Introduction
Angiogenesis is a key event in the promotion of
cancer, being a process required for tumor cell proliferation,
invasion and metastasis (1–3). Hepatocellular carcinoma (HCC) has been
recognized to be a hypervascular tumor, and its prognosis is
associated with angiogenesis (4). A
number of anti-angiogenesis drugs have been approved by the US Food
and Drug Administration and are being used in cancer therapy
(5). In the treatment of HCC,
anti-angiogenic therapy is considered among the most promising
strategies. However, in clinical practice, anti-angiogenic therapy
alone appears to be insufficient for improving patient survival,
for overcoming the occurrence of resistance and for producing
enduring clinical responses (6,7).
Furthermore, immunosuppression occurs as a result of the
establishment of hypoxia during anti-angiogenic therapy, and thus
anti-angiogenic drugs are typically used in combination with
chemotherapeutic and immune-enhancing agents (7–10).
Therefore, the identification of an anti-angiogenic lead compound
with the properties of minimal toxicity and immunoprotection that
may serve as a natural chemopreventative agent is becoming
increasingly important (11).
Osthole, 7-methoxy-8-(3-methyl-2-butenyl) coumarin,
is a bioactive coumarin derivative that may be extracted from a
number of medicinal plants, including Cnidium monnieri (L.)
Cusson. It has a history of use in traditional Chinese medicine for
the treatment of eczema, cutaneous pruritus, trichomonas vaginalis
infection and sexual dysfunction. More recent studies have revealed
that osthole possesses antitumor effects by inhibiting tumor cell
growth and inducing apoptosis (12–15).
Previous studies by our group have demonstrated that osthole may
effectively inhibit tumor growth in various HCC cell lines and
models with no or minimal toxicity via the induction of apoptosis,
and in tumor-bearing mice with HCC, via the enhancement of
antitumor immune responses mediated by T cells (16,17).
However, it is currently unknown whether osthole has
anti-angiogenic activity in HCC.
Anti-angiogenic drugs aimed at blocking vessel
growth in cancer are based on the targeting of vascular endothelial
growth factor (VEGF)-VEGF receptor (VEGFR) signaling (5,18). A high
level of VEGF expression has been identified in HCC (19). Nuclear factor-κB (NF-κB) has been
reported to be a critical regulator of the VEGF pathway (20). When cells are stimulated by external
factors, they trigger a series of enzyme-linked reactions. The
activation of NF-κB promotes tumor necrosis factor (TNF)-α to
produce VEGF and positively regulate the expression of mRNA and
proteins in the VEGF pathway (21). A
previous study by our group also identified that osthole
significantly suppressed NF-κB activity in a time- and
dose-dependent manner in hepatoma cells (16).
In the present study, an orthotopic mouse model of
HCC was established, and the effects of osthole on microvessel
density (MVD) in tumor and adjacent tissues, and on tumor growth
were examined to investigate its potential anti-HCC role as an
inhibitor of angiogenesis; furthermore, the effects of osthole on
the expression of NF-κB and VEGF in the tumor and adjacent tissues
was determined to examine the potential mechanism underlying its
anti-angiogenic effect.
Materials and methods
Chemicals and reagents
Osthole
(C15H16O3;molecular weight,
244.29; purity, ≥99%) was purchased from National Institutes for
Food and Drug Control, Beijing, China and dissolved in corn oil
prior to use. All other chemicals and reagents were purchased from
Sigma-Aldrich; Merck KGaA (Darmstadt, Germany).
Cell culture
The murine HCC Hepa1-6 cell line was a gift from Dr
Limin Zheng (School of Life Sciences, Sun Yat-Sen University,
Guangzhou, China). Cells were maintained in high glucose Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), supplemented with 10% heat-inactivated
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) at 37°C
in a humidified atmosphere containing 5% CO2.
Animal model and treatment
The male C57/BL6 mice [n, 88; age, 6–8 weeks;
weight, 20–22 g; provided by the animal experiment center of Matt
Albert Technology Co., Ltd., Suzhou, China; animal certificate no.
SCXK (JING) 2014-0004] were used for the establishment of an
orthotopic mouse model of HCC following adaptive feeding for 3
days. An orthotopic transplanted model of murine HCC was
established as described by previous studies (22,23). The
model was induced by intrahepatic implantation of 2×106
Hepa1-6 cells into the left liver lobes of mice, and the animals
were maintained in laminar flow cabinets under pathogen-free
conditions, and Small Animal Diagnostic Ultrasound (IVIS Lumina;
Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, USA) was
performed at early and late time points to confirm the presence of
a tumor and for assessment of tumor burden (24). When the tumor weight reached >10%
of the body weight of the experimental mice, the mice were
sacrificed by cervical dislocation. All experimental protocols and
procedures were performed in accordance with the EU Directive
2010/63/EU for animal experiments. The present study was approved
by the Local Ethics Committee of Suzhou Hospital of Traditional
Chinese Medicine (Suzhou, China). The mice were administrated with
osthole according to programme A and B.
Programme A was as follows: The mice bearing tumors
at 8 days that were in the initial tumor stages were randomly
distributed into five groups and each group consisted of 8 mice.
They were treated with 61 mg/kg (0.25 mmol/kg), 122 mg/kg (0.5
mmol/kg) and 244 mg/kg (1.00 mmol/kg) osthole intraperitoneally
(i.p.) in 0.2-ml corn oil, with corn oil alone as the model control
group and cisplatin (5 mg/kg; Qilu Pharmaceutical Co., Ltd., Jinan,
China) as the chemotherapy control once every other day for 2
weeks. Programme B was as follows: The mice bearing tumors at 13
days that were at the rapid growth stage of the tumor were randomly
distributed into five groups and each group consisted of 8 mice.
The mode of administration was the same as that of Programme A.
Additionally, 8 C57BL/6 mice without tumor cell inoculation were
treated with corn oil alone as the normal control group.
Assessment of tumor weight
A total of 24 h after the last treatment
administration at day 14, mice were sacrificed. The tumor tissues
and adjacent tissues were weighed and collected. Tumor inhibition
rate (%)=(mean tumor weight of model control group-mean tumor
weight of experiment group)/mean tumor weight of model control
group ×100.
