Introduction
Although the incidence of cervical cancer has
decreased in developed countries due to the wide spread use of pap
smear screening and the launch of HPV vaccinations, it is still the
second most commonly diagnosed cancer and the third leading cause
of cancer death among women of less developed countries.
Epidemiological study estimated that there were 527,600 new cases
and 265,700 deaths worldwide in 2012 (1). The major treatment strategy for cervical
cancer at an early stage of disease is radical surgery. Radiation
therapy is often used when patients are diagnosed at advanced
stages or under unresectable conditions. Chemotherapy is usually
used concurrently with radiation and inevitably regarded as an
adjuvant therapy modality (2).
New therapeutic strategies for cervical cancer are
currently being investigated. Among them, target therapy,
particularly the anti-vascular endothelial growth factors agents,
have been added to current chemotherapeutic regimens to treat
advanced and recurrent cervical cancer with significant benefits
being reported (3). In addition, the
value of complementary and alternative medicines is also being
investigated. For example, some herbal medicines have been reported
to effectively induce apoptosis of cervical cancer cells and to
have the potential for cancer treatment (4–7).
Butein (2′,3,4,4′-tetrahydroxychalcone) is a plant
polyphenol and an bioactive component extractable from the
heartwood of Dalbergia odorifera, Caragana jubata, and
Rhus verniciflua Stokes, and the stem bark of cashews
(Semecarpus anacardium). These plant extracts have been
reported to exert various pharmacological effects including
anti-oxidant and anti-inflammatory activity, and they have long
been used as a traditional herbal medicine in Asian countries
(8–11)
and claimed to have therapeutic potentials for chronic diseases,
such as liver tuberculosis, obesity, diabetes and hypertension
(12). Mechanistically, butein has
been shown to be a specific protein tyrosine kinase inhibitor by
repressing autophosphorylation level of epidermal growth factor
receptor in HepG2 cells (13). In the
context of butein-exhibited anti-cancer activity, previous in
vitro pharmacological studies have shown that butein inhibits
cell proliferation and induces apoptosis in numerous types of tumor
cells, including lung (14), liver
(15), pancreas (16), colon (17), bladder (18), prostate (19), breast (16), melanoma tumors (20), as well as ovarian (21) and cervical cancers (22). Moreover, butein is found to suppress
migration and invasion of bladder (14), breast, and pancreatic cancer cells
(15). In vivo studies further
confirm that butein is able to inhibit the growth of prostate
(19), breast and pancreatic tumors
(16) in human tumor xenograft nude
mouse models. In addition, butein also inhibits pulmonary
metastasis of B16F10 melanoma cells in mice, mainly via decreasing
angiogenic factor production (23).
The effect of butein on cervical carcinogenesis has
been demonstrated by the studies using HPV18-containing HeLa cells.
It is previously reported that butein suppresses proliferation and
migration of HeLa cells, and inhibits growth of xenograft tumor
cell in nude mouse model (22).
Furthermore, butein combination treatment increases the sensitivity
of HeLa cells to cisplatin in both in vitro and in
vivo settings, while the underlying mechanism involves
inhibition of AKT and MAPK pathways (24). Despite previous studies demonstrating
involvement of apoptosis regulators in butein-induced cell death of
cervical cancer cells, little is known about whether it also
modulates the expression of inhibitor of apoptosis (IAP) proteins.
The aim of this study was to examine its inhibitory ability on the
growth of two human cervical cancer cell lines, C-33A (HPV
negative) and SiHa (HPV16 positive) cells. The effects of butein on
cytotoxicity, pro-apoptotic caspase activation and expression
changes of various apoptosis regulators were examined in these two
cervical cancer cell lines.
Materials and methods
Reagents
Butein pure compound was purchased from Enzo Life
Sciences (Farmingdale, NY, USA; cat no. ALX-350-246). Butein was
stocked at 30 mM in dimethylsulfoxide (DMSO) and stored at −80°C
until use. The concentrations of DMSO in solvent control and all
butein-treated groups were equal to or <1% and showed no notable
effect on cell viability. Dulbecco's modified Eagle's medium (DMEM;
cat no. 11995-065), fetal bovine serum (FBS; cat no. 10437-028) and
gentamicin (cat no. 15750-060) were obtained from Gibco (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Primary antibodies
against the Bcl-2 family proteins [Bax (1:1,000; cat no. 5023); Bak
(1:1,000; cat no. 12105); Bcl-xL (1:1,000; cat no. 2764)], cellular
IAP 1 (cIAP-1; 1:1,000; cat no. 7065), x-linked IAP (XIAP; 1:1,000;
cat no. 2045), survivin (1:1,000; cat no. 2808), poly (ADP-ribose)
polymerase (PARP; 1:1,000; cat no. 9532) and actin (1:5,000; cat
no. 3700) were purchased from Cell Signaling Technology Inc.
(Danvers, MA, USA). Goat anti-rabbit and goat anti-mouse secondary
antibodies were purchased from Cell Signaling Technology (1:2,000;
cat no. 7074) and Perkin-Elmer (NEF822001EA; 1:2,000; Boston, MA,
USA), respectively.
Cell lines and cell culture
Two human cervical cancer cell lines were examined
in this study. The C-33A cell line was obtained from the National
Health Research Institute Cell Bank (Hsinchu, Taiwan), and the SiHa
cell line was obtained from the American Type Culture Collection
(Manassas, VA, USA). Both cell lines were cultured in DMEM medium
supplemented with 10% FBS and gentamicin (50 µg/ml). Both cell
lines were maintained in a standard cell culture incubator at 37°C
and in a humidified 95% air/5% CO2 atmosphere.
3-(4,5-Dimethylthiazol-2-yl)-2,5-iphenyltetrazolium bromide) (MTT)
cytotoxicity assay
Cervical cancer cells were trypsinized and seeded
onto a 96-well plate at 5,000 cells per well for 24-h incubation.
