Introduction
Uterine cervical cancer (UCC) is a tumor of
epithelial cells located in the cervix that affects women around
the world (1). An estimate of the
overall disease incidence for the year 2012 (2) revealed the notification of 528 thousand
new cases, with a record of 266 thousand deaths worldwide, making
this malignant neoplasm the fourth most common cancer among women,
representing the fourth cause of cancer death among women worldwide
(3). UCC is considered the second
most common cancer and the third leading cause of death by cancer
among women in less developed countries, with higher incidence and
mortality rates in sub-Saharan Africa, Melanesia, Latin America,
and the Caribbean (2).
The natural history of the UCC clearly shows the
close relationship between the disease and persistent infection
with genotypes of high-risk human papillomavirus (HR-HPV), since
the viral genome sequences are detected in tumor cells of virtually
all cases of the disease (4). Despite
its high prevalence due to its infectious nature, UCC stands out
among the malignant neoplasms with the greatest potential of
prevention and cure (5). The wide
variation in incidence and mortality rates of UCC in different
geographic regions reflects the economic, social, and cultural
differences (6), as well as the
availability of screening tests, which allow early detection and
removal of precancerous lesions (7,8).
In most cases the infection is transient and
asymptomatic without the presence of injury, or has only low-grade
lesions that regress to spontaneous healing in a period of up to
two years (9). However, in a
proportion of women infected with oncogenic genotypes of the virus,
the symptoms disappear but the virus is not eliminated; rather, it
remains in the form of persistent infection (10). A number of women with persistent
infection develop cervical intraepithelial neoplasia (CIN), which
can progress to UCC. The progression to cancer occurs when the
virus integrates the chromosomes of the host cell and depends on
other factors such as epigenetic events triggered by physical,
chemical, and environmental factors, as well as host
characteristics including nutritional status, genetic background,
and immune status (11,12).
The absence of a cure for HPV infection seems to be
related to escape mechanisms used by HPV to evade the host immune
response. HR-HPV avoids immunological recognition by inactivating
the cells of the innate immune response, downregulating the type I
interferon production and inducing peripheral tolerance of
cytotoxic T lymphocytes (CTL) (13,14). One
possible explanation for this is the fact that the infection is
restricted to the epithelium, with no occurrence of viremia with
the spread of the virus, nor lyses, degeneration or necrosis of
cells, or damaging signs to alert the immune system (15). Thus, there is no activation of an
inflammatory response with the necessary cytokine production for
recognition of the virus as a pathogenic agent. This hinders the
proper activation of the local immune response, which may favor an
induction of immune tolerance to the virus and which therefore
becomes undetectable after long periods of time, promoting the
immune evasion of the infected cells (16).
HPV infection induces weak immunity due to the
action of viral proteins that activate multiple mechanisms to
prevent initiating a robust immune response. The most notable among
these mechanisms are the depletion of Langerhans cells (LCs)
mediated by viral protein E6; the downregulating of MHC molecules
of class I by E5, avoiding the attack of CTL; and the blocking of
the signaling pathway of type I interferon by E7 (17). Furthermore, the downregulating of IFN
expression by viral proteins E6 and E7 results in an absence of
co-stimulatory signals by inflammatory cytokines, including IFN for
recognizing antigens that can induce immunological tolerance rather
than an effective immune response (18).
Thus, the curing of HPV-induced lesions is dependent
on a cell-mediated immune response, requiring interactions between
CD4+ and CD8+ T lymphocytes. LCs are probably responsible for
triggering an anti-HPV immune response, but the function of these
cells is interrupted by HPV at different levels (19). The epithelium of the uterine cervix is
a very poor microenvironment in immunological mediators, and
therefore appears to be immunologically hyporesponsive to HPV since
this virus induces a local immune deficiency by the depletion of
CD4+ and LCs, as well as the downregulating of the cytokine
production (20). Additionally, the
vaginal epithelium has a dendritic cell (DC) subset that does not
express Langerin (Lang-DC), exerting the downregulation of CTL in
the cervical mucosa by a mechanism that may involve interleukin
(IL)-17, and IL-10 to a lesser degree (21).
When the antigen presentation occurs in the absence
of cytokines that favor the response of the Th1 type,
differentiation into other types of response can occur, including
Th2 or Th17. This results in a less-efficient T-cell response,
promoting viral persistence and increasing the risk of cervical
lesions of different degrees, as well as the progression to cancer
(22). Furthermore, there may be
unbalanced activation of Th17 and regulatory T cells (Treg), which
can compromise the efficiency of the immune response, reducing
their suppressive action on the tumor, and also possibly favoring
tumor development and progression (23–25).
Persistent HPV infection may affect cell
differentiation of the adaptive immune response including CD4+/CD8+
T cells and Treg cells, resulting in immune tolerance of the host
to the infected cells. Thus, a compromised cellular immune
response, an abnormal imbalance between type 1 T helper (Th1) and
Th2 cells, infiltration of Treg cells, and the downregulation of
activating and maturing DCs may contribute to the progression of
HPV-associated premalignant lesions for UCC (16). It has been reported that high
expression levels of IL-17, which is the main cytokine of the Th17
response, are associated with poor UCC prognosis, but the role of
this kind of response in tumor progression is still ambiguous
(26). This review was performed to
analyze recent advances on the immunological and molecular
mechanisms involved in the immune response of the Th17 type, and
their role in the progression of HPV-associated lesions for
cancer.
The literature search was conducted using the
electronic databases of PubMed (National Institutes of Health,
Bethesda, MD, USA; www.ncbi.nlm.nih.gov/pubmed), Scopus (Elsevier,
Amsterdam, Netherlands; www.scopus.com/scopus/home.url), and Web of Knowledge
(Thomson Reuters, New York, NY, USA; www.webofknowledge.com), using the following keywords:
Th17, Th17 cells, IL-17, HPV Infection, Cervical Cancer, HPV
infection, HPV-associated cervical lesions. The databases retrieved
thousands of articles and those thought to be most relevant were
selected, which included recent studies published in impact
journals conducted by groups with recognized expertise in the areas
of immunity and cancer, and mainly cervical cancer.
Differentiation of T helper cells
After the first contact with its specific antigen
through antigen-presenting cells (APC), the specific clone of the
naïve TCD4+ cell acquires the potential to differentiate into
effector T cells with a variety of different phenotypes of each
other (27,28). These different types of cells, called
T-helper cells, acquire the ability to produce and secrete
cytokines through a differentiation process triggered in response
to multiple signals of the environment, including cytokines
(29). These cells are crucial for
mounting an adaptive immune response that protects the host against
both the infection caused by pathogens and the development of
tumors (30). However, the responses
induced by these cell types may also trigger immunopathological
mechanisms that result in tissue damage, similar to those observed
in the infections caused by intracellular pathogens such as
viruses, autoimmune disorders, and allergies (31).
Only two types of T-helper cells were initially
identified. One was the Th1 cell, described by Mosmann et al
(32), which develops in response to
IL-12, signaling, producing and especially secreting IFN-γ and
regulating cell-mediated protective immunity against intracellular
pathogens (33). The other type
identified was the Th2 cell, described by Murphy et al
(34), which develops in response to
IL-4, signaling, producing and secreting IL-5 and IL-13 and
regulating protective immunity against extracellular pathogens
(33). This dichotomy of Th1/Th2
prevailed in the field of immune regulation until recently, when
new phenotypes of T-helper cells were identified (27,28).
The enormous complexity of the cell-mediated immune
response revealed by experimental studies had already indicated
that this model (based on only two subtypes of Th cross-regulatory
cells) would be insufficient to explain the various aspects of
initiation, regulation, and fine-tuning of several types of immune
responses triggered by the host in response to the environmental
stimuli (28). Later, a new type of
Th cell was discovered, called regulatory T or Treg that expresses
Foxp3, a transcription factor, and represents a negative regulation
mechanism of immune-mediated inflammation to prevent
self-destructive immune responses, including autoimmune and
auto-inflammatory disorders, allergies, and cancer (35,36).
In the last 10 years, three additional Th cell
subtypes were identified and named according to the type of
cytokine secreted by each of them (28). One of them was Th17, a subtype of Th
that produces and secretes high levels of IL-17 (33), in addition to other inflammatory
cytokines such as IL-21 and IL-22, and are involved in tumor
progression by promoting angiogenesis and immunosuppressive
activities. However, Th17 cells may also act by mediating antitumor
immune responses by promoting recruitment of immune cells to the
tumor site, activating effector CD8+ T cells against the tumors, or
even reverting to the Th1 phenotype by producing IFN-γ which
promotes further activation of CD8+ T cells. Thus, these cells have
an ambiguous function in relation to the tumors (37). The others subtypes are Th9 cells,
which produces and secretes IL-9 (38), and Th22, which produces and secretes
IL-22 (39). This shows that adaptive
cell-mediated immune response involves a complex network of
interactions between cells with different phenotypes through a
suite of mediators, mainly cytokines.
The differentiation of naïve CD4+ T helper cells in
the Th17 cell is stimulated by the combined action of TGF-β and of
pro-inflammatory cytokines such as IL-1β, IL-6, IL-21, and IL-23,
which play a central role in generation of these cells (40). The TGF-β signaling appears to play a
critical role in the differentiation of Th17, since TGF-β
inhibition substantially reduces the generation of these cells. It
has been discussed whether TGF-β is in fact necessary for
generating Th17, as it has been shown that murine T cells can be
differentiated in Th17 using IL-1β, IL-6, and IL-23 in the absence
of exogenous TGF-β. However, treatment with anti-TGF-β antibody
inhibited this differentiation, suggesting the involvement of
endogenous TGF-β in the differentiation process (41).
What differentiates the lineages of TCD4 cell from
each other is the signature transcription factor that each them
express. Thus, Tregs are marked by the expression of FOXP3 induced
during its maturation in the thymus, or in the periphery induced by
TGF-β and retinoic acid. On the other hand, the signature
transcription factor for Th17 cells, retinoid-related orphan
receptor γ t(RORγt), is also induced by TGF-β, thus linking the
differentiation of Treg and Th17 lineages. Therefore, in the
absence of a second signal emitted by a proinflammatory cytokine,
FOXP3 can inhibit RORγt function, and leads to differentiation of
TCD4 in Treg. However, when that same cell receives the second
signal from a proinflammatory cytokine, such as IL-6, the FOXP3
function is inhibited and the cell differentiates in Th17.
Therefore, the balance between the FOXP3 and RORγt function is that
it determines the fate of CD4+ T cells and the type of immune
response that will be generated (42).