Immunohistochemistry (IHC) assay and
analysis
Tumor and adjacent tissue specimens from the
harvested livers were fixed for 24 h at 25°C in 4% formalin
followed by embedding in paraffin. Tissue sections (4-µm thick)
were subjected to IHC. For deparaffinization, sections were washed
in xylene for 5 min each a total of three times, then in two washes
of 100% ethanol for 10 min each, in two washes of 95% ethanol for
10 min each and washed sections twice in dH2O for 5 min
each. For antigen retrieval, slides were brought to a boil in 10 mM
sodium citrate buffer, (pH 6.0) and maintained at a sub-boiling
temperature for 10 min and cooled for 30 min. In order to block
endogenous peroxidase blocking, sections were incubated in 3%
hydrogen peroxide for 10 min and were washed in dH2O
twice for 5 min each, and blocked each section with 100–150 µl
blocking solution (1% w/v BSA, TBS) for 2 h at 25°C. The sections
were subjected to staining procedures with purified monoclonal
anti-rabbit CD34 primary antibody (dilution, 1:250; cat. no.
ab81289; Abcam, Cambridge, UK) at 4°C overnight. Subsequently,
biotinylated goat anti-rabbit immunoglobulin secondary antibodies
(dilution, 1:2,000; cat. no. ab205718; Abcam, Cambridge, UK) was
incubated for 1 h at 25°C. Rabbit specific HRP conjugate (cat. no.
D110117; Sangon Biotech Co., Ltd., Shanghai, China) was incubated
for 15 min at 25°C. The visualization signals were developed using
3,3′-diaminobenzidine chromogen substrate (Boster Biological
Technology, Pleasanton, CA, USA) and the slides were counterstained
with Meyer's hematoxylin for 1~2 min at 25°C and dehydrated through
ethanol and xylene series (25). The
sections were observed with a light microscope (Nikon Corporation,
Tokyo, Japan) at ×200 magnification.
ImageJ software (version 1.8.0; National Institutes
of Health, Bethesda, MD, USA) was used to quantify MVD based on
CD34 staining. The percentage of positively stained area was
calculated using a color deconvolution for separating the staining
components in ≥5 fields per section (26). The results are presented as the
percentage of the treated group compared with the model or normal
group. All sections were analyzed and evaluated independently by
two double blinded pathologists (ZL and YQ), and the results were
reconfirmed when inconsistent.
ELISA assays of VEGF expression in
tumor and adjacent non-tumor tissues
Tumor and adjacent non-tumor liver tissue specimens
of HCC-bearing mice treated with corn oil or osthole were
homogenized on ice with PBS (50 mg tissue and 500 µl PBS).
Homogenates were centrifuged at 3,000 × g for 10 min at 4°C and the
supernatants (100 ml) were used for analysis. The concentrations of
VEGF in the tissue homogenate were quantitatively measured using