The cells were then exposed to butein at various concentrations (0,
10, 30, 100 and 300 µM) for different incubation periods (24, 48,
and 72 h). Cells that were not treated with butein were used as
controls. After a volume of 50 µl of 1 mg/ml MTT solution (cat no.
M5655) was added and incubated for 4 h at 37°C, the solution was
removed and 100 µl of DMSO (cat no. D4540; both Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) was added to lyse the cells. The
MTT-formazan product was read with a microplate reader (Labsystems,
Helsinki, Finland) at 550 nm. The cytotoxic effect at each
concentration of butein was determined by calculating the
IC50 value with the control group as 100%. Each assay
was performed in triplicate.
Mitochondrial transmembrane potential
(MTP) assay
MTP assay was performed using BD™ MitoScreen (JC-1),
Flow Cytometry Mitochondrial Membrane Potential Detection kit (cat
no. 551320; BD Biosciences, Franklin Lakes, NJ, USA) according to
the manufacturer's instructions. The aggregation of fluorescent
lipophilic cationic probe JC-1 in mitochondria is sensitive for MTP
detection. C-33A and SiHa cells were seeded onto 6-cm dishes for 24
h and exposed to different concentrations of butein (0, 10, 30, and
100 µM) for another 24 h. In brief, cells were harvested, washed,
and re-suspended in PBS. The cells were then incubated with JC-1
working solution for 10 min at 37°C. The cells were then washed
with assay buffer and analyzed using Accuri C6 flow cytometer
(Beckton Dickinson, Ann Arbor, MI, USA). The aggregated dye located
in the unaffected mitochondria emitted red florescence, whereas in
cells with damaged MTP, the monomeric dye remained in cytoplasm
showed diffuse green fluorescence (25).
Caspase-3, −8, and −9 activity
assay
The caspase activity was detected using caspase-3,
−8 and −9 colorimetric assay kit (cat nos. BF3100, BF4100 and
BF10100; R&D Systems Inc., Minneapolis, MN, USA) according to
the manufacturer's protocol. Briefly, C-33A and SiHa cells were
treated with butein at various concentrations (0, 10, 30, and 100
µM) and cultured for 24 h. The cells were then harvested, washed
twice with PBS, and centrifuged at 150 g for 5 min. Cell lysates
were then incubated on ice for 10 min, and centrifuged at 10,000 g
for 1 min. The lysates (100 µg of total protein) were added to the
reaction mixtures in a final volume of 50 µl, containing 5 µl of
colorimetric substrate peptides specific for either caspase-3
(DEVD-pNA), caspase-8 (IETD-pNA), or caspase-9 (LEHD-pNA). The
mixtures were incubated at 37°C for an additional 2 h. Finally, the
absorbance at 405 nm was measured on a microplate reader, and the
respective non-butein treated cells were used as controls. The
results were shown as the induction fold over the values of
controls.
Cytochrome c assay
A cytochrome c ELISA kit (cat no.
ALX-850-261; Enzo Life Sciences Inc.) was used to examine the
release of cytochrome c from mitochondria in butein treated
cells. Briefly, cervical cancer cells were incubated with three
concentrations of butein for 24 h. The cells were then collected
and washed with PBS, permeabilized with digitonin-containing buffer
on ice for 5 min. The cells were centrifuged, and the supernatants
were collected and further diluted with the assay buffer. The
samples were then placed on a cytochrome c ELISA plate and
allowed to incubate for 1 h at 500 rpm. The cytochrome c
conjugate was added and incubated for an additional 30 min. The
substrate solution for color development was added, and incubated
at room temperature for 45 min. After adding stop solution, the
concentration of cytochrome c in the cytosol was measured
and read at 405 nm on a microplate reader.
Western blot analysis
C-33A and SiHa cells were seeded in a 10-cm dish.
After butein treatment, total cellular proteins were collected by
lysing the cell pellets in RIPA buffer. A BCA protein assay kit
(cat no. 71285; Novagen; Merck KGaA) was used to determine the
protein concentrations using bovine serum albumin as standards.
Sodium dodecyl sulfate (SDS) polyacrylamide gel electrophoresis,
electrotransferration, and immunodetection were performed as
previously described (5). Equal
amount of total protein (50 µg) in each lane was initially loaded
for gel resolution. An enhanced chemiluminescence detection system
(cat no. NEL103001EA; Perkin Elmer Inc.) was used to detect
immunoreactive signals, followed by densitometric analysis using
Image J software (NIH, Bethesda, MD, USA). Signals for the
housekeeping gene product β-actin were regarded internal controls.
Three experiments were independently performed.
Statistical analysis
All quantitative data are shown as mean ± SD. The
statistical significance of difference was calculated using
Mann-Whitney U test when comparing between two groups or
using the Kruskal-Wallis test followed by Dunn's post hoc test when
comparing more than two groups. SPSS version 22 (IBM Corp., Armonk,
NY, USA)was used for all statistical analyses. P<0.05 was
considered to indicate a statistically significant difference.
Results
Cytotoxic effect of butein in C-33A
and SiHa cervical cancer cells
An MTT-based cytotoxicity assay was used to
determine the cytotoxicity of butein in cervical cancer cells. The
results revealed that butein reduced the viability of C-33A and
SiHa cells in a dose- and time-dependent manner. Compared to the
cells that were not treated with butein, a progressive decrease in
the cell viability of the C-33A and SiHa cells was observed with
increasing dosage of butein (Fig. 1A and
B). After treatment with butein at 30 µM for 24 h, the
viability of C-33A cells decreased to 48% of the controls
(P<0.05), and further decreased to 5.3% at 72 h (P<0.01). The
viability of SiHa cells also attenuated to 62 and 22% of the
controls when exposed to butein at 100 µM for 24 and 72 h,
respectively (P<0.05). The IC50 values of
butein-treated C-33A and SiHa cells are presented in Table I. The difference in IC50
values between these two cell lines (P<0.05) suggested that
butein induced higher cytotoxicity in C-33A cells than that in SiHa
cells.
