Introduction
Cell migration serves crucial roles in numerous
biological and pathological processes, including embryonic
morphogenesis, tissue repair and cancer metastasis (1–3). During
the complicated migratory processes, one of the most important
steps is the formation of lamellipodia, which are broad, flat
protrusions at the leading edge of cells that have the ability to
sense the surrounding environment, and drive and guide cell
locomotion (4,5). Lamellipodia formation requires the
assembly of the actin cytoskeleton and the motility of membranes
(6). Lipid rafts, liquid-ordered
plasma membrane microdomains, are in principle well suited to serve
major roles in regulating membrane motility. Lipid rafts accumulate
at the leading edges in migrating fibroblast-like cells and
regulate cell motility by selectively excluding or including
proteins (7–9). Furthermore, when the integrity of lipid
rafts is disrupted, the migration of multiple cancer cells is
inhibited (10–12). However, whether lipid rafts influence
lamellipodia formation of cancer cells has not been described.
Beyond actin polymerization, the generally known
basic mechanism of lamellipodia formation, the adhesion of membrane
protrusions to the extracellular matrix (ECM) is also necessary for
the formation of lamellipodia. It has been reported that
lamellipodia that do not establish stable adhesions become
retracted towards the cell body (13). Integrin, a major cell surface
receptor, mediates the adhesion between cells and the ECM, and
serves important roles in cell migration. Integrin transmits
signals into cells and generates positive feedback to control
lamellipodia formation (14–16). However, the core function of integrin
is to nucleate the formation of focal adhesions at the
lamellipodia, which physically link the actin cytoskeleton to the
ECM and generate the traction to pull the cell body forwards
(17). In addition, integrin
recycling is believed to be linked to the migration of cells.
Generally, integrins are internalized at the rear of the migrating
cell and are recycled to the leading edge, thus resulting in high
ras-related C3 botulinum toxin substrate activity and
lamellipodia-like protrusions (18).
Recently, integrins have been shown to be localized in lipid rafts,
and the disruption of lipid rafts inhibits the internalization and
function of integrins (19–21). In our previous study, it was found
that lipid rafts regulate the internalization of β3 integrin
through sarcoma protein kinase-rhodopsin (Rho)-Rho-associated
protein kinase (ROCK)-mediated actin cytoskeleton dynamics in
migrating human melanoma A375 cells (22). In A375 cell spreading, lipid rafts
control β1 integrin clustering via the recruitment and modification
of certain adaptor proteins (23).
However, the role and the association of lipid rafts and integrins
in lamellipodia formation in human melanoma A375 cells remain
unclear.
Based on our previous results (22,23), the
present study aimed to investigate the association between lipid
rafts and the lamellipodia formation of A375 cells and determine
whether lipid rafts can control the lamellipodia formation of A375
cells by regulating β1 and β3 integrin distribution in the cell
membrane.
Materials and methods
Cell culture
Human melanoma A375 cells were purchased from the
Cell Bank of the Type Culture Collection of the Chinese Academy of
Science (Shanghai, China) and cultured in Dulbecco's modified
Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS) at
37°C in 5% CO2.
Antibodies and regents
Antibodies to β1 integrin (clone TDM29; 1:200; cat.
no. CBL481) and β3 integrin (clone LM609, 1:200; cat. no. MAB1976)
were purchased from EMD Millipore (Billerica, MA, USA).
Tetramethylrhodamine or fluorescein isothiocyanate-conjugated goat
anti-mouse IgG antibody (1:250; cat. nos. T5393 and F9006,
respectively), methyl-β cyclodextrin (MβCD, cat. no. C4555),
cytochalasin D (CD, cat. no. C2618) and cholesterol (cat. no.
C8667) were purchased from Sigma-Aldrich; Merck KGaA (Darmstadt,
Germany). Rhodamine-conjugated phalloidin (cat. no. R415) and Alexa
Fluor® 488-conjugated cholera toxin subunit B (cat. no.
C22841) were purchased from Molecular Probes; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). DMEM was purchased from Thermo
Fisher Scientific, Inc. and FBS was obtained from Gibco; Thermo
Fisher Scientific, Inc.
Analysis of lamellipodia formation at
wound edges
A375 cells were grown to confluence on glass
coverslips and scratch wounded with the narrow end of a 10-µl
pipette tip. The cells were then washed twice with
phosphate-buffered saline and incubated with fresh 2% FBS/DMEM with
or without 5 mM MβCD at 37°C. After 3 h, the MβCD was removed, and
fresh medium containing 1 mM cholesterol was added for 6 h to allow
the integrity of the lipid rafts to recover. The formation of
lamellipodia at the wound edges was investigated with phase
contrast microscopy (Nikon Corporation, Tokyo, Japan), and the
percentage of cells displaying lamellipodia was calculated.
Immunofluorescence
A375 cells were grown to confluence on glass
coverslips and scratch wounded. The cells were then treated with 5
mM MβCD for 0, 1, 2 and 3 h, or treated with 0.05 µg/ml CD for 1 h.
