwartz GK and Shah MA: Targeting the
cell cycle: A new approach to cancer therapy. J Clin Oncol.
23:9408–9421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pardee AB: A restriction point for control
of normal animal cell proliferation. Proc Natl Acad Sci USA.
71:1286–1290. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sherr CJ: The Pezcoller lecture: Cancer
cell cycles revisited. Cancer Res. 60:3689–3695. 2000.PubMed/NCBI
|
|
23
|
Watson NC, Di YM, Orr MS, Fornari FA Jr,
Randolph JK, Magnet KJ, Jain PT and Gewirtz DA: Influence of
ionizing radiation on proliferation, c-myc expression and the
induction of apoptotic cell death in two breast tumour cell lines
differing in p53 status. Int J Radiat Biol. 72:547–559. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lavhale MS, Kumar S, Mishra SH and
Sitasawad SL: A novel triterpenoid isolated from the root bark of
Ailanthus excelsa Roxb (Tree of Heaven), AECHL-1 as a potential
anti-cancer agent. PLoS One. 4:e53652009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang ZH, Peng ZL, Duan ZL and Liu H:
Expression and clinical significance of autophagy gene Beclin 1 in
cervical squamous cell carcinoma. Sichuan Da Xue Xue Bao Yi Xue
Ban. 37:860–863. 2006.(In Chinese). PubMed/NCBI
|
|
26
|
Xu MY, Lee DH, Joo EJ, Son KH and Kim YS:
Akebia saponin PA induces autophagic and apoptotic cell death in
AGS human gastric cancer cells. Food Chem Toxicol. 59:703–708.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mizushima N, Ohsumi Y and Yoshimori T:
Autophagosome formation in mammalian cells. Cell Struct Funct.
27:421–429. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Česen MH, Pegan K, Spes A and Turk B:
Lysosomal pathways to cell death and their therapeutic
applications. Exp Cell Res. 318:1245–1251. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamamoto A, Tagawa Y, Yoshimori T,
Moriyama Y, Masaki R and Tashiro Y: Bafilomycin A1 prevents
maturation of autophagic vacuoles by inhibiting fusion between
autophagosomes and lysosomes in rat hepatoma cell line, H-4-II-E
cells. Cell Struct Funct. 23:33–42. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Klionsky DJ, Elazar Z, Seglen PO and
Rubinsztein DC: Does bafilomycin A1 block the fusion of
autophagosomes with lysosomes? Autophagy. 4:849–850. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mehdi S: Cell-penetrating inhibitors of
calpain. Trends Biochem Sci. 16:150–153. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Seglen PO, Grinde B and Solheim AE:
Inhibition of the lysosomal pathway of protein degradation in
isolated rat hepatocytes by ammonia, methylamine, chloroquine and
leupeptin. Eur J Biochem. 95:215–225. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Phadwal K, Watson AS and Simon AK:
Tightrope act: Autophagy in stem cell renewal, differentiation,
proliferation, and aging. Cell Mol Life Sci. 70:89–103. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gozuacik D and Kimchi A: Autophagy as a
cell death and tumor suppressor mechanism. Oncogene. 23:2891–2906.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tanida I, Ueno T and Kominami E: LC3
conjugation system in mammalian autophagy. Int J Biochem Cell Biol.
36:2503–2518. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Seaman MN, Marcusson EG, Cereghino JL and
Emr SD: Endosome to Golgi retrieval of the vacuolar protein sorting
receptor, Vps10p, requires the function of the VPS29, VPS30, and
VPS35 gene products. J Cell Biol. 137:79–92. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pattingre S, Tassa A, Qu X, Garuti R,
Liang XH, Mizushima N, Packer M, Schneider MD and Levine B: Bcl-2
antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell.
122:927–939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mizushima N and Komatsu M: Autophagy:
Renovation of cells and tissues. Cell. 147:728–741. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang P, Sun D and Jiang F: Ailanthone
promotes human vestibular schwannoma cells apoptosis and autophagy
by down-regulation of miR-21. Oncol Res. Jan 3–2018.(Epub ahead of
print). View Article : Google Scholar
|