Introduction
The global incidence of breast cancer has been
increasing since the end of 1980s (1). According to the ‘Global Cancer Report
2014’ issued by the World Health Organization, there were 1.677
million new cases of breast cancer in females globally in 2012,
with China accounting for 11.2% of cases (2). At present, breast cancer has the highest
incidence rate in China of any malignant tumor for women (3). The majority of patients with breast
cancer are the middle or late stages upon initial diagnosis, thus
the curative effects are often unsatisfactory (4). The primary cause of mortality for
patients with breast cancer is metastasis (5). However, the underlying molecular
mechanism of breast cancer metastasis remains unresolved.
Therefore, it is considered important to study and understand the
underlying molecular mechanism associated with breast cancer
progression and metastasis, in order to provide a scientific basis
for the development of effective treatment strategies for breast
cancer, and to improve the survival rate of patients with breast
cancer.
In recent years, a study revealed that the
occurrence and development of breast cancer is associated with the
abnormal expression of microRNA (miRNA) (6). miRNA is a class of endogenous non-coding
RNA. It widely exists in eukaryotic cells and has a regulatory
effect on cell proliferation, apoptosis, migration and invasion
(7–9).
It also serves an important function in the occurrence and
development of tumors (10–12). A number of studies have revealed that
miR-30a is downregulated in a variety of tumor tissues, including
ovarian cancer (13), lung cancer
(14), hepatocellular carcinoma
(15) and gastric cancer (16). Sestito et al (13) reported that the expression level of
miR-30a in ovarian cancer tissue was significantly reduced, and
overexpression of miR-30a in ovarian cancer cells was able to
decrease the activity of phosphoinositide 3-kinase/protein kinase B
and reduce the activation of the mitogen-activated protein kinase
signaling pathway, thus inhibiting the proliferation and invasion
of ovarian cancer cells, and downregulating the levels of
epithelial-mesenchymal transition (EMT)-associated markers in
ovarian cancer cells. Tang et al (14) revealed that miR-30a expression levels
were significantly decreased in lung cancer tissues and that
miR-30a expression level was closely associated with the
development and prognosis of lung cancer. Liu et al
(17) revealed that in hepatocellular
carcinoma, miR-30a was able to regulate the expression level of
Snail, which was associated with EMT, and inhibit the migration and
invasion of hepatocellular carcinoma. Liu et al (18) reported that miR-30a expression was
abnormal in gastric carcinoma tissue, and miR-30a was able to alter
Vimentin expression by regulating the expression of runt-related
transcription factor 3, which was associated with EMT and therefore
involved in the occurrence and development of gastric cancer.
Thus far, there are few studies on the change in
miR-30a expression in breast cancer or its association with the
development, proliferation and invasion of breast cancer.
Therefore, in the present study, miR-30a expression in breast
cancer tissue and breast cancer cell lines was detected using
reverse transcription-polymerase chain reaction (RT-PCR), and
miR-30a mimic and anti-miR-30a were transfected into the breast
cancer cell line SK-BR-3 to observe the effects of miR-30a
expression on the proliferation and invasion of SK-BR-3 cells. Zinc
finger protein Snai 1, the target gene of miR-30a, was verified and
the effect of miR-30a on the expression level of Snail protein was
detected in order to identify a possible novel therapeutic target
for the treatment of breast cancer.
Materials and methods
Clinical data
The tumor tissue and paracancerous tissue of
patients with breast cancer (n=43) who were treated in the No. 254
Hospital of the People's Liberation Army, (Tianjin, China) between
June 2015 and February 2016 were collected. These patients were
aged between 19 and 58 years old, with an mean age of 31.42±21.46
years. Tumor tissue specimens and adjacent normal tissue specimens
from patients were surgically collected. All specimens were
verified as breast cancer tissues by two pathologists at No. 254
Hospital of the People's Liberation Army following surgery, while
normal tissues were confirmed to have no cancer invasion. The
specimens were stored in liquid nitrogen within 10 min of being
obtained. Tumor (T) staging was defined by the length of the tumor
according to the following criteria: Tl≤2 cm; 2 cm <T2≤5 cm;
T3>5 cm. Tumors directly invading skins and chest walls (costa,
intercostal muscles, serratus anterior muscle) were defined as T4,
without regard to their size. In total, there were 16 cases of T1,
12 cases of T2, 9 cases of T3 and 6 cases of T4. Written informed
consent was obtained from all patients. The present study was
approved by the ethics committee of the No. 254 Hospital of the
People's Liberation Army.
