Introduction
A defining characteristic of malignant brain tumors
is their invasive growth and easy recurrence (1,2), and
therefore more effective treatment modalities are urgently
required. A previous study revealed the existence of tumor stem
cells (TSCs), which have the same functions as neural stem cells
inside tumor cells including the ability to self-proliferate
(3). There are a number of
similarities between TSCs and neural stem cells that suggest the
origin of TSCs (4), a variety of
solid tumors, including brain tumors (5) and expression of numerous similar
proteins (6). The discovery of TSCs
has provided novel options for the treatment of brain tumors
(7), including therapies targeting
specific signaling pathways and transcription factors, and niche
therapies targeting cancer stem cells (8,9). TSCs
serve major roles inside tumor tissues, acting as a source for the
proliferation and differentiation of tumors. Even in the presence
of regulatory factors, TSCs continue to proliferate and
differentiate; additionally, the next generation of cells retains
the characteristics of the precursors. These characteristics
include an unlimited proliferation ability, which promotes the
rapid formation of tumor bodies as well as rapid reoccurrence
following surgery, radiotherapy, and chemotherapy (10).
Nestin, a class VI intermediate filament protein,
was originally detected in neural stem cells during development,
and has been shown to be expressed in the transformed cells of
various human malignancies (11). It
is hypothesized to contribute to the aggressive behavior of these
cells and facilitate tumor growth (12).
Cluster of differentiation (CD) 133 is one of the
most commonly used markers of cancer stem cells (CSCs), which are
characterized by their ability for self-renewal and tumorigenicity
(13). Over-expression of CD133 is
associated with solid cancers, many studies have correlated the
overexpression of CD133 with survival, recurrence, metastasis or
therapy resistance (14).
TSCs can regulate the conduction pathways involved
in cell proliferation and differentiation; these pathways include,
Notch (15), Wnt, sonic hedgehog
(SHH), and Bmi-1 (16), which have
important roles in regulating the growth of neural stem cells. A
previous study has demonstrated the important roles of Wnt and the
SHH signaling conduction pathways in regulating the proliferation
of neural stem cells (17). When
tissues are damaged, the self-protective roles of the Wnt and SHH
signaling conduction pathways are activated to repair the damaged
tissues, after which the corresponding pathways are disrupted
through strict regulation. However, when certain genes, including
casein kinase 1 epsilon (18) and sil
(19), are mutated, the Wnt and SHH
pathways can become abnormally activated, enabling cells to undergo
unlimited proliferation and differentiation. The blocking of
differentiation and maturation can induce tumors and abnormalities
in the Wnt and SHH pathways can lead to glioblastoma (20); however, one study has identified that
signal transduction pathway inhibitors can shrink tumors in mice
until they have disappeared and induce rapid cell apoptosis of
medulloblastoma in vitro (21).
The Notch signaling pathway has been conserved
throughout evolution and is involved in cell growth, proliferation,
and differentiation. It not only serves important roles in cell
fate during organ development, but also can be reactivated in
disease states, participating in cell apoptosis, regeneration, and
differentiation through cross-talk with other signaling pathways,
which can affect the occurrence and development of various diseases
(22,23). However, few studies have examined
these issues, and understanding the activities of these pathways is
important for the development of novel cancer therapeutics.
The Notch signaling pathway is associated with
tumorigenesis, and γ-secretase inhibitor II (GSI II) factor
inhibits the activation of Notch1, which is a member of the Notch
signaling pathway (24). In the
present study, TSCs from the human brain tumor cell line U87 were
identified, and then added GSI II to the human brain glioma cell
line U87 for co-culture to observe the growth of TSCs.
Materials and methods
Materials
The cell culture medium contained Dulbecco's
modified Eagles medium combined with Ham's F-12 nutrient mixture
(DMEM/F12, Sigma Aldrich; Merck KGaA, Darmstadt, Germany) and 10%
fetal bovine serum (FBS; Wuhan Boster Biological Technology, Ltd.,
Wuhan, China). Additives included 2% B27, 2% N2, epidermal growth
factor 20 mg/l−1, and basic fibroblast growth factor 20
mg/l−1 (Wuhan Boster Biological Technology, Ltd.).
Antibodies included rabbit anti-human Nestin antibody (cat. no.
n5413; 1:100; Sigma-Aldrich; Merck KGaA), mouse anti-human CD133
antibody (cat. no. o43490; 1:500; Bioworld Technology, Inc., St.
Louis Park, MN, USA), Cy3-labeled goat anti-mouse antibody (cat.
no. BA103; 1:100; Wuhan Boster Biological Technology, Ltd.),
fluorescein isothiocyanate (FITC)-labeled goat anti-rabbit antibody
(cat. no. BA1105; 1:50; Wuhan Boster Biological Technology, Ltd.),
and rabbit anti-human glial fibrillary acidic protein antibody
(GFAP; cat. no. PAB18435; 1:500; Abnova, Tapei, Taiwan). Other
materials included the tumor cell line U87 (U-87 MG
ATCC® HTB14™; cat. no. CBP60299; Nanjing Ke Bai
Biological Technology Co., Ltd., Nanjing, China), 0.01%
poly-lysine, 0.25% trypsin, GSI II (Sigma-Aldrich, Merck KGaA), and
Triton-X100 (Sigma-Aldrich, Merck KGaA).
Culture of TSCs
The tumor cell line U87 (2×106) was added
to the culture media (DMEM/F12) of neural stem cells and cultured
in a humidified incubator at 37°C with 5% CO2. The media
was intermittently exchanged (2–3 days) for 2-week culture period
until spherical TSCs were observed, and immunofluorescence was
performed for identification.