Subsequent to being washed with phosphate-buffered saline, the
cells were fixed with 10% formaldehyde for 10 min at 22°C and
permeabilized with 0.1% Triton X-100 for 3 min. The cells were then
blocked in 3% bovine serum albumin for 1 h at 37°C, incubated with
the aforementioned primary antibodies for 1 h at 22°C and
subsequently incubated with fluorochrome-conjugated secondary
antibody for 45 min at 22°C. F-actin was labeled with
rhodamine-conjugated phalloidin, and the lipid raft marker,
ganglioside GM1, was labeled with Alexa Fluor 488-conjugated
cholera toxin subunit B. The coverslips were mounted and observed
under a confocal microscope (Olympus Corporation, Tokyo,
Japan).
Statistical analysis
Data shown represent the mean ± standard deviation
from three independent experiments. Statistical comparisons were
performed using one-way analysis of variance followed by Tukey's
test. P<0.05 was considered to indicate a statistically
significant difference. Statistical analysis was performed using
SPSS version 19.0 software (IBM Corp, Armonk, NY, USA).
Results
Lipid rafts regulate lamellipodia
formation in A375 cells
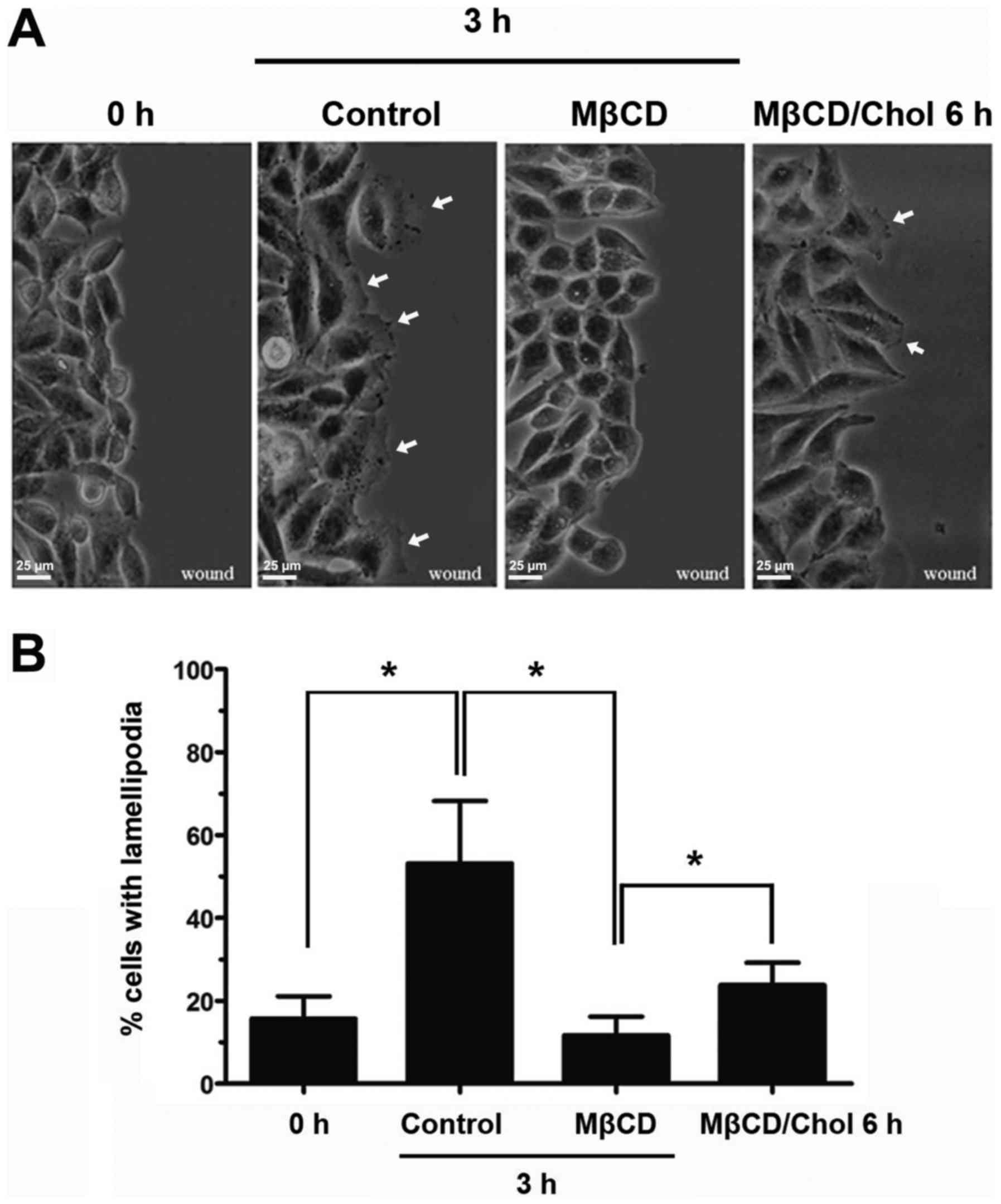
To investigate the role of lipid rafts in
lamellipodia formation in A375 cells, a cell migration model was
established by scratch wound healing, and 5 mM MβCD was used to
deplete cholesterol and disrupt the integrity of the lipid rafts
(22). At 3 h post-wounding, 50% of
cells without MβCD treatment extended broad lamellipodia towards
the scratch area (Fig. 1A and B).