Cells and reagents
Normal mammary epithelial MCF-10A cells, low
invasive breast cancer MCF-7 cells, moderate metastatic and
invasive breast cancer SK-BR-3 cells (19) and high metastatic and invasive human
breast cancer MDA-MB-231 cells were all purchased from the
Preclinical Medicine Institute of the Chinese Academy of Medical
Sciences (Beijing, China). These cells were cultured in Dulbecco's
modified Eagle's medium containing 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Horseradish
peroxidase (HRP)-conjugated goat anti mouse immunoglobulin G was
purchased from OriGene Technologies, Inc. (Rockville, MD, USA). The
transfection reagents used were Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) and TRIzol® (Life
Technologies; Thermo Fisher Scientific, Inc.). Negative control
(NC), miRNA-30a mimic, anti-NC and anti-miRNA-30a were purchased
from (Ambion; Thermo Fisher Scientific, Inc.). Boyden chambers used
in the invasion assay were purchased from EMD Millipore (Billerica,
MA, USA). The RT-PCR primers were synthesized by Sangon Biotech
Co., Ltd. (Shanghai, China). A luciferase assay kit was purchased
from Promega Corporation (Madison, WI, USA). Antibodies against
Snail, E-cadherin, N-cadherin and Vimentin were purchased from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
RT-PCR
Total RNA were extracted from the tissue samples
using TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) cDNAs
were synthesized with the Prime-Script RT reagent kit (Takara Bio,
Inc., Otsu, Japan), SYBR-Green (Takara Bio, Inc., Otsu, Japan) was
used as the fluorophore, according to the manufacturer's protocol.
The thermocycling conditions were as follows: 95°C for 5 min, 95°C
for 10 sec, 60°C for 20 sec, for a total of 40 cycles. Primer
sequences were as follows: Snail forward primer,
5′-GTGGGATGGCTGCCAGC-3′; reverse primer,
5′-TGCAGGACTCTAATCCAAGTTTAC-3′; forward primer of internal control
U6, 5′-CTCGCTTCGGCAGCACA-3′; reverse primer,
5′-AACGCTTCACGAATTTGCGT-3′. The obtained cDNA were identified by
standard agarose gel electrophoresis.
Clone formation test
A total of 1×104 SK-BR-3 cells were
seeded into each well of 6-well plate, and incubated overnight at
37°C, then transfected with NC, miR-30a mimic, anti-NC and
anti-miR-30a at a concentration of 200 nmol/l. Lipofectamine 2000
or Oligofectamine (Invitrogen; Thermo Fisher Scientific, Inc.) was
used for transfection according to the manufacturer's protocol. At
48 h following transfection, cells were used for further
experiments. Primer sequences were as follows: NC sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′; miR-30a mimic sense,
5′-UGUAAACAUCCUCGACUGGAAG-3′ and antisense,
5′-UCCAGUCGAGGAUGUUUACAUU-3′; anti-NC, 5′-CAGUACUUUUGUGUAGUACAA-3′;
anti-miR-30a, 5′-CUUCCAGUCGAGGAUGUUUACA-3′. The mimic was purchased
from Guangzhou RiboBio Co., Ltd. (Guangzhou, China). Cells were
inoculated in six-well plates at a density of 5×103
cells per well, and cultured for 72 h. The formed clones were fixed
with 70% methanol at 4°C overnight, and stained with 20% methanol
containing 0.1% crystal violet for 30 min at 37°C. Colonies that
contained >50 cells were counted under a light microscope
(magnification, ×100; Eclipse TiU, Nikon Corporation., Tokyo,
Japan).
Invasion test
The invasive ability of cells was detected using
Boyden chambers. The cells used in this experiment were stained
with 0.1% trypan blue (Beijing Solarbio Science and Technology Co.,
Ltd., Beijing, China) for 5 min at room temperature counted under
the a light microscope (magnification, ×200) (Eclipse TiU; Nikon
Corporation) in 5 randomly selected fields, and then suspended. A
total of 1×105 cells were placed in the upper chamber.
Matrigel (5–10 µg in 50 µl per well) was pre-coated in a 24-well
plate, and dried overnight in a laminar airflow hood and serum-free
medium (StemPro®MSC SFM; Invitrogen: Thermo Fisher
Scientific, Inc.) was added up to 300 µl. A total of 500 µl
serum-free medium (StemPro®MSC SFM; Invitrogen: Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(Thermo Fisher Scientific Inc.) was added to the lower chamber.
After 12 h of incubation, the non-invasive cells were removed from
the upper chamber with a cotton swab. The cells in the lower
chamber were fixed with 5% glutaraldehyde for 5 min at 4°C and
stained with 0.1% crystal violet for 30 min at 37°C. The number of
invaded cells was counted under the microscope (×200 magnification)
(Eclipse TiU, Nikon Corporation) in 5 randomly selected fields.
