Introduction
Cervical cancer, which is often induced by human
papilloma virus (HPV) infection, is one of the most serious
life-threatening diseases among women in developing countries
(1,2).
Recently, two prophylactic vaccines, Cervarix and Gardasil, against
the high-risk strains HPV-16 and HPV-18 have been developed and are
used in more than 100 countries worldwide (3). However, these vaccines offer no benefit
for patients that are already infected with HPV, have pre-cancerous
lesions or cervical cancer (4). The
long-term use of routine approaches to treat cervical cancer,
including surgical removal, radiotherapy and chemotherapy, further
damage the health of patients. Accordingly, seeking effective
treatment agents from natural compounds to prevent and treat
cervical cancer without these adverse effects is clinically
urgent.
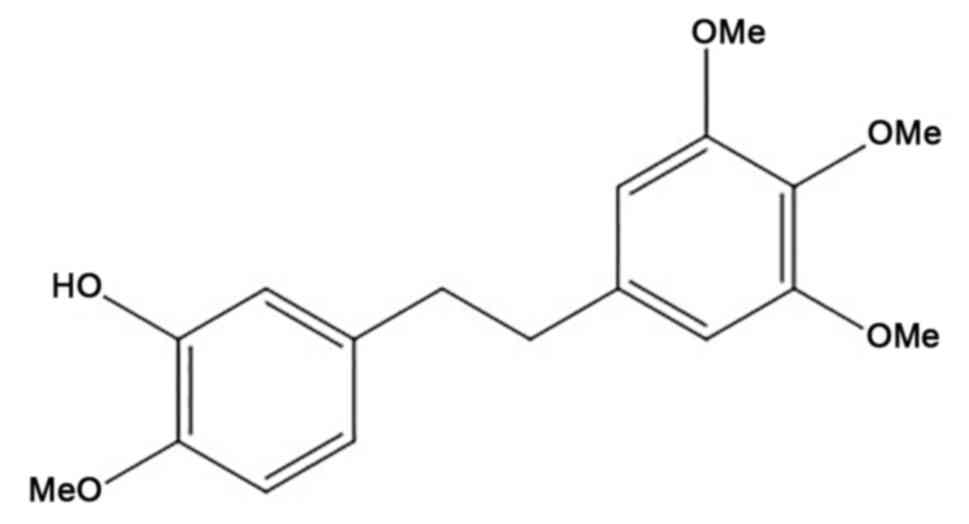
Erianin
[2-Methoxy-5-(2-(3,4,5-trimethoxyphenyl)-ethyl)-phenol] (Fig. 1) is a natural bibenzyl compound
present in Dendrobium chrysotoxum Lindl., which is commonly
known as Shihu in China; it has been used as a herbal drug for
thousands of years in traditional Chinese medicine (TCM), where it
is often used as antipyretic and analgesic medication (5). Previous studies have demonstrated that
erianin can elicit multiple pharmacological effects, including
anti-oxidative (6) and antitumor
activity (7). Erianin has been
reported to inhibit cell proliferation and induce apoptosis in
human promyelocytic leukemia HL-60 cells (8), and reverse multidrug resistance in
B16/hMDR-1 cells (9). However, the
effect of erianin on cervical cancer HeLa cells and the underlying
molecular mechanisms behind these effects remain unclear. Thus, in
the present study, the anticancer effects of erianin in HeLa cells
compared with paclitaxel, a frequently used chemotherapeutic drug,
were examined. Additionally, the involvement of tumor protein p53
and extracellular signal-regulated kinase (ERK) signaling were also
investigated as potential molecular mechanism. The present study
aimed to elucidate the effects and potential mechanism of erianin
on cervical cancer HeLa cells in vitro.
Materials and methods
Reagents
Erianin and paclitaxel (PTX) were purchased from
Chengdu Must Bio-Technology Co. Ltd. (Chengdu, China), purity
>98%, and dissolved in dimethyl sulfoxide (DMSO) (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) for use.
Cell culture
HeLa cells, human cervical cancer cell line, were
obtained from American Type Culture Collection (Manassas, VA, USA).
HeLa cells were cultured in Dulbecco's modified Eagle's medium
(DMEM) (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
with 10% fetal bovine serum (FBS) (Gibco; Thermo Fisher Scientific,
Inc.) at 37°C in a humidified atmosphere of 5% CO2.
Cell viability assay
The cytotoxic activity of erianin on HeLa cells was
assessed using an MTT assay. HeLa cells were treated with erianin
at various concentrations (3.9, 7.8, 15.7, 31.4 or 157.0 µM) for
different time points (24, 48 or 72 h), and then incubated with MTT
(0.5 mg/ml) at 37°C for 4 h. Meanwhile, HeLa cells were treated
with PTX (7.81, 15.63, 31.25, 62.50, 125.0 and 250.0 nM) for 48 h
as the positive groups. The purple formazan crystals were dissolved
in 0.15 ml of DMSO. Following shaking, the plates read on an
automated microplate spectrophotometer at 570 nm. Assays were
performed in triplicate in three independent experiments.
Cell cycle assay
Following exposure to erianin (3.9, 7.9, 15.7 and
31.4 µM) and 0.05 µM PTX for 12, 24, 36 and 48 h, the control
(without treatment) and experimental groups (3.0×105
cells in a 60-mm dish) were harvested using 0.25% trypsin, washed
with ice-cold PBS and collected by centrifugation at 400 × g for 15
min at 4°C. The cells were suspended in PBS and fixed with 70%
ethanol (v/v) overnight at 4°C. The following day, subsequent to
three washes with ice-cold PBS, the cells were treated with 100 µl
RNase at 37°C for 30 min and stained with 400 µl propidium iodide
(PI) (Nanjing KeyGen Biotech Co. Ltd., Nanjing, China) for 30 min.
Next, cell cycle analysis was performed by flow cytometry (Coulter
Epics XL; Beckman Coulter, Inc., Brea, CA, USA). The data were
analyzed using MultiCycle software (version 6-16-03-F32; Beckman
Coulter, Inc.).