However, 5 mM MβCD-treated A375 cells appeared to have lost the
ability to form lamellipodia protrusions (Fig. 1A). The percentage of cells extending
lamellipodia following MβCD treatment was decreased to <20% of
cells (Fig. 1B). When cholesterol was
added to the MβCD-treated A375 cells to rescue lipid raft
integrity, the strongly suppressed lamellipodia reformed (Fig. 1A and B). These results indicated that
intact lipid rafts are indispensable in lamellipodia formation in
A375 cells.
Lipid rafts are asymmetrically
distributed to the leading edge in migrating A375 cells
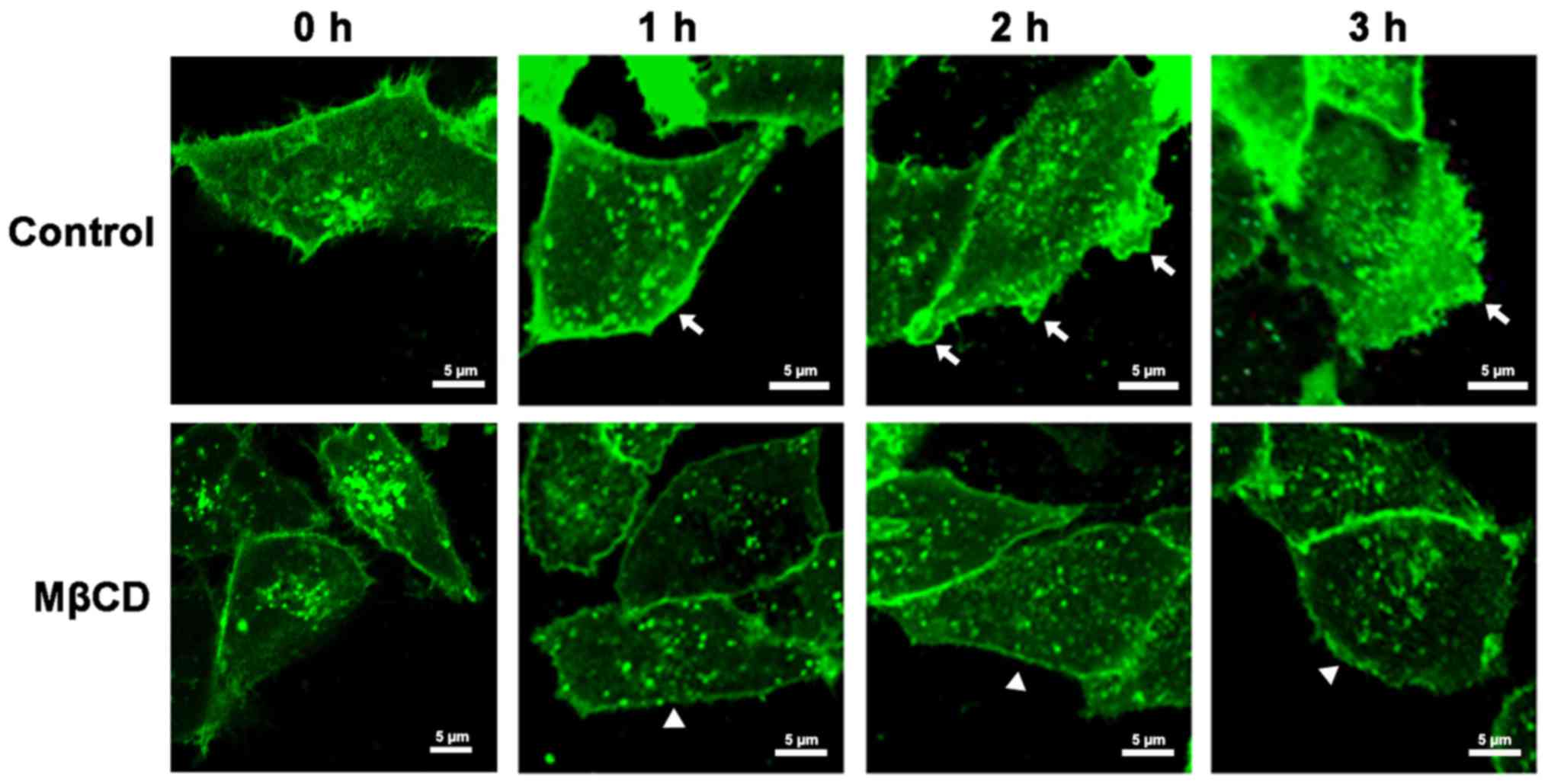
To further investigate how lipid rafts influence the
lamellipodia formation of A375 cells, the distribution of lipid
rafts in migrating cells was first detected. A375 cells were
wounded and subjected to immunostaining for ganglioside GM1, which
is raft-enriched and is regarded as a marker to identify lipid
rafts. The results showed that lipid rafts were asymmetrically
distributed in the cell membrane during the formation of the
lamellipodia. When the cells were initially wounded (0 h; Fig. 2), GM1 was homogeneously distributed in
the cytoplasm and at the cell periphery of A375 cells. At 1 h
post-wounding, GM1 aggregated at the leading edge of A375 cells,
and this was followed by the formation of small, scattered
lamellipodia (2 h) (Fig. 2). At 3 h
post-wounding, broad, flat lamellipodia formed towards the scratch
area, and this was accompanied by increased GM1 at the leading edge
of the lamellipodia (Fig. 2).