The role of GAT3 in Th17 cell differentiation was
analyzed in an experimental murine model of arthritis induced with
methylated bovine serum albumin, in both wild type and in CD2
T-cell-specific GATA-3 (CD2-GATA-3)-transgenic mice. It was found
that wild-type mice developed severe joint inflammation with
massive inflammatory cell infiltration and bone erosion, while the
CD2-GATA-3 transgenic mice only presented mild joint inflammation,
accompanied by systemic and local reductions in the numbers of
IL-17+ IFN-γ- and IL-17+ IFN-γ +, but not of IL-17-IFN-γ cells and
CD4+ T cells, by inducing cytokine expression with Th2 profile. In
addition, overexpression of GATA-3 resulted in reduced expression
of the gene encoding RORγt, the signature transcription factor of
Th17 cells. These results indicate that GATA-3 protects against
joint inflammation and bone erosion by inhibiting the
differentiation of Th17, but not Th1 cells in experimental
arthritis in mice (43).
Ovarian tumor cells, tumor-derived fibroblasts and
antigen presenting cells (APCs) have also been shown to secrete
cytokines such as IL-1β, IL-6, TNF-α and TGF-β, which create a
favorable environment for differentiation and expansion of IL-17
producing cells. It further shows that IL-1β is required for the
differentiation and expansion of human Th17 cells, while IL-6 and
IL-23 may also play a role in the expansion of Th17 memory cells,
although IL-23 levels are low or undetectable in ovarian cancer
(44).
In humans, Th17 differentiation is induced in the
presence of IL-1β, IL-6 and TGF-β cytokines, while the IL-23
cytokine is responsible for the maintenance of Th17 cell survival.
Effector Th17 cells express various cytokines and cell surface
markers, including IL-17A, IL-17F, IL-22, IL-26, CCR6 and TNF-α. It
has been shown that the RORC2 transcription factor in human cells
plays a critical role in Th17 cell differentiation. The
differentiation process of Th17 cells is controlled by many
transcription factors, including RORγt, interferon regulatory
factor 4 (IRF4), runt-related transcription factor 1 (RUNX1), basic
leucine zipper ATF-like transcription factor (BATF) and signal
transducer and activator of transcription 3 (STAT3) (45).
It was found that TGF-β induces differentiation of
Th17 cells by reversing the RORγt suppression related to the
retinoic acid receptor (RAR), mediated by SKI-SMAD4. This has been
proven since SMAD4-deficient T cells differentiate into Th17 cells
in the absence of TGFβ signaling in a RORγt-dependent manner,
unlike in wild-type T cells. Expression of ectopic SMAD4 suppresses
RORγt expression and prevents differentiation of Th17 cells from
SMAD4-deficient T cells. However, TGF-β neutralizes SMAD4-mediated
suppression, since in order to suppress the RORγt function, and
SMAD4 needs to interact with SKI, which is a transcription
repressor that is degraded after the cell is stimulated by TGFβ
(46).
It has been shown that phosphatase and tensin
homologue (PTEN) play an important role in the differentiation of
Th17 cells, promoting the blockade of IL-2 expression. In an in
vitro study it was shown that Th17-specific Pten
deletion (Ptenfl/flIl17acre)
impairs the differentiation of Th17 cells and improves the symptoms
of experimental autoimmune encephalomyelitis, a model of Th17
mediated autoimmune disease. The Pten-deficiency upregulates IL-2
expression and STAT5 phosphorylation, but downregulates STAT3
phosphorylation, which results in inhibition of Th17 cell
differentiation (47). In another
study it was shown that reduced expression of RFX1 transcription
factor in CD4+ T cells from patients with systemic lupus
erythematosus determine overexpression of IL-17A, which increased
the histone H3 acetylation and decreased DNA and H3K9
tri-methylation. This indicates that the differentiation of CD4 + T
cells into Th17 cells may also be influenced by epigenetic
modifications of the DNA. As the functions of RFX1 downstream of
STAT3 and p-Stat3 can inhibit RFX1 expression, a non-canonical
pathway that regulates differentiation of Th17 cells exists
(48).
In humans the differentiation of naïve CD4+ T helper
cells in the Th17 cell occurs in the presence of IL-1β and IL-6,
depending on the transcription factor STAT3. The cytokine IL-23
produced by dendritic cell activity increases and stabilizes the
cells with phenotype Th17 and helps these cells acquire effector
functions. The Th17 cells secrete IL-17, IL-21, and IL-22, which
play a significant role in the immune response against pathogens,
but which also contribute to the pathogenesis of inflammatory and
autoimmune disease. Furthermore, a Th17 cell subpopulation has been
found in many types of cancers, but it is not yet clear whether the
Th17 cell promotes or inhibits tumor progression (49). A representative scheme is proposed in
Fig. 1 to summarize the mechanisms
and cytokines involved in the differentiation process of naïve CD4+
T cells (Th0) in Th17 cells, and the possible role of these cells
in tumor progression.
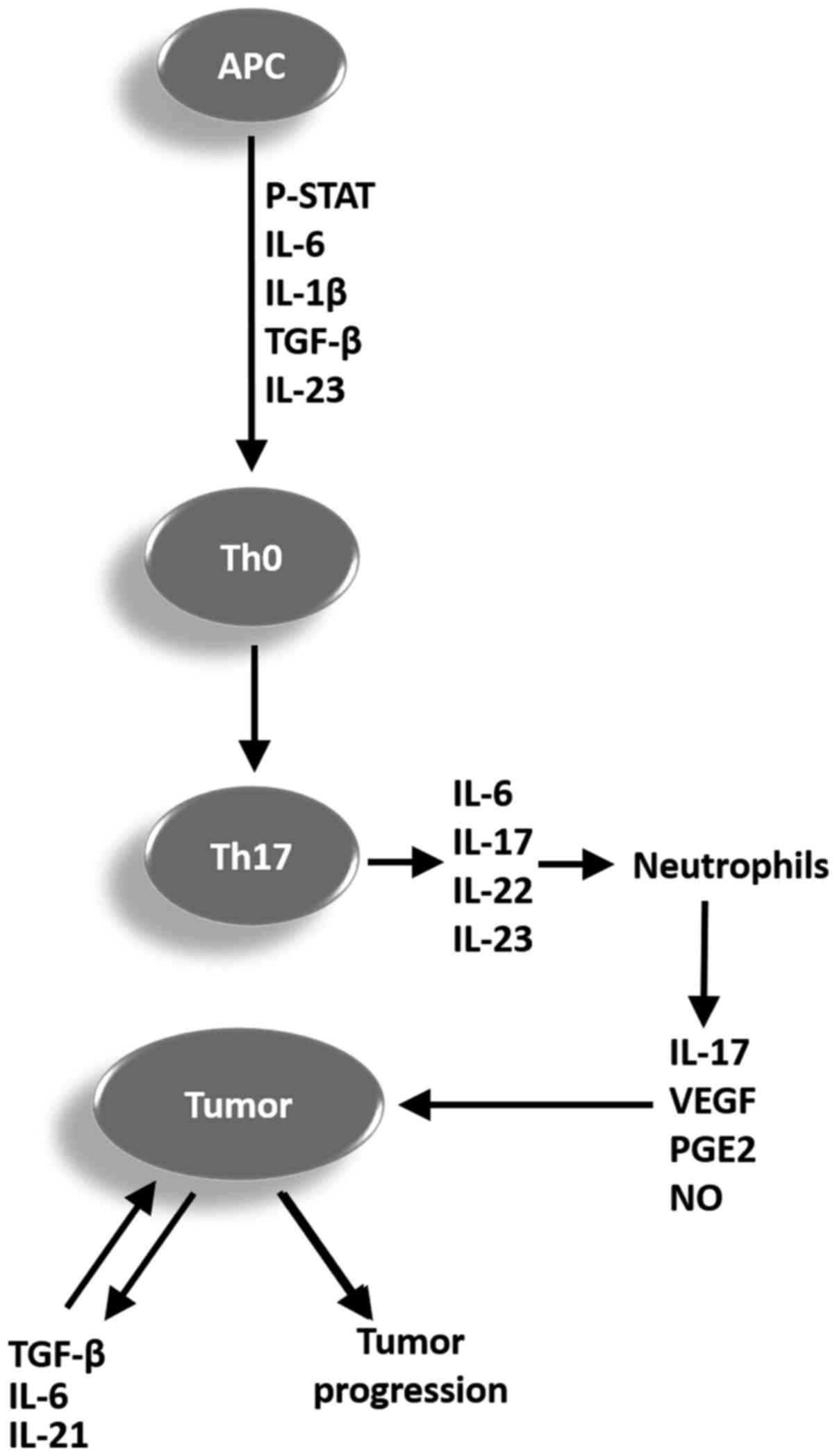 | Figure 1.Representative scheme summarizing the
mechanisms and cytokines involved in the differentiation process of
Th0 in Th17 cells, and the potential function of these cells in
tumor progression. APC, antigen-presenting cell; p-STAT,
phosphorylated signal transducer and activator of transcription;
IL, interleukin; TGF-β, tumor growth factor β; Th, T helper cell;
VEGF, vascular endothelial growth factor; PGE2, prostaglandin E2;
NO, nitric oxide; Th0, CD4+ T cells. |
Biology and functions of T helper 17
cells
The Th17 cell belongs to a subset of T-helper cells,
characterized as producers of tumor necrosis factor-α (TNFα) and
IL-17, which indeed represent a family of cytokines composed of
IL-17A, IL-17B, IL-17C, IL-17D, IL-17E, and IL-17F, in addition to
producing IL-6, IL-21, IL-22, and IL-23 (50). Cells with this phenotype were
identified as a new lineage of T-helper cells with a key role in
host defense against extracellular infectious agents by mediating
effector mechanisms through its cytokines, but it is also
associated with chronic inflammatory disease, autoimmune disorders,
and cancer (51).
An extensive list of diseases has been associated
with the major product of the Th17 cell, the cytokine IL-17, but
the definitive demonstration of its contribution to the
pathogenesis of these diseases is still little known. It is
believed that IL-17 has a regulatory function of the host defense
and of chronic inflammation, but its action could result in damage
to tissues and autoimmunity. Thus, IL-17 establishes a link between
innate and adaptive immunity with disease, and may have
physiological or pathological effects on the immune system.
Evidence supports the notion that the production of unregulated
IL-17 and IL-21 by Th17 cells may contribute to the pathogenesis of
autoimmune diseases (50).