commercially available ELISA kits (cat. no. 110518007; eBioscience;
Thermo Fisher Scientific, Inc.) according to manufacturer's
protocols.
Western blot analysis
Liver tumor and adjacent tissues were lysed on ice
using nuclear protein and cytosolic protein extraction kits
(Beyotime Institute of Biotechnology) for 30 min. The lysates were
centrifuged at 14,000 × g for 10 min at 4°C and supernatants were
collected. The nuclear proteins were quantified using a
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology). Equal amount of protein (100 µg) were loaded onto
10% SDS-PAGE and transferred onto polyvinylidene difluoride
membranes. The membranes were blocked with 5% skimmed dried milk
buffered for 2 h at 25°C and incubated with primary antibodies
against NF-κB p65 rabbit mAb (dilution, 1:1,000; cat. no. 0009;
Cell Signaling Technology, Inc., Danvers, MA, USA) and IκB-α
antibody (dilution, 1:1,000; cat. no. 0010; Cell Signaling
Technology, Inc.) overnight at 4°C. Next, the membranes were washed
and incubated with horseradish peroxidase-conjugated Goat
Anti-Rabbit IgG [H+L] secondary antibody (dilution, 1:2,000; cat.
no. 0822WB; Cell Signaling Technology, Inc.) for 2 h at 25°C. Blots
were washed four times with tris-buffered saline with Tween-20 and
detected by a fluorescence visible imaging system (ProteinSimple,
San Jose, CA, USA), according to the manufacturer's protocol. The
quantification was normalized to the corresponding value of GAPDH
(dilution, 1:1,000; cat. no. 0006; Cell Signaling Technology, Inc.)
expression used ImageJ software (version 1.8.0; National Institutes
of Health, Bethesda, MD, USA).
Statistical analysis
All data represent ≥3 independent experiments and
the results of the experimental studies are expressed as the mean ±
standard error of the mean. Statistical significance of differences
was analyzed using the Student's t-test or one-way analysis of
variance, followed by the Bonferroni's or Dunnett's post hoc tests,
and correlation analysis was assessed using Pearson's correlation
coefficient. Statistical analyses were performed using GraphPad
Prism 5.0 Software (GraphPad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Osthole suppresses tumor growth in
orthotopic HCC-bearing mice
To establish an orthotopic mouse model of HCC, HCC
Hepa1-6 cells were injected into the left liver lobes of mice. As
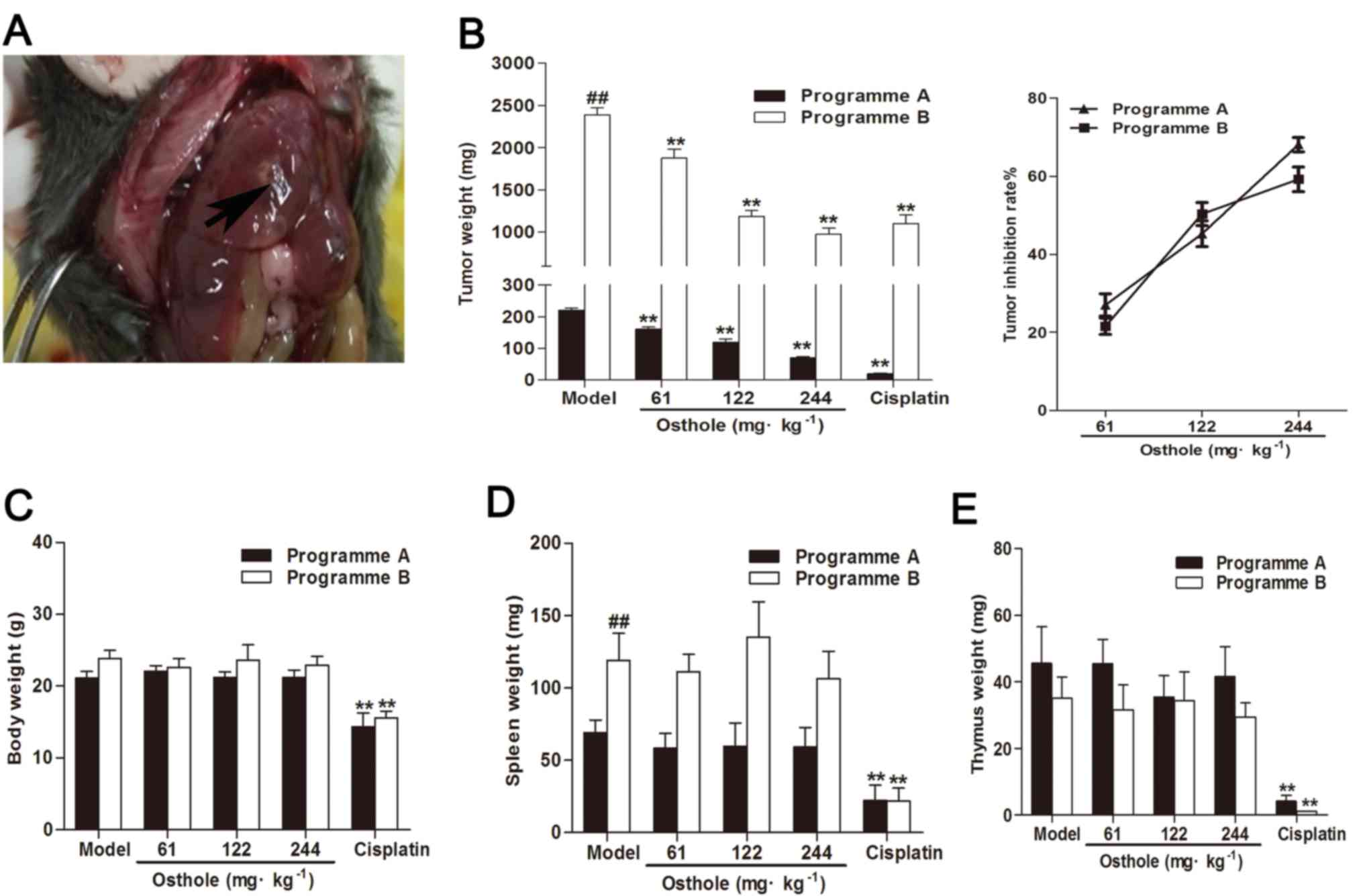
demonstrated in Fig. 1A, the mice
developed internal liver tumors. Subsequently, the antitumor
effects of osthole on an orthotopic mouse model of HCC were studied
throughout different stages of tumor growth. On days 8 and 13 after
the establishment of tumor xenografts (via programmes A and B,