Cell apoptosis assay
For apoptotic cells analysis, HeLa cells were
treated with various concentrations of erianin (3.9, 7.9, 15.7,
31.4) µM and 0.05 µM PTX for 12, 24, 36 and 48 h. The cells were
then collected and washed with ice-cold PBS. Next, early apoptosis
was measured using an Annexin V-Fluorescein Isothiocyanate
(FITC)/PI apoptosis detection kit (BD Pharmingen; BD Biosciences,
Franklin Lakes, NJ, USA). The cells were suspended with 100 µl of
binding buffer and stained with 5 µl annexin V-FITC and 5 µl PI for
20 min at room temperature in the dark and then 400 µl binding
buffer was added. The proportion of apoptotic cells were determined
by flow cytometry and observed using the ImageXpress®
Micro XLS High Content Screening system (Molecular Devices, LLC,
Sunnyvale, CA, USA), with Hoechest 33342 staining.
Western blot assay
Cells were treated with 3.9, 7.8, 15.7 or 31.4 µM
erianin or 0.05 µM PTX for 48 h, lysed using lysis buffer
radioimmunoprecipitation assay (RIPA; 3 ml; cat. no. AR0102; Wuhan
Boster Biological Technology, Ltd., Wuhan, China) buffer with
phenylmethane sulfonyl fluoride (40 µl; cat. no. AR1778; Wuhan
Boster Biological Technology, Ltd.) and a protease inhibitor
cocktail (cat. no. P8340; 30 µl Sigma-Aldrich; Merck KGaA) and the
lysates were measured for protein concentrations with a total
protein quantitative assay (Coomassie brilliant blue method) kit
(cat. no. A405-2; Nanjing Jiancheng Bioengineering Institute,
Nanjing, China) as described previously (10), according to the manufacturer's
protocols. Protein samples (20 µg) were separated by 10% SDS-PAGE
and then transferred to a polyvinylidene fluoride membrane. The
membrane was blocked for 2 h with 5% non-fat dry skimmed milk.
Following incubation overnight at 4°C with antibodies against tumor
protein p53 (hereafter p53) (dilution, 1:1,000; cat. no. ab131442;
Abcam, Cambridge, UK), ERK1/2 (dilution, 1:1,000; cat. no. ab50011;
Abcam), phosphorylated ERK1/2 (dilution, 1:1,000; cat. no. ab36991;
Abcam), caspase-3 (dilution, 1:1,000; cat. no. 9665; Cell Signaling
Technology, Inc., Danvers, MA, USA), B-cell lymphoma-2 (Bcl-2)
(dilution, 1:1,000; cat. no. ab136285; Abcam), Bcl-2-associated X
(Bax) (dilution, 1:1,000; cat. no. ab32503; Abcam), GAPDH
(dilution, 1:1,000; cat. no. BA2913; Wuhan Boster Biological
Technology, Ltd.) and β-actin (dilution, 1:1,000; cat. no. ab3280,
Abcam), the membrane was incubated with horseradish
peroxidase-conjugated anti-rabbit antibody (dilution, 1:5,000; cat.
no. RA1054; Wuhan Boster Biological Technology, Ltd.) at 37°C for 2
h. Finally, the reaction was visualized using SuperLumia enhanced
chimiluminescence kit (cat. no. K22020; Abbkine Scientific Co.,
Ltd, Wuhan, China) and detected using a Molecular
Imager® Gel Doc™ XR+ (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). The integrated density of the bands was
quantified using Image Lab 5.1 software (Bio-Rad Laboratories,
Inc.).
Bioinformatics methods
The data concerning the association between p53
expression and survival ratios in patients with cervical cancer
were acquired from The Cancer Genome Atlas (TCGA) database
(https://cancergenome.nih.gov/). The
interaction regulatory function of p53, ERK1/2, Bax and Bcl-2 was
analyzed using Pathway Commons (http://www.pathwaycommons.org/) (11).
Statistical analysis
All experiments were independently repeated three
times. All quantitative data were presented as the mean ± standard
deviation and analyzed using SPSS 19.0 software (IBM Corp., Armonk,
NY, USA). The half-maximal inhibitory concentration
(IC50) and combination index values were calculated
using a logit regression model. One-way analysis of variance with a
least significant difference post-hoc test was used to compare mean
values between the control and treatment groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of erianin on the
proliferation of HeLa cells
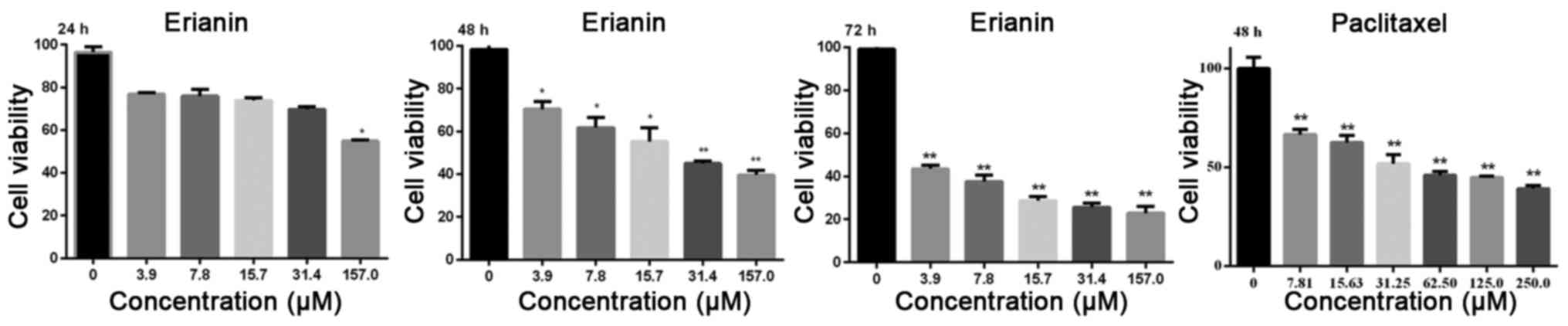
To evaluate the inhibitory effect of erianin on HeLa
cells, cell viability was assessed using an MTT assay after 24, 48,
or 72 h of treatment. The results showed that erianin inhibited
HeLa cells growth in a dose- and time-dependent manner. At doses of
3.9, 7.8, 15.7, 31.4 and 157.0 µM, erianin elicited a significant
inhibition in HeLa cell viability after a 48-h treatment
(P<0.05). The IC50 values of erianin and PTX after 48
h were 8.3±1.3 and 0.055±0.0017 µM, respectively (Fig. 2).