Forecasting possible target genes of
miR-30a using bioinformatics methods
According to Sequence complementarity (the 706–713
site in the 3′untranslated region (3′UTR) of Snail mRNA was a
possible site of action of miR-30a), Snail was selected as the
target gene of miR-30a using the target gene prediction software
TargetScans (http://www.targetscan.org/vert_71/).
Verification of target gene by
luciferase reporter gene test
The renilla luciferase gene with the 3′UTR was
PCR-amplified from the pRL-SV40 plasmid (Promega Corporation) and
inserted into the 3′UTR site of pMIR-REPORT. Luciferase expression
constructs were generated by cloning of the 3′UTRs of Snail into
the modified pMIR vector (pMIR-RL). Amplified 3′ UTR were cloned
downstream of the firefly luciferase coding region in the
pMIR-REPORTTM (Ambion; Thermo Fisher Scientific, Inc.). The
pMIR-luciferase reporter vectors (100 ng) were transfected into
SK-BR-3 cells in 6-well plates together with miR-30a
oligonucleotides (100 nM) by using Lipofectamine 2000. After 72 h
of culture, luciferase activity was detected with Dual-Luciferase
Reporter assay system (Promega Corporation) according to the
manufacturer's protocol. Firefly luciferase activity was normalized
to that of Renilla luciferase.
Western blot analysis
The protein concentration was determined by one
method of quantification (LI-COR Biosciences, Lincoln, NE, USA), a
total of 50 µg protein was loaded in to each well and were
separated on 8% SDS-PAGE and 5% concentrated gel, then transferred
to polyvinylidene difluoride membranes in a half-dry state and
blocked at room temperature for 2 h with Tris-Buffered Saline and
Tween-20 (TBST) containing 5% bovine serum albumin (BSA, Abcam,
Cambridge, UK). The primary antibody (1:800; rabbit polyclonal
anti-iNOS, sc-650; Santa Cruz Biotechnology) was added and
incubated at 4°C overnight with agitation. The membrane was washed
with 0.1% TBST three times, for 5 min each time. HRP-conjugated
secondary antibody (1:10,000; goat anti-rabbit IgG (H+L)-HRP
conjugate, cat. no. 170-6515, Bio-Rad Laboratories, Inc.) was added
and incubated at room temperature for 1 h. Following a wash with
0.1% TBST, the bands on the membrane were colored with Supersignal
West Femto HRP-sensitive chemiluminescence substrate (Pierce;
Thermo Fisher Scientific, Inc.). Actin was used as an internal
control. All the experiments were repeated at least three times.
The immunoreactive proteins were detected using the ECL Plus
Western Blotting Detection system (GE Healthcare, Chicago, IL,
USA), visualized by Molecular Imager® VersaDoc™ MP
Imaging systems (Bio-Rad Laboratories, Hercules, CA, USA) and the
densitometry were measured by the professional image analysis
software, ImagePro Plus 6.0 (IPP6.0, Media Cybernetics, Bethesda,
USA).
Statistical analysis
Statistical analysis was conducted using SPSS
software (version 17.0; SPSS, Inc., Chicago, IL, USA). Measurement
data are expressed as mean ± standard deviation. Comparisons
between data in two groups were conducted using Student's t-test.
Multi-group comparisons were conducted using univariate analysis of
variance. Inter-group comparisons were conducted using
Student-Newman-Keuls (SNK) method. P<0.05 was considered to
indicate a statistically significant difference.
Results
RT-PCR
MiR-30a expression levels in breast cancer tissues
and cell lines are demonstrated by the RT-PCR results (Fig. 1A). The relative expression level of
miR-30a was 5.43±0.51 in normal tissues; 4.06±0.69 in stage T1
breast cancer tissues; 3.11±0.33 in stage T2 breast cancer tissues;
1.32±0.39 in stage T3 breast cancer tissues; and 0.71±0.23 in stage
T4 breast cancer tissues. With the increase of pathological grade
of breast cancer, miR-30a expression level gradually decreased
(F=223.806, P<0.001). As presented in Fig. 1B, the results of RT-PCR revealed that
the relative expression level of miR-30a was 4.22±0.41 in normal
mammary epithelial cell MCF-10A; 3.07±0.39 in the low invasive
breast cancer cell line MCF-7; 2.16±0.31 in the moderately
metastatic and invasive breast cancer cell line SK-BR-3; and
1.08±0.19 in highly metastatic and invasive human breast cancer
cell line MDA-MB-231. With the increase of the malignant degree of
breast cancer cells, expression level of miR-30a decreased
(F=47.260, P<0.001).