It has been shown in co-cultures that LPS was the
most potent stimulator for activating loaded macrophages of antigen
to conduct the polarization for Th17 response in both CD4+ cells
and CD45RO+ memory cells, while IFN-γ/LPS preferably activates
macrophages to the Th1 phenotype. Antigen-loaded dendritic cells
were more efficient than macrophages in conducting the response of
the Th1 type. However, the ability of Th17 to polarize these two
types of antigen-presenting cells was equivalent and showed to be
strongly dependent on IL-1β. This clearly shows that human
macrophages activated in different ways also have different ways to
present antigens, and this affects adaptive immunity in
vivo, particularly in the conducting polarization to Th17 in
inflamed tissues (52).
Because of the accumulation of Th17 cells in many
types of tumors compared to healthy tissue, the Th17 cells were
associated with both good and bad prognosis regarding tumors,
although the intensity is different from one tumor type to another.
The high plasticity of Th17 cells compared to other cells that
exhibit either protective or pathogenic functions can contribute to
versatile functions of the Th17 cell in the tumor context. Thus, on
one hand Th17 cells can promote tumor growth by angiogenesis
induction mediated by IL-17 and exert an immunosuppressive
function; on the other hand, Th17 cells can lead to an antitumor
immune response through recruitment of immune cells to the
microenvironment of the tumor and activate CD8+ T effector cells
against the tumor, in addition to being able to act directly
through conversion to the Th1 phenotype producing IFN-γ, which in
turn activates more CD8+ cells against the tumor (53).
Despite some advances in knowledge about the role of
Th17 cells in tumor biology, many aspects remain unclear. The
greatest challenge faced by researchers is the plasticity of these
cells within the tumor microenvironment, where they can be
converted into other subsets of Th cells, including Th1 and Treg
cells. Thus, it is necessary to identify the molecular mechanisms
and factors that act in the tumor microenvironment which are
responsible for plasticity and reciprocity in the conversion of
Th1, Th17 and Treg (53). Evidence
suggests that there is a dynamic interaction and mutual conversion
between Treg and Th17 cells in the tumor microenvironment in which
the number of these cells seems to be mutually linked to tumor
progression (54,55). Furthermore, two different
subpopulations of Th17 cells have been found infiltrated in the
tumors, including Th17 cells producing IL-17 and IFN-γ, and Th17
cells producing IL-17 and IL-10. It is possible that the generation
of these two types of Th17 cells is due to variations in the
cellular circumstances and tumor stage (53).
It has been shown that receptors for IL-17 and IL-22
are widely expressed in various epithelial tissues. Therefore, the
cytokines secreted by Th17 cells mediate a critical crosstalk
between the immune system and tissues, and play an essential role
in immunity (56). It has been shown
that Th17 cells have a high degree of functional plasticity, being
endowed with the ability to be converted into Th1 cells producing
IL-17 and IFN-γ, or IL-17 and IL-10, or into Treg cells (57,58).
Because of its high plasticity, Th17 cells exhibit the ability to
shift rapidly to the Th1 phenotype in the inflammatory sites. Thus,
in these places there is usually a dichotomous blend of classic Th1
cells and non-classical Th1 cells derived from Th17 expressing
CD161. Initial studies in this field show evidence suggesting that
the non-classical Th17-derived Th1 cell appears to play a more
important role than the classical Th1 in the maintenance of chronic
inflammation and various autoimmune diseases (59).
Studies conducted in patients with inflammatory
disease of the intestinal tract showed that cytokine IL-23 has an
important role as an inhibitor in the generation of Treg cells
(60). Furthermore, signaling by
IL-23 together with IL-12 both produced by monocyte-derived
dendritic cells (61) promotes IFN-γ
secretion from Th17 cells, thus determining the plasticity of
Th17/Th1 (62). A similar situation
could occur in relation to tumors. This plasticity of Th17 cells
can lead to the conversion of these cells in two subsets of
non-classical Th1 cells, expressing CD161. One of them produces and
secretes IL-17 and IFN-γ, promoting the activation of CD8+ T
lymphocytes with cytotoxic activity, contributing to tumor
regression. The other subset of non-classical Th1 cells produces
IL-17 and IL-10, and unlike the first this subset suppresses the
activity of CD8+ T cells and promotes chronic inflammation, which
may favor tumor progression (57).
The contribution of Th17 cells for immunity against
tumors in humans remains largely unknown. Since its discovery, Th17
cells have been extensively studied in tumors in both animal and
human models. The obtained results suggest the importance of Th17
cells in immunity against tumors, but the functional role of these
cells in tumors remains controversial. The knowledge obtained from
studies in murine tumor models suggests that Th17 cells can exert
pro-or antitumor action, depending on the tumor model used, the
source of IL-17 and Th17 cells, as well as the specific cytokine
and conditions within the tumor (57).
In the tumor microenvironment, cells with the Th17
phenotype may be converted into Treg cells when activated by tumor
antigens in the presence of factors such as IL-2, retinoic acid
(RA), and indoleamine 2,3-dioxygenase (IDO) produced by tumor
cells, thereby producing TGF-β, IL-6, and IL-10. In turn, the Treg
cells being stimulated by cytokines such as IL-6, IL-21, and TGF-β
produced by tumor cells can be converted into Th17 cells, producers
of IL-17, IL-21, and IL-22. The role of Th17 in any study is to
promote or inhibit tumor growth. Therefore, extensive research on
Th17 cells in patients with different tumor types and in different
stages is critical for advancing knowledge in this area (57).
In Fig. 2 a
representative scheme is proposed for summarizing the role of
different cell types, mechanisms, and cytokines involved in the
plasticity process and reciprocity in conversion of Th1, Th17, and
Treg cells for a better understanding of ambiguous functions of the
Th17 cells in the clinical course of tumors, which can contribute
to the regression or progression of tumors.
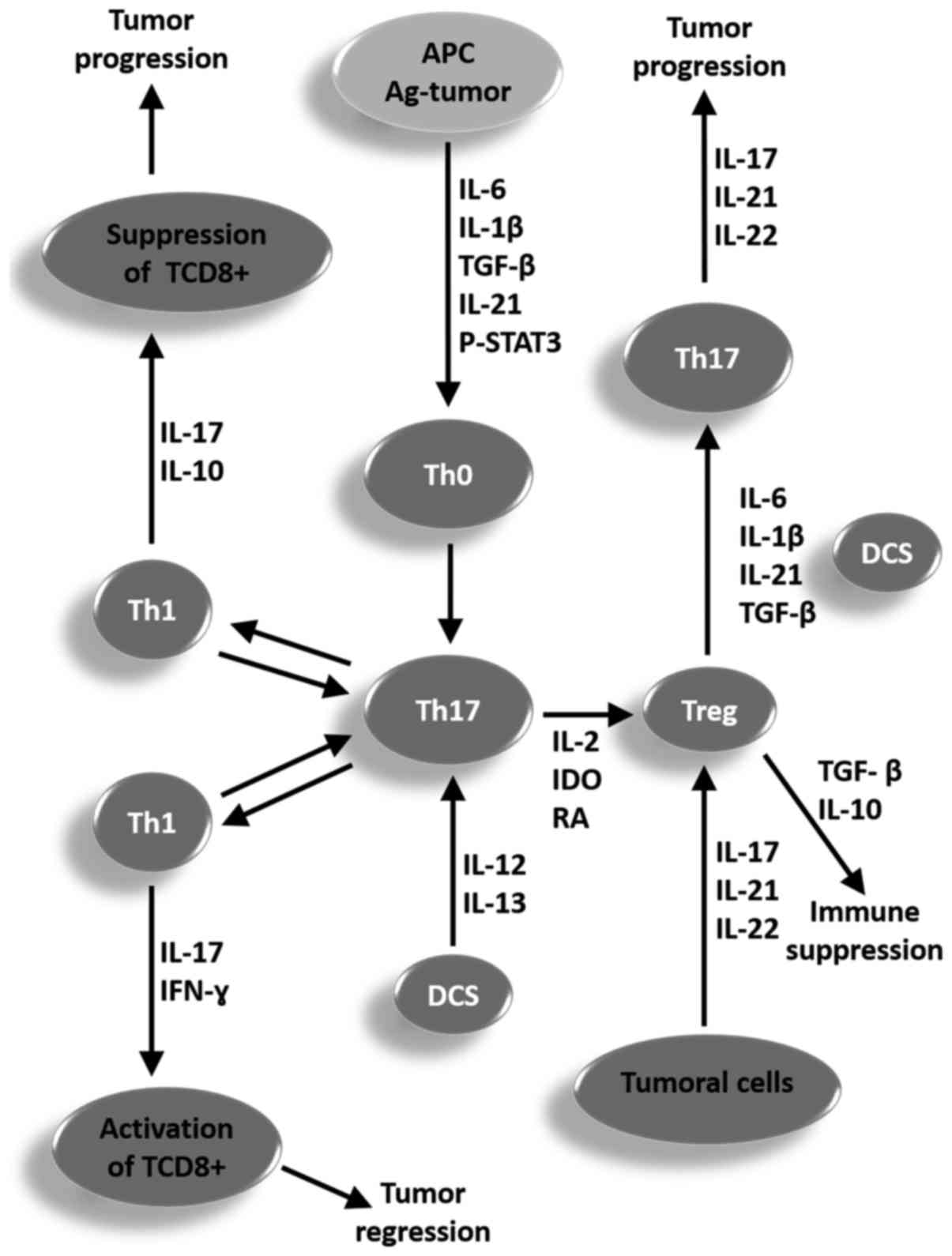 | Figure 2.Representative scheme proposed for
summarizing the function of different cell types, mechanisms and
cytokines involved in the plasticity process and reciprocity in the
conversion of Th1, Th17 and Treg cells for a better understanding
of the ambiguous functions of the Th17 cells in the clinical course
of a tumor, which may contribute to the regression or progression
of a tumor. APC, antigen-presenting cell; p-STAT, phosphorylated
signal transducer and activator of transcription; IL, interleukin;
TGF-β, tumor growth factor β; Th, T helper cell; IDO, indoleamine
2,3-dioxygenase; DCS, dendritic cells; Treg, T regulatory cells;
IFN-γ, type II interferon. |
The subcutaneous injection of lung carcinoma cells
in mice followed by intraperitoneal injection of IL-17 or PBS
showed that the animals inoculated with IL-17 presented
significantly greater tumors with higher levels of metastasis, the
largest infiltration of CCR2+ macrophages and the largest number of
vascular endothelial cells expressing CD34 when compared to animals
injected with PBS. Furthermore, higher levels of mRNA expression of
the vascular endothelial growth factor (VEGF), matrix
metalloproteinase (MMP)2, MMP9, and TNF-α, and lower expression
levels of the migration inhibitory factor and thrombospondin-1 were
observed in animals inoculated with IL-17. The authors concluded
that IL-17 can promote tumor growth by increasing angiogenesis,
metastasis, and the number of CCR2+ macrophages (63). On the other hand, mice with
pre-malignant oral lesions induced by carcinogenic agents presented
a differentiation tendency of the CD4+ helper cells to the
phenotypes Th1 and Th17, as well as drainage of these spleen cells
to the lymph nodes with a shift to the Treg phenotype, followed by
cancer progression. The treatment of the animals with premalignant
oral lesions with an inhibitor of the TGF-β receptor plus IL-23 not
only maintained the Th17 phenotype, but also increased the number
of these cells in the spleen and regional lymph nodes, and slowed
the progression of pre-malignant lesions to cancer (64).