However, when MβCD was added following wounding, GM1 remained
evenly distributed and lamellipodia formation was inhibited (1–3 h;
Fig. 2). These results indicated that
the aggregation of lipid rafts at the leading edge in cell
membranes contributes to lamellipodia formation.
Lipid rafts recruit β1 and β3
integrins to lamellipodia
Lipid rafts function as scaffold-like platforms for
protein recruitment and signal transfer. Therefore, we speculated
that lipid rafts may spatially concentrate the proteins closely
associated with lamellipodia formation to the leading edges of
cells. Integrins are believed to be crucial adhesion molecules in
cell membranes that mediate lamellipodia formation (15). Our previous data have shown that β1
and β3 integrins are highly expressed in melanoma A375 cells and
are closely associated with cell migration (23). Thus, the present study sought to
investigate whether lipid rafts regulate lamellipodia formation by
affecting the distribution of β1 and β3 integrin.
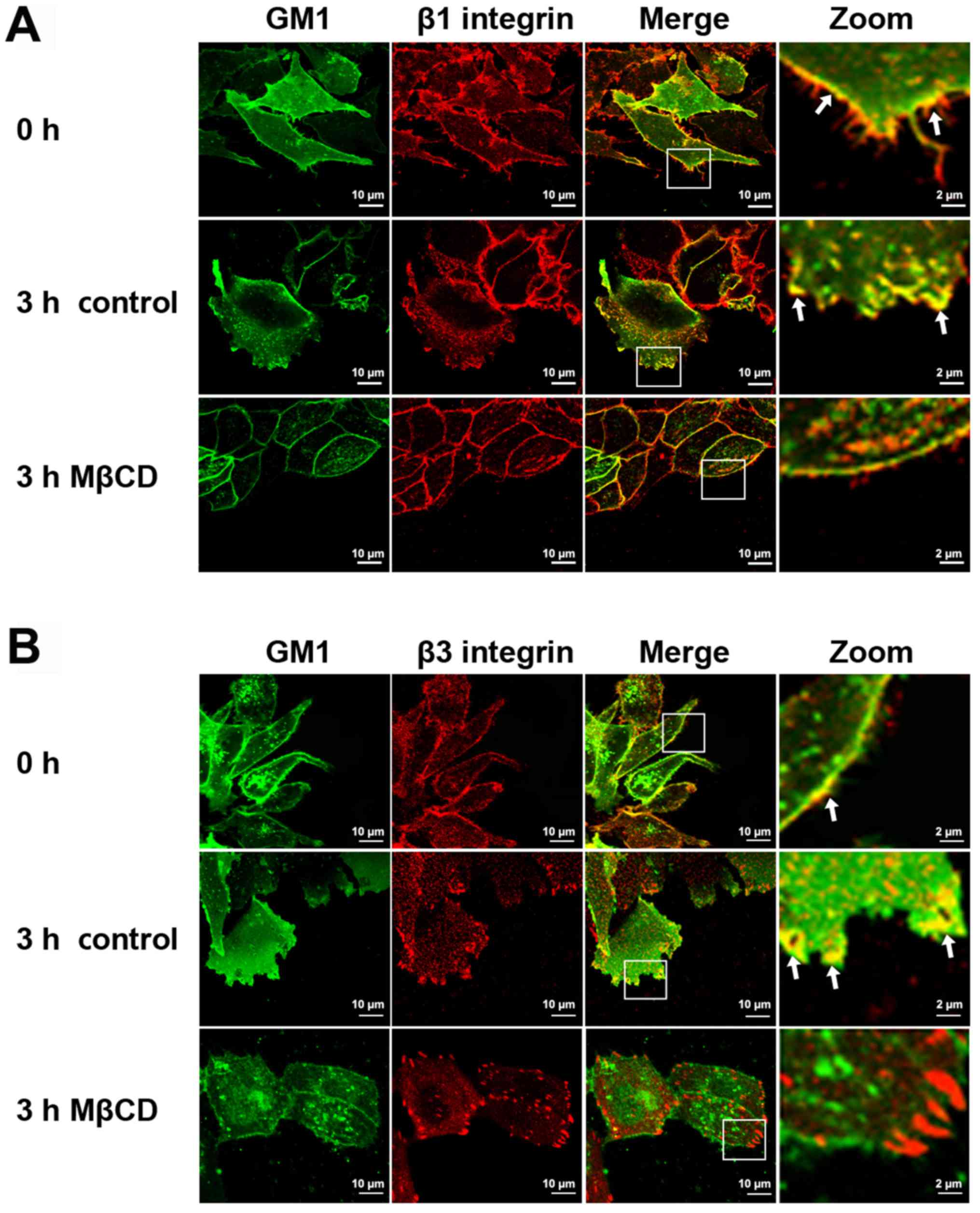
Immunofluorescence staining revealed the different distributions of
β1 and β3 integrins in the lamellipodia of A375 cells prior to and
following MβCD treatment. In control migrating A375 cells, β1
integrin, which showed a dispersed distribution, colocalized with
GM1 mainly at the front edges of the lamellipodia (Fig. 3A). However, β3 integrin, which had a
spot-like distribution in the lamellipodia, colocalized with GM1
within the cell membrane surrounding the front edge of the
lamellipodia (Fig. 3B). Following
MβCD treatment for 3 h, the morphology of the cells became spindle
shaped. Meanwhile, the asymmetrical distribution of β1 integrin
disappeared and became an even distribution in the periphery of the
cells (Fig. 3A). By contrast, β3
integrin was visible as larger foci following MβCD treatment and
was distributed at the cell periphery (Fig. 3B). These results indicated that β1 and
β3 integrins, two important proteins that may serve different roles
in lamellipodia formation, are recruited to lamellipodia by lipid
rafts.