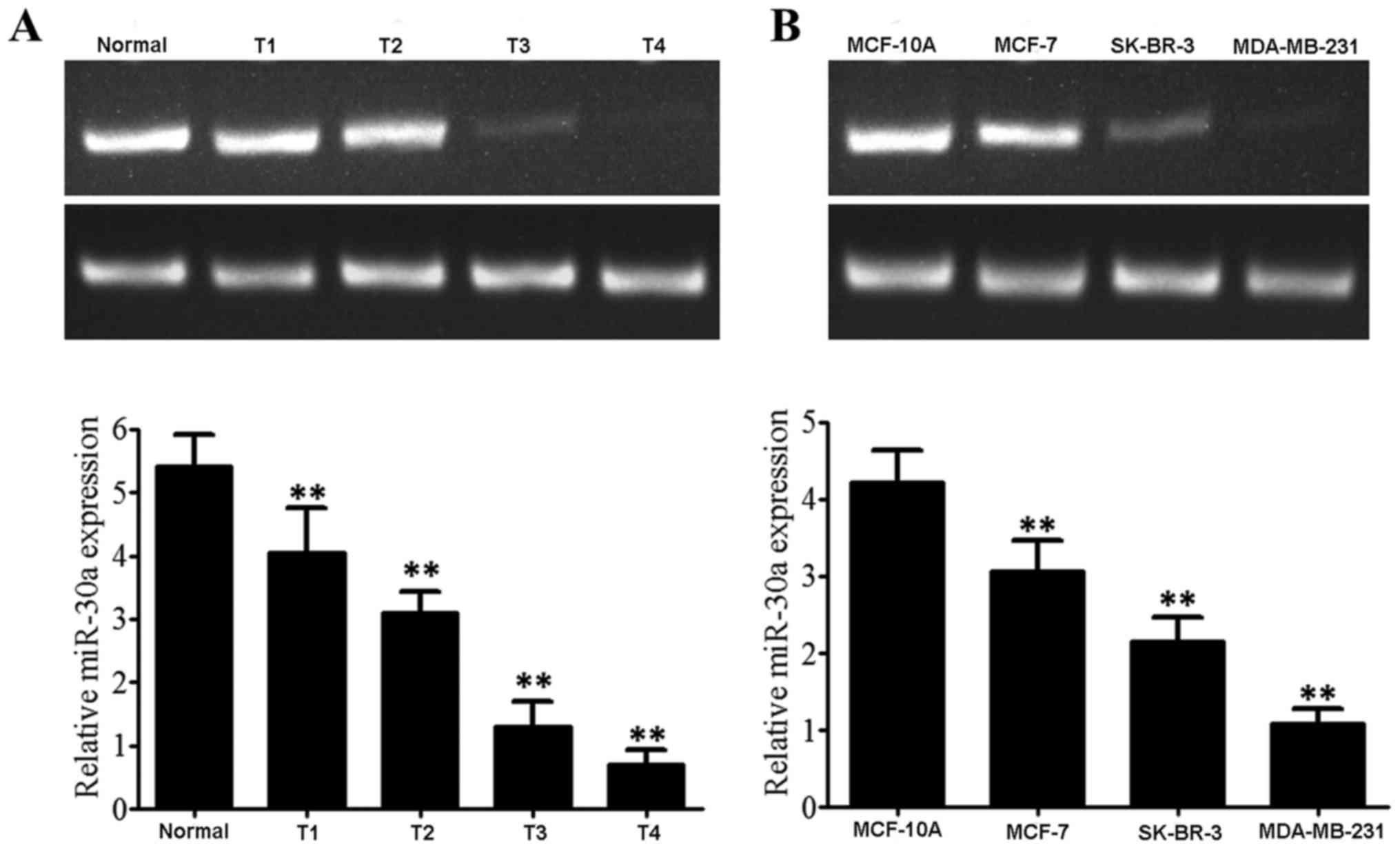 | Figure 1.The expression of miR-30a mRNA in
breast cancer tissues and cells. (A) miR-30a mRNA expression in
breast cancer tissues: 1, Paracancerous tissue; 2, T1; 3, T2; 4,
T3; 5, T4; (B) miR-30a expression in breast cancer cells, 1,
MCF-10A cells; 2, MCF-7 cells; 3, SK-BR-3 cells; 4, MDA-MB-231
cells; **P<0.01 vs. negative control. miRNA, microRNA. |
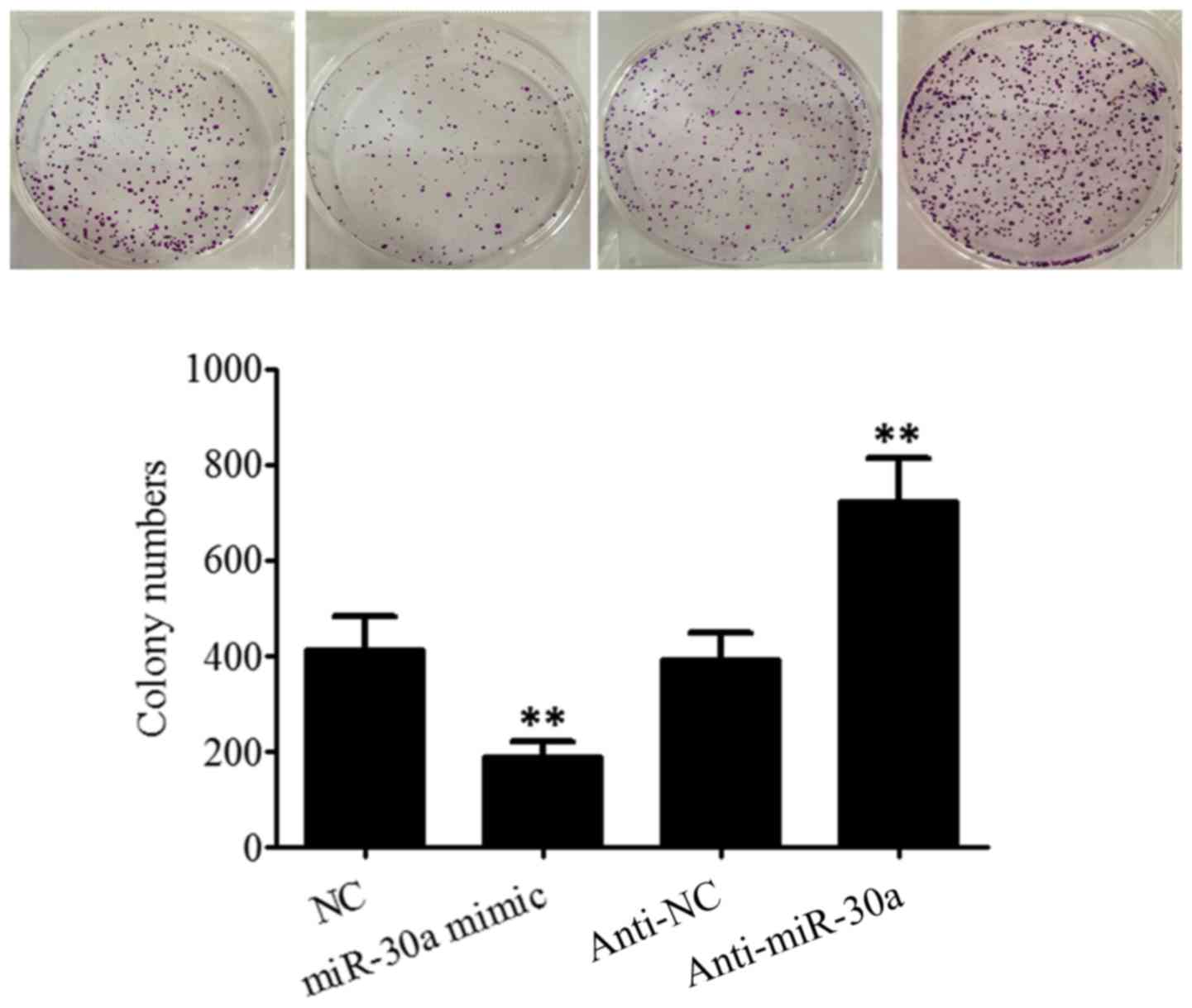
Effect of miR-30a on the proliferation
of breast cancer cells
The effect of miR-30a on the proliferation of
SK-BR-3 breast cancer cells was detected using a clone formation
test. The results (Fig. 2) revealed
that the number of clones of SK-BR-3 cells was 415.63±69.22 in the
NC group; 189.74±31.37 in the miR-30a mimic group; 393.68±57.16 in
the anti-NC group; and 725.82±89.57 in the anti-miR-30a group.
Transfection with miR-30a mimic significantly inhibited the
proliferation of SK-BR-3 cells (t=5.148, P=0.003), while
transfection with anti-miR-30a significantly increased the
proliferation of SK-BR-3 cells (t=5.414, P=0.003). This suggests
that, miR-30a was able to inhibit the proliferation of SK-BR-3
breast cancer cells. It was also identified that, transfection with
miR-30a inhibited the proliferation of MDA-MB-231 human breast
cancer cells which exhibit high invasive and metastatic abilities.
Inhibiting miR-30a expression was observed to promote the
proliferation of breast cancer MCF-7 cells which exhibit a low
invasive ability (data not shown).