Role of Th17 cells in UCC
Evidence of Th17 cells in playing some role in the
development of cervical cancer is the fact that accumulation of
these infiltrates in the tumor tissue has been associated with a
greater chance of tumor recurrence. Also, it was found that Th17
cells infiltrated into the tumor had an activated phenotype with
increased expression of CCR6, the chemokine receptor CCL20.
Correspondingly, the level of CCL20 in tumor tissues was
significantly higher when compared to normal and non-tumor tissues.
In addition, levels of this chemokine were positively associated
with the amount of Th17 cells (65).
The local immune responses in women with CIN or UCC
were evaluated in two separate studies involving Chinese women. In
the first it was observed that TCD4+ cells expressing Foxp3
accumulated around the tumor in a significantly higher proportion
than found around the CIN. Also, the patients with UCC who had
metastasis of the lymph nodes showed a greater proportion of T
cells expressing Foxp3 compared with those without metastasis
(66). The second study revealed that
patients with CIN and UCC had a greater number of Th17 cells
compared to healthy women. A significant increase in Treg cells and
plasmatic concentrations of cytokines IL-17 and IL-10 was observed
in patients with UCC, but not in those who had CIN. The prevalence
of Th17 cells in the cancer patients was associated with the
clinical stage of disease, the presence of metastasis and
vasoinvasion, while the increase in Treg cells was associated with
the degree of tumor differentiation. Surprisingly, an important
imbalance was found in the Th17/Treg relation, both in patients
with UCC as well as in those with CIN. The imbalance in the
Th17/Treg ratio was significantly higher in women with cancer that
had developed metastasis or vasoinvasion compared with those that
had not. These results reinforce the hypothesis that the imbalance
of Th17/Treg may be involved in the promotion and progression of
UCC (23).
Patients with CIN or UCC have been shown to have a
higher proportion of Th17 cells and Foxp3-expressing T cells among
lymphocytes infiltrated into the tumor when compared to healthy
women with no history of cervical lesions. When comparing the
Th17/T cell ratio expressing Foxp3 among the infiltrated
lymphocytes decreased in the sick women, it was markedly reduced in
lymphocytes infiltrated in the tissues of healthy women. The
cytokine IL-6, TGF-β and IL-10 concentrations were significantly
higher in UCC patients than those in healthy women. This reinforces
the hypothesis that an imbalance of Th17/Foxp3-expressing T cells
plays a critical role in UCC, and that Th17 cells can promote
initiation and progression of this tumor (67).
A higher proportion of Th17 cells and
Foxp3-expressing T cells were found in patients with UCC compared
with the proportion found in healthy women. The ratio of
Th17/Foxp3-expressing T cells among the tumor-infiltrating
lymphocytes (TILs) was more markedly decreased when compared to
normal cervical tissues. Furthermore, the average concentrations of
the cytokines IL-6, TGF-β, and IL-10 were significantly higher in
patients with UCC compared with healthy women. Interestingly, the
amount of Th17 cells in the tumor environment has been positively
correlated with microvessel density. Thus, the imbalance in the
ratio of Th17/Foxp3-expressing T cells may play a critical role in
both promotion and progression of the UCC, where Th17 cells may
contribute to tumor progression by promoting angiogenesis (67).
It has been shown that patients with CIN and UCC
have significantly lower IFN-γ levels than those in the control
group, observing an inverse relationship to the number of Treg
cells, thereby suggesting that Treg cells suppress IFN-γ production
by NK cells and CTL activity. The participation of Th17 cells in
the development of the UCC could be through the IL-17 secretion,
which in turn induces angiogenesis and promotes tumor growth. The
levels of TGF-β, IL-6, IL-10, IL-17 and IL-23 were significantly
higher in patients with CIN and UCC than those in the control
group, with a positive correlation between the percentages of Th17
cells and levels of IL-6, IL-23, and IL-17. The Treg percentage of
cells was positively correlated with elevated levels of TGF-β,
IL-6, and IL-10. This suggests that TGF-β cytokine, IL-6, IL-17,
and IL-23 promote an imbalance in the Th17/Treg relationship, and
this alteration may be a possible mechanism by which Th17 cells
contribute to the development of UCC (68). Once that Treg cells are highly
expressed in UCC, it can be inferred that they play a crucial role
in the development and progression of the disease, since Treg cells
are correlated with the growth, metastasis, and vasoinvasion in UCC
(69). Considering that Th17 cells
can be converted into Treg cells due to their high plasticity, this
could be a possible mechanism of participation of Th17 cells in the
promotion and progression of this tumor type.
Immunopathological analysis of patients with CIN2 or
CIN3 HPV-associated lesions and murine skin with epithelial
hyperplasia and protein-induced HPV16 E7, used as a chronic injury
model induced by HPV, showed a high expression of IL-17 and IL-23,
a major inducer of IL-17. Using the HPV16 E7 transgenic skin graft
animal model, it was demonstrated that IL-17 was able to
efficiently inhibit the immune response against the graft. It was
also found that IL-17 was predominantly produced by CD4(+) T cells
in human CIN2 or CIN3 lesions, and by CD4(+) and γδ T cells in
murine hyperplasic skin lesions. IL-23 and IL-1β, but not IL-18,
induced the production of IL-17 in E7 transgenic skin, showing that
IL-17 induces an immunosuppressive effect in epithelial hyperplasia
associated with HPV. This suggests that blocking IL-17 in the
persistent viral infections can promote the antiviral immunity and
prevent progression to cancer (70).
The active form of the STAT3 transcription factor,
called phosphorylated-STAT-3 (p-STAT3) (71), is found in several types of cancers
including UCC, and it induces differentiation and maturation of
Th17 cells, activating the production of proinflammatory cytokines
such as IL-17, IL-6, IL-22, and IL-23 (72). IL-17 promotes positive feedback of
Stat3, which induces the IL-17 expression even more, in turn
inducing production of the vascular VEGF, prostaglandin E2 (PGE2)
and nitric oxide, which then promotes angiogenesis resulting in the
formation of new blood vessels and tumor growth (57). Furthermore, IL-17 favors accumulation
of neutrophils and increases the resistance of tumor cells to
apoptosis. Thus, IL-17 appears to promote a mechanism of
carcinogenesis by activating the Stat3 signaling pathway, which
triggers the proliferation, migration, and survival of tumor cells
by inducing apoptosis resistance and angiogenesis, facilitating the
growth and progression of tumors (73).
It has been reported that HPV infection induces
constitutive Stat3 activity and a high level of IL-17 expression,
and that the joint action of these two factors may create a
pro-inflammatory response in the colorectal epithelium, which in
turn may promote carcinogenesis and possibly facilitate the
progression of colorectal cancer (74). As HR-HPV infection is the necessary
cause of UCC, it is possible that a similar mechanism may
contribute to development of the disease. Moreover, studies in
humans and animals have shown a positive correlation between
increased expression levels of IL-17 in the mucosa of the uterine
cervix and development of UCC in patients infected with HPV 16 and
18 (the major oncogenic types of HPV) in relation to other
genotypes of the virus (70,75).
In UCC, IL-17 appears to be predominantly produced
by innate myeloid cells such as neutrophils which promote tumor
growth and are correlated with poor prognosis, while Th17 cells
(also producers of IL-17) are correlated with increased survival of
the patients, suggesting that these cells are part of a tumor
suppressor immune response. As IL-6 and IL-23 are also correlated
with poor prognosis of the disease, it has been proposed that these
cytokines may induce IL-17 production by myeloid cells, while Th17
cells somehow counteract this response elicited by IL-6 and IL-32.
These data support the hypothesis that the different types of cells
expressing IL-17 perform different roles in the tumor
microenvironment and have different effects, as indicated by the
correlation between the prevalence of Tregs and the increased
survival of tumor cells (26).
The markers CD3E, IL-6, and VEGFA, and a high
proportion of IL-6/IL-17 combined with low IL-5 expression were
identified as the main factors leading to poor prognosis in UCC.
IL-6 is produced by infiltrating both tumor cells and immune cells,
and directly promotes tumor growth. High levels of IL-17 produced
by Th17 cells, and of IL-5 produced by Th2 cells, have been
correlated with a more favorable prognosis, especially when
compared to levels of expression of Il-6. On the other hand, high
expression of IL-6 and VEGFA, with the latter being an inducer of
angiogenesis, were together correlated with a poor prognosis. IL17
secreted by Th17 cells could counteract the tumor-promoter effect
of IL6, and even more in combined action with the Th2 response
characterized by the production of IL5 (26,76).
Cells producing IL-17
IL-17A is the characteristic cytokine of a unique
subset of CD4+ T cells called Th17 cells (77). However, other cell types including
CD8+αβ, T cells γδ, T cells, LTi cells, neutrophils, eosinophils,
basophils, mast cells, epithelial cells, endothelial cells and
neurons can also produce IL-17A (78–80).
Moreover, ovarian tumor cells, tumor-derived fibroblasts and APCs
may also contribute to the production of IL-17 by secreting
cytokines such as IL-1β, IL-6, TNF-α and TGF-β, which create a
favorable environment for the differentiation and expansion of
IL-17 producing cells (44). IL-17
binds and signals through IL-17 (IL-17R) receptor complexes
composed of IL-17RA transmembrane type 1 and IL-17RC, protein
subunits that are widely expressed in epithelial cells, endothelial
cells and fibroblasts. IL-17/IL-17R binding results in the
secretion of inflammatory mediators, including IL-1, IL-6, IL-8,
IL-23, TNF and various chemokines that further stimulate the
inflammatory cascade (77,79). Studies have shown that IL-17A
represents the prototype member of the IL-17 cytokine family and is
mainly produced by Th17 cells. Unlike IL-17A, IL-17B is widely
expressed in various tissue types and its biological function is
associated with pathologies, particularly to tumors (81).