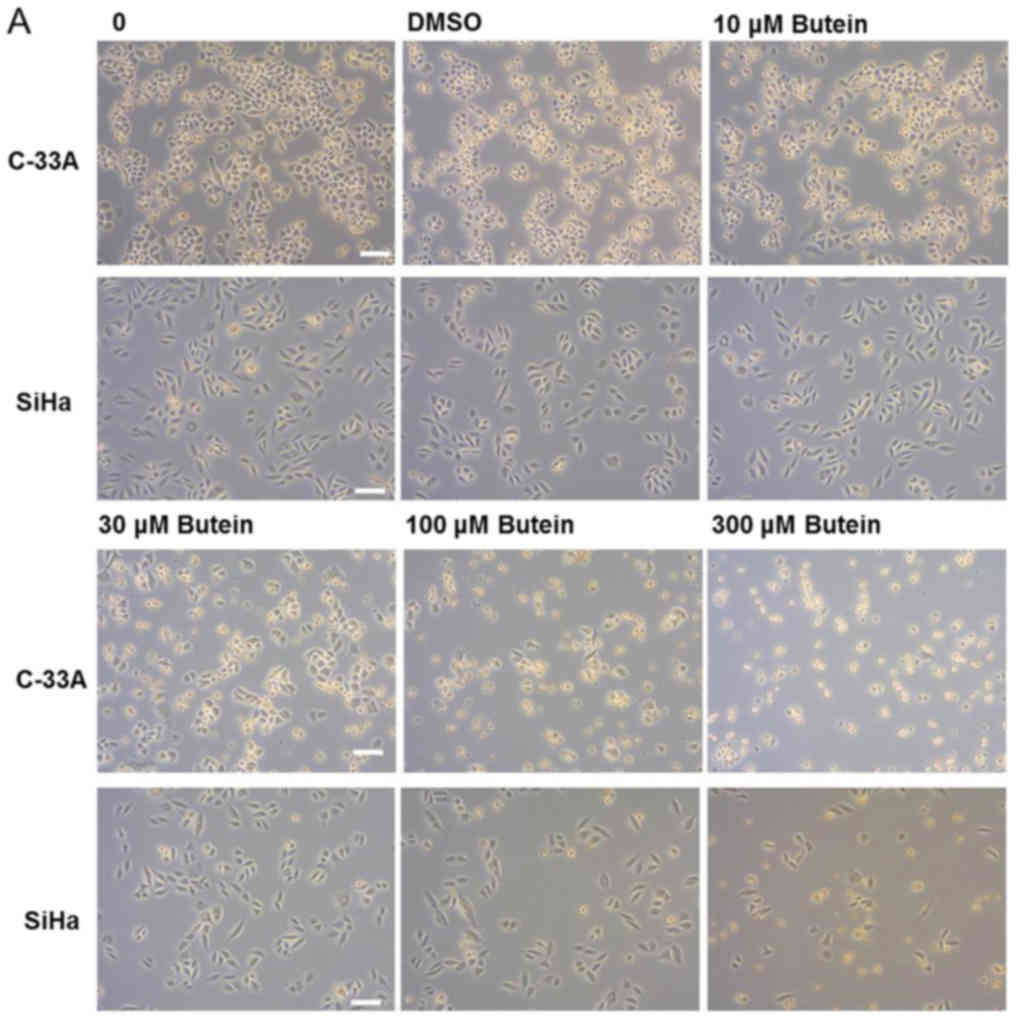 | Figure 1.Cytotoxic effect of butein in C-33A
and SiHa cervical cancer cells. (A) Phase-contrast photomicrograph
of cervical cancer cells, C-33A and SiHa, treated with various
concentrations of butein for 24 h. (B) Both cells were treated with
0, 10, 30, 100 and 300 µM of butein for 24, 48 and 72 h and
subjected to an MTT-based cytotoxicity assay. Scale bar, 100 µm.
Data are presented as means ± SD from three independent
experiments. Asterisks indicate a significant difference compared
with the control group (*P<0.05 and **P<0.01). DMSO,
dimethylsulfoxide; MTT,
3-(4,5-dimethylthiazol-2-yl)-2,5-iphenyltetrazolium bromide. |
 | Table I.Cytotoxicity of butein in C-33A and
SiHa cervical cancer cells with various treatment durations. |
Table I.
Cytotoxicity of butein in C-33A and
SiHa cervical cancer cells with various treatment durations.