Actin cytoskeleton is responsible for
lipid raft-mediated β1 and β3 integrin distribution in
lamellipodia
Given the aforementioned results, the mechanism of
lipid raft-mediated β1 and β3 integrin recruitment in lamellipodia
was further investigated. The actin cytoskeleton has been reported
to associate with lipid rafts, and to regulate their structure and
organization (24). In addition,
integrins connect the ECM with the actin cytoskeleton inside the
cell. Thus, we speculated that the actin cytoskeleton may be
responsible for lipid raft-mediated β1 and β3 integrin distribution
in lamellipodia. To verify this hypothesis, the colocalization
between β1 integrin and actin, and β3 integrin and actin was first
detected. β1 integrin colocalized with the submembranous cortical
actin cytoskeleton at the forefront of the lamellipodia. However,
β3 integrin and actin colocalized at the ends of the stress fibers,
which were inside the leading edge (Fig.
4A). These results indicated that the actin cytoskeleton was
associated with the β1 and β3 integrins. Next, the cells were
treated with CD to disrupt the actin cytoskeletal arrangement, and
it was found that the colocalization between GM1 and β1 integrin,
and GM1 and β3 integrin, in the lamellipodia disappeared following
CD treatment (Fig. 4B and C).
Together, the results suggest that the actin cytoskeleton may be
responsible for lipid raft-mediated β1 and β3 integrin distribution
in lamellipodia.
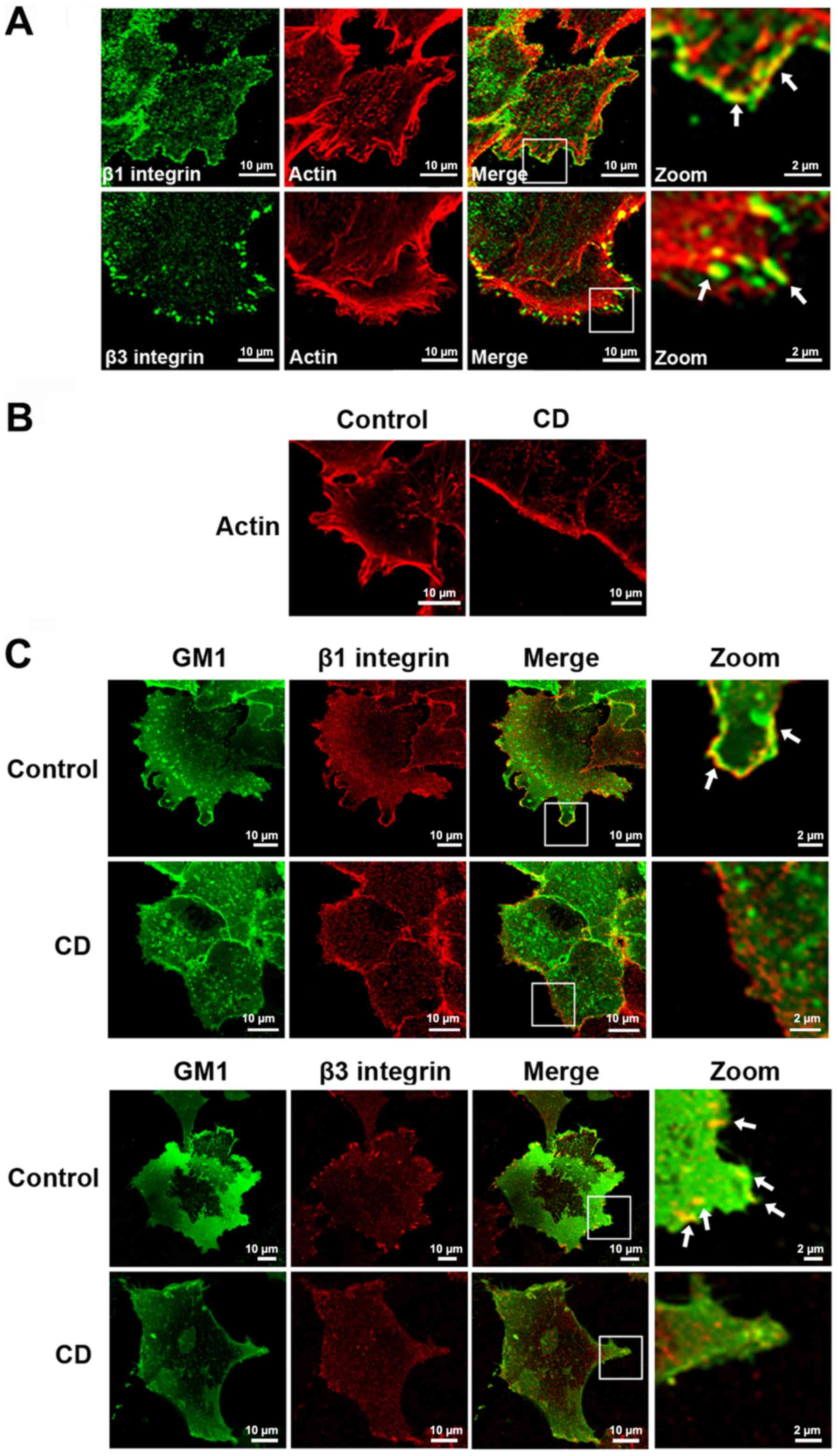 | Figure 4.Lipid raft-mediated β1 and β3 integrin
distribution in lamellipodia requires an intact actin cytoskeleton.