IL-17A is believed to be an immune mediator that
evolutionarily emerged as an innate cytokine and after evolved and
developed as an adaptive cytokine, since it is more abundantly
expressed by thymus-dependent lymphocytes, including adaptive
memory Th2 cells, Th9 cells, CD8+αβ T cells, as well as the innate
γδ T cells, invariant natural killer T (iNKT) cells and lymphoid
tissue-inducing (LTi) like cells. However, regardless of their
evolutionary past, the importance of IL-17A production by innate
cell populations and the role that these cells play in shaping the
immune response are becoming clearer (79,80).
The differentiation of virgin T cells into Th17
cells appears to involve TGF-β, IL-6, IL-1β, IL-21, and IL-23
signals. In addition, IL-1α or IL-1β act in synergy with IL-23 to
promote IL-17 secretion from memory T cells. Induction or function
of Th17 cells is regulated by cytokines secreted by other T cell
subtypes, including IFN-γ, IL-4, IL-10 and high concentrations of
TGF-β. The main function of IL-17 secreting T cells is to mediate
inflammation, stimulating the production of inflammatory cytokines
such as TNF-α, IL-1β and IL-6, and inflammatory chemokines that
promote the recruitment of neutrophils and macrophages (82).
It has been shown that activated CD8+ T cells in the
presence of the IL-6 or IL-21 cytokines plus TGF-β, analogously to
the CD4+ T cells, differentiate into Tc17 producer cells of IL-17.
These cells have suppressed cytotoxic function together with low
levels of CTL markers such as: T-box transcription factor
eomesodermin, granzyme B and IFN-γ. Instead, these cells express
hallmark molecules of Th17 programming including RORγt, RORα, IL-21
and IL-23R. The expression of RORγt master regulator type 17 is
associated with generation of Tc17, since its overexpression
stimulates the production of IL-17 in the presence of IL-6 or
IL-21. Both the positive regulation of the type 17 programming as
well suppression of differentiation into CTL are dependent on STAT3
(83).
Differentiated Tc17 cells from naïve CD8+ T cells
did not possess cytotoxic molecules and do not show strong
cytotoxicity. However, when Tc17 effector cells were cultured in
the presence of IL-12, they were converted to IFN-γ producing Tc17
cells, which mainly consisted of IL-17/IFN-γ (Tc17/IFN-γ). Tc17
cells converted with IL-12 also acquired cytotoxic function in
addition to the production capacity of IFN-γ, in addition to
showing strong antitumor activity in both in vitro and in
vivo (84). In contrast, in
another study involving patients with UCC or CIN a greater
proportion of Tc17 cells was found in both peripheral blood and
cervical tissues of women with these lesions when compared with
healthy women. In addition, the number of Tc17 cells in UCC tissues
was significantly higher than in the tissues of CIN. The increase
in the number of Tc17 in patients with UCC was related to the
status of pelvic lymph node metastases and increased microvessel
density. Significant correlations between the infiltration of Tc17
cells and Th17 cells or Foxp3 expressing T cells were observed in
UCC and CIN tissues. This indicates that the infiltration of Tc17
cells into the UCC is associated with tumor progression accompanied
by increased cell infiltration of Th17 and of Tregs, and further
promoted tumor vasculogenesis (85).
Cells expressing receptors for the IL-17
family
There is a wide variety of cells expressing
receptors for the cytokine IL-17 family, including T helper type 2,
T helper type 9, basophils, fibroblasts, adipose cells, epithelial
cells, endothelial cells and chondrocytes (80). The receptors of IL-17 cytokine are
mediators of the biological functions of the IL-17 cytokine family
composed of six members ranging from IL-17A to IL-17F to
pro-inflammatory inducing programs (86), in addition to a heterodimeric species
consisting of disulfide-linked IL-17A and IL-17F (87,88). These
cytokines signal through a heterodimeric receptor complex
consisting of the IL-17RA and IL-17RC chains, which is detected in
various cell types (86,89). Although these dimers stimulate many
overlapping signaling pathways, the degree of induction varies
among species with the IL-17A homodimer eliciting more robust
responses than the heterodimer or the IL-17F homodimer (87).
The cytokine receptor family IL-17 is currently
constituted by five members (IL-17RA, IL-17RB, IL-17RC, IL-17RD and
IL-17RE) which share sequence homology and which interact with six
ligands (from IL-17A to IL-17F). All members of this family have a
fibronectin III-like domain in their extracellular part and an
SEF/IL-17R domain (SEFIR) in their intracellular portion (89). The IL-17RA receptor identified there
is often the IL-17 receptor. However, current evidence indicates
that IL-17RA is probably a common receptor used by all members of
the IL-17 family. On the other hand, the other receptors such as
IL-17RB, IL-17RC and IL-17RE were identified as being specific for
IL-17E, IL-17A, IL-17F and IL-17C, respectively. Also IL-17RD,
originally identified as a negative regulator in fibroblast growth
factor signaling, has recently been identified as a negative
regulator of the IL-17A signaling (90,91).
Cytokines IL-17A and IL-17F were believed to be predominantly
secreted by Th17 cells, have been shown to signal through a
heteromeric receptor complex. This complex consists of IL-17RA and
IL-17RC, which are unique transmembrane proteins ubiquitously
expressed in various cell types including epithelial cells,
fibroblasts and astrocytes (92).
Studies have shown that IL-17A and IL-17F are capable of inducing
expression of proinflammatory genes, alone or in synergy with TNFα,
IL-6, G-CSF, IL-1β, CXCL1, CCL20 and matrix metalloproteases
(93), promoting the activation of
NF-κB, MAPK and C/EBP cascades (94).
In addition, other studies have shown that the cytokine IL-17 also
activates the cell signaling pathways Jak-Stat and Jak-PI3K
(95,96). It has also been shown that in addition
to altering gene expression, stimulation by IL-17 can stabilize
mRNAs by promoting the splicing factor disengagement of the mRNA,
preventing its degradation (97,98).
It has been demonstrated that the expression levels
of IL-17RB receptor of IL-17 are increased in gastric cancer
tissues and that IL-17B/IL-17RB signaling plays a critical role in
the progression of this type of tumor. It has also been found that
this signaling is associated with poor prognosis in patients with
breast cancer. In addition, overexpression of IL-17RB was
associated with increased malignancy, metastatic development and
reduced survival in patients with pancreatic cancer. It has been
also observed that the signal triggered by the interaction of
IL-17B with its receptor IL-17RB positively regulates Mesenchymal
stem cells (MSCs) through the activation of the AKT/β-catenin
pathway in gastric cancer. This suggests that IL-17B increases the
progression of gastric cancer by the activation of MSCs (81).
Role of IL-17 in UCC
Potential tumorigenic effect
IL-17 is an inflammatory cytokine with several
functions contributing to host defenses, but also participating in
the pathology of autoimmune diseases, chronic inflammatory diseases
and cancer (70). Reports of
abnormally high levels of IL-17 in a large group of malignant
tumors in humans suggest the involvement of this cytokine in the
tumorigenesis process, according to the currently obtained evidence
(99). One of the first studies to
produce evidence of the role of IL-17 as a tumorigenic factor in
UCC was obtained by Tartour et al (100), in which it was observed that cell
lines derived from the tumor cultured with IL-17 showed increased
levels of IL-6 and IL-8 expression, two inflammatory cytokines that
are usually upregulated in cancer, although no direct effect on
cell proliferation was observed.
In a study conducted by Feng et al (101), evidence was found that elevated
levels of IL-17A can be used as a marker of prognosis in patients
with UCC. This family of cytokines is associated with increased
ability to promote migration and invasion of tumor cells by
increasing the expression and activity of the MMP2 and MMP9 enzymes
in virtue of its capacity to downregulate expression of the
inhibitors of these enzymes (TIMP-1 and TIMP-2) of the signaling
pathway p38/NF-κB (101). In
addition, higher concentrations of IL-17 were more frequent in the
fresh homogenate of tumor tissue obtained from women with UCC
infected with HPV16 or 18 when compared to women infected with
other HPV genotypes. On the other hand, higher levels of macrophage
and granule colony stimulating factor IL-10 and IL-15 were found in
tumors obtained from women infected with different genotypes of
HPV16 and 18 (75).
It was shown that the stroma cells of UCC
predominantly express CCL20, and that this molecule is correlated
with the infiltration of CD4+ cells producing IL-17 in the tumor.
In addition, cells of the UCC seem to be able to emit signals that
stimulate fibroblasts to produce high levels of CCL20 and attract
CD4+ cells producing IL-17 expressing CCR6. It was also found that
IL-6 produced by the tumor cells constitutes an important mediator
of paracrine induction of CCL20, activating the transcription of
the mRNA to this protein in the fibroblast. Thus, the cervical
neoplastic cells modulate the tumor microenvironment by instructing
fibroblasts to support Th17 cell infiltration by IL-6 induction in
a paracrine manner (102).
In patients with UCC, the accumulation of Th17 cells
with activated phenotype and with a marked increase in the
expression of receptor CCR6 was found within the tumor. As
expected, the levels of CCL20 in the tumor tissue were
significantly higher compared to those found in normal tissue, with
a strongly positive correlation with the presence of Th17 cells. In
addition, in vitro migration assay revealed that CCL20
showed an effective chemotaxis to attract Th17 circulating cells.
This suggests that Th17 cells are recruited into the tumor,
preferably by means of the CCR6-CCL20 pathway (65).
The potential role of IL-17 in the modulation of the
carcinogenic phenotype in the UCC was further reinforced by the
finding that IL-17 is produced in vivo by tumor infiltrated
CD4+ cells in patients with disease progression (100). In addition, a tendency for gradual
increase in the Th17 ratio in the total CD4+ cells with the
worsening of the lesion was shown in patients with cervical lesions
associated with HPV, along with the same being observed in relation
to IL-17 levels in serum and in cervical tissue homogenate
supernatant, suggesting that Th17 and IL-17 cells may contribute to
the progression of cervical lesions associated with HPV (103).
The possible tumorigenic role of IL-17 has aroused
great interest in investigating the polymorphisms in the cytokine
gene and its possible association with the increased risk of
cancer, including UCC. A case-control study involving 264 patients
with UCC and 264 healthy women found a close relationship between
the single-nucleotide (SNP) polymorphism common in the IL-17 gene
and the disease. The study revealed that the polymorphic genotype
AA rs2275913 was strongly associated with increased risk of UCC
compared to wild type genotype GG in patients infected with HPV-16
or HPV-18 compared to the control group (99).