| Cell line | 24 h (µM) | 48 h (µM) | 72 h (µM) |
|---|
| C-33A |
79.88±7.45a |
9.95±3.38a |
8.30±1.32a |
| SiHa | 185.00±23.48 | 30.54±2.57 | 24.29±5.09 |
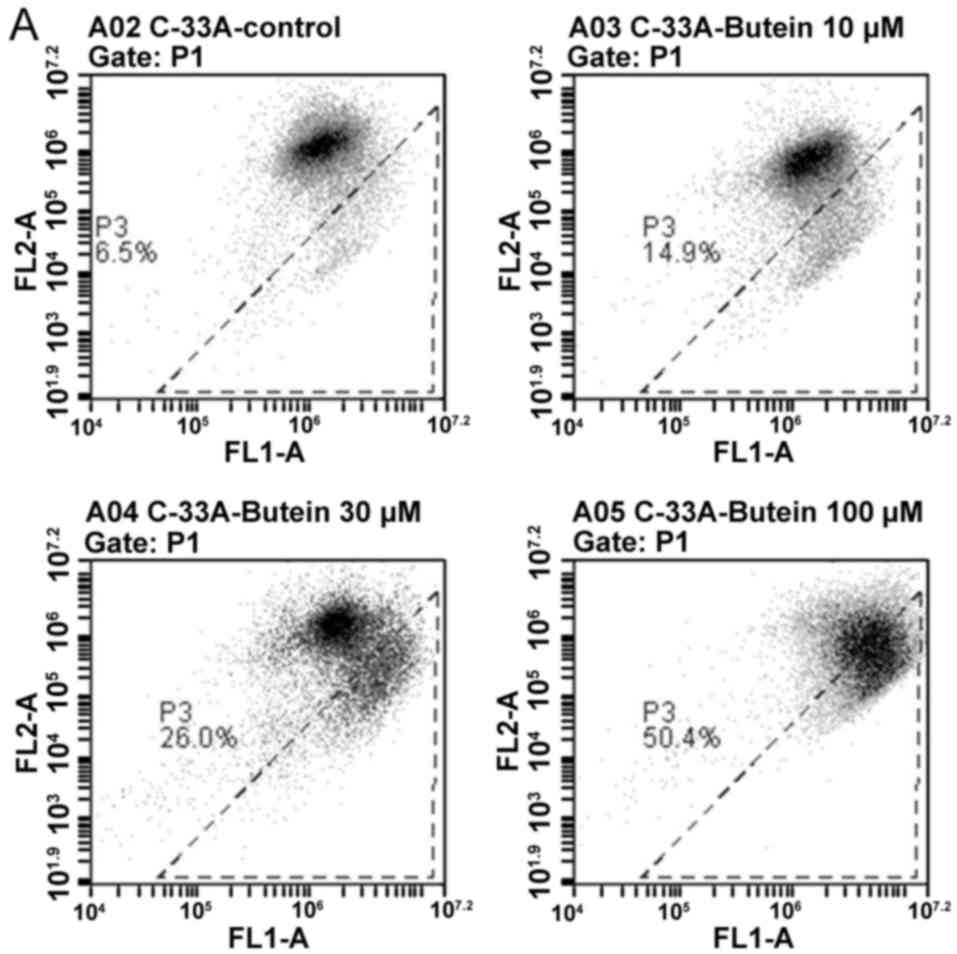
Damaging effect of butein on MTP
To measure the disrupting effect of butein on MTP in
cervical cancer cells, the cells were stained with JC-1, which
undergoes a potential-dependent aggregation in mitochondria and
emits red fluorescence. On the contrary, the monomeric JC-1 dye
remains in cytosol and emits green fluorescence, reflecting
mitochondrial membrane depolarization. The representative results
of flow cytometry shown in Fig. 2
clearly indicated that a dose-dependent increase in the percentage
of green fluorescence-positive cells was noted in the
butein-treated cervical cancer cells, from 6.5% in the controls to
50.4 % in the C-33A cells, and from 6.4% in the controls to 27.1%
in the SiHa cells, respectively (Fig. 2A
and B) (P<0.05). The results indicated that butein
significantly induced a loss of MTP in the cervical cancer
cells.
Involvement of extrinsic and intrinsic
pathways in butein-induced apoptosis
To clarify the pharmacologic action of butein on the
apoptosis of cervical cancer cells, the regulators of extrinsic and
intrinsic pro-apoptotic pathways were examined in butein-treated
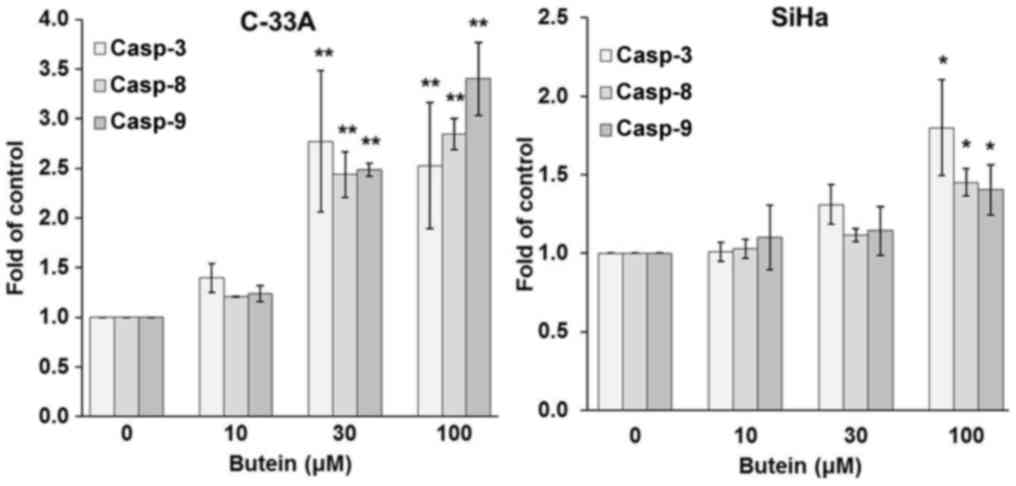
C-33A and SiHa cells. The caspase activity assay revealed that
butein at doses equal to and higher than 30 µM resulted in
significant increases in the activities of caspase-3, −8, and −9 in
C-33A, while butein at concentrations up to 100 µM prominently
increased the activities of all these caspases in SiHa cells
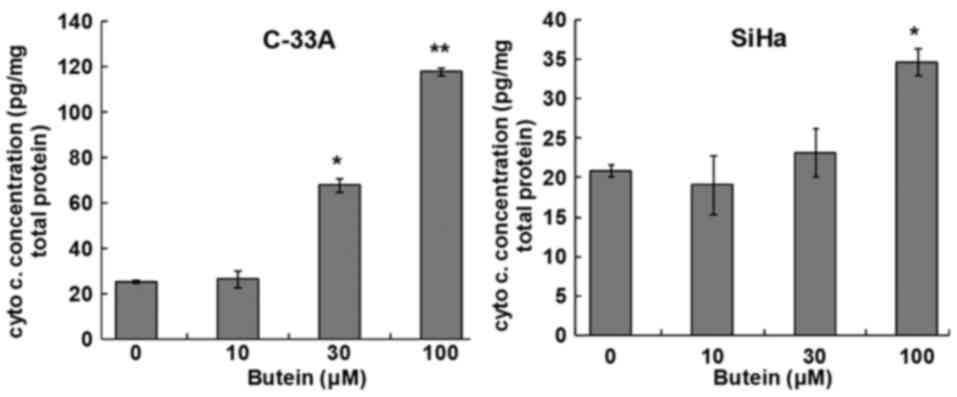
(Fig. 3). The results of cytochrome
c assay demonstrated that butein also dose-dependently
increased cytosolic contents of cytochrome c in both
cervical cancer cell lines (Fig. 4).