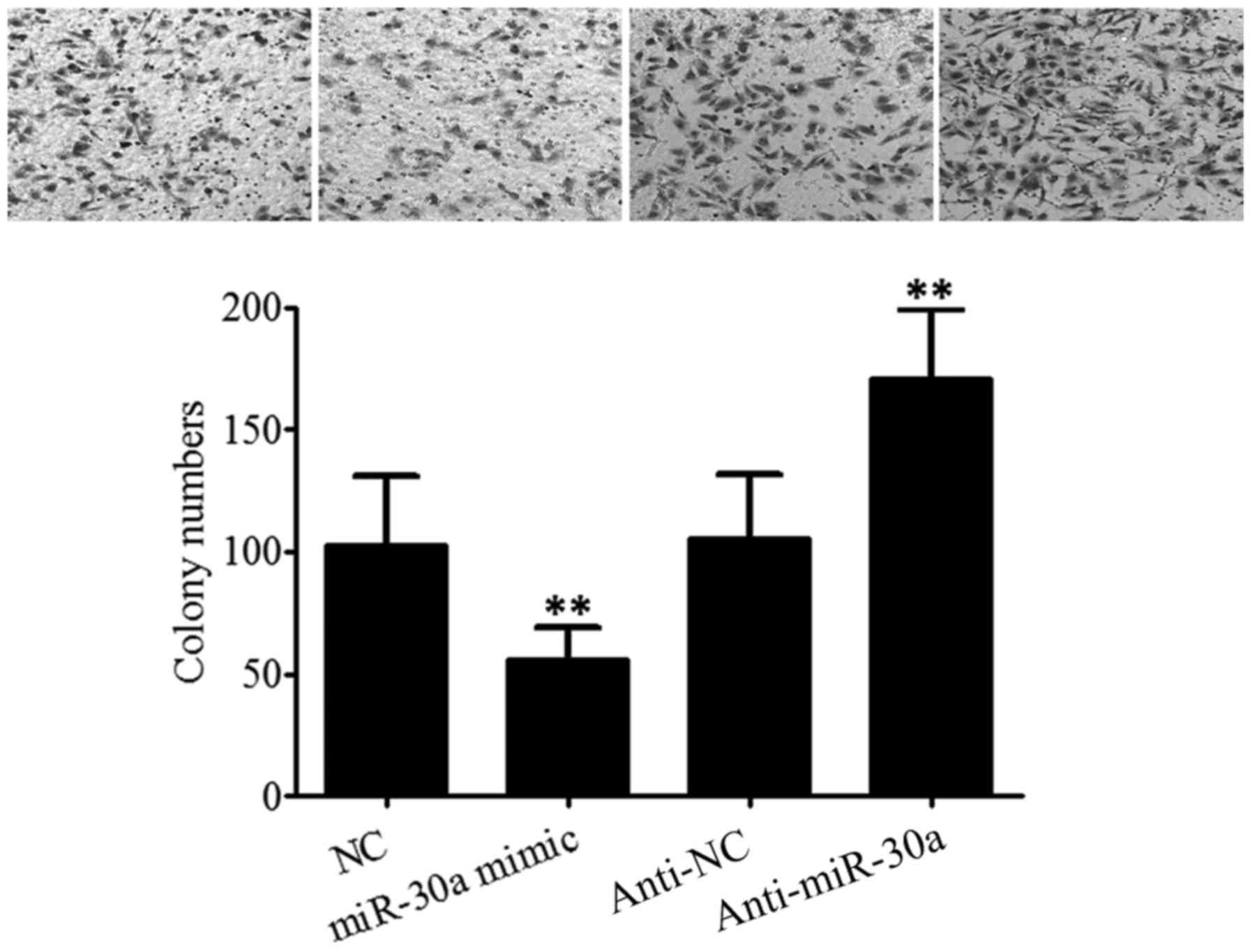
Effect of miR-30a on the invasive
ability of breast cancer cells
As presented in Fig.
3, the results of the cell invasion assay revealed that the
quantity of invaded SK-BR-3 cells was 102.6± 28.5 in the NC group,
55.7±13.6 in the miR-30a mimic group, 105.5±25.9 in the anti-NC
group and 171.2±27.2 in the anti-miR-30a group. Transfection with
miR-30a mimic significantly inhibited the invasive ability of
SK-BR-3 cells (t=2.572, P=0.031), while transfection with
anti-miR-30a significantly increased the invasive ability of
SK-BR-3 cells (t=2.989, P<0.020). This suggests that miR-30a was
able to inhibit the invasion of SK-BR-3 cells. The results of the
present study suggest that transfecting miR-30a may inhibit the
invasiveness of MDA-MB-231 cells which exhibit high invasive and
metastatic abilities. Furthermore, inhibition of miR-30a expression
promoted the invasion of breast cancer MCF-7 cells which exhibit
low invasive ability (data not shown).