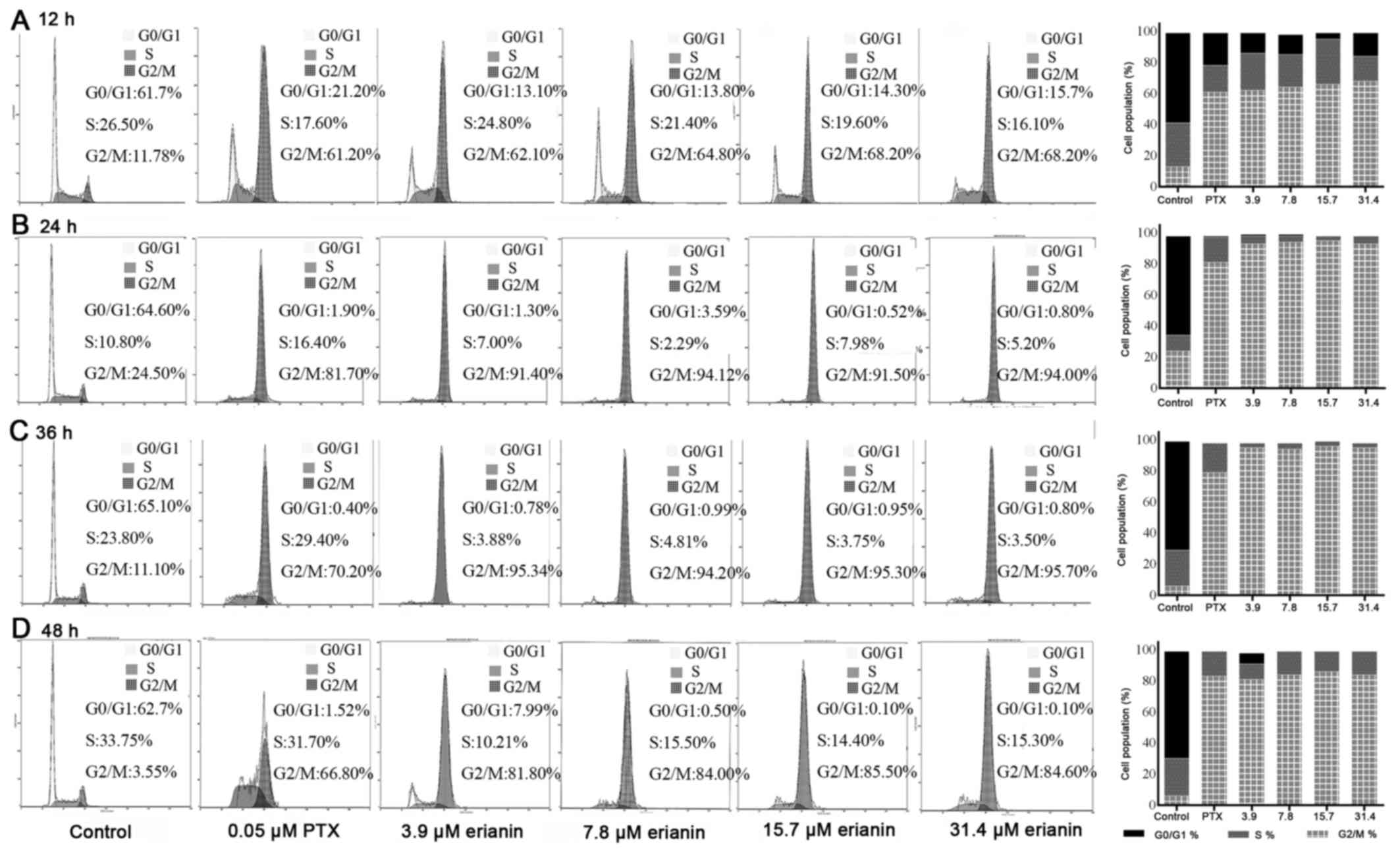
Effects of erianin on cell cycle of HeLa cells. To
investigate whether the inhibition in cell proliferation induced by
erianin could be attributed to the induction of cell cycle arrest,
cell cycle analysis was performed. This analysis revealed that
erianin induced cell cycle arrest in a time- and dose-dependent
manner (Fig. 3). Compared with the
control, the proportion of cells in G0/G1
phase following treatment with erianin and PTX decreased
significantly after 12 h of treatment (P<0.01), whereas the
proportion of cells in G2/M phase increased
significantly (P<0.01) (Fig. 3A),
indicating that erianin could at least partly inhibit HeLa cells
proliferation via G2/M checkpoint arrest.