These findings suggested that extrinsic and intrinsic signaling
pathways were both involved in the butein-induced apoptosis.
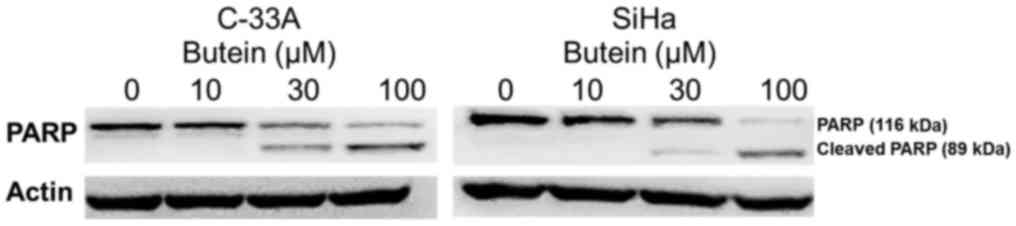
Additionally, western blot analysis indicated that exposure of
C-33A and SiHa cells to butein at 30 µM or higher resulted in the
proteolytic cleavage of PARP (Fig.
5), further supporting that butein induced apoptosis of both
cervical cancer cells.
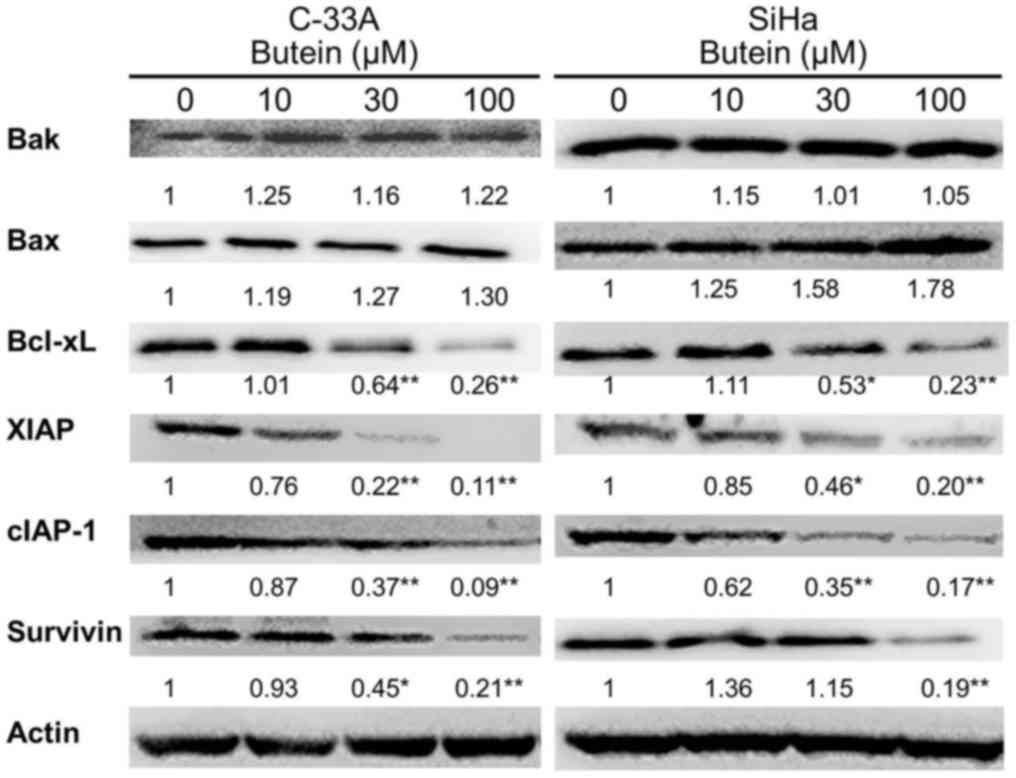
Modulation of butein on anti- and
pro-apoptotic regulator expression
To assess whether Bcl-2 family proteins are affected
in the butein-elicited apoptosis, both anti- and pro-apoptotic
members of the Bcl-2 family members were measured in butein-treated
C-33A and SiHa cells. Western blot analysis clearly showed that
butein induced a significant downregulation of the anti-apoptotic
Bcl-xL protein at 30 µM in C-33A and SiHa cells, although butein at
100 µM did not significantly change the expressions of Bak and Bax
pro-apoptotic proteins (Fig. 6) and
anti-apoptotic Bcl-2 in both cell lines (data not shown).
Downregulation effect of butein on IAP
proteins
Western blot analysis further showed that the IAP
proteins in both C-33A and SiHa cells were downregulated after
treatment with butein (Fig. 6). The
expressions of XIAP and cIAP-1 in both C-33A and SiHa cells
significantly decreased when exposed to butein at 100 µM
(P<0.01). Downregulation of survivin was also noted in C-33A and
SiHa cells with butein treatment at 100 µM (P<0.05). The cIAP-2
expression levels in both cells were not affected by butein
treatment (data not shown).
Discussion
Butein is a member of the chalcone family of open
chain flavonoids that are extracted from medicinal plants and known
to modulate protein tyrosin kinase activity. Butein has recently
attracted attention due to its anticancer activity. Numerous in
vitro and in vivo studies have shown that an increasing
number of cancers could be inhibited by butein (15–21). These
studies have identified that butein induces cancer cell death
through induction of apoptosis. A clinical trial reported that the
flavonoid Rhus verniciflua, which mainly contains butein,
could decrease the tumor size of gastric cancer in an elderly
patient and was well-tolerated (26).