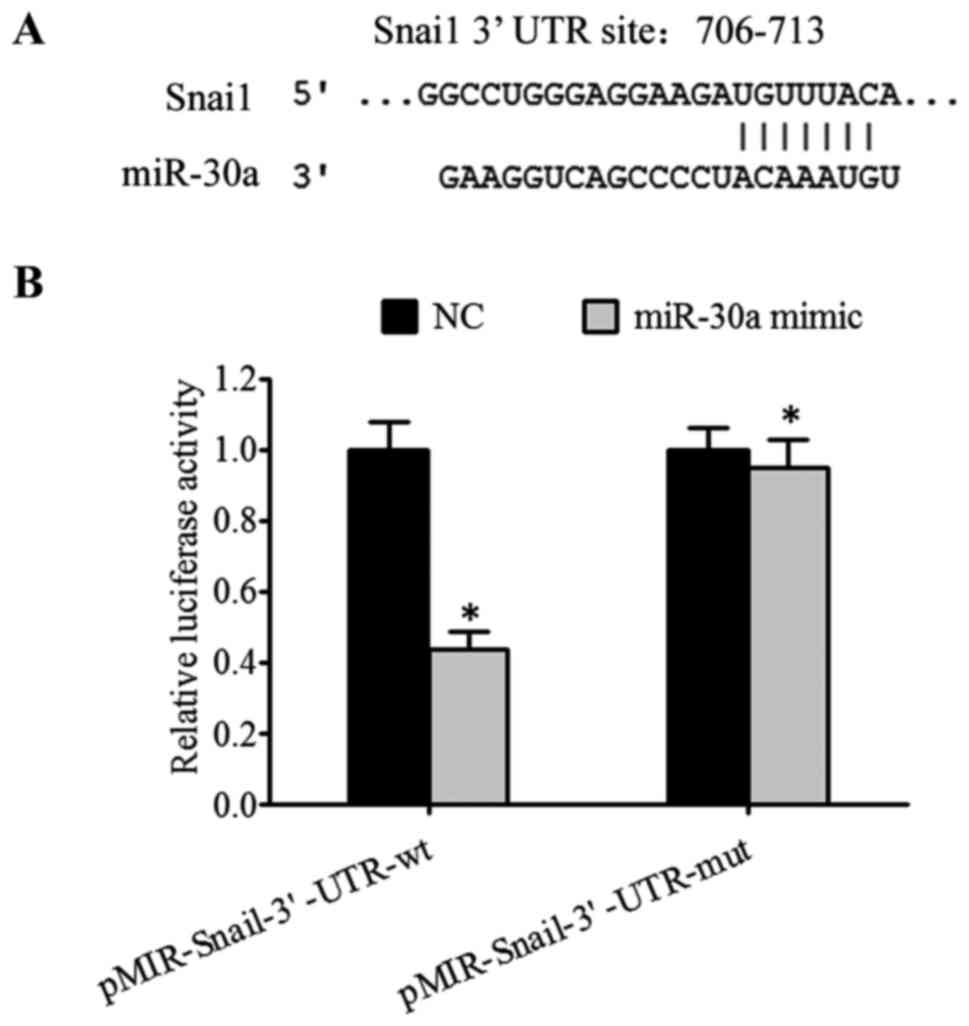
Forecasting miR-30a target gene by
TargetScan software
The target gene prediction software TargetScans
indicated that Snail may be the target gene of miR-30a. The
predicted binding site of miR-30a, Snail 3′-UTR, is presented in
Fig. 4A. The results of a luciferase
reporter gene assay revealed that following co-transfection with
wild-type pMIR-Snail 3′-UTR vector and miR-30a, luciferase activity
was significantly reduced (t=10.337, P<0.001), however,
following co-transfection with mutant pMIR-Snail 3′-UTR vector and
miR-30a, luciferase activity did not change significantly. The
luciferase activity did not significantly change (P>0.05)
following co-transfection of wild type pMIR- Snai1 3′-UTR vector
and miR-30b, miR-30c, miR-30d and miR-30e. This suggests that there
is a direct interaction between miR-30a and Snail (Fig. 4B).