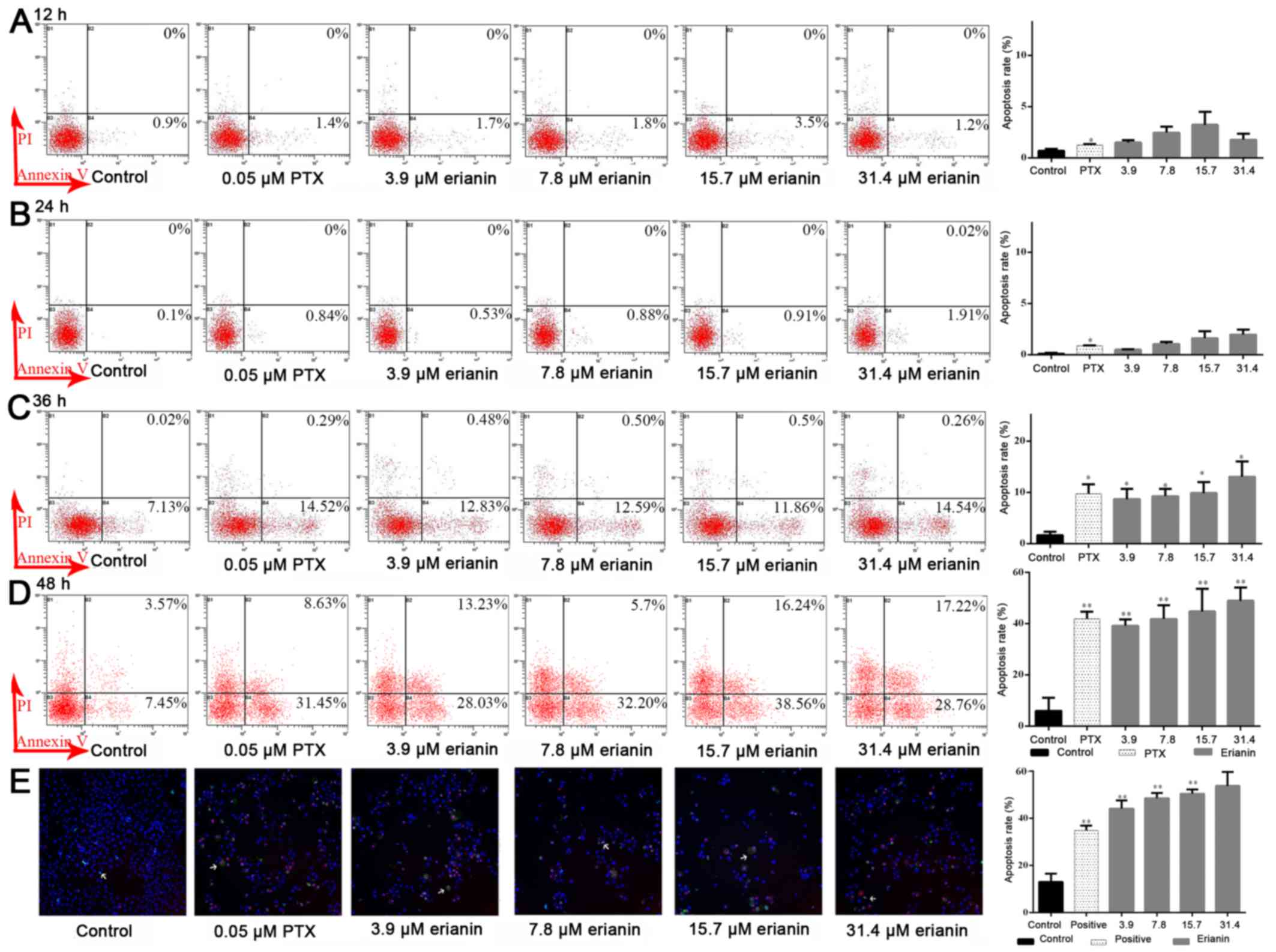
Effects of erianin on apoptosis of
HeLa cells
Induction of apoptosis is an central mechanism by
which anticancer drugs can inhibit cancer cell growth. Flow
cytometry analysis demonstrated that erianin at a dose of 7.8 µM
induced apoptosis rate compared with the control, 0.22±0.26 vs.
1.52±0.29% at 12 h, 0.10±0.1 vs. 8.12±0.92% at 24 h, 1.83±0.71 vs.
11.53±1.22% at 36 h and 5.99±5.12 vs. 41.83±5.32% at 48 h,
respectively (Fig. 4). As shown in
Fig. 4D, erianin could increase rates
of early apoptosis in HeLa cells, from 5.99±5.12% in the control
(untreated cells at 48 h) to 48.91±5.22% in cells treated with 31.4
µM erianin after 48 h of treatment (P<0.05). Therefore, the
results of the present study demonstrated that erianin induced
apoptosis in HeLa cells in a time- and dose-dependent manner.
Effects of erianin on the expression
of p53, ERK1/2, caspase-3, Bcl-2 and Bax proteins
It has been reported that the p53 and ERK pathways
are involved in mitochondrial-based apoptosis (12,13).
Therefore, whether erianin induced mitochondrial-based apoptosis
through the inhibition of ERK1/2 signaling and activation of p53
was examined. The results of western blot analysis revealed that
erianin could reduce the levels of p53 expression and
phosphorylated-ERK1/2 expression without decreasing total ERK1/2
expression, indicating that erianin could affect ERK1/2
phosphorylation in a concentration-dependent manner, compared with
the control (P<0.05; Fig. 5A and
B). Further study indicated that administration of erianin for
48 h evidently promoted caspase-3 cleavage, upregulated the
expression of Bax and downregulated the expression of Bcl-2
(P<0.05; Fig. 5C and D), Note
that, since the effect of erianin on p-ERK1/2 and caspase-3
expression at a concentration of 1.9 µM was similar to that at 3.9
µM, these data were therefore not included in the figure. A
concentration of 3.9 µM as the minimum concentration was selected
for the further investigation.