This trend was confirmed in four other similar
case-control studies, all involving Chinese women. The first study
analyzed 352 patients with UCC and 352 healthy women in the control
group. It was demonstrated that women with genotype GA and AA for
SNP polymorphism rs2275913 in the IL-17A gene presented a higher
risk of UCC compared with those with the GG genotype. A significant
association was found between the rs2275913 polymorphism and
genital infection by HPV 16 or 18 and risk of developing UCC
(25). The second study looked at 311
cases of UCC and 463 controls without the disease and found that
the frequency of genotype AA for the SNP polymorphism rs2275913 was
significantly higher than that found for the GG genotype in the
patients with UCC compared to the control group. It was also found
that this genetic variation not only increased the susceptibility
for disease, but also its severity (104). In the third study which analyzed 216
cases of UCC and 432 controls without the disease, it was
demonstrated that patients with the AA genotypic variant of SNP
polymorphism rs2275913 showed a significantly higher risk of
developing UCC compared to those with wild-type genotype GG
(105). The fourth study included
306 cases of UCC and 354 controls free of the disease also showed
that women with the AA genotype variant of the polymorphism
rs2275913 had a significantly higher risk of developing UCC
compared to the wild type genotype GG. In addition, in the latter
study it also was found that the A allele of the rs699947
polymorphism of the IL-17 gene was also associated with an
increased risk for UCC in patients infected with HPV 16 or 18
(106). On the other hand, in a
population-based case-control study involving American women, it
was found that the SNP polymorphism rs2275913 of the gene encoding
IL-17 was positively associated with UCC for HPV 16 (107).
Cancer cell proliferation
Although it has been shown that IL-17 did not exert
any in vitro action on the proliferation of cells derived
from cervical cancer, a significant increase in tumor size was
observed when these cells were transfected with a cDNA encoding
IL-17 and transplanted to nude mice when compared to the parent
tumor. It has been found that the action of IL-17 on inducing tumor
growth occurred by increasing IL-6 expression which promotes the
recruitment of macrophages to the tumor site (100).
The phenotypic analysis of cervical cancer cells by
means of immunohistochemistry and immunofluorescence revealed that
IL-17 is predominantly expressed by neutrophils (66%), mast cells
(23%) and innate lymphoid cells (8%), and to a lesser extent only
4% by Th17 cells. A similar distribution was also observed in the
tumor epithelium. A number above the median total of cells
expressing IL-17 in the early stages of the tumor presented an
independent factor of poor prognosis, being associated with less
survival. A high number of neutrophils and the low number of mast
cells also showed a tendency of reduced survival. On the other
hand, cells expressing IL-17 and neutrophils were correlated with
the absence of vasoinvasion. IL-17 has been shown to enhance cell
proliferation or tightness of cervical cancer cell lines, which may
be a mechanism for tumorigenesis in the early stage of tumor
development. This study suggests that IL-17 mainly expressed by
neutrophils promotes tumor growth, being correlated with poor
prognosis in the early stage of the disease. Surprisingly, a high
number of Th17 cells presented as a factor associated with a
favorable prognosis to improve patient survival, suggesting that
Th17 cells are part of a tumor suppressor immune response (26).
Induction of metastasis
Evaluation of IL-17A expression levels and its
clinical significance in tissue samples obtained from 50 cases of
cervical cancer showed that IL-17A expression did not correlate
with patient age, FIGO stage or tumor size, but was significantly
correlated with depth of tumor invasion and status of lymphatic
metastasis. In addition, IL-17A increased the motility of cancer
cells through the upregulation expression of MMP2 and MMP9 and
decreased the expression of their natural endogenous inhibitors
TIMP-1 and TIMP-2. This suggests that IL-17A acts by increasing
tumor cell motility in cervical cancer by being involved in the
remodeling of the extracellular matrix (ECM), thus contributing to
the invasion and metastasis malignant cells, since the cancer cells
need degrading of ECM during the process of metastasis to invade
blood or lymph vessels to reach other tissues and organs, and then
generate a new tumor (101).
Induction of angiogenesis
Inflammation and angiogenesis are two
characteristics of lung adenocarcinoma and are both related to
IL-17. Microvessel density was positively associated with the
expression of IL-17, IL-6, IL-8 and VEGF, reinforcing the
hypothesis that IL-17 contributes to the development of this tumor
by inducing angiogenesis through its stimulatory action of IL-6,
IL-8 and VEGF (108). Interestingly,
UCC bears some resemblance to lung adenocarcinoma, and it was found
that IL-17 presented tumorigenic action in UCC-derived cell lines
as it induced increased expression of IL-6 and IL-8 (61). Furthermore, VEGF expression levels are
also elevated in both precursor and UCC lesions (109), and have been associated with poor
survival in patients with this type of tumor (110). Thus, although it has not yet been
reported, it is likely that IL-17 may also play a role in the
induction of angiogenesis in UCC.
Conclusion
The role of the Th17 immune response in the
development of UCC, as in other types of cancers, has shown to be
ambiguous; in some cases protecting against tumors while in others
promoting them. Th17 may indirectly exert a protective function
against tumors by producing cytokines which promote the recruitment
of effector CD8+ T cells for the tumor site, or directly becoming a
Th1 cell producing IFN-γ that activates CD8+ T cells against
tumors. In other conditions, Th17 cells may act in promoting tumor
growth by producing cytokines such as IL-6, IL-17, and IL-23, which
recruit neutrophils for tumor sites where they produce more IL-17
that leads to chronic inflammation and production of VEGF that
induces angiogenesis. This creates a favorable microenvironment for
tumors or exerts the suppressive function of immune response
against the tumor.
One possible explanation for this ambiguous function
of Th17 cells in relation to UCC is the high plasticity presented
by these cells, which can convert into non-classical Th1 cells,
producers of IL-17 and IFN-γ, or IL-17 and IL-10, or further into T
regulatory cells, according to the types of cytokines present in
the tumor microenvironment. The involvement of Th17 cells in the
tumorigenesis process appears to not only be related to their
ability to produce and secrete IL-17, but also by the secretion of
other cytokines that attract cells to the tumor site. Thus, the
contradictory role of Th17 cells may be due to the other cells that
also produce IL-17. The high levels of IL-17 expression found in
patients with UCC appear to be primarily produced by myeloid cells
of the innate immune response, such as neutrophils. This cytokine
would be responsible for the promotion of tumor growth, and is
related to poor prognosis of the disease.
On the other hand, the IL-17 produced by Th17 cells
was correlated with improved prognosis and the highest survival of
patients, suggesting that cells with this phenotype are part of the
tumor suppressor immune response. It has been proposed that IL-6
and IL-23, whose presence is correlated with poor prognosis in UCC,
would be responsible for activating neutrophils to produce IL-17,
while the IL-17 produced by Th17 cells antagonizes the response
triggered by IL-6 and IL-23. This strengthens the hypothesis that
there are different types of cells producing IL-17, performing
different roles and producing different effects in the tumor
microenvironment.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of data and materials
Not applicable.
Authors' contributions
JJPA, TAAMF and JVF drafted the manuscript. TAAMF,
JMGA, RNOC, DCFL, FLB, VSA and JVF revised the manuscript
critically for important intellectual content. FLB, VSA and JVF
approved the final version to be published.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Catarino R, Petignat P, Dongui G and
Vassilakos P: Cervical cancer screening in developing countries at
a crossroad: Emerging technologies and policy choices. World J Clin
Oncol. 6:281–290. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bosch FX and de Sanjosé S: The
epidemiology of human papillomavirus infection and cervical cancer.
Dis Markers. 23:213–227. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lima EG, de Lima DB, Miranda CA, de Sena
Pereira VS, de Azevedo JC, de Araújo JM, de Medeiros Fernandes TA,
de Azevedo PR and Fernandes JV: Knowledge about HPV and screening
of cervical cancer among women from the metropolitan region of
Natal, Brazil. ISRN Obstet Gynecol. 2013:9304792013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin Y and Zhan FB: Geographic variations
of racial/ethnic disparities in cervical cancer mortality in Texas.
South Med J. 107:281–288. 2014.PubMed/NCBI
|
|
7
|
Forman D, de Martel C, Lacey CJ,
Soerjomataram I, Lortet-Tieulent J, Bruni L, Vignat J, Ferlay J,
Bray F, Plummer M and Franceschi S: Global burden of human
papillomavirus and related diseases. Vaccine. 30 Suppl 5:F12–F23.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vaccarella S, Lortet-Tieulent J, Plummer
M, Franceschi S and Bray F: Worldwide trends in cervical cancer
incidence: Impact of screening against changes in disease risk
factors. Eur J Cancer. 49:3262–3273. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Castellsagué X: Natural history and
epidemiology of HPV infection and cervical cancer. Gynecol Oncol.
110 3 Suppl 2:S4–S7. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bodily J and Laimins LA: Persistent of
human papillomavirus infection: Keys to malignant progression.
Trends Microbiol. 19:33–39. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Muñoz N, Castellsagué X, de González AB
and Gissman L: Chapter 1-HPV in the etiology of human cancer.
Vaccine. 24 Suppl 3:S3/1–10. 2006. View Article : Google Scholar
|
|
12
|
Saavedra KP, Brebi PM and Roa JC:
Epigenetic alterations in preneoplastic and neoplastic lesions of
the cervix. Clin Epigenetics. 4:132012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Daud II, Scott ME, Ma Y, Shiboski S,
Farhat S and Moscicki AB: Association between toll-like receptor
expression and human papillomavirus type 16 persistence. Int J
Cancer. 128:879–886. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mora-García ML and Monroy-García A: Immune
response in cervical cancer. Strategies for the development of
therapeutic vaccines. Rev Med Inst Mex Seguro Soc. 53 Suppl
2:S206–S211. 2015.PubMed/NCBI
|
|
15
|
Stanley MA: Immune responses to human
papilloma viruses. Indian J Med Res. 130:266–276. 2009.PubMed/NCBI
|
|
16
|
Song D, Li H, Li H and Dai J: Effect of
human papillomavirus infection on the immune system and its role in
the course of cervical cancer. Oncol Lett. 10:600–606. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Iwasaki A: Antiviral immune responses in
the genital tract: Clues for vaccines. Nat Rev Immunol. 10:699–711.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sasagawa T, Takagi H and Makinoda S:
Immune responses against human papillomavirus (HPV) infection and
evasion of host defense in cervical cancer. J Infect Chemother.