(A) Migrating A375 cells were stained for F-actin (red), β1
integrin and β3 integrin (green), respectively. (B) A375 cells
treated with 0.05 µg/ml CD for 1 h, or left untreated, were stained
for F-actin. (C) A375 cells treated with 0.05 µg/ml CD for 1 h, or
left untreated, were stained for GM1 (green), β1 integrin and β3
integrin (red) antibody, respectively. The images were obtained
with confocal microscopy (×60 magnification). The right panel shows
magnified views of the boxed area in the merged images. The arrows
in the images indicate the colocalization. Scale bar, 10 µm. CD,
cytochalasin D; GM1, monosialotetrahexosyl ganglioside. |
Discussion
Lipid rafts, detergent-resistant membrane domains
enriched in cholesterol and sphingolipids, have been implicated in
cancer progression, including the migration and invasion of cancer
cells (25,26). However, the role of lipid rafts in
lamellipodia formation remains obscure. In the present study, by
observing the morphological changes in lipid raft-disrupted A375
cells, it was determined that intact lipid rafts are indispensable
in lamellipodia formation in melanoma cells.
One of the most critical functions of lipid rafts is
to act as platforms for localizing signaling proteins and eliciting
signal transduction (9). This
characteristic has been extensively reported, particularly in T
cells and other leukocytes (27–30).
Several cell surface receptor proteins have been reported to be
localized to lipid rafts and to execute their functions through
association with lipid rafts (28,31,32). In
resting cells, lipid rafts are evenly distributed over the entire
cell surface. However, when the cells are stimulated, lipid rafts
move laterally and coalesce into larger aggregated patches, thus
resulting in the concentration and redistribution of
raft-associated proteins, and efficient and sustained signal
transduction (33). In the present
study, using immunofluorescence assays, it was found that as A375
cells migrated to the scratches, lipid rafts aggregated at the
leading edge of the cells with an asymmetrical distribution, which
was critical for lamellipodia formation.
Given these results and the function of lipid rafts
as platforms, we speculated that proteins associated with lipid
rafts and lamellipodia formation may be recruited to the leading
edges of A375 cells along with the asymmetrical distribution of
lipid rafts. β1 and β3 integrins have been found to be membrane
raft-associated proteins (23). As
adhesion molecules, β1 and β3 integrins regulate the migration and
lamellipodia formation of cancer cells. Thus, the present study
examined whether the asymmetrical distribution of lipid rafts could
aggregate β1 and β3 integrin into the lamellipodia of A375 cells.
The results showed that in migrating A375 cells, β1 and β3
integrins colocalized with GM1 in the lamellipodia. However, MβCD
treatment changed the distributions of β1 and β3 integrins in the
cells and inhibited lamellipodia formation, thus suggesting that
lipid rafts are involved in the recruitment of β1 and β3 integrin
in the lamellipodia. In the process of these experiments, a notable
phenomenon of β1 and β3 integrin being differently distributed in
A375 cells became apparent. Subsequent research on the
colocalization of β1 and β3 integrins with the actin cytoskeleton
also confirmed that β1 integrin was distributed mainly along the
leading edge of the lamellipodia, thereby determining the shape of
the cell, whereas β3 integrin was distributed inside the leading
edge of the lamellipodia and colocalized with actin at the ends of
stress fibers. Despite belonging to the same family, the different
distribution characteristics of β1 and β3 integrin suggested that
they have different functions. In fact, distinct functions of β1
and β3 integrin in different cell types or even at different stages
of one cell type have been reported. For example, β1 integrin is
widely distributed on various cell types and is involved in the
physiological processes of cell proliferation, survival and
differentiation (34). However, β3
integrin is mainly expressed on the surface of cancer cells and
platelets, and it participates in the migration and invasion of
tumor cells, and the coagulation of platelets (35). In cell migration, β1 integrin promotes
random migration, whereas β3 integrin promotes persistent migration
in the same epithelial cell background (36). In the assembly of focal adhesion, β1
integrin has been shown to be the core component of focal adhesion
in epithelial cells isolated from human breast tumors and in
spreading human melanoma A375 cells (23,37),
whereas in migrating A375 cells, focal adhesions largely consist of
β3 integrin (22). However, to the
best of our knowledge, this is the first study to describe the
distinct distributions of the two integrins in the lamellipodia
formation of A375 cells.