Immunofluorescence
TSCs (1×105) were seeded onto slides with
polylysine pretreatment, 1 ml of 10% acetone was added, and the
slides were placed at −20°C for 10 min. Next, the slices were
soaked with 0.1% Triton-X100 at room temperature for 1 h, gently
rinsed and soaked in 5% goat serum (Wuhan Boster Biological
Technology, Ltd.) at 20°C for 1 h. The CD133 and Nestin antibodies
were added to different groups of slides labeled Group A (CD133)
and B (Nestin), and then incubated at 4°C for 24 h. Next, 1 ml 1%
Cy3-labeled goat anti-mouse antibody was added to group A and 1 ml
1% FITC-labeled goat anti-rabbit antibody was added to group B.
Following an incubation at room temperature overnight, the slices
were mounted with glycerol and cell staining was observed under a
fluorescence microscope (magnification, ×200). To the control
group, neural stem cell culture medium was added to replace the
CD133 and Nestin antibodies; the other steps were identical to
those described previously.
Differentiation induction of TSCs
Cultured TSCs were transferred into 10%
FBS-containing 24-well culture plates, and cultured at 37°C for ~10
days to observe their growth. When the TSCs clusters appeared as
differentiated wall-adherent cells after ~10 days,
immunofluorescence was performed to identify the cells; the primary