In the present study, we examined whether butein has
an anti-tumor effect on cervical cancer. Our results revealed that
butein reduced cell viability in the two cervical cancer cells,
C-33A and SiHa, in a dose- and time-dependent manner. Consistent
with our findings, the cytotoxicity of butein in HeLa cells has
been previously demonstrated. Butein inhibited cervical cancer cell
viability by the PI3K/AKT/mTOR and ERK/p38 MAPK pathway (22,24). This
study evaluated the in vitro chemosensitivity of two
cervical cancer cell lines with difference HPV infection status
after treatment with butein. The significantly different
IC50 values between two tumor cells in response to
butein suggest that butein is more sensitive in inhibiting HPV
negative than that HPV16 positive cervical cancer cells.
The characteristic products of apoptosis, caspase-3
and the proteolytic cleavage of PARP, were increased in a
dose-dependent manner in the butein-treated cervical cancer cells.
Therefore, apoptosis appears to be an important mechanism by which
butein exerts its anti-tumor effect on cervical cancer cells.
Caspase activation cascade is a major signaling pathway involved in
apoptotic cell death. There are two distinct routes to activate the
caspase cascade, one from the cell surface (extrinsic pathway) and
the other from mitochondria (intrinsic pathway). The extrinsic
pathway involves activation of cell membrane receptors, adaptor
proteins, and caspase-8, while activation of the intrinsic pathway
from mitochondria requires Apaf-1, caspase-9 and cytosolic
cytochrome c. Both routes can activate down-stream caspase-3
and lead to cell apoptosis (27).
Consistent with the results observed in the butein-treated C-33A
cells, Bai et al, reported that butein induced HeLa cells
apoptosis through increasing caspase-3, −8 and −9 activities
(22).
The Bcl-2 family proteins regulate apoptosis through
the intrinsic mitochondrial pathway (28), and the ratio of pro-apoptotic to
anti-apoptotic Bcl-2 proteins is known to critically regulate the
apoptotic processes. In this study, butein decreased the expression
of anti-apoptotic Bcl-xL protein in both C-33A and SiHa cells,
suggesting that butein may reduce the ratio of anti-apoptotic to
pro-apoptotic Bcl-2 proteins subsequently leading to mitochondrial
change. In previous reports, butein decreased Bcl-2 anti-apoptotic
and increased pro-apoptotic expression in various cancer cells
(12). In addition, the results of
MTP, cytochrome c, and caspase assays revealed that butein
damaged MTP, increased the level of cytosolic cytochrome c,
and increased the activity of caspase-9. These findings further
support that butein induces apoptosis via the intrinsic pathway.
The caspase assay revealed that the activity of caspase-8 was
increased, and therefore butien-induced apoptosis in the cervical
cancer cells may also involve the extrinsic pathway.
IAP proteins are a family of proteins that play an
important role in carcinogenesis because they can cause cancer cell
proliferation and survival by inhibiting apoptotic induction and
providing resistance to programmed cell death in cancer cells
(29). Recently, therapeutic
strategies have been designed to target the IAP proteins as a
potential cancer treatment (30–33). There
are eight members in human IAP proteins, of which XIAP is the most
physiologically relevant direct inhibitor of caspase-3, −7, and −9
(34). Another two IAP protein
members, cIAP-1 and cIAP-2, function as E3 ligases and promote RIP1
ubiquitination, which is associated with the pro-survival kinase
TAK1 and facilitates cancer cell survival. Conversely,
deubiquitinated RIP1 functions as a proapoptotic adaptor that binds
to and activates caspase-8 and thereby induces cancer cell
apoptosis (35). Recent evidence
indicates that higher expression of IAP proteins has been
considered as one of characteristics of cancer stem-like cells and
proposed as a therapeutic target through simultaneous activation of
caspase-3/7 and autophagy flux (36).
Survivin is involved in controlling cell division and inhibiting
apoptosis (37). Upregulation of
survivin has been reported to confer chemoresistance and contribute
to tumor cell survival in various types of carcinomas, and
therefore to be responsible for a poor prognosis (38). In this study, the downregulated
expression of XIAP, cIAP-1, and survivin by butein strongly
suggests that butein may induce apoptosis in cervical cancer cells
through inhibiting the IAP protein expression and simultaneous
activation of pro-apoptotic caspases. Intriguingly, targeting
XIAP/caspase-7 complex has been found to effectively kill
caspase-3-deficient malignancies, providing an opportunity to treat
the resistant tumors with caspase-3 downregulation (39). Whether butein treatment also triggers
caspase-7 activation via XIAP interaction in cervical cancer cells
awaits further elucidation.
In addition to its pro-apoptotic activity, butein
has long been known to modulate immune activity of the hosts with
chronic diseases (8,9,11). The
molecular mechanism underlying the butein-induced anti-inflammatory
effect has been identified to involve NF-κB signaling activation in
many types of cells (40). In the
context of its antitumor activity, butein is reported to
downregulate metastatic protease and angiogenic factor expression
of prostate cancer cells (41) and
chemokine receptor expression and function of breast and pancreatic
tumor cells (16), mainly through
inactivating NF-κB signaling. Increasing evidence also supports the
notion that anti-inflammatory drugs may also confer anti-neoplastic
effect against uterine cervical cancer or HPV-dependent neoplasia
(42). This raises the possibility
that the combination strategy of using anti-inflammatory and
conventional anti-tumor drugs may confer higher sensitivity to some
types of tumors. In fact, butein has been known to sensitize
cisplatin-induced cytotoxicity in HeLa cervical cancer cells
(24) and TRAIL-induced hepatoma cell
apoptosis (43). Further elucidation
on the in vivo efficacy of butein and the combination
therapy with conventional anti-tumor agent in cervical cancer
treatment may warrant its translation to a clinical setting.