respectively), the mice were treated intraperitoneally with osthole
at 61, 122 and 244 mg/kg for 2 weeks, with corn oil treatment as
the model control and cisplatin treatment (5 mg/kg) as the positive
control. The mean tumor weight of programme A mice in the model
control group was 220.60±19.05 mg, and of programme B mice was
2,390.80±232.57 mg. Compared with the model control group, the
osthole treatment groups (61, 122 and 244 mg/kg) exhibited
significantly suppressed tumor growth (P<0.01); the tumor
inhibition rates of osthole at 61, 122 and 244 mg/kg were 27.02,
45.24 and 68.09%, respectively, in the programme A mice, and 21.58,
50.31 and 59.21%, respectively, in the programme B mice (Fig. 1A). These results demonstrated that
osthole inhibited tumor growth in orthotopic HCC-bearing mice.
Additionally, in the present study, cisplatin
treatment decreased body, thymus and spleen weight in orthotopic
HCC-bearing mice (P<0.01), whereas treatment with osthole had no
effect on body, thymus and spleen weight in the mice (Fig. 1B-D). This indicated that osthole
exerted no apparent toxicity in the orthotopic HCC-bearing mice,
which was similar to the model treatment.
Osthole suppresses angiogenesis in the
tumor tissues of orthotopic HCC-bearing mice
Based on the antitumor effect of osthole, the effect
of osthole on angiogenesis in tumor tissue was subsequently
determined. Programmes A and B were used to investigate the effects
of osthole on the different stages of blood vessel development in
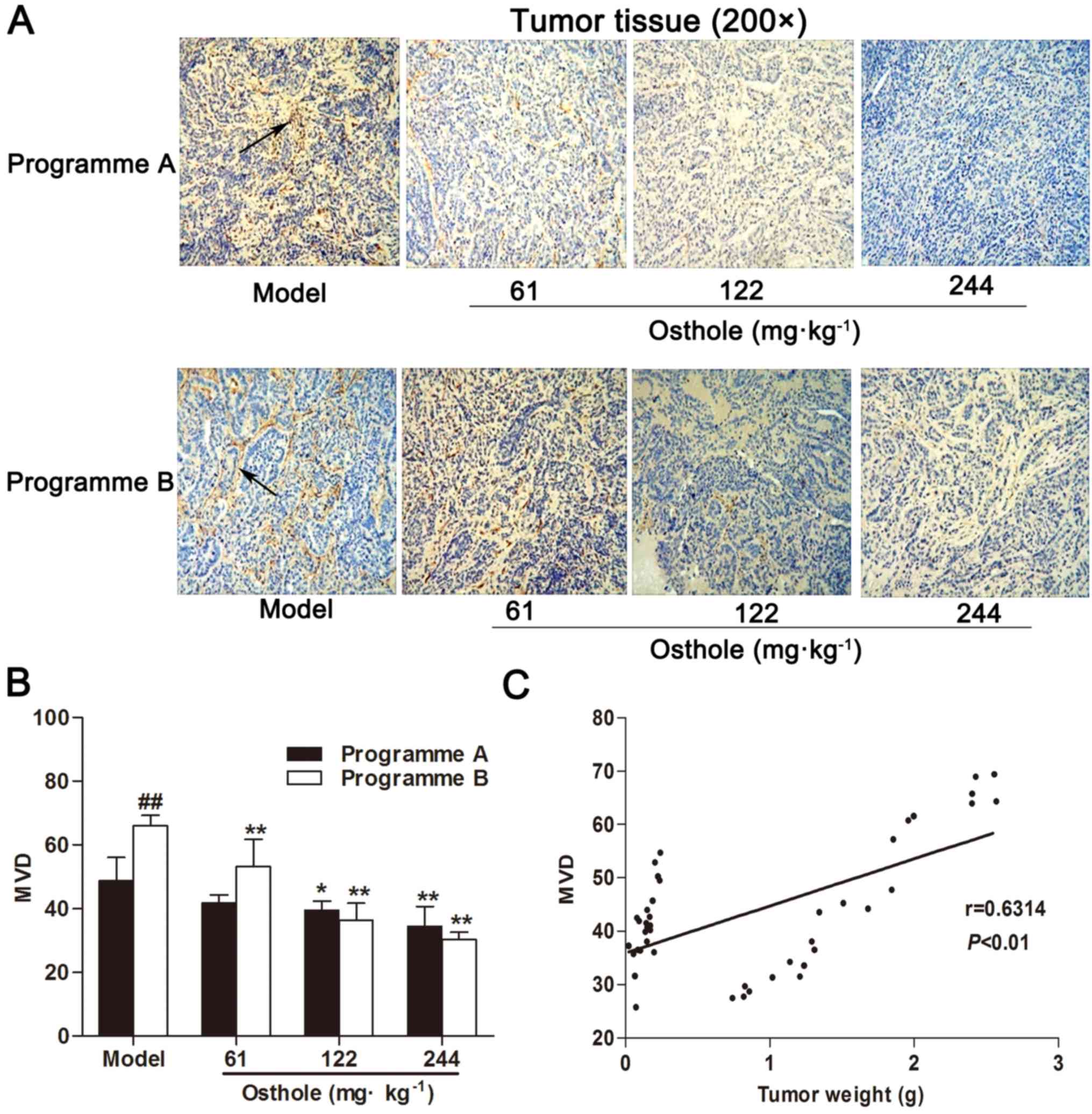
the process of tumor growth. IHC for CD34 expression was performed
on tumor tissue specimens. Compared with the model control group,
MVD in the HCC tissues of the osthole treatment groups was
significantly decreased (P<0.05 and P<0.01; Fig. 2A and B). Furthermore, MVD in HCC
tissues was increased during the progression of tumor growth, and
MVD was correlated with tumor weight (r=0.6314, P<0.01; Fig. 2C). These data suggested that osthole
suppressed angiogenesis in the tumor tissues of orthotopic
HCC-bearing mice.
Effects of osthole on VEGF and NF-κB
signaling in the tumor tissues of orthotopic HCC-bearing mice
The NF-κB-mediated VEGF signaling pathway
(NF-κB/VEGF) has been reported to be one of the mechanisms
regulating tumor angiogenesis (4,27).
Therefore, the present study determined the expression levels of
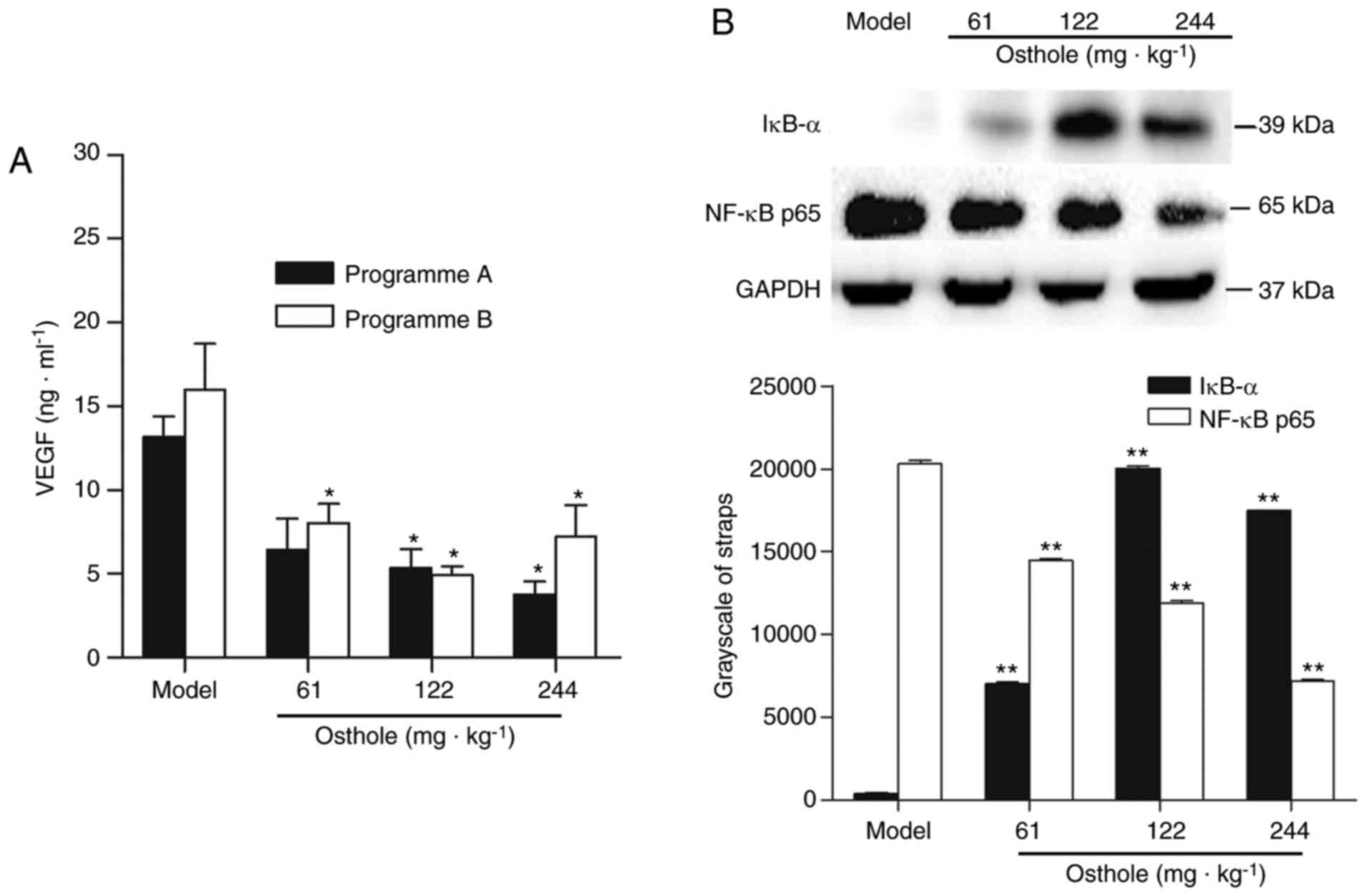
VEGF and NF-κB in HCC tumor tissues. The effects of osthole on the
expression of VEGF in the tumor tissues were analyzed by ELISA. The
concentration of VEGF in tumor tissue homogenate was 13.17±3.51
ng/ml in the programme A tumor-bearing mice and 16.01±9.63 ng/ml in
the programme B tumor-bearing mice (Fig.