antibody was changed to GFAP, and the secondary antibody was FITC,
and the remaining steps were conducted as described previously.
Impact of GSI II on TSC growth
Cultured U87 cells were gently dispersed using a
pipette and the concentration was adjusted to 105
cells/ml with 10% FBS. Next, 2 ml cell suspension was added to each
well of 6-well culture plates (the bottom of each plate had been
evenly coated with 0.01% poly-lysine); culture was conducted in a
humidified incubator at 37°C with 5% CO2 for 2 days
until the cells had completely adhered to the walls, after which
the culture solution was removed. Stem cell culture medium (2 ml)
was added and the four plates were labeled A1, A2, B1 and B2. Next,
0.01 MPBS was added to A1 and A2 as the control group, and GSI II
(0.2 µg per well) was added to B1 and B2 to observe changes in
growth and the number of cell clusters in each culture plate was
recorded. After a 20-day culture in the previously described
culture conditions, the culture plates were directly stained as
previously described.
Statistical analysis
The number of Nestin-positive cell clusters was
recorded and SPSS 10.0 (SPSS, Inc., Chicago, IL, USA) was used to
perform an analysis of variance followed by Fisher's least
significant difference to compare the differences between different
groups.
Results
Cells differentiated from U87
cells
The U87 cells were extracted and cultured in neural
stem cell culture medium; after 2 days, wall-adherent cells began
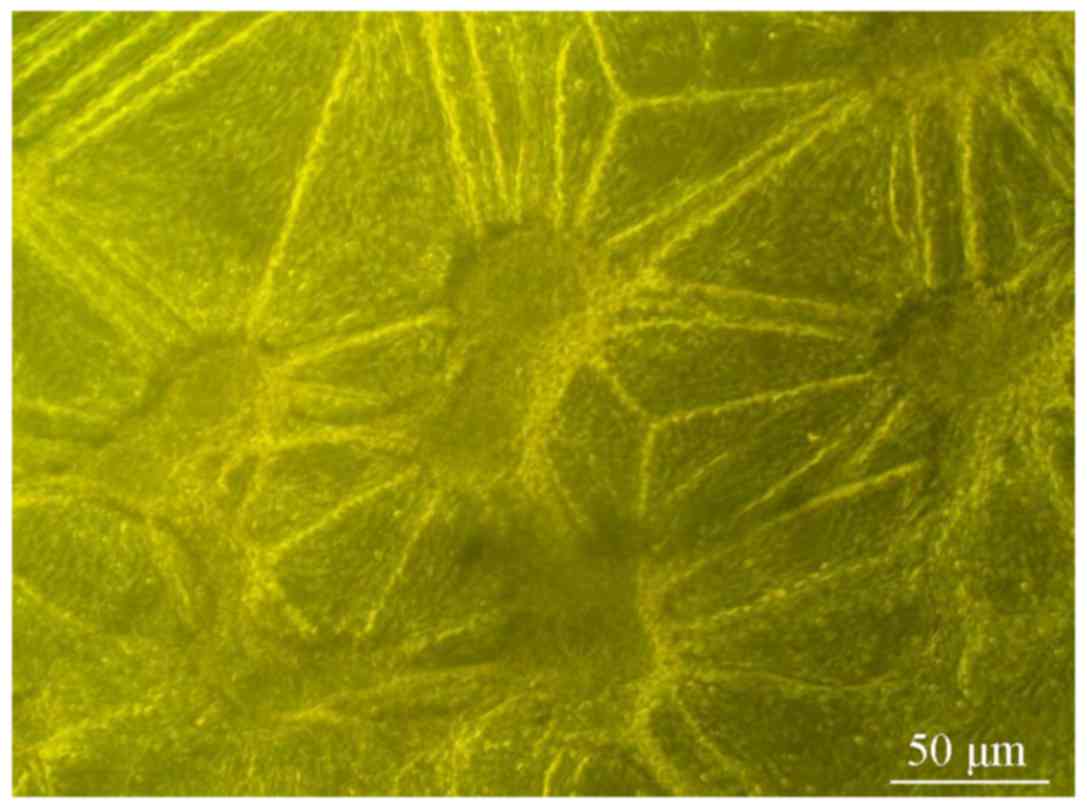
to grow in a cluster shape. After 4 days, the cell clusters
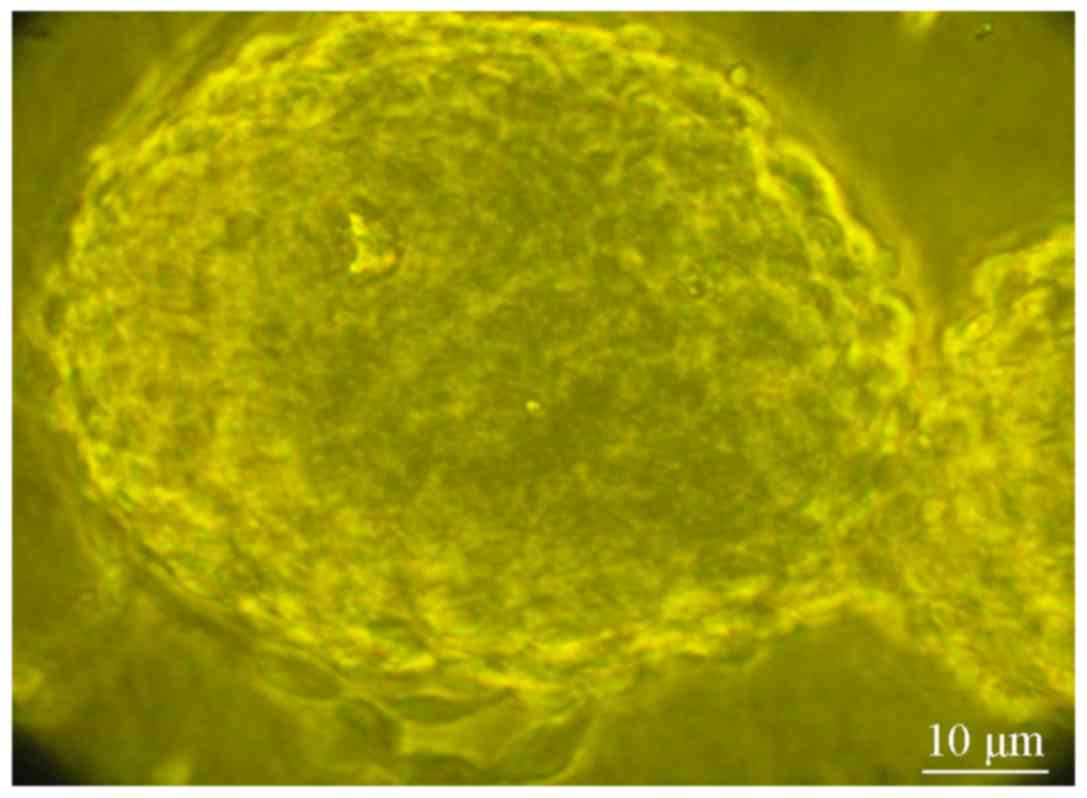
gradually increased in volume and began to appear spherical in
shape (Fig. 1). The spherical cells
were re-dispersed and seeded into neural stem cell culture medium.
Several days later, spherical cell structures had reformed.
Positive staining under a fluorescence
microscope
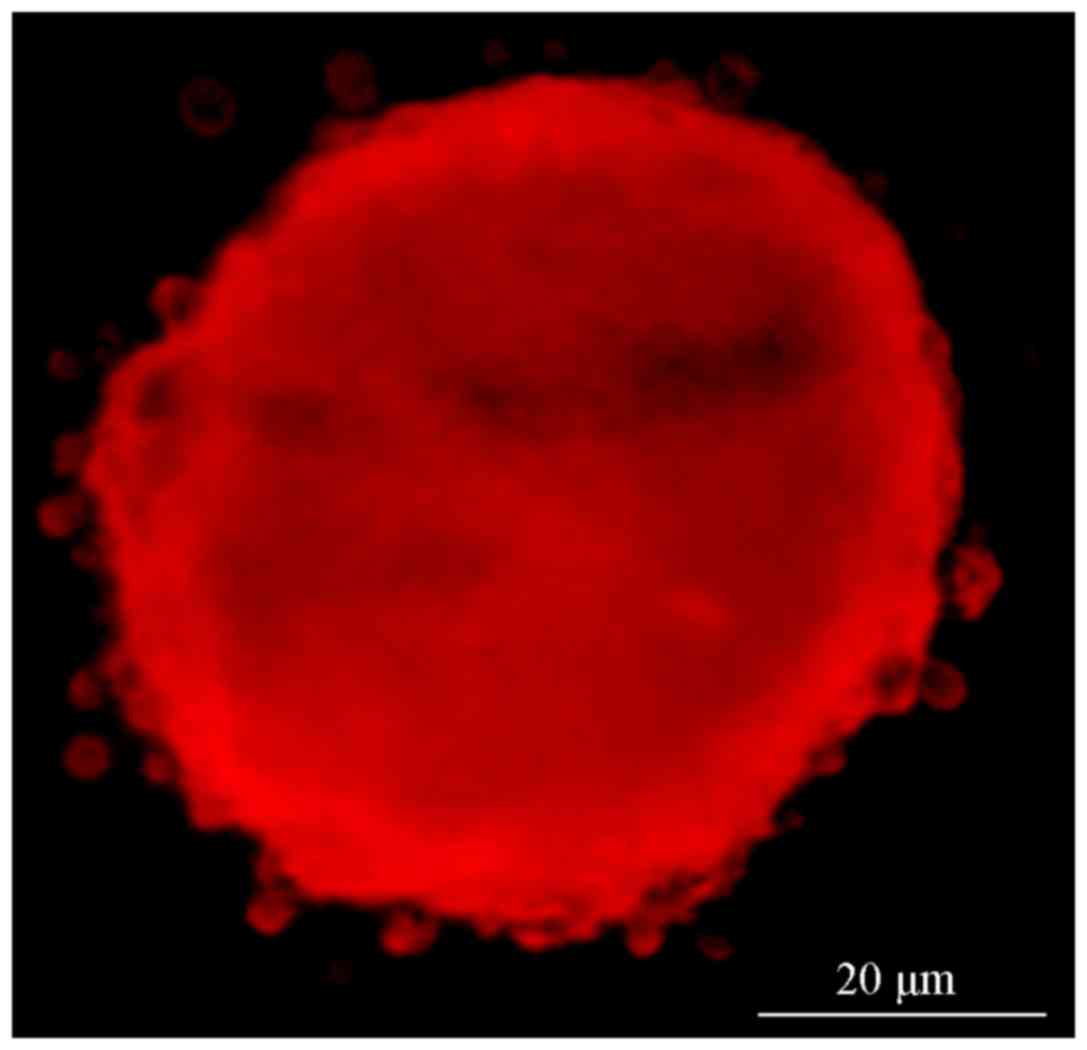
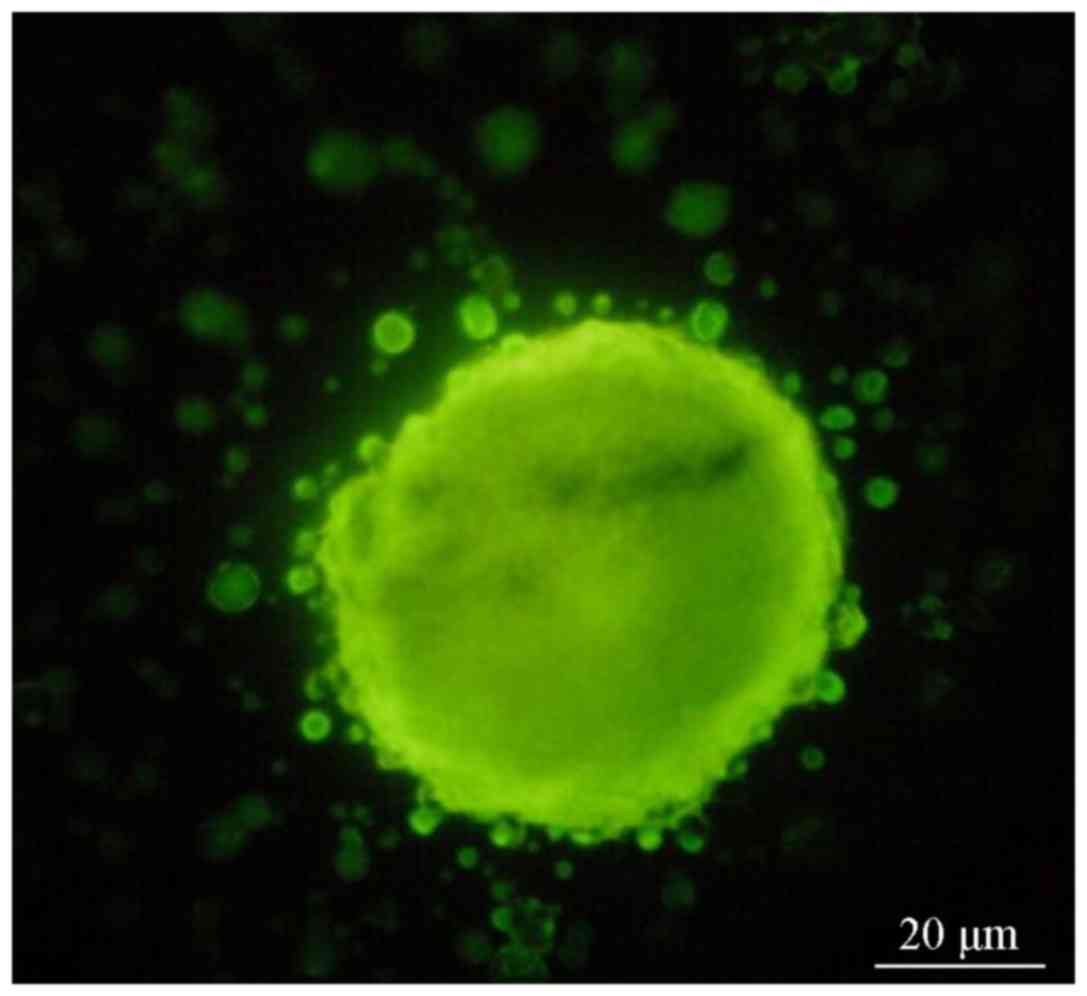
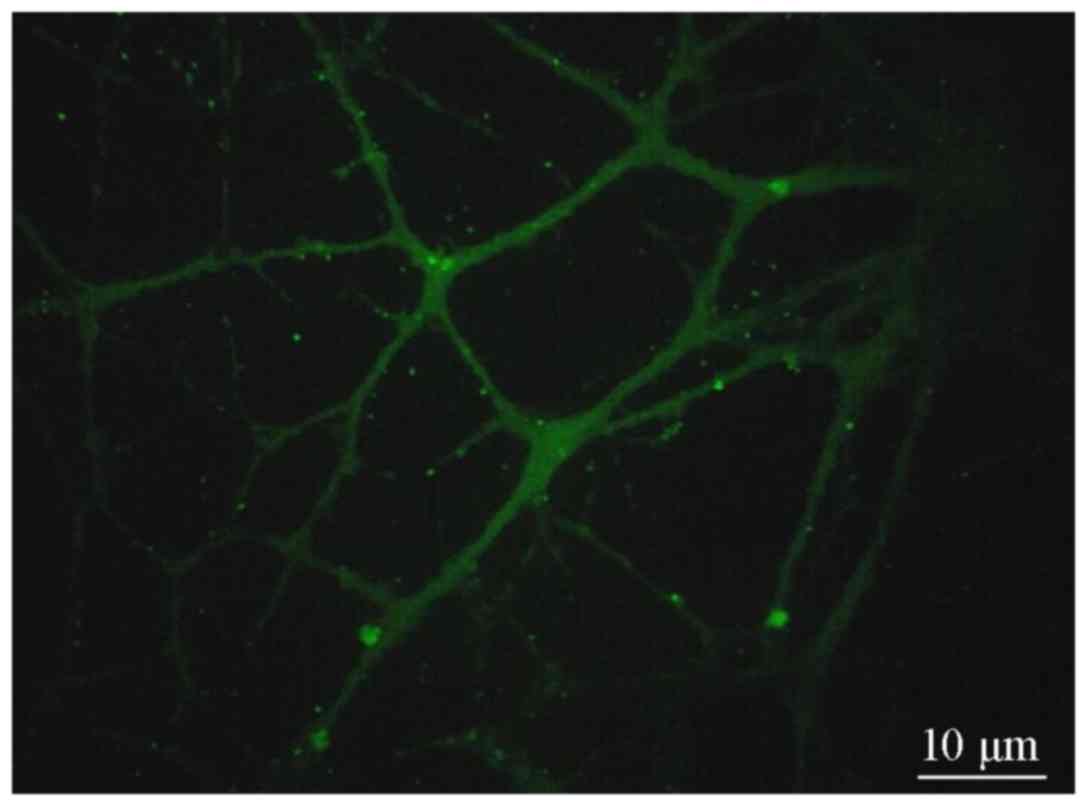
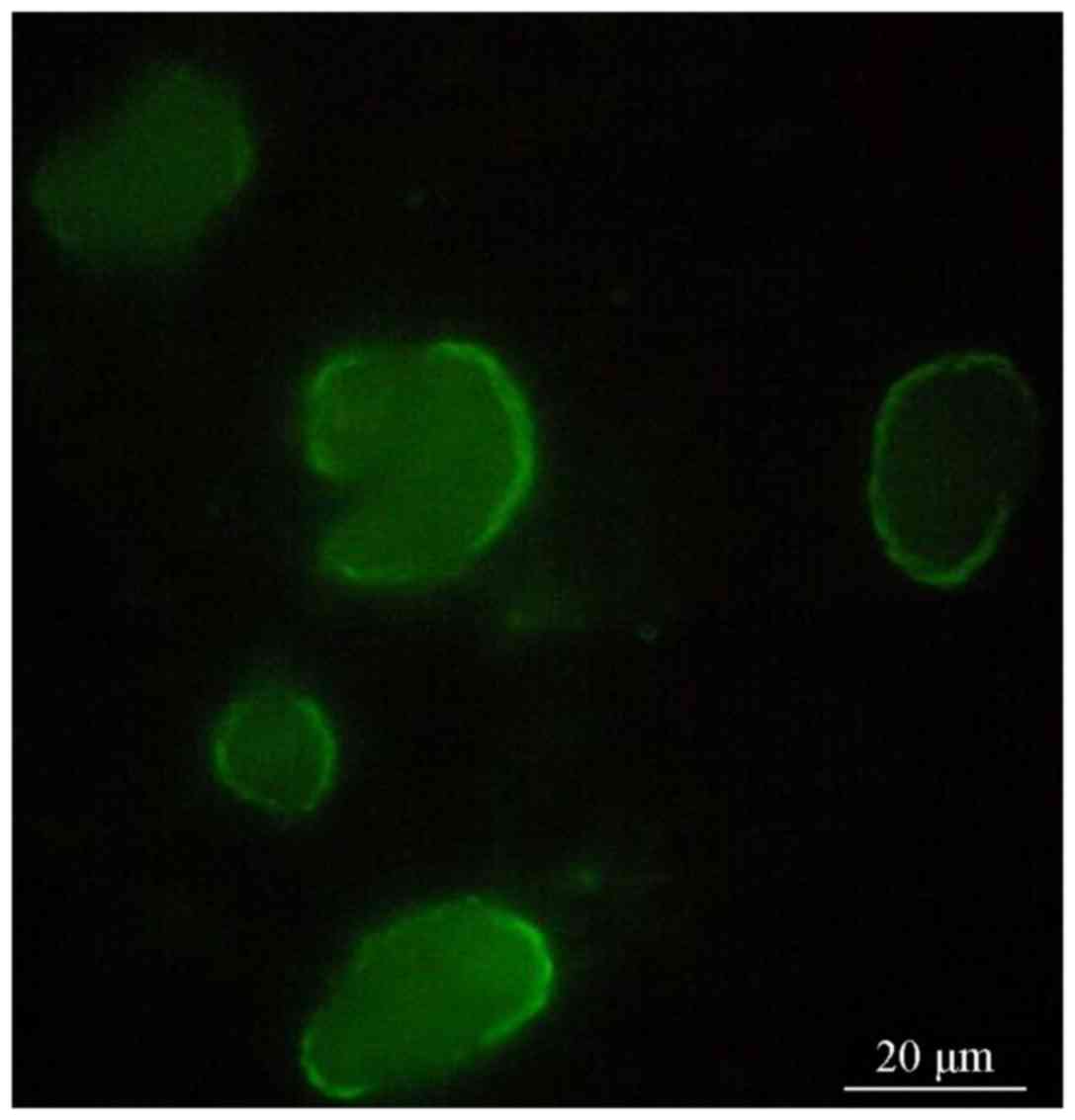
Fluorescence microscopy revealed that the cell
clusters in groups A and B exhibited red (Fig. 2) and green fluorescence, (Fig. 3), indicating that the CD133 and Nestin
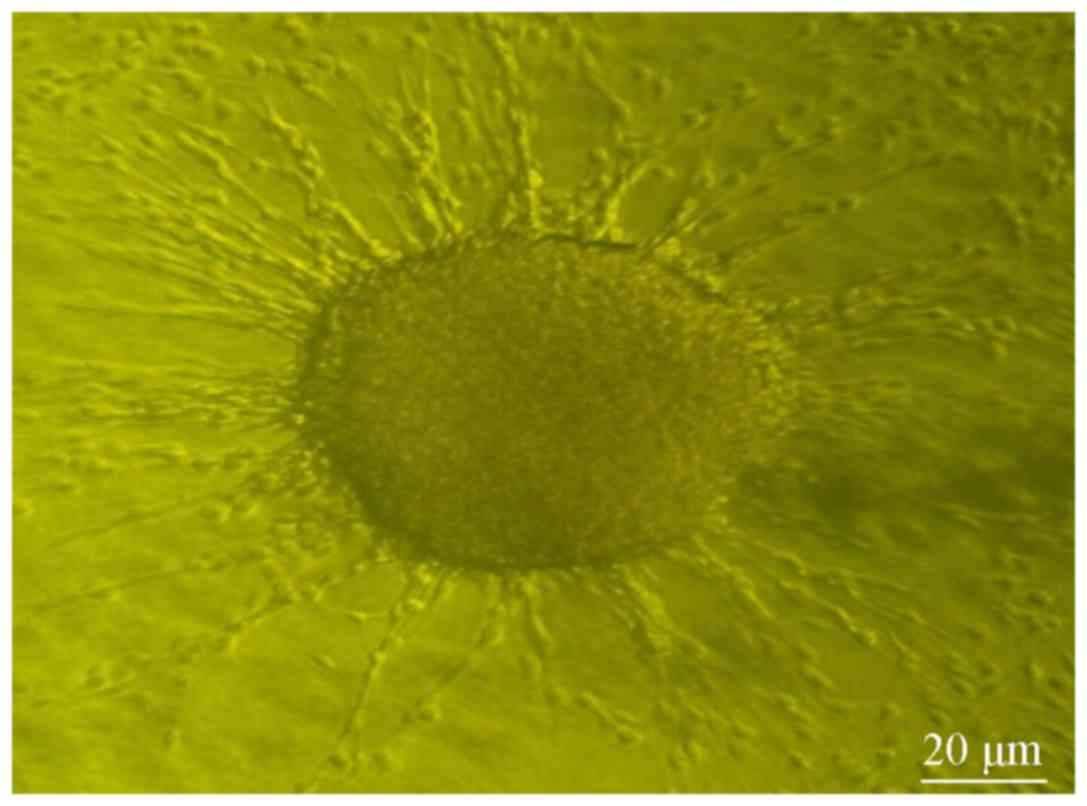
antigens were located inside the cells. The cell clusters were then
placed into FBS medium for 10-day culture. Some cell clusters
became wall-adherent again and presented a spindle shape; the cells
also showed a high proliferation rate (Fig. 4). The wall-adherent cells, which were
differentiated near the adjacent cell clusters, gradually exhibited
cord-like connections (Fig. 5). The