18:807–815. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stanley M: Immunobiology of HPV and HPV
vaccines. Gynecol Oncol. 109 Suppl 2:S15–S21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Blaskewicz CD, Pudney J and Anderson DJ:
Structure and function of intercellular junctions in human cervical
and vaginal mucosal epithelia. Biol Reprod. 85:97–104. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hervouet C, Luci C, Rol N, Rousseau D,
Kissenpfennig A, Malissen B, Czerkinsky C and Anjuère F: Langerhans
cells prime IL-17-producing T cells and dampen genital cytotoxic
responses following mucosal immunization. J Immunol. 184:4842–4851.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kemp TJ, Hildesheim A, García-Piñeres A,
Williams MC, Shearer GM, Rodriguez AC, Schiffman M, Burk R, Freer
E, Bonilla J, et al: Elevated systemic levels of inflammatory
cytokines in older women with persistent cervical human
papillomavirus infection. Cancer Epidemiol Biomarkers Prev.
19:1954–1959. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Ma D, Zhang Y, Tian Y, Wang X,
Qiao Y and Cui B: The imbalance of Th17/Treg in patients with
uterine cervical cancer. Clin Chim Acta. 412:894–900. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Paradkar PH, Joshi JV, Mertia PN, Agashe
SV and Vaidya RA: Role of cytokines in genesis, progression and
prognosis of cervical cancer. Asian Pac J Cancer Prev.
15:3851–3864. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cong J, Liu R, Wang X, Sheng L, Jiang H,
Wang W, Zhang Y, Yang S and Li C: Association between
interluekin-17 gene polymorphisms and the risk of cervical cancer
in a Chinese population. Int J Clin Exp Pathol. 8:9567–9573.
2015.PubMed/NCBI
|
|
26
|
Punt S, Fleuren GJ, Kritikou E, Lubberts
E, Trimbos JB, Jordanova ES and Gorter A: Angels and demons: Th17
cells represent a beneficial response, while neutrophil IL-17 is
associated with poor prognosis in squamous cervical cancer.
Oncoimmunology. 4:e9845392015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dong C: Targeting Th17 cells in immune
diseases. Cell Res. 24:901–903. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schmitt E, Klein M and Bopp T: Th9 cells,
new players in adaptive immunity. Trends Immunol. 35:61–68. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kaplan MH: Th9 cells: Differentiation and
disease. Immunol Rev. 252:104–115. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Goldszmid RS, Dzutsev A and Trinchieri G:
Host immune response to infection and cancer: Unexpected
commonalities. Cell Host Microbe. 15:295–305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Damsker JM, Hansen AM and Caspi RR: Th1
and Th17 cells: Adversaries and collaborators. Ann N Y Acad Sci.
1183:211–221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mosmann TR, Cherwinski H, Bond MW, Giedlin
MA and Coffman RL: Two types of murine helper T cell clone. I.
Definition according to profiles of lymphokine activities and
secreted proteins. J Immunol. 136:2348–2357. 1986.PubMed/NCBI
|
|
33
|
Wilson JN, Boniface K, Chan JR, McKenzie
BS, Blumenschein WM, Mattson JD, Basham B, Smith K, Chen T, Morel
F, et al: Development, cytokine profile and function of human
interleukin 17-producing helper T cells. Nat Immunol. 8:950–957.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Murphy KM and Reiner SL: The lineage
decisions of helper T cells. Nat Rev Immunol. 2:933–944. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gavin MA, Rasmussen JP, Fontenot JD, Vasta
V, Manganiello VC, Beavo JA and Rudensky AY: Foxp3-dependent
programme of regulatory T-cell differentiation. Nature.
445:771–775. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Josefowicz SZ, Lu LF and Rudensky AY:
Regulatory T cells: Mechanisms of differentiation and function.
Annu Rev Immunol. 30:531–564. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Asadzadeh Z, Mohammadi H, Safarzadeh E,
Hemmatzadeh M, Mahdian-Shakib A, Jadidi-Niaragh F, Azizi G and
Baradaran B: The paradox of Th17 cell functions in tumor immunity.
Cell Immunol. 322:15–25. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Veldhoen M, Uyttenhove C, van Snick J,
Helmby H, Westendorf A, Buer J, Martin B, Wilhelm C and Stockinger
B: Transforming growth factor-beta ‘reprograms’ the differentiation
of T helper 2 cells and promotes an interleukin 9-producing subset.
Nat Immunol. 9:1341–1346. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Eyerich S, Eyerich K, Pennino D, Carbone
T, Nasorri F, Pallotta S, Cianfarani F, Odorisio T, Traidl-Hoffmann
C, Behrendt H, et al: Th22 cells represent a distinct human T cell
subset involved in epidermal immunity and remodeling. J Clin
Invest. 119:3573–3585. 2009.PubMed/NCBI
|
|
40
|
Maddur MS, Miossec P, Kaveri SV and Bayry
J: Th17 cells: Biology, pathogenesis of autoimmune and inflammatory
diseases, and therapeutic strategies. Am J Pathol. 181:8–18. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Peters A, Lee Y and Kuchroo VK: The many
faces of Th17 cells. Curr Opin Immunol. 23:702–706. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ziegler SF and Buckner JH: FOXP3 and the
regulation of Treg/Th17 differentiation. Microbes Infect.
11:594–598. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
van Hamburg JP, Mus AM, de Bruijn MJ, de
Vogel L, Boon L, Cornelissen F, Asmawidjaja P, Hendriks RW and
Lubberts E: GATA-3 protects against severe joint inflammation and
bone erosion and reduces differentiation of Th17 cells during
experimental arthritis. Arthritis Rheum. 60:750–759. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Miyahara Y, Odunsi K, Chen W, Peng G,
Matsuzaki J and Wang RF: Generation and regulation of human CD4+
IL-17-producing T cells in ovarian cancer. Proc Natl Acad Sci USA.
105:15505–15510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nalbant A and Eskier D: Genes associated
with T helper 17 cell differentiation and function. Front Biosci
(Elite Ed). 8:427–435. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang S, Takaku M, Zou L, Gu AD, Chou WC,
Zhang G, Wu B, Kong Q, Thomas SY, Serody JS, et al: Reversing
SKI-SMAD4-mediated suppression is essential for TH17 cell
differentiation. Nature. 551:105–109. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kim HS, Jang SW, Lee W, Kim K, Sohn H,
Hwang SS and Lee GR: PTEN drives Th17 cell differentiation by
preventing IL-2 production. J Exp Med. 214:3381–3398. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhao M, Tan Y, Peng Q, Huang C, Guo Y,
Liang G, Zhu B, Huang Y, Liu A, Wang Z, et al: IL-6/STAT3 pathway
induced deficiency of RFX1 contributes to Th17-dependent autoimmune
diseases via epigenetic regulation. Nat Commun. 9:5832018.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Karczmarczyk A, Karp M and Giannopoulos K:
The role of Th17 cells in tumor immunity Znaczenie limfocytów Th17
w odporności przeciwnowotworowej. Acta Haematol Polonica.
45:155–160. 2014. View Article : Google Scholar
|
|
50
|
Shabgah AG, Fattahi E and Shahneh FZ:
Interleukin-17 in human inflammatory diseases. Postepy Dermatol
Alergol. 31:256–261. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Fu B, Tian Z and Wei H: Th17 cells in
human recurrent pregnancy loss and pre-eclampsia. Cell Mol Immunol.
11:564–570. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Arnold CE, Gordon P, Barker RN and Wilson
HM: The activation status of human macrophages presenting antigen
determines the efficiency of Th17 responses. Immunobiology.
220:10–19. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Guéry L and Hugues S: Th17 cell plasticity
and functions in cancer immunity. Biomed Res Int. 2015:3146202015.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Komatsu N, Okamoto K, Sawa S, Nakashima T,
Oh-hora M, Kodama T, Tanaka S, Bluestone JA and Takayanagi H:
Pathogenic conversion of Foxp3+ T cells into TH17 cells in
autoimmune arthritis. Nat Med. 20:62–68. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gagliani N, Vesely Amezcua MC, Iseppon A,
Brockmann L, Xu H, Palm NW, de Zoete MR, Licona-Limón P, Paiva RS,
Ching T, et al: Th17 cells transdifferentiate into regulatory T
cells during resolution of inflammation. Nature. 523:221–225. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Eyerich S, Eyerich K, Cavani A and
Schmidt-Weber C: IL-17 and IL-22: Siblings, not twins. Trends
Immunol. 31:354–361. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ye J, Livergood RS and Peng G: The role
and regulation of human Th17 cells in tumor immunity. Am J Pathol.
182:10–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gálvez J: Role of Th17 cells in the
pathogenesis of human IBD. ISRN Inflamm. 2014:9284612014.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Annunziato F, Cosmi L, Liotta F, Maggi E
and Romagnani S: Human T helper type 1 dichotomy: Origin, phenotype
and biological activities. Immunology. 144:343–351. 2015.
View Article : Google Scholar
|
|
60
|
Barnes MJ and Powrie F: Regulatory T cells
reinforce intestinal homeostasis. Immunity. 31:401–411. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Davidson MG, Alonso MN, Yuan R, Axtell RC,
Kenkel JA, Suhoski MM, González JC, Steinman L and Engleman EG:
Th17 cells induce Th1-polarizing monocyte-derived dendritic cells.
J Immunol. 191:1175–1187. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Morrison PJ, Ballantyne SJ and Kullberg
MC: Interleukin-23 and T helper 17-type responses in intestinal
inflammation: From cytokines to T-cell plasticity. Immunology.
133:397–408. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wei L, Wang H, Yang F, Ding Q and Zhao J:
Interleukin-17 potently increases non-small cell lung cancer
growth. Mol Med Rep. 13:1673–1680. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Young MR, Levingston CA and Johnson SD:
Treatment to sustain a Th17-type phenotype to prevent skewing
toward Treg and to limit premalignant lesion progression to cancer.
Int J Cancer. 138:2487–2498. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yu Q, Lou XM and He Y: Preferential
recruitment of Th17 cells to cervical cancer via CCR6-CCL20
pathway. PLoS One. 10:e01208552015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wu MY, Kuo TY and Ho HN:
Tumor-infiltrating lymphocytes contain a higher proportion of
FOXP3(+) T lymphocytes in cervical cancer. J Formos Med Assoc.
110:580–586. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hou F, Li Z, Ma D, Zhang W, Zhang Y, Zhang
T, Kong B and Cui B: Distribution of Th17 cells and
Foxp3-expressing T cells in tumor-infiltrating lymphocytes in
patients with uterine cervical cancer. Clin Chim Acta.