Next, the present study detected how lipid rafts
recruit β1 and β3 integrin to the lamellipodia. Lipid rafts and the
actin cytoskeleton have been reported to be closely associated. In
our previous study, it was found that lipid rafts regulate the
dynamics of the actin cytoskeleton. When the integrity of lipid
rafts is disrupted, A375 cells form strong stress fibers, thus
suggesting inhibition of the depolymerization of the actin
cytoskeleton (22). By contrast,
other studies have indicated that the organization, structure and
function of lipid rafts requires an intact actin cytoskeleton
(24). Thus, the present study
investigated the role of the actin cytoskeleton in lipid
raft-mediated β1 and β3 integrin distribution in the lamellipodia.
Immunofluorescence assays showed that the actin cytoskeleton
colocalized with β1 and β3 integrins. When the arrangement of the
actin cytoskeleton was disrupted by CD, the distribution of β1 and
β3 integrin in lamellipodia, and their colocalization with GM1
disappeared. These results demonstrated that lipid rafts recruit β1
and β3 integrin to lamellipodia via the actin cytoskeleton.
In summary, the present data indicated that lipid
rafts recruit β1 and β3 integrin to the leading edge in melanoma
A375 cells, thereby facilitating lamellipodia formation, in a
manner dependent on the intact actin cytoskeleton. These findings
provide novel insight into the association between lipid rafts and
lamellipodia formation.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (no. 81402416) and the Henan
Scientific and Technological Research Projects (no.
182102311136).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XZ created the study concept and designed the
experiments. JB performed the experiments and wrote the manuscript.
RW analyzed the data and edited the manuscript. All the authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ECM
|
extracellular matrix
|
|
GM1
|
monosialotetrahex-osylganglioside
|
|
MβCD
|
methyl-β, cyclodextrin
|
|
CD
|
cytochalasin D
|
References
|
1
|
Reig G, Cerda M, Sepúlveda N, Flores D,
Castañeda V, Tada M, Härtel S and Concha ML: Extra-embryonic tissue
spreading directs early embryo morphogenesis in killifish. Nat
Commun. 8:154312017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang W, Li P, Li W, Jiang J, Cui Y, Li S
and Wang Z: Osteopontin activates mesenchymal stem cells to repair
skin wound. PLoS One. 12:e01853462017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hammer A and Diakonova M: Tyrosyl
phosphorylated serine-threonine kinase PAK1 is a novel regulator of
prolactin-dependent breast cancer cell motility and invasion. Adv
Exp Med Biol. 846:97–137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krause M and Gautreau A: Steering cell
migration: Lamellipodium dynamics and the regulation of directional
persistence. Nat Rev Mol Cell Biol. 15:577–590. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Small JV, Stradal T, Vignal E and Rottner
K: The lamellipodium: Where motility begins. Trends Cell Biol.
12:112–120. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bisi S, Disanza A, Malinverno C, Frittoli
E, Palamidessi A and Scita G: Membrane and actin dynamics interplay
at lamellipodia leading edge. Curr Opin Cell Biol. 25:565–573.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gómez-Móuton C, Abad JL, Mira E, Lacalle
RA, Gallardo E, Jiménez-Baranda S, Illa I, Bernad A, Mañes S and
Martínez-A C: Segregation of leading-edge and uropod components
into specific lipid rafts during T cell polarization. Proc Natl
Acad Sci USA. 98:9642–9647. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Golub T, Wacha S and Caroni P: Spatial and
temporal control of signaling through lipid rafts. Curr Opin
Neurobiol. 14:542–550. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Simons K and Toomre D: Lipid rafts and
signal transduction. Nat Rev Mol Cell Biol. 1:31–39. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bi J, Wang R, Zhang Y, Han X, Ampah KK,
Liu W and Zeng X: Identification of nucleolin as a
lipid-raft-dependent β1-integrin-interacting protein in A375 cell
migration. Mol Cells. 36:507–517. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jeon JH, Kim SK, Kim HJ, Chang J, Ahn CM
and Chang YS: Lipid raft modulation inhibits NSCLC cell migration
through delocalization of the focal adhesion complex. Lung Cancer.
69:165–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Raghu H, Sodadasu PK, Malla RR, Gondi CS,
Estes N and Rao JS: Localization of uPAR and MMP-9 in lipid rafts
is critical for migration, invasion and angiogenesis in human
breast cancer cells. BMC Cancer. 10:6472010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Borm B, Requardt RP, Herzog V and Kirfel
G: Membrane ruffles in cell migration: Indicators of inefficient
lamellipodia adhesion and compartments of actin filament
reorganization. Exp Cell Res. 302:83–95. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu S, Calderwood DA and Ginsberg MH:
Integrin cytoplasmic domain-binding proteins. J Cell Sci.