3A). Compared with the model control treatment, osthole
decreased the expression of VEGF in tumor tissues (P<0.05;
Fig. 3A). To further elucidate the
molecular basis of the anti-angiogenic effect of osthole, the
effect of osthole on NF-κB activity was investigated. The protein
expression of IκB-α and NF-κB p65 was detected by western blot
analysis. Compared with the model control treatment, the expression
of IκB-α was increased, while that of NF-κB p65 was decreased, in
tumor tissues following osthole treatment (P<0.01; Fig. 3B). Taken together, these results
demonstrated that the downregulation of NF-κB and VEGF was involved
in the anti-angiogenesis effect of osthole in the tumor tissues of
orthotopic HCC-bearing mice.
Effects of osthole on MVD, and VEGF
and NF-κB signaling in the adjacent tissues of orthotopic
HCC-bearing mice
Within the tumor microenvironment, the interaction
of malignant tumor cells and non-malignant cells makes the
microenvironment supportive for the growth of the tumor (28). It has been identified that the
components of the tumor microenvironment serve an important role in
the development of tumor angiogenesis and may modulate tumor
angiogenesis in various ways (29).
In the present study, IHC results showed that there was no
expression of CD34-positive microvessels in normal liver tissue
(Fig. 4A). Adjacent tissue was a
distance of 0.5 cm from the tumor tissue. Compared with the normal
control group, MVD in the adjacent tissues of the model control
group was significantly increased (P<0.01; Fig. 4B), the expression of VEGF was
marginally increased (normal, 20.67±2.13 ng/ml; model, 23.43±1.37
ng/ml; Fig. 4C) and the levels of
IκB-α were decreased while those of NF-κB p65 were increased
(P<0.05; Fig. 4D). The
upregulation of NF-κB and VEGF expression in adjacent tissues was
consistent with the development of HCC.
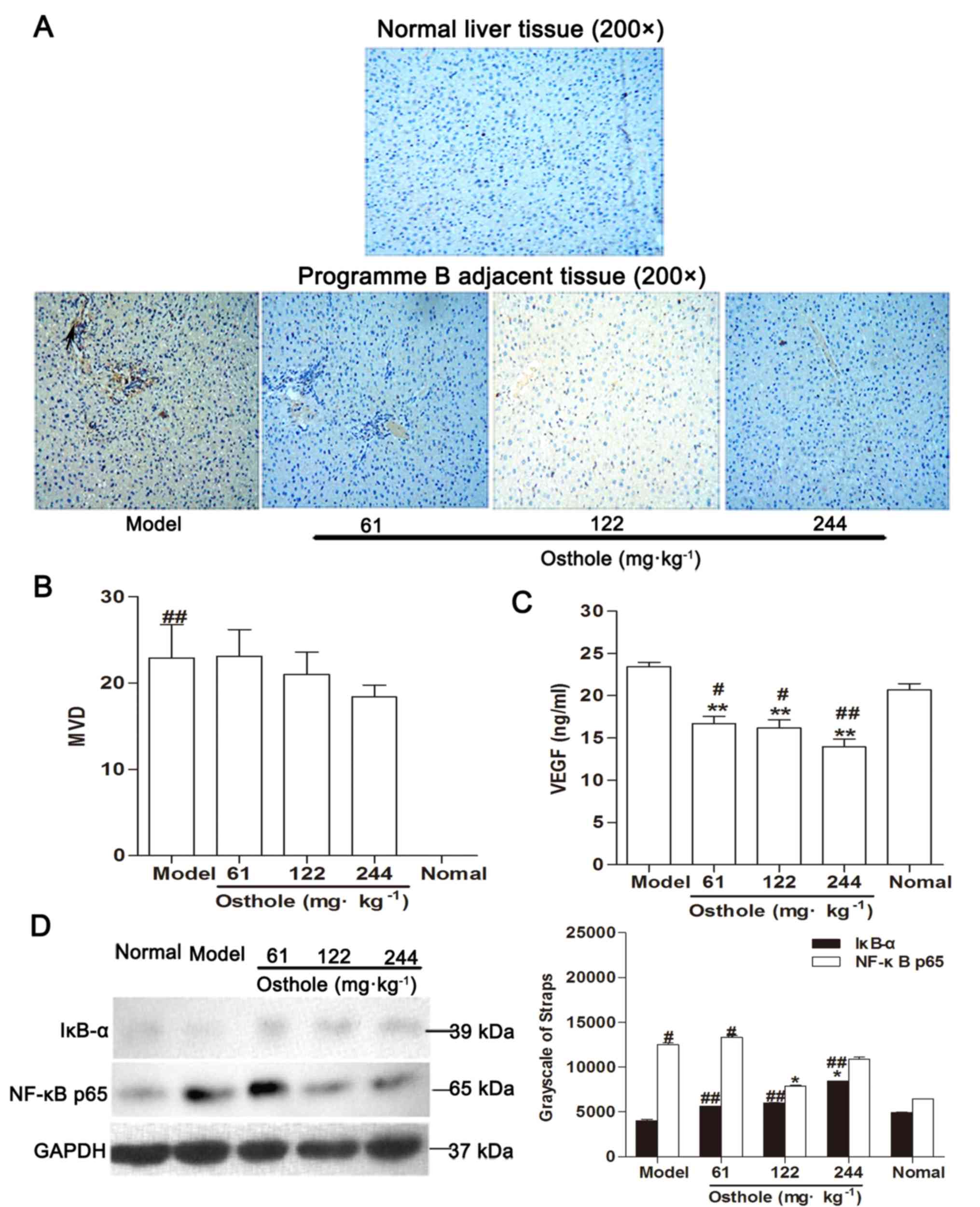 | Figure 4.Effects of osthole on MVD, VEGF and
NF-κB signaling in adjacent tissues of orthotopic hepatocellular
carcinoma-bearing mice. (A) Immunohistochemical staining for blood
vessels with CD34 (arrow) was performed on adjacent tissues from
osthole-treated (61, 122 and 244 mg/kg) mice and normal liver
tissue sections. (B) Compared with the model control group, the
staining indicated that the MVD was decreased in the
osthole-treated mice. (C) Expression levels of VEGF in adjacent
tissues were measured by ELISA. (D) Adjacent tissue lysates were
prepared and quantified. Protein expression of IκB-α and NF-κB p65
were detected by western blot analysis. Equal loading was confirmed
by stripping immunoblots and reprobing for GAPDH. Statistical
analysis of IκB-α and NF-κB p65 quantification. Data are presented
as the mean ± standard error of the mean. *P<0.05, **P<0.01,
compared with the model control group; #P<0.05,
##P<0.01, compared with the normal control group.