413:1848–1854. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen Z, Ding J, Pang N, Du R, Meng W, Zhu
Y, Zhang Y, Ma C and Ding Y: The Th17/Treg balance and the
expression of related cytokines in Uygur cervical cancer patients.
Diagn Pathol. 8:612013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Hou F, Ma D and Cui B: Treg cells in
different forms of uterine cancer. Clin Chim Acta. 415:337–340.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Gosmann C, Mattarollo SR, Bridge JA,
Frazer IH and Blumenthal A: IL-17 suppresses immune effector
functions in human papillomavirus-associated epithelial
hyperplasia. J Immunol. 193:2248–2257. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Shukla S, Mahata S, Shishodia G, Pandey A,
Tyagi A, Vishnoi K, Basir SF, Das BC and Bharti AC: Functional
regulatory role of STAT3 in HPV16-mediated cervical carcinogenesis.
PLoS One. 8:e678492013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Backert I, Koralov SB, Wirtz S, Kitowski
V, Billmeier U, Martini E, Hofmann K, Hildner K, Wittkopf N, Brecht
K, et al: STAT3 activation in Th17 and Th22 cells controls
IL-22-mediated epithelial host defense during infectious colitis. J
Immunol. 193:3779–3791. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Gagliani N, Hu B, Huber S, Elinav E and
Flavell RA: The fire within: Microbes inflame tumors. Cell.
157:776–783. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Li YX, Zhang L, Simayi D, Zhang N, Tao L,
Yang L, Zhao J, Chen YZ, Li F and Zhang WJ: Human papillomavirus
infection correlates with inflammatory Stat3 signaling activity and
IL-17 level in patients with colorectal cancer. PLoS One.
10:e01183912015. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Vidal AC, Skaar D, Maguire R, Dodor S,
Musselwhite LW, Bartlett JA, Oneko O, Obure J, Mlay P, Murphy SK
and Hoyo C: IL-10, IL-15, IL-17, and GMCSF levels in cervical
cancer tissue of Tanzanian women infected with HPV16/18 vs.
non-HPV16/18 genotypes. Infect Agent Cancer. 10:102015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Liang W and Ferrara N: The complex role of
neutrophils in tumor angiogenesis and metastasis. Cancer Immunol
Res. 4:83–91. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Gaffen SL: An overview of IL-17 function
and signaling. Cytokine. 43:402–407. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Bettelli E, Korn T, Oukka M and Kuchroo
VK: Induction and effector functions of T(H)17 cells. Nature.
453:1051–1057. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Cua DJ and Tato CM: Innate IL-17-producin
cells: The sentinels of the immune system. Nat Rev Immunol.
10:479–489. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Pappu R, Ramirez-Carrozzi V and Sambandam
A: The interleukin-17 cytokine family: Critical players in host
defence and inflammatory diseases. Immunology. 134:8–16. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Bie Q, Jin C, Zhang B and Dong H: IL-17B:
A new area of study in the IL-17 family. Mol Immunol. 90:50–56.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Mills KH: Induction, function and
regulation of IL-17-producing T cells. Eur J Immunol. 38:2636–2649.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Huber M, Heink S, Grothe H, Guralnik A,
Reinhard K, Elflein K, Hünig T, Mittrücker HW, Brüstle A, Kamradt T
and Lohoff M: A Th17-like developmental process leads to CD8(+)
Tc17 cells with reduced cytotoxic activity. Eur J Immunol.
39:1716–1725. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Tajima M, Wakita D, Satoh T, Kitamura H
and Nishimura T: IL-17/IFN-γ double producing CD8+ T (Tc17/IFN-γ)
cells: A novel cytotoxic T-cell subset converted from Tc17 cells by
IL-12. Int Immunol. 23:751–759. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Zhang Y, Hou F, Liu X, Ma D, Zhang Y, Kong
B and Cui B: Tc17 cells in patients with uterine cervical cancer.
PLoS One. 9:e868122014. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Hu Y, Shen F, Crellin NK and Ouyang W: The
IL-17 pathway as a major therapeutic target in autoimmune diseases.
Ann N Y Acad Sci. 1217:60–76. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Liang SC, Long AJ, Bennett F, Whitters MJ,
Karim R, Collins M, Goldman SJ, Dunussi-Joannopoulos K, Williams
CM, Wright JF and Fouser LA: An IL-17F/A heterodimer protein is
produced by mouse Th17 cells and induces airway neutrophil
recruitment. J Immunol. 179:7791–7799. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wright JF, Guo Y, Quazi A, Luxenberg DP,
Bennett F, Ross JF, Qiu Y, Whitters MJ, Tomkinson KN,
Dunussi-Joannopoulos K, et al: Identification of an interleukin
17F/17A heterodimer in activated human CD4+ T cells. J Biol Chem.
282:13447–13455. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Gaffen SL: Structure and signalling in the
IL-17 receptor family. Nat Rev Immunol. 9:556–567. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Ron D, Fuchs Y and Chorev DS: Know thy
Sef: A novel class of feedback antagonists of receptor tyrosine
kinase signaling. Int J Biochem Cell Biol. 40:2040–2052. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Mellett M, Atzei P, Horgan A, Hams E,
Floss T, Wurst W, Fallon PG and Moynagh PN: Orphan receptor IL-17RD
tunes IL-17A signalling and is required for neutrophilia. Nat
Commun. 3:11192012. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Trajkovic V, Stosic-Grujicic S, Samardzic
T, Markovic M, Miljkovic D, Ramic Z and Stojkovic Mostarica M:
Interleukin-17 stimulates inducible nitric oxide synthase
activation in rodent astrocytes. J Neuroimmunol. 119:183–191. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Ruddy MJ, Wong GC, Liu XK, Yamamoto H,
Kasayama S, Kirkwood KL and Gaffen SL: Functional cooperation
between interleukin-17 and tumor necrosis factor-alpha is mediated
by CCAAT/enhancer-binding protein family members. J Biol Chem.
279:2559–2567. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhu S and Qian Y: IL-17/IL-17 receptor
system in autoimmune disease: Mechanisms and therapeutic potential.
Clin Sci (Lond). 122:487–511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Huang F, Kao CY, Wachi S, Thai P, Ryu J
and Wu R: Requirement for both JAK-mediated PI3K signaling and
ACT1/TRAF6/TAK1-dependent NF-kappaB activation by IL-17A in
enhancing cytokine expression in human airway epithelial cells. J
Immunol. 179:6504–6513. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Saleh A, Shan L, Halayko AJ, Kung S and
Gounni AS: Critical role for STAT3 in IL-17A-mediated CCL11
expression in human airway smooth muscle cells. J Immunol.
182:3357–3365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Hartupee J, Liu C, Novotny M, Sun D, Li X
and Hamilton TA: IL-17 signaling for mRNA stabilization does not
require TNF receptor-associated factor 6. J Immunol. 182:1660–1666.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Sun D, Novotny M, Bulek K, Liu C, Li X and
Hamilton TA: Treatment with IL-17 prolongs the half-life of
chemokine CXCL1 mRNA via the adaptor TRAF5 and the
splicing-regulatory factor SF2 (ASF). Nature Immunol. 12:853–860.
2011. View Article : Google Scholar
|
|
99
|
Lv Q, Zhu D, Zhang J, Yi Y, Yang S and
Zhang W: Association between six genetic variants of IL-17A and
IL-17F and cervical cancer risk: A case-control study. Tumour Biol.
36:3979–3984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Tartour E, Fossiez F, Joyeux I, Galinha A,
Gey A, Claret E, Sastre-Garau X, Couturier J, Mosseri V, Vives V,
et al: Interleukin 17, a T-cell-derived cytokine, promotes
tumorigenicity of human cervical tumors in nude mice. Cancer Res.
59:3698–3704. 1999.PubMed/NCBI
|
|
101
|
Feng M, Wang Y, Chen K, Bian Z, Jinfang Wu
and Gao Q: IL-17A promotes the migration and invasiveness of
cervical cancer cells by coordinately activating MMPs expression
via the p38/NF-κB signal pathway. PLoS One. 9:e1085022014.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Walch-Rückheim B, Mavrova R, Henning M,
Vicinus B, Kim YJ, Bohle RM, Juhasz-Böss I, Solomayer EF and Smola
S: Stromal fibroblasts induce CCL20 through IL6/C/EBPβ to support
the recruitment of Th17 cells during cervical cancer progression.
Cancer Res. 75:5248–5259. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Xue J, Wang Y, Chen C, Zhu X, Zhu H and Hu
Y: Effects of Th17 cells and IL-17 in the progression of cervical
carcinogenesis with high-risk human papillomavirus infection.
Cancer Med. 7:297–306. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Quan Y, Zhou B, Wang Y, Duan R, Wang K,
Gao Q, Shi S, Song Y, Zhang L and Xi M: Association between IL17
polymorphisms and risk of cervical cancer in Chinese women. Clin
Dev Immunol. 2012:2582932012. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Li L, Tian YL, Lv XM, Yu HF, Xie YY, Wang
JD and Shi W: Association analysis of IL-17A and IL-17F
polymorphisms in Chinese women with cervical cancer. Genet Mol Res.
14:12178–12183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Sun LX, Wang XB and Huang XJ: Association
analysis of rs2275913G>A and rs763780T>C interleukin 17
polymorphisms in Chinese women with cervical cancer. Genet Mol Res.
14:13612–13617. 2015.PubMed/NCBI
|
|
107
|
Hardikar S, Johnson LG, Malkki M,
Petersdorf EW, Galloway DA, Schwartz SM and Madeleine MM: A
population-based case-control study of genetic variation in
cytokine genes associated with risk of cervical and vulvar cancers.
Gynecol Oncol. 139:90–96. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Huang Q, Duan L, Qian X, Fan J, Lv Z,
Zhang X, Han J, Wu F, Guo M, Hu G, et al: IL-17 promotes angiogenic
factors IL-6, IL-8, and Vegf production via Stat1 in lung
adenocarcinoma. Sci Rep. 6:365512016. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Mandic A, Knezevic Usaj S and Ivkovic
Kapicl T: Tissue expression of VEGF in cervical intraepithelial
neoplasia and cervical cancer. J BUON. 19:958–964. 2014.PubMed/NCBI
|
|
110
|
Zhang J, Liu J, Zhu C, He J, Chen J, Liang
Y, Yang F, Wu X and Ma X: Prognostic role of vascular endothelial
growth factor in cervical cancer: A meta-analysis. Oncotarget.
8:24797–24803. 2017.PubMed/NCBI
|