113:3563–3571. 2000.PubMed/NCBI
|
|
15
|
Saravanan C, Liu FT, Gipson IK and
Panjwani N: Galectin-3 promotes lamellipodia formation in
epithelial cells by interacting with complex N-glycans on
alpha3beta1 integrin. J Cell Sci. 122:3684–3693. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hamill KJ, Hopkinson SB, Jonkman MF and
Jones JC: Type XVII collagen regulates lamellipod stability, cell
motility, and signaling to Rac1 by targeting bullous pemphigoid
antigen 1e to alpha6beta4 integrin. J Biol Chem. 286:26768–26780.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Maziveyi M and Alahari SK: Cell matrix
adhesions in cancer: The proteins that form the glue. Oncotarget.
8:48471–48487. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Paul NR, Jacquemet G and Caswell PT:
Endocytic trafficking of integrins in cell migration. Curr Biol.
25:R1092–R1105. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vassilieva EV, Gerner-Smidt K, Ivanov AI
and Nusrat A: Lipid rafts mediate internalization of beta1-integrin
in migrating intestinal epithelial cells. Am J Physiol Gastrointest
Liver Physiol. 295:G965–G976. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee JL, Wang MJ, Sudhir PR and Chen JY:
CD44 engagement promotes matrix-derived survival through the
CD44-SRC-integrin axis in lipid rafts. Mol Cell Biol. 28:5710–5723.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Runz S, Mierke CT, Joumaa S, Behrens J,
Fabry B and Altevogt P: CD24 induces localization of beta1 integrin
to lipid raft domains. Biochem Biophys Res Commun. 365:35–41. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang R, Bi J, Ampah KK, Ba X, Liu W and
Zeng X: Lipid rafts control human melanoma cell migration by
regulating focal adhesion disassembly. Biochim Biophys Acta.
1833:3195–3205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang R, Bi J, Ampah KK, Zhang C, Li Z,
Jiao Y, Wang X, Ba X and Zeng X: Lipid raft regulates the initial
spreading of melanoma A375 cells by modulating β1 integrin
clustering. Int J Biochem Cell Biol. 45:1679–1689. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chichili GR and Rodgers W:
Cytoskeleton-membrane interactions in membrane raft structure. Cell
Mol Life Sci. 66:2319–2328. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Costantini F and Barbieri G: The HLA-DR
mediated signalling increases the migration and invasion of
melanoma cells, the expression and lipid raft recruitment of
adhesion receptors, PD-L1 and signal transduction proteins. Cell
Signal. 36:189–203. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang YF, Jan YH, Liu YP, Yang CJ, Su CY,
Chang YC, Lai TC, Chiou J, Tsai HY, Lu J, et al: Squalene synthase
induces tumor necrosis factor receptor 1 enrichment in lipid rafts
to promote lung cancer metastasis. Am J Respir Crit Care Med.
190:675–687. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lingwood D and Simons K: Lipid rafts as a
membrane-organizing principle. Science. 327:46–50. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Setiadi H and McEver RP: Clustering
endothelial E-selectin in clathrin-coated pits and lipid rafts
enhances leukocyte adhesion under flow. Blood. 111:1989–1998. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dykstra M, Cherukuri A, Sohn HW, Tzeng SJ
and Pierce SK: Location is everything: Lipid rafts and immune cell
signaling. Annu Rev Immunol. 21:457–481. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kiely JM, Hu Y, García-Cardeña G and
Gimbrone MA Jr: Lipid raft localization of cell surface E-selectin
is required for ligation-induced activation of phospholipase C
gamma. J Immunol. 171:3216–3224. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Leitinger B and Hogg N: The involvement of
lipid rafts in the regulation of integrin function. J Cell Sci.
115:963–972. 2002.PubMed/NCBI
|
|
32
|
Rossy J, Schlicht D, Engelhardt B and
Niggli V: Flotillins interact with PSGL-1 in neutrophils and, upon
stimulation, rapidly organize into membrane domains subsequently
accumulating in the uropod. PLoS One. 4:e54032009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Simons K and Sampaio JL: Membrane
organization and lipid rafts. Cold Spring Harb Perspect Biol.
3:a0046972011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Riopel MM, Li J, Liu S, Leask A and Wang
R: β1 integrin-extracellular matrix interactions are essential for
maintaining exocrine pancreas architecture and function. Lab
Invest. 93:31–40. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kuphal S, Bauer R and Bosserhoff AK:
Integrin signaling in malignant melanoma. Cancer Metastasis Rev.
24:195–222. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Danen EH, van Rheenen J, Franken W,
Huveneers S, Sonneveld P, Jalink K and Sonnenberg A: Integrins
control motile strategy through a Rho-cofilin pathway. J Cell Biol.
169:515–526. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Goel HL, Pursell B, Standley C, Fogarty K
and Mercurio AM: Neuropilin-2 regulates α6β1 integrin in the
formation of focal adhesions and signaling. J Cell Sci.
125:497–506. 2012. View Article : Google Scholar : PubMed/NCBI
|