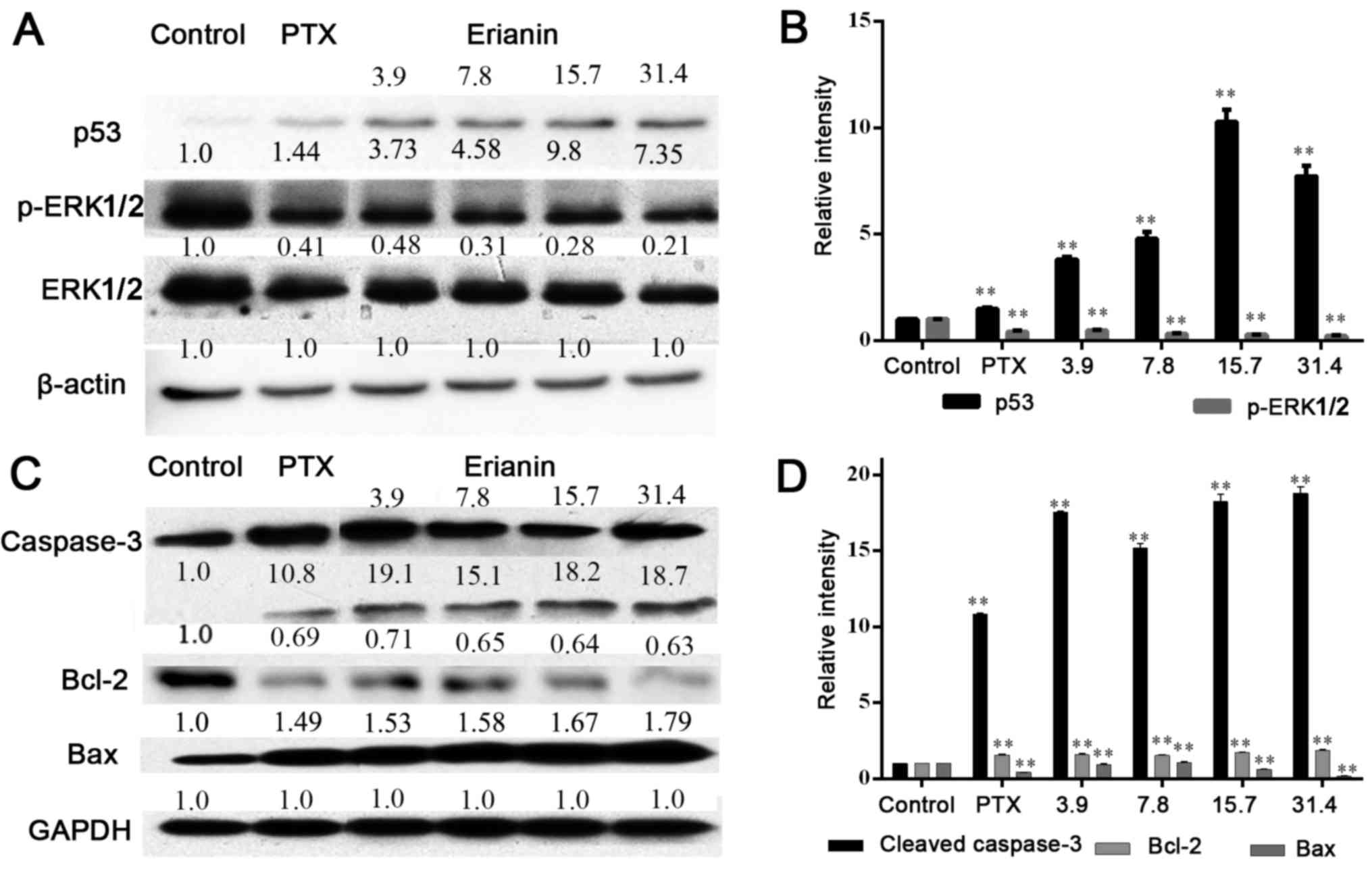 | Figure 5.Effect of erianin on p53/ERK1/2
signaling. (A) The levels of p53, ERK1/2 and its phosphorylated
protein were tested after treatment for 48 h by 0.05 µM PTX and
erianin at concentrations of 3.9, 7.8, 15.7 and 31.4 µM for HeLa
cells, with (B) quantification of this expression. (C) The levels
of caspase-3, Bcl-2 and Bax proteins were measured following
treatment with compounds, with (D) quantification of this
expression. **P<0.01. p53, tumor protein p53; p-ERK1/2,
phosphorylated extracellular signal-regulated kinase 1/2; PTX,
paclitaxel; Bcl-2, B-cell lymphoma-2; Bax, Bcl-2-assocated X; PTX,
paclitaxel. |
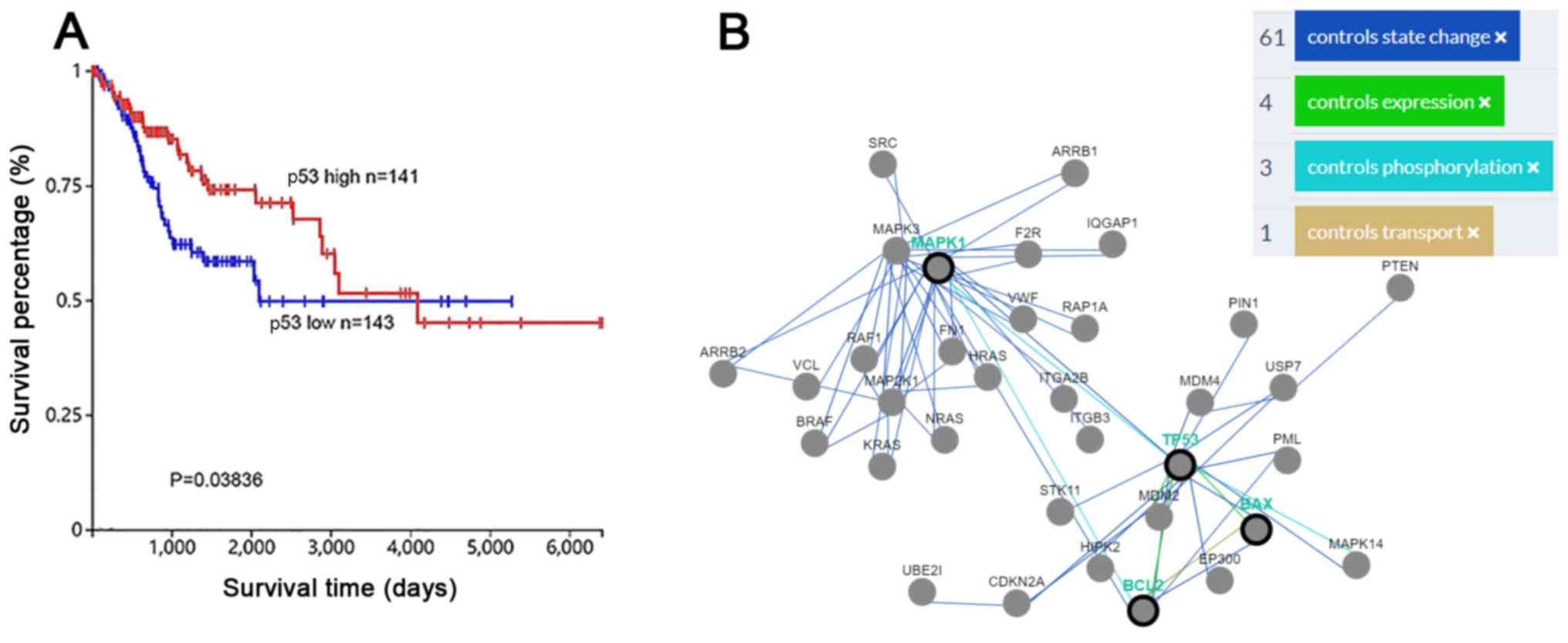
ERK and p53 expression are associated
with mitochondrial-based apoptosis in cervical cancer
p53 is an essential tumor suppressor gene in various
types of cancer (14). According to
data from TGCA, high expression of p53 was positively associated
with the survival rates in patients with cervical cancer (P=0.038)
(https://cancergenome.nih.gov/),
indicating that promoting p53 expression may represent a promising
approach to treating cervical cancer (Fig. 6A). Furthermore, data from Pathway
Commons (http://www.pathwaycommons.org/) in cervical cancer
revealed that p53 serves essential roles in numerous different
pathways, including apoptosis. Specially, p53 could control the
expression and activation of Bcl-2 and Bax, and control
phosphorylation of ERK1/2. The activation of the ERK1/2 pathway
could control the activity of Bcl-2 (Fig.