differentiated cells were positive for GFAP staining (Fig. 6) and showed the same characteristics
as glial cells.
Comparison of TSC growth rates between
the 2 groups
The proliferation rates of cells in groups B1 and
B2, which were incubated with GSI II, were significantly lower than
those in the control groups A1 and A2 (P<0.01, Table I). Single stem cell clusters were
positive for Nestin staining (Fig.
7), which confirmed the cells were tumor stem cells.
 | Table I.The numbers of Nestin-positive cell
clusters of the 2 groups. |
Table I.
The numbers of Nestin-positive cell
clusters of the 2 groups.
| Group |
Numbers of
Nestin-positive cell clusters | mean ± SD | t | P-value |
|---|
| Control group
(A1+A2) | 13 | 12 | 13 | 10 | 14 | 12 | 16 | 9 | 14 | 12 | 6 | 14 | 16 | 8 | 11 | 10 | 15 | 7 | 11.78±2.96 | 2.83 | <0.05 |
| Experimental group
(B1+B2) | 11 | 10 | 9 | 10 | 12 | 10 | 7 | 11 | 13 | 9 | 11 | 11 | 7 | 9 | 6 | 8 | 6 | 9 |
9.39±2.96 |
|
|
Discussion
Brain tumors have been suggested to originate from
glial cells containing gene mutations that undergo differentiation
and maturation (25). The study of Hu
et al (26) suggested that
TSCs are present inside tumor tissues and that these cells can
proliferate and differentiate into tumor cells. The study of
Alamgeer et al (27) extracted
and cultured tumor cells from lung and breast cancer and found TSCs
showed differentiation ability. Fong et al (28) cultured spherical TSCs from mouse TSCs
and found that both TSC types could grow in a suspension in culture
media and retained the ability to differentiate into tumor cells
after several passages. In the study of Lucarelli et al
(29) tumor tissues were extracted
from high-degree malignant tumors and the cell clusters that
exhibited a similar growth state as stem cells were cultured. These
previous studies only indirectly revealed the existence of TSCs, as
no specific antigens were available for TSCs. At present, there are
two principal markers that can be used for this purpose, including
CD133, which is a transmembrane protein specific to neural stem
cells and the majority of previous studies have used CD133 in order
to identify glioma stem cells (28,29). CD133
is highly expressed in glioma cells, and the ratio of CD133+ cells
in glioma was demonstrated to be associated with the pathological
type and prognosis (30). The other
antigen used is against Nestin, a cytoskeletal protein that
supports neural stem cells and is specific to neural stem cells
(31). The U87 cell line was used in
the present study, after several passages, neural stem cells were
mixed with U87 cells, and these cells were used for the
immunofluorescent identification of cell clusters as TSCs.