MVD, microvessel density; VEGF, vascular endothelial growth factor;
NF-κB, nuclear factor-κB; IκB-α, inhibitor of κB-α. |
Compared with the model control group, MVD in the
adjacent tissues of the osthole-treated groups exhibited a
decreasing trend (Fig. 4B) and the
expression of VEGF was also decreased (P<0.01; Fig. 4C). Additionally, western blot analysis
revealed that the expression levels of IκB-α were increased while
those of NF-κB p65 were decreased in the adjacent tissues with
osthole treatment (P<0.05; Fig.
4D). These data were consistent with the effects of osthole on
the tumor tissues.
Discussion
The aim of the present study was to investigate the
effect of osthole on the inhibition of angiogenesis in an
orthotopic mouse model of HCC. A novel finding of the present study
was that osthole inhibited angiogenesis in the orthotopic mouse
model of HCC in a dose-dependent manner. Furthermore, in tumor and
adjacent tissues, it was noted that osthole not only downregulated
the expression of VEGF, but also reduced NF-κB activity, which
supported the anti-angiogenesis effect of osthole. Additionally, it
was also demonstrated that osthole inhibited tumor growth in the
orthotopic mouse model of HCC in a dose-dependent manner. The
results of the present study suggested that the anti-angiogenesis
effect of osthole participates in its suppression of tumor
growth.
HCC mouse models are currently established by
chemical induction, transgenesis and transplantation, with
orthotopic transplantation mouse models considered optimal for
imitating the growth of tumors and observing tumor morphology and
microenvironment (30,31). The orthotopic mouse model of HCC can
be established by the injection of HCC cells or the tumor tissues
into the mouse liver, and the growth of tumors can be monitored
using imaging techniques (24,32,33).
The present study directly injected the HCC Hepa1-6 cell line into
the left liver lobes of mice to generate internal liver tumors, and
Small Animal Diagnostic Ultrasound was performed at early (7 or 8
days) and late (20–27 days) time points to confirm the presence of
a tumor and for the assessment of tumor burden, respectively. In
the present study, it was observed that the maximum ratio of tumor
weight (2.571 g) to body weight (26.1 g) was 9.85%, indicating that
the orthotopic mouse model of HCC was appropriate. The present
study established the orthotopic mouse model of HCC to investigate
neovascularization in tumor and adjacent tissues, as well as its
association with tumor growth. At the initial stage of tumor growth
(7 or 8 days after injection of hepatoma cells), intratumoral
neovascularization was mostly absent, while at the rapid growth
stage of HCC (12 or 13 days after injection of hepatoma cells),
intratumoral neovascularization was abundant. A relatively high
dose of osthole was used based on our previous study that aimed to
achieve therapeutic effects, in which osthole-treated mice
exhibited no apparent signs of toxicity. The current orthotopic
mouse model exhibited similar reactions to the xenograft HCC model
in our previous study (16). In the
present study, similarities and differences in the effects of
osthole on the growth and angiogenesis of tumor tissues at the
initial and rapid growth stages of tumor development were observed.
MVD may be involved in the ability of tumor tissue to induce
angiogenesis (34). CD34 is
considered to be a sensitive and specific marker of microvessels in
HCC (35). The present study examined
MVD in tissues based on CD34 expression in microvessels as a
surrogate marker. The results demonstrated that the antitumor and
anti-angiogenesis effects of osthole at the early and middle stages
of HCC were similar. Additionally, there was a lack of
CD34-positive microvessels in normal liver tissue, while the tumor
and adjacent tissues exhibited marked increases in CD34-MVDs, which
was similar to a study undertaken by Dai et al (36). Furthermore, MVD was positively
correlated with tumor weight, which supported the importance of
angiogenesis for tumor growth. In order to grow beyond minimal
size, tumors are required to induce the growth of angiogenesis;
conversely, therapeutic inhibition of angiogenesis leads to
inhibition of tumor growth (1,37). While
the results from the correlation between MVD and tumor weight
exhibited a low r-value. The small sample size and osthole
treatment may have altered the r value obtained in the present
study.
To date, whether osthole exerted anti-angiogenesis
activity in HCC has remained largely unknown. In the present study,
MVD in tumor and adjacent tissues was significantly decreased by
treatment with osthole in a dose-dependent manner. Meanwhile, the
expression of VEGF in tumor and adjacent tissues was markedly
increased during tumor progression, but was significantly decreased
by osthole administration. In addition, the results of the present
study indicated that osthole inhibited the activity of NF-κB and
decreased MVD in tumor and adjacent tissues, which is supported by
previously published data demonstrating the importance of NF-κB in
angiogenesis (38–42). To the best of our knowledge, the
present study was the first to demonstrate the anti-angiogenesis
efficacy of osthole, which may be, at least in part, responsible
for its inhibition of tumor growth.
Anti-angiogenic approaches aimed at blocking vessel
growth in cancer led to the approval of therapeutics targeting VEGF
or VEGFR (8,18). The regulation of VEGF involves
multiple signaling pathways (3).
VEGF-C is the target gene of NF-κB and may enhance the binding
capacity of NF-κB (20). In a
previous study, following the activation of NF-κB, VEGF expression
was increased and angiogenesis was promoted, and this in turn
resulted in an increase in MVD (43).