6B).
Discussion
An epidemiological study indicated that in 2008,
cervical cancer was the third-most common cancer in women worldwide
and the gynecological malignancy with the highest morbidity, with
an estimated 529,000 new cases occurring, resulting in 274,000
mortalities (15). Treatments for
cervical cancer, including surgery, chemotherapy, radiation
therapy, and chemotherapy combined with radiation therapy, can have
a curative effect; however, these treatments are accompanied by
adverse effects and recurrence within a short time (16–20).
Accordingly, seeking effective chemotherapeutical agents to
increase the curative rate, reduce the risk of recurrence and
metastasis, and improve patient quality of life is highly
desirable.
The development of antitumor drugs, particularly
those based on natural products, is receiving considerable
attention. Erianin, a naturally occurring product isolated from
Dendrobium chrysotoxum Lindl., was reported to exert
antitumor (21–23) and anti-oxidative effects (8). However, the anti-cervical cancer
activity and the potential mechanisms of erianin have, to the best
of our knowledge, not been assessed. The present study aimed to
elucidate the effects of erianin on cell growth and apoptosis, and
clarify the potential mechanism by which this occurred, which was
associated with regulation of the ERK1/2 signaling and
mitochondrial pathways.
Investigating the molecular mechanisms of cancer
cell growth is critically important. Cell cycle dysregulation is a
major factor in cancer cell growth (24). ERKs have a key role in promoting cell
survival and cellular proliferation (25,26) and
have been considered to be significant targets for cancer
therapeutics (27). Evidence
indicates that erianin affects cell cycle progression, evidenced by
inhibition of gastric carcinoma SGC-7901 cell proliferation by
blocking progression to S phase (28), and arresting progression at the
G2/M phase in hepatocellular carcinoma Huh7 cells
(23) and human colorectal cancer
SW480 cells (29). In the present
study, erianin exhibited potent anti-proliferative activities
against HeLa cells (IC50=8.3±1.3 µM; Fig. 2) at 48 h, induced the arrest of HeLa
cells at the G2/M phase (Fig.
3), and suppressing the phosphorylation of ERK1/2.
As a tumor suppressor, p53 has a pivotal role in
cell cycle progression, the DNA damage response and apoptosis
(12). Previous studies confirmed
that p53 was one of the most commonly mutation genes in cancer
(30–32). Additionally, the upregulation of p53
in cancer cells may prevent cancer cell proliferation by promoting
cell cycle arrest and apoptosis (33–36). Liu
et al (37) revealed that the
p53 pathway in human cervical cancer cells is activated by
reversion-inducing-cysteine-rich protein with Kazal motifs
overexpression, which induces cancer cell apoptosis and reduces
migration. In the present study, erianin treatment of HeLa cells
could promote the activation of p53.
Apoptosis maintains the healthy survival/death
balance in metazoan cells. Defects in apoptosis can cause cancer
autoimmunity, whereas enhanced apoptosis may cause degenerative
diseases (38). Mitochondria have a
notable role in the intrinsic pathway of mammalian apoptosis. In
various types of cancer, inhibitors of apoptosis are highly
expressed whereas apoptosis promoters are mostly inactivated,
resulting in a certain degree of drug resistance. Thus,
reactivation of the normal apoptosis response in cancer cells
through regulating apoptotic regulators is a desirable treatment
approach (39,40). Developing therapeutic approaches has
involved modifying the activity of Bcl-2 family proteins to
reactivate apoptosis, eradicating cancer cells (41–43). It is
generally recognized that Bcl-2, regarded as an anti-apoptotic
protein, was suppressed and pro-apoptotic proteins like Bax was
overexpressed following cleavage of caspase-family proteins once
cytochrome C was released from the mitochondria into the cytoplasm
(44,45). Evidence indicates provided that
baicalein and betulinic acid induced apoptosis in HeLa cells by
downregulating Bcl-2 expression and upregulating that of expression
and caspase-8. When HeLa cells were exposed to erianin, the
apoptotic rate was increased and expression of Bax was upregulated,
caspase-3 was activated and Bcl-2 levels reduced, which was
consistent with the results of previous studies (46,47),
indicating that apoptosis induced by erianin was associated with
the activation of the proteolytic caspase family and the Bcl-2
family.