This in vitro study demonstrated that butein
can decrease viability and induce apoptosis in cervical cancer
cells at relatively low concentrations. The butein-induced
apoptosis mechanistically involves activation of both extrinsic and
intrinsic pathways, which may be regulated by inhibition of the IAP
proteins. These findings suggest that butein may be a promising
agent for the treatment of cervical cancer. Further in vivo
and clinical studies are needed to verify its anticancer effect. In
addition, the possible interactions of butein with other currently
used anticancer drugs should be investigated.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from Show
Chwan Memorial Hospital, Taiwan (grant no. RA-16001) and part of
this content was presented at 2016 IGCS biennial meeting, Lisbon,
Portugal.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PYY, DNH and FSL designed the study. PYY and YHK
performed the experiments. PYY, DNH and ICL analyzed the data. ICL
performed the statistical analysis. All authors read and approved
the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HPV
|
human papillomavirus
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
FBS
|
fetal bovine serum
|
|
DMSO
|
dimethyl sulfoxide
|
|
PBS
|
phosphate buffered saline
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-iphenyltetrazolium bromide
|
|
PARP
|
poly (ADP-ribose) polymerase
|
|
IAP
|
inhibitors of apoptosis protein
|
|
XIAP
|
X-linked inhibitor of apoptosis
protein
|
|
cIAP
|
cellular inhibitor of apoptosis
protein
|
|
MTP
|
mitochondrial transmembrane
potential
|
|
SDS
|
sodium dodecyl sulfate
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hacker NF and Vermorken JB: Cervical
cancerBerek and Hacker's Gynecologic Oncology. 5 edition. Berek JS
and Hacker NF: Lippincott Williams and Wilkins; Philadelphia: pp.
336–378. 2015
|
|
3
|
Tewari KS, Sill MW, Long HJ III, Penson
RT, Huang H, Ramondetta LM, Landrum LM, Oaknin A, Reid TJ, Leitao
MM, et al: Improved survival with bevacizumab in advanced cervical
cancer. N Engl J Med. 370:734–743. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gao H, Lamusta J, Zhang WF, Salmonsen R,
Liu Y, O'Connell E, Evans JE, Burstein S and Chen JJ: Tumor Cell
Selective Cytotoxicity and Apoptosis Induction by an Herbal
Preparation from Brucea javanica. N Am J Med Sci (Boston). 4:62–66.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takara K, Horibe S, Obata Y, Yoshikawa E,
Ohnishi N and Yokoyama T: Effects of 19 herbal extracts on the
sensitivity to paclitaxel or 5-fluorouracil in HeLa cells. Biol
Pharm Bull. 28:138–142. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang PY, Hu DN and Liu FS: Cytotoxic
effect and induction of apoptosis in human cervical cancer cells by
Antrodia camphorata. Am J Chin Med. 41:1169–1180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou Y, Liu YE, Cao J, Zeng G, Shen C, Li
Y, Zhou M, Chen Y, Pu W, Potters L and Shi YE: Vitexins,
nature-derived lignan compounds, induce apoptosis and suppress
tumor growth. Clin Cancer Res. 15:5161–5169. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chan SC, Chang YS, Wang JP, Chen SC and
Kuo SC: Three new flavonoids and antiallergic, anti-inflammatory
constituents from the heartwood of Dalbergia odorifera. Planta Med.
64:153–158. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jung CH, Kim JH, Hong MH, Seog HM, Oh SH,
Lee PJ, Kim GJ, Kim HM, Um JY and Ko SG: Phenolic-rich fraction
from Rhus verniciflua Stokes (RVS) suppress inflammatory response
via NF-kappaB and JNK pathway in lipopolysaccharide-induced RAW
264.7 macrophages. J Ethnopharmacol. 110:490–497. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee JC, Lim KT and Jang YS: Identification
of Rhus verniciflua Stokes compounds that exhibit free radical
scavenging and anti-apoptotic properties. Biochim Biophys Acta.
1570:181–191. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Song Z, Shanmugam MK, Yu H and Sethi G:
Butein and its role in chronic diseases. Adv Exp Med Biol.
928:419–433. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Padmavathi G, Roy NK, Bordoloi D, Arfuso
F, Mishra S, Sethi G, Bishayee A and Kunnumakkara AB: Butein in
health and disease: A comprehensive review. Phytomedicine.
25:118–127. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang EB, Zhang K, Cheng LY and Mack P:
Butein, a specific protein tyrosine kinase inhibitor. Biochem
Biophys Res Commun. 245:435–438. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Ma C, Qian M, Wen Z, Jing H and Qian
D: Butein induces cell apoptosis and inhibition of cyclooxygenase-2
expression in A549 lung cancer cells. Mol Med Rep. 9:763–767. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rajendran P, Ong TH, Chen L, Li F,
Shanmugam MK, Vali S, Abbasi T, Kapoor S, Sharma A, Kumar AP, et
al: Suppression of signal transducer and activator of transcription
3 activation by butein inhibits growth of human hepatocellular
carcinoma in vivo. Clin Cancer Res. 17:1425–1439. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chua AW, Hay HS, Rajendran P, Shanmugam
MK, Li F, Bist P, Koay ES, Lim LH, Kumar AP and Sethi G: Butein
downregulates chemokine receptor CXCR4 expression and function
through suppression of NF-κB activation in breast and pancreatic
tumor cells. Biochem Pharmacol. 80:1553–1562. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yit CC and Das NP: Cytotoxic effect of
butein on human colon adenocarcinoma cell proliferation. Cancer
Lett. 82:65–72. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang L, Chen W and Li X: A novel
anticancer effect of butein: Inhibition of invasion through the
ERK1/2 and NF-kappa B signaling pathways in bladder cancer cells.
FEBS Lett. 582:1821–1828. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Khan N, Adhami VM, Afaq F and Mukhtar H:
Butein induces apoptosis and inhibits prostate tumor growth in
vitro and in vivo. Antioxid Redox Signal. 16:1195–1204. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cui Z, Song E, Hu DN, Chen M, Rosen R and
McCormick SA: Butein induces apoptosis in human uveal melanoma
cells through mitochondrial apoptosis pathway. Curr Eye Res.