By contrast, NF-κB signaling blockade significantly inhibited the
expression of the major pro-angiogenic molecules VEGF and
interleukin-8 in vitro and in vivo, and thereby
decreased MVD (44). The results of
the present study demonstrated that the expression of NF-κB and
VEGF in tumor and adjacent tissues was markedly increased during
tumor progression, but was significantly decreased by osthole
administration. This indicated that the mechanism involved in the
anti-angiogenesis effect of osthole on HCC is associated with
attenuation of the downregulation of NF-κB and VEGF. However, the
present study did not provide further evidence linking the
inhibitory effects of osthole on the angiogenesis with the
NF-κB/VEGF signaling pathway, which represents a limitation of the
current study. Further research is required to investigate the role
of the NF-κB/VEGF signaling pathway in the inhibitory effects of
osthole on angiogenesis in vivo and in vitro by
pharmacological and genetic interference with NF-κB. In addition,
NF-κB is involved in inflammatory responses, where it triggers the
secretion of pro-inflammatory cytokines, and thereby induces VEGF
secretion. Previous studies have observed that osthole interfered
with the NF-κB signaling pathway during inflammation (45,46), which
not only indicated the importance of the NF-κB signaling pathway to
osthole-induced inhibition of angiogenesis, but also suggested that
the anti-angiogenesis effect of osthole is associated with the
inhibition of inflammatory responses. Therefore, NF-κB may be the
underlying mechanism of the inhibitory effects of osthole on the
angiogenesis and inflammatory responses, which requires further
study.
To the best of our knowledge, the present study was
the first to demonstrate that osthole inhibited angiogenesis and
tumor growth in mice bearing orthotopic HCC. The results of the
present study indicated that osthole attenuates angiogenesis to
inhibit the growth of HCC. Suppression of the NF-κB/VEGF signaling
pathway may be, at least in part, involved in the anti-angiogenesis
effect of osthole on HCC. In combination with our previous study,
osthole exhibited antitumor effects through the induction of
apoptosis, inhibition of angiogenesis and enhancement of antitumor
immune responses, suggesting that it may be a promising leading
compound for the treatment of HCC.
Acknowledgements
No applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81303276), the
Project Fund of Suzhou City (grant no. SYSD2015173) and research
funds from the Suzhou Hospital of Traditional Chinese Medicine
Youth Fund (grant no. YQN2014003).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
All authors contributed to the study concept and
design, and the interpretation of the data. FY, LRZ, GRJ, ML, GQL
and QY acquired and analyzed the data. FY, LRZ and GRJ drafted the
manuscript. LRZ and GRJ reviewed the manuscript for important
intellectual content. All authors revised the article and approved
the final version for publication. LRZ is responsible for the
integrity of the work as a whole.
Ethics approval and consent to
participate
All the animal experimental procedures were
conducted in compliance with the Directive 2010/63/EU. The study
was approved by the Local Ethics Committee of Suzhou Hospital of
Traditional Chinese Medicine (Suzhou, China).
Patient consent for publication
No applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Detmar M: Tumor angiogenesis. J Investig
Dermatol Symp Proc. 5:20–23. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yance DR Jr and Sagap SM: Targeting
angiogenesis with integrative cancer therapies. Integr Cancer Ther.
5:9–29. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao Y and Adjei AA: Targeting
angiogenesis in cancer therapy: Moving beyond vascular endothelial
growth factor. Oncologist. 20:660–673. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu HB, Zhang HF, Zhang X, Li DY, Xue HZ,
Pan CE and Zhao SH: Resveratrol inhibits VEGF expression of human
hepatocellular carcinoma cells through a NF-kappa B-mediated
mechanism. Hepatogastroenterology. 57:1241–1246. 2010.PubMed/NCBI
|
|
5
|
Rajabi M and Mousa SA: The role of
angiogenesis in cancer treatment. Biomedicines. 5:E342017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Berqers G and Hanahan D: Modes of
resistance to anti-angiogenic therapy. Nat Rev Cancer. 8:592–603.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li C, Liu T, Bazhin AV and Yang Y: The
sabotaging role of myeloid cells in anti-angiogenic therapy:
Coordination of angiogenesis and immune suppression by hypoxia. J
Cell Physiol. 232:2312–2322. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hillen F and Griffioen AW: Tumour
vascularization sprouting angiogenesis and beyond. Cancer
Metastasis Rev. 26:489–502. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chouaib S, Messai Y, Couve S, Escudier B,
Hasmim M and Noman MZ: Hypoxia promotes tumor growth in linking
angiogenesis to immune escape. Front Immunol. 3:212012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
McDonald PC, Chafe SC and Dedhar S:
Overcoming hypoxia-mediated tumor progression: Combinatorial
approaches targeting ph regulation, angiogenesis and immune
dysfunction. Front Cell Dev Biol. 4:272016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Y and Martin RC II: Herbal medicine and
hepatocellular carcinoma: Applications and challenges. Evid Based
Complement Alternat Med. 2011:5412092011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang LL, Wang MC, Chen LG and Wang CC:
Cytotoxic activity of coumarins from the fruits of Cnidium
monnieri on leukemia cell lines. Planta Med. 69:1091–1095.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Riviere C, Goossens L, Pommery N, Fourneau
C, Delelis A and Henichart JP: Antiproliferative effects of
isopentenylated coumarins isolated from phellolophium
madagascariense baker. Nat Prod Res. 20:909–916. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu X, Zhang Y, Qu D, Jiang T and Li S:
Osthole induces G2/M arrest and apoptosis in lung cancer A549 cells
by modulating PI3K/Akt pathway. J ExpClin Cancer Res. 30:33–39.
2011. View Article : Google Scholar
|
|
15
|
Chou SY, Hsu CS, Wang KT, Wang MC and Wang
CC: Antitumor effects of osthole from Cnidium monnieri: An
in vitro and in vivo study. Phytother Res. 21:226–230. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang L, Jiang G, Yao F, He Y, Liang G,
Zhang Y, Hu B, Wu Y, Li Y and Liu H: Growth inhibition and
apoptosis induced by osthole, a natural coumarin, in hepatocellular
carcinoma. PLoS One. 7:e378652012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang L, Jiang G, Yao F, Liang G, Wang F,
Xu H, Wu Y, Yu X and Liu H: Osthole promotes anti-tumor immune
responses in tumor-bearing mice with hepatocellular carcinoma.
Immunopharmacol Immunotoxicol. 37:301–307. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Potente M, Gerhardt H and Carmeliet P:
Basic and therapeutic aspects of angiogenesis. Cell. 146:873–887.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Turlin B, Le Quilleuc D, Leroyer P,
Brissot P, Deugnier Y and Loréal O: High vascular endothelial
growth factor(VEGF) expression in chemically-induced hepatic
microcancers in mice. J Hepatol. 37:620–624. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tong Q, Zheng L, Lin L, Li B, Wang D,
Huang C and Li D: VEGF is upregulated by hypoxia-induced mitogenic
factor via the PI-3K/Akt-NF-kappaB signaling pathway. Respir Res.