Paclitaxel exerts an anticancer function and has
been manufactured into albumin-bound paclitaxel
(nab-paclitaxel), which received US Food and Drug
Administration approval for the treatment of metastatic breast
cancer, non-small cell lung cancer and pancreatic cancer (48). The National Comprehensive Cancer
Network recommends paclitaxel combined with carboplatin or
cisplatin for treatment of recurrent or metastatic cervical cancer.
Therefore, in the current study paclitaxel was used as positive
control agent and was compared with erianin in terms of effects on
HeLa cell proliferation, cell cycle and apoptosis. The results of
this analysis revealed that erianin arrested progression at the
G2/M phase and enhanced the apoptotic rate, similar to
paclitaxel, indicating that erianin has the potential to serve as
an anti-cervical cancer drug.
In conclusion, erianin is a promising anticancer
compound, owing to its ability to inhibit growth of HeLa by
arresting progression at the G2/M phase transition and
inducing apoptosis. The potential mechanism of action involves
regulation of the ERK1/2 signaling and mitochondrial pathways.
Acknowledgements
Not applicable.
Funding
The present study was financially supported by the
National Natural Science Foundation of China (grant no.
J1310034).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
ML performed the cell cycle and apoptosis
experiments. YH performed the cell viability experiments. GH
participated in statistical analyses and wrote the manuscript. XX
and CP designed the research and wrote the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to publish
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tsikouras P, Zervoudis S, Manav B, Tomara
E, Iatrakis G, Romanidis C, Bothou A and Galazios G: Cervical
cancer: Screening, diagnosis and staging. J BUON. 21:320–325.
2016.PubMed/NCBI
|
|
2
|
Bonneau C, Perrin M, Koskas M, Genin AS
and Rouzier R: Epidemiology and risk factors for cancer of the
uterus. Rev Prat. 64:774–779. 2014.(In French). PubMed/NCBI
|
|
3
|
Castle PE and Maza M: Prophylactic HPV
vaccination: Past, present, and future. Epidemiol Infect.
144:449–468. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tang J and Hao F: The present situation
and the future of vaccines for cervical cancer. Immunol J.
26:546–550. 2010.
|
|
5
|
Chinese Pharmacopoeia Committee, .
Pharmacopoeia of China. Chin Med Sci Technol Press; pp. 1–93.
2015
|
|
6
|
Ng TB, Liu F and Wang ZT: Antioxidative
activity of natural products from plants. Life Sci. 66:709–723.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma GX, Xu GJ and Xu LS: Inhibitory effects
of Dendrobium chrysotoxum and its constituents on the mouse HePA
and ESC. J Chin Pharm Univ. 25:188–189. 1994.
|
|
8
|
Li YM, Wang HR and Liu GQ: Erianin induces
apoptosis in human leukemia HL-60 cells. Acta Pharmacol Sin.
22:1018–1022. 2001.PubMed/NCBI
|
|
9
|
Ma GX and LeBlanc GA: The activity of
erianin and chrysotoxine from Dendrobium chrysotoxum to reverse
multidrug resistance in B16/hMDR-1 Cells. J Chin Pharm Sci.
7:142–146. 1998.
|
|
10
|
Zhang W, Yuan W, Xu N, Li J and Chang W:
Icariin improves acute kidney injury and proteinuria in a rat model
of pregnancy-induced hypertension. Mol Med Rep. 16:73982017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cerami EG, Gross BE, Demir E, Rodchenkov
I, Babur O, Anwar N, Schultz N, Bader GD and Sander C: Pathway
Commons, a web resource for biological pathway data. Nucleic Acids
Res. 39:D685–D690. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wawryk-Gawda E, Chylińska-Wrzos P,
Lis-Sochocka M, Chłapek K, Bulak K, Jędrych M and Jodłowska-Jędrych
B: P53 protein in proliferation, repair and apoptosis of cells.
Protoplasma. 251:525–533. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang T, Xu F, Sheng Y, Zhang W and Chen Y:
A targeted proteomics approach to the quantitative analysis of
ERK/Bcl-2-mediated anti-apoptosis and multi-drug resistance in
breast cancer. Anal Bioanal Chem. 408:7491–7503. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Robles AI and Harris CC: p53-mediated
apoptosis and genomic instability diseases. Acta Oncol. 40:696–701.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Long HJ 3rd, Bundy BN, Grendys EC Jr,
Benda JA, McMeekin DS, Sorosky J, Miller DS, Eaton LA and Fiorica
JV; Gynecologic Oncology Group Study, : Randomized phase III trial
of cisplatin with or without topotecan in carcinoma of the uterine
cervix: A gynecologic oncology group study. J Clin Oncol.
23:4626–4633. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kurtz JE, Freyer G, Joly F, Gladieff L,
Kaminski MC, Fabbro M, Floquet A, Hardy-Bessard AC, Raban N,
Ray-Coquard I and Pujade-Lauraine E; GINECO Group, France, .
Combined oral topotecan plus carboplatin in relapsed or advanced
cervical cancer: A GINECO phase I–II trial. Anticancer Res.
32:1045–1049. 2012.PubMed/NCBI
|
|
18
|
Xiong Y, Liang LZ, Cao LP, Min Z and Liu
JH: Clinical effects of irinotecan hydrochloride in combination
with cisplatin as neoadjuvant chemotherapy in locally advanced
cervical cancer. Gynecol Oncol. 123:99–104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peng R and Zhao CQ: Chemotherapy drugs in
the application and research progress of cerical cancer treatment.