37:730–739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang PY, Hu DN, Lin IC and Liu FS: Butein
shows cytotoxic effects and induces apoptosis in human ovarian
cancer cells. Am J Chin Med. 43:769–782. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bai X, Ma Y and Zhang G: Butein suppresses
cervical cancer growth through the PI3K/AKT/mTOR pathway. Oncol
Rep. 33:3085–3092. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lai YW, Wang SW, Chang CH, Liu SC, Chen
YJ, Chi CW, Chiu LP, Chen SS, Chiu AW and Chung CH: Butein inhibits
metastatic behavior in mouse melanoma cells through VEGF expression
and translation-dependent signaling pathway regulation. BMC
Complement Altern Med. 15:4452015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang L, Yang X, Li X, Li C, Zhao L, Zhou
Y and Hou H: Butein sensitizes HeLa cells to cisplatin through the
AKT and ERK/p38 MAPK pathways by targeting FoxO3a. Int J Mol Med.
36:957–966. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cossarizza A, Baccarani-Contri M,
Kalashnikova G and Franceschi C: A new method for the
cytofluorimetric analysis of mitochondrial membrane potential using
the J-aggregate forming lipophilic cation
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolcarbocyanine
iodide (JC-1). Biochem Biophys Res Commun. 197:40–45. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee SH, Choi WC, Kim KS, Park JW, Lee SH
and Yoon SW: Shrinkage of gastric cancer in an elderly patient who
received Rhus verniciflua Stokes extract. J Altern Complement Med.
16:497–500. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cho SG and Choi EJ: Apoptotic signaling
pathways: Caspases and stress-activated protein kinases. J Biochem
Mol Biol. 35:24–27. 2002.PubMed/NCBI
|
|
28
|
Frenzel A, Grespi F, Chmelewskij W and
Villunger A: Bcl2 family proteins in carcinogenesis and the
treatment of cancer. Apoptosis. 14:584–596. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mace PD, Shirley S and Day CL: Assembling
the building blocks: structure and function of inhibitor of
apoptosis proteins. Cell Death Differ. 17:46–53. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
de Almagro MC and Vucic D: The inhibitor
of apoptosis (IAP) proteins are critical regulators of signaling
pathways and targets for anti-cancer therapy. Exp Oncol.
34:200–211. 2012.PubMed/NCBI
|
|
31
|
Fulda S and Vucic D: Targeting IAP
proteins for therapeutic intervention in cancer. Nat Rev Drug
Discov. 11:109–124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Varfolomeev E and Vucic D: Inhibitor of
apoptosis proteins: Fascinating biology leads to attractive tumor
therapeutic targets. Future Oncol. 7:633–648. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang S: Design of small-molecule Smac
mimetics as IAP antagonists. Curr Top Microbiol Immunol.
348:89–113. 2011.PubMed/NCBI
|
|
34
|
Eckelman BP, Salvesen GS and Scott FL:
Human inhibitor of apoptosis proteins: Why XIAP is the black sheep
of the family. EMBO Rep. 7:988–994. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bertrand MJ, Milutinovic S, Dickson KM, Ho
WC, Boudreault A, Durkin J, Gillard JW, Jaquith JB, Morris SJ and
Barker PA: cIAP1 and cIAP2 facilitate cancer cell survival by
functioning as E3 ligases that promote RIP1 ubiquitination. Mol
Cell. 30:689–700. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen SM, Li YY, Tu CH, Salazar N, Tseng
YY, Huang SF, Hsieh LL and Lui TN: Blockade of inhibitors of
apoptosis proteins in combination with conventional chemotherapy
leads to synergistic antitumor activity in medulloblastoma and
cancer stem-like cells. PLoS One. 11:e01612992016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mita AC, Mita MM, Nawrocki ST and Giles
FJ: Survivin: Key regulator of mitosis and apoptosis and novel
target for cancer therapeutics. Clin Cancer Res. 14:5000–5005.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zaffaroni N and Daidone MG: Survivin
expression and resistance to anticancer treatments: Perspectives
for new therapeutic interventions. Drug Resist Updat. 5:65–72.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lin YF, Lai TC, Chang CK, Chen CL, Huang
MS, Yang CJ, Liu HG, Dong JJ, Chou YA, Teng KH, et al: Targeting
the XIAP/caspase-7 complex selectively kills caspase-3-deficient
malignancies. J Clin Invest. 123:3861–3875. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sung B, Cho SG, Liu M and Aggarwal BB:
Butein, a tetrahydroxychalcone, suppresses cancer-induced
osteoclastogenesis through inhibition of receptor activator of
nuclear factor-kappaB ligand signaling. Int J Cancer.
129:2062–2072. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Moon DO, Choi YH, Moon SK, Kim WJ and Kim
GY: Butein suppresses the expression of nuclear factor-kappa
B-mediated matrix metalloproteinase-9 and vascular endothelial
growth factor in prostate cancer cells. Toxicol In Vitro.
24:1927–1934. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Soriano-Hernandez AD, Madrigal-Perez D,
Galvan-Salazar HR, Martinez-Fierro ML, Valdez-Velazquez LL,
Espinoza-Gómez F, Vazquez-Vuelvas OF, Olmedo-Buenrostro BA,
Guzman-Esquivel J, Rodriguez-Sanchez IP, et al: Anti-inflammatory
drugs and uterine cervical cancer cells: Antineoplastic effect of
meclofenamic acid. Oncol Lett. 10:2574–2578. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Moon DO, Kim MO, Choi YH and Kim GY:
Butein sensitizes human hepatoma cells to TRAIL-induced apoptosis
via extracellular signal-regulated kinase/Sp1-dependent DR5
upregulation and NF-kappaB inactivation. Mol Cancer Ther.
9:1583–1595. 2010. View Article : Google Scholar : PubMed/NCBI
|