TSCs can proliferate and differentiate into tumor
cells, and if the intermediate link is blocked, tumor growth can be
inhibited (32). Notch1 is highly
expressed in glioma cells and is closely associated with a tumor's
pathological grade (33). GSI II
effectively inhibits Notch1 signal activation (34). In the present study, GSI II was
demonstrated to effectively inhibit TSC generation in U87 cells
(P<0.05); additionally, a recent study demonstrated that the
resistance of TSCs towards the tumor necrosis factor related
apoptosis-inducing ligand could be induced, and its activation may
increase the effects of chemotherapeutic drugs (35).
Although a previous study demonstrated that the U87
cell line is not authentic to the tumor of the original cell
established in the University of Uppsala in 1968, it is also a
glioblastoma cell line (36). The
purpose of the present study was to isolate and cultivate TSCs, and
examine the effect of GSI II on the growth and differentiation of
TSCs. Therefore, the U87 cell line from ATCC or the U87 cell line
from Uppsala University could be used in the present study and
without affecting the results.
The present study provides a foundation for
developing therapeutics for the treatment of glioma. The selective
killing of TSCs in brain tumors may prevent tumor recurrence and
metastasis, providing a means for curing or preventing tumor
recurrence and metastasis.
Acknowledgements
Not applicable.
Funding
This study was funded by Shandong Provincial Natural
Science Foundation (grant no. ZR2012HM084).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XDD and CQD conceived and designed the study. XDD,
CQD, FW, WSD, MMY, QHM and PS performed the experiments. XDD and
MMY conducted the statistical analysis. XDD and CQD wrote the
paper. PS reviewed and edited the manuscript. All authors read and
approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lowenstein PR and Castro MG: Evolutionary
basis of a new gene- and immune-therapeutic approach for the
treatment of malignant brain tumors: From mice to clinical trials
for glioma patients. Clin Immunol. 189:43–51. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Baker GJ, Yadav VN, Motsch S, Koschmann C,
Calinescu AA, Mineharu Y, Camelo-Piragua SI, Orringer D, Bannykh S,
Nichols WS, et al: Mechanisms of glioma formation: Iterative
perivascular glioma growth and invasion leads to tumor progression,
VEGF-independent vascularization, and resistance to antiangiogenic
therapy. Neoplasia. 16:543–561. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nakano I and Kornblum HI: Methods for
analysis of brain tumor stem cell and neural stem cell
self-renewal. Methods Mol Biol. 568:37–56. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Turner JD and Sanai N: A brain tumor stem
cell origin for glioblastoma endothelium. World Neurosurg.
75:574–575. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Khan IS and Ehtesham M: Isolation and
characterization of stem cells from human central nervous system
malignancies. Adv Exp Med Biol. 853:33–47. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bryukhovetskiy A, Shevchenko V, Kovalev S,
Chekhonin V, Baklaushev V, Bryukhovetskiy I and Zhukova M: To the
novel paradigm of proteome-based cell therapy of tumors: Through
comparative proteome mapping of tumor stem cells and
tissue-specific stem cells of humans. Cell Transplant. 23 Suppl
1:S151–S170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Toda M: Identification of antigen against
brain tumor stem cell and application for treatment of glioma. Gan
To Kagaku Ryoho. 37:1016–1018. 2010.(In Japanese). PubMed/NCBI
|
|
8
|
Tanaka S, Louis DN, Curry WT, Batchelor TT
and Dietrich J: Diagnostic and therapeutic avenues for
glioblastoma: No longer a dead end? Nat Rev Clin Oncol. 10:14–26.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lathia JD, Li M, Hall PE, Gallagher J,
Hale JS, Wu Q, Venere M, Levy E, Rani MR, Huang P, et al: Laminin
alpha 2 enables glioblastoma stem cell growth. Ann Neurol.
72:766–778. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sherriff J, Tamangani J, Senthil L,
Cruickshank G, Spooner D, Jones B, Brookes C and Sanghera P:
Patterns of relapse in glioblastoma multiforme following
concomitant chemoradiotherapy with temozolomide. Br J Radiol.
86:201204142013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Neradil J and Veselska R: Nestin as a
marker of cancer stem cells. Cancer Sci. 106:803–811. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tampaki EC, Nakopoulou L, Tampakis A,
Kontzoglou K, Weber WP and Kouraklis G: Nestin involvement in
tissue injury and cancer-a potential tumor marker? Cell Oncol
(Dordr). 37:305–315. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yiming L, Yunshan G, Bo M, Yu Z, Tao W,
Gengfang L, Dexian F, Shiqian C, Jianli J, Juan T and Zhinan C:
CD133 overexpression correlates with clinicopathological features
of gastric cancer patients and its impact on survival: A systematic
review and meta-analysis. Oncotarget. 6:42019–42027. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kemper K, Grandela C and Medema JP:
Molecular identification and targeting of colorectal cancer stem
cells. Oncotarget. 1:387–395. 2010.PubMed/NCBI
|
|
15
|
Garcia A and Kandel JJ: Notch: A key
regulator of tumor angiogenesis and metastasis. Histol Histopathol.