Chin Pharm. 24:1143–1146. 2013.
|
|
20
|
Jin ZH, Liao GH and Jiang N: Clinical
analyses on 91 recurrence or metastasis cases of young women
cervical cancer. Pract J Cancer. 21:502–503,511. 2006.
|
|
21
|
Sun J, Fu X, Wang Y, Liu Y, Zhang Y, Hao T
and Hu X: Erianin inhibits the proliferation of T47D cells by
inhibiting cell cycles, inducing apoptosis and suppressing
migration. Am J Transl Res. 8:3077–3086. 2016.PubMed/NCBI
|
|
22
|
Yu Z, Zhang T, Gong C, Sheng Y, Lu B, Zhou
L, Ji L and Wang Z: Erianin inhibits high glucose-induced retinal
angiogenesis via blocking ERK1/2-regulated HIF-1α-VEGF/VEGFR2
signaling pathway. Sci Rep. 6:343062016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Su P, Wang J, An JX, Zhu Q, Lu X and Tang
Y: Inhibitory effect of erianin on hepatocellular carcinoma (HCC)
Huh7 cells. Chin J Appl Environ Biol. 17:662–665. 2011.
|
|
24
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 2011.144:646–674. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
El-Baba C, Mahadevan V, Fahlbusch FB,
Mohan SS, Rau TT, Gali-Muhtasib H and Schneider-Stock R:
Thymoquinone-induced conformational changes of PAK1 interrupt
prosurvival MEK-ERK signaling in colorectal cancer. Mol Cancer.
13:2012014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song X, Wang Y, Du H, Fan Y, Yang X, Wang
X, Wu X and Luo C: Overexpression of HepaCAM inhibits cell
viability and motility through suppressing nucleus translocation of
androgen receptor and ERK signaling in prostate cancer. Prostate.
74:1023–1033. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bai LX, Mao R, Wang J, Ding L, Jiang S,
Gao C, Kang H, Chen X, Sun X and Xu J: ERK1/2 promoted
proliferation and inhibited apoptosis of human cervical cancer
cells and regulated the expression of c-Fos and c-Jun proteins. Med
Oncol. 32:572015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei H, Shenglin M, Lingbin D, et al:
Experimental study of erianin inducing apotosis in gastric
carcinoma SGC-7901. Chin Cancer. 17:499–501. 2008.(In Chinese).
|
|
29
|
Cui XQ, Su P, Zhu QY, An JX, Wang J, Wu J
and Tang YX: Molecular mechanism of apoptosis of human colorectal
cancer SW480 cells induced by Erianin. Chin J Appl Environ Biol.
17:512–516. 2011.
|
|
30
|
Hoogervorst EM, Van SH and De VA:
Nucleotide excision repair- and p53-deficient mouse models in
cancer research. Mutat Res. 574:3–21. 2015. View Article : Google Scholar
|
|
31
|
Haga S, Nakayama M, Tatsumi K, Maeda M,
Imai S, Umesako S, Yamamoto H, Hilgers J and Sarkar NH:
Overexpression of the p53 gene product in canine mammary tumors.
Oncol Rep. 8:1215–1219. 2001.PubMed/NCBI
|
|
32
|
Park YK, Chi SG, Kim YW, Park HR and Unni
KK: P53 mutations in Ewing's sarcoma. Oncol Rep. 8:533–537.
2001.PubMed/NCBI
|
|
33
|
Moskovits N, Kalinkovich A, Bar J, Lapidot
T and Oren M: p53 attenuates cancer cell migration and invasion
through repression of SDF-1/CXCL12 expression in stromal
fibroblasts. Cancer Res. 66:10671–10676. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Khan S, Chib R, Shah BA, Wani ZA, Dhar N,
Mondhe DM, Lattoo S, Jain SK, Taneja SC and Singh J: A cyano
analogue of boswellic acid induces crosstalk between p53/PUMA/Bax
and telomerase that stages the human papillomavirus type 18
positive HeLa cells to apoptotic death. Eur J Pharmacol.
660:241–248. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen ZL, Gu PQ, Liu K, Su YJ and Gao LJ:
The globular heads of the C1q receptor regulate apoptosis in human
cervical squamous carcinoma cells via a p53-dependent pathway. J
Transl Med. 10:1–12. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou MJ, Chen FZ, Chen HC, Wan XX, Zhou X,
Fang Q and Zhang DZ: ISG15 inhibits cancer cell growth and promotes
apoptosis. Int J Mol Med. 39:446–452. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu Y, Li L, Liu Y, Geng P, Li G, Yang Y
and Song H: RECK inhibits cervical cancer cell migration and
invasion by promoting p53 signaling pathway. J Cell Biochem.
4:3058–3066. 2018. View Article : Google Scholar
|
|
38
|
Hassan M, Watari H, AbuAlmaaty A, Ohba Y
and Sakuragi N: Apoptosis and molecular targeting therapy in
cancer. Biomed Res Int. 2014:1508452014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kaufmann SH and Vaux DL: Alterations in
the apoptotic machinery and their potential role in anticancer drug
resistance. Oncogene. 22:7414–7430. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schmitt CA, Rosenthal CT and Lowe SW:
Genetic analysis of chemoresistance in primary murine lymphomas.
Nat Med. 6:1029–1035. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kang MH and Reynolds CP: Bcl-2 inhibitors:
Targeting mitochondrial apoptotic pathways in cancer therapy. Clin
Cancer Res. 15:1126–1132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Patel MR, Masood A, Patel PS and
Chanan-Khan AA: Targeting the Bcl-2. Curr Opin Oncol. 21:516–523.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Leibowitz B and Yu J: Mitochondrial
signaling in cell death via the Bcl-2 family. Cancer Biol Ther.
9:417–422. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hu Y, Ding L, Spencer DM and Núñez G:
WD-40 repeat region regulates Apaf-1 self-association and
procaspase-9 activation. J Biol Chem. 273:33489–33494. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jiang XJ and Wang XD: Cytochrome
C-mediated apoptosis. AnnuRev Biochem. 73:87–106. 2014. View Article : Google Scholar
|
|
46
|
Peng Y, Guo C, Yang Y, Li F, Zhang Y,
Jiang B and Li Q: Baicalein induces apoptosis of human cervical
cancer HeLa cells in vitro. Mol Med Rep. 11:2129–2134. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xu T, Pang Q, Zhou D, Zhang A, Luo S, Wang
Y and Yan X: Proteomic investigation into betulinic acid-induced
apoptosis of human cervical cancer HeLa cells. PLoS One.
9:e1057682014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kundranda MN and Niu J: Albumin-bound
paclitaxel in solid tumors: Clinical development and future
directions. Drug Des Devel Ther. 9:3767–3777. 2015. View Article : Google Scholar : PubMed/NCBI
|