7:372006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Q, Lu Y, Proulx ST, Guo R, Yao Z,
Schwarz EM, Boyce BF and Xing L: Increased lymphangiogenesis in
joints of mice with inflammatory arthritis. Arthritis Res Ther.
9:R1182007. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kwon OJ, Kim PH, Huyn S, Wu L, Kim M and
Yun CO: A hypoxia- and {alpha}-fetoprotein-dependent oncolytic
adenovirus exhibits specific killing of hepatocellular carcinomas.
Clin Cancer Res. 16:6071–6082. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wei D, Li Q, Wang XL, Wang Y, Xu J, Feng
F, Nan G, Wang B, Li C, Guo T, et al: Oncolytic Newcastle disease
virus expressing chimeric antibody enhanced anti-tumor efficacy in
orthotopic hepatoma-bearing mice. J ExpClin Cancer Res.
34:1532015.
|
|
24
|
McFadden DG, Vernon A, Santiago PM,
Martinez-McFaline R, Bhutkar A, Crowley DM, McMahon M, Sadow PM and
Jacks T: p53 constrains progression to anaplastic thyroid carcinoma
in a Braf-mutant mouse model of papillary thyroid cancer. Proc Natl
Acad Sci U S A. 111:E1600–E1609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Meng J, Liu Y, Han J, Tan Q, Chen S, Qiao
K, Zhou H, Sun T and Yang C: Hsp90β promoted endothelial
cell-dependent tumor angiogenesis in hepatocellular carcinoma. Mol
Cancer. 16:722017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim HY, Kim J, Thi Ha HT, Bang OS, Lee WS
and Hong S: Evaluation of anti-tumorigenic activity of BP3B against
colon cancer with patient-derived tumor xenograft model. BMC
Complement Altern Med. 16:4732016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen H, Zhang J, Luo J, Lai F, Wang Z,
Tong H, Lu D, Bu H, Zhang R and Lin S: Antiangiogenic effects of
oxymatrine on pancreatic cancer by inhibition of the NF-κB-mediated
VEGF signaling pathway. Oncol Rep. 30:589–595. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hui L and Chen Y: Tumor microenvironment:
Sanctuary of the devil. Cancer Lett. 368:7–13. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ribatti D and Vacca A: The role of
microenvironment in tumor angiogenesis. Genes Nutr. 3:29–34. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Leenders MW, Nijkamp MW and Rinkes Borel
IH: Mouse models in liver cancer research: A review of current
literature. World J Gastroenterol. 14:6915–6923. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rao Q, You A, Guo Z, Zuo B, Gao X, Zhang
T, Du Z, Wu C and Yin H: Intrahepatic tissue implantation
represents a favorable approach for establishing orthotopic
transplantation hepatocellular carcinoma mouse models. PLoS One.
11:e01482632016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li B, Zhang Y, Wu W, Du G, Cai L, Shi H
and Chen S: Neovascularization of hepatocellular carcinoma in a
nude mouse orthotopic liver cancer model: A morphological study
using X-ray in-line phase-contrast imaging. BMC Cancer. 17:732017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Woodfield SE, Shi Y, Patel RH, Jin J,
Major A, Sarabia SF, Starosolski Z, Zorman B, Gupta SS, Chen Z, et
al: A novel cell line based orthotopic xenograft mouse model that
recapitulates human hepatoblastoma. Sci Rep. 7:177512017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis - correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
El-Assal ON, Yamanoi A, Soda Y, Yamaguchi
M, Igarashi M, Yamamoto A, Nabika T and Nagasue N: Clinical
significance of microvessel density and vascular endothelial growth
factor expression in hepatocellular carcinoma and surrounding
liver: Possible involvement of vascular endothelial growth factor
in the angiogenesis of cirrhotic liver. Hepatology. 27:1554–1562.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dai L, Peng XX, Tan EM and Zhang JY:
Tumor-associated antigen CAPERα and microvessel density in
hepatocellular carcinoma. Oncotarget. 7:16985–16995. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cao Y and Langer R: A review of Judah
Folkman's remarkable achievements in biomedicine. Proc Natl Acad
Sci USA. 105:13203–13205. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Aggarwal BB, Vijayalekshmi RV and Sung B:
Targeting inflammatory pathways for prevention and therapy of
cancer: Short-term friend, long-term foe. Clin Cancer Res.
15:425–430. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schmidt D, Textor B, Pein OT, Licht AH,
Andrecht S, Sator-Schmitt M, Fusenig NE, Angel P and
Schorpp-Kistner M: Critical role for NF-kappaB-induced JunB in VEGF
regulation and tumor angiogenesis. EMBO J. 26:710–719. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ismail HA, Lessard L, Mes-Masson AM and
Saad F: Expression of NF-kappaB in prostate cancer lymph node
metastases. Prostate. 58:308–313. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ghosh S and Karin M: Missing pieces in the
NF-kappaB puzzle. Cell. 109 Suppl:S81–S96. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Javan H, Szucsik AM, Li L, Schaaf CL,
Salama ME and Selzman CH: Cardiomyocyte p65 nuclear factor-κB is
necessary for compensatory adaptation to pressure overload. Circ
Heart Fail. 8:109–118. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang B, Wang D, Ji TF, Shi L and Yu JL:
Overexpression of lncRNA ANRIL up-regulates VEGF expression and
promotes angiogenesis of diabetes mellitus combined with cerebral
infarction by activating NF-κB signaling pathway in a rat model.
Oncotarget. 8:17347–17359. 2017.PubMed/NCBI
|
|
44
|
Xiong HQ, Abbruzzese JL, Lin E, Wang L,
Zheng L and Xie K: NF-kappaB activity blockade impairs the
angiogenic potential of human pancreatic cancer cells. Int J
Cancer. 108:181–188. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yu C, Li P, Qi D, Wang L, Qu HL, Zhang YJ,
Wang XK and Fan HY: Osthole protects sepsis-induced acute kidney
injury via down-regulating NF-κB signal pathway. Oncotarget.
8:4796–4813. 2017.PubMed/NCBI
|
|
46
|
Li YQ, Wang JY, Qian ZQ, Li YL, Li WN, Gao
Y and Yang DL: Osthole inhibits intimal hyperplasia by regulating
the NF-κB and TGF-β1/Smad2 signalling pathways in the rat carotid
artery after balloon injury. Eur J Pharmacol. 811:232–239. 2017.
View Article : Google Scholar : PubMed/NCBI
|