27:151–156. 2012.PubMed/NCBI
|
|
16
|
Ye L, Wang C, Yu G, Jiang Y, Sun D, Zhang
Z, Yu X, Li X, Wei W, Liu P, et al: Bmi-1 induces radioresistance
by suppressing senescence in human U87 glioma cells. Oncol Lett.
8:2601–2606. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wilson NH and Stoeckli ET: Sonic Hedgehog
regulates Wnt activity during neural circuit formation. Vitam Horm.
88:173–209. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Foldynová-Trantírková S, Sekyrová P,
Tmejová K, Brumovská E, Bernatík O, Blankenfeldt W, Krejcí P,
Kozubík A, Dolezal T, Trantírek L and Bryja V: Breast
cancer-specific mutations in CK1epsilon inhibit Wnt/beta-catenin
and activate the Wnt/Rac1/JNK and NFAT pathways to decrease cell
adhesion and promote cell migration. Breast Cancer Res. 12:R302010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Izraeli S, Lowe LA, Bertness VL, Campaner
S, Hahn H, Kirsch IR and Kuehn MR: Genetic evidence that Sil is
required for the Sonic Hedgehog response pathway. Genesis.
31:72–77. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jin X, Jeon HM, Jin X, Kim EJ, Yin J, Jeon
HY, Sohn YW, Oh SY, Kim JK, Kim SH, et al: The ID1-CULLIN3 axis
regulates intracellular SHH and WNT signaling in glioblastoma stem
cells. Cell Rep. 16:1629–1641. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Filbin MG, Dabral SK, Pazyra-Murphy MF,
Ramkissoon S, Kung AL, Pak E, Chung J, Theisen MA, Sun Y,
Franchetti Y, et al: Coordinate activation of Shh and PI3K
signaling in PTEN-deficient glioblastoma: New therapeutic
opportunities. Nat Med. 19:1518–1523. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou QZ, Zhang G, Long HB, Lei F, Ye F,
Jia XF, Zhou YL, Kang JP and Feng DX: Effect of spinal cord
extracts after spinal cord injury on proliferation of rat embryonic
neural stem cells and Notch signal pathway in vitro. Asian Pac J
Trop Med. 7:562–567. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen CY, Liao W, Lou YL, Li Q, Hu B, Wang
Y and Deng ZF: Inhibition of Notch signaling facilitates the
differentiation of human-induced pluripotent stem cells into neural
stem cells. Mol Cell Biochem. 395:291–298. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Saito N, Fu J, Zheng S, Yao J, Wang S, Liu
DD, Yuan Y, Sulman EP, Lang FF, Colman H, et al: A high Notch
pathway activation predicts response to γ secretase inhibitors in
proneural subtype of glioma tumor-initiating cells. Stem Cells.
32:301–312. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Glass R and Synowitz M: CNS macrophages
and peripheral myeloid cells in brain tumours. Acta Neuropathol.
128:347–362. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hu M, Xiang FX and He YF: Are cancer stem
cells the sole source of tumor? J Huazhong Univ Sci Technolog Med
Sci. 34:621–625. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alamgeer M, Peacock CD, Matsui W, Ganju V
and Watkins DN: Cancer stem cells in lung cancer: Evidence and
controversies. Respirology. 18:757–764. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fong D, Yeh A, Naftalovich R, Choi TH and
Chan MM: Curcumin inhibits the side population (SP) phenotype of
the rat C6 glioma cell line: Towards targeting of cancer stem cells
with phytochemicals. Cancer Lett. 293:65–72. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lucarelli G, Galleggiante V, Rutigliano M,
Vavallo A, Ditonno P and Battaglia M: Isolation and
characterization of cancer stem cells in renal cell carcinoma.
Urologia. 82:46–53. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lehnus KS, Donovan LK, Huang X, Zhao N,
Warr TJ, Pilkington GJ and An Q: CD133 glycosylation is enhanced by
hypoxia in cultured glioma stem cells. Int J Oncol. 42:1011–1017.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brazel CY, Alaythan AA, Felling RJ,
Calderon F and Levison SW: Molecular features of neural stem cells
enable their enrichment using pharmacological inhibitors of
survival-promoting kinases. J Neurochem. 128:376–390. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tamura K, Aoyagi M, Ando N, Ogishima T,
Wakimoto H, Yamamoto M and Ohno K: Expansion of CD133-positive
glioma cells in recurrent de novo glioblastomas after radiotherapy
and chemotherapy. J Neurosurg. 119:1145–1155. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fang KM, Lin TC, Chan TC, Ma SZ, Tzou BC,
Chang WR, Liu JJ, Chiou SH, Yang CS and Tzeng SF: Enhanced cell
growth and tumorigenicity of rat glioma cells by stable expression
of human CD133 through multiple molecular actions. Glia.
61:1402–1417. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim SH and Singh SV: Mammary cancer
chemoprevention by withaferin A is accompanied by in vivo
suppression of self-renewal of cancer stem cells. Cancer Prev Res
(Phila). 7:738–747. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang X, Chen T, Zhang J, Mao Q, Li S,
Xiong W, Qiu Y, Xie Q and Ge J: Notch1 promotes glioma cell
migration and invasion by stimulating β-catenin and NF-κB signaling
via AKT activation. Cancer Sci. 103:181–190. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xiao YG, Wang W, Gong D and Mao ZF:
γ-Secretase inhibitor DAPT attenuates intimal hyperplasia of vein
grafts by inhibition of Notch1 signaling. Lab Invest. 94:654–662.
2014. View Article : Google Scholar : PubMed/NCBI
|