Snail expression in breast cancer
tissues and cell lines
The results of western blotting (Fig. 5A) revealed that the relative
expression level of Snail was 1.00±0.08 in normal tissues,
1.65±0.19 in stage T1 breast cancer tissues, 2.13±0.33 in stage T2
breast cancer tissues, 2.71±0.29 in stage T3 breast cancer tissues
and 3.30±0.18 in stage T4 breast cancer tissues. An increase in the
expression level of Snail was associated with increased tumor grade
(F=372.022, P<0.001). The results of western blotting (Fig. 5B) revealed that the relative
expression level of Snail was 1.00±0.13 in normal mammary
epithelial cells MCF-10A, 1.82±0.21 in low invasive breast cancer
cells MCF-7, 2.61±0.19 in moderately metastatic and invasive breast
cancer cells SK-BR-3 and 3.25±0.35 in highly metastatic and
invasive human breast cancer cells MDA-MB-231. Snail expression
levels were associated with increased levels of malignancy
(F=51.938, P<0.001).
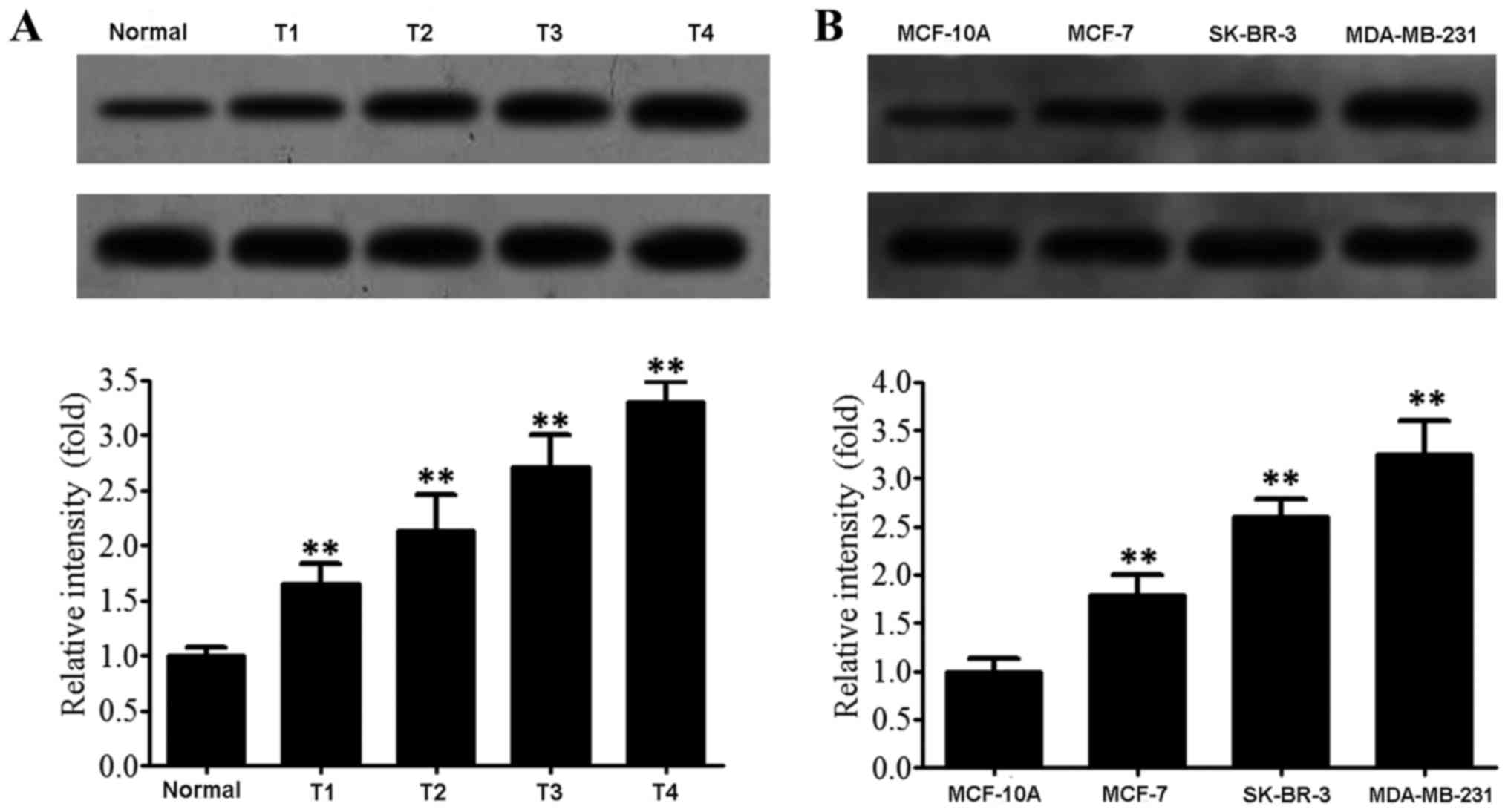 | Figure 5.Snail expression in breast cancer
tissues and cells. (A) Snail expression in breast cancer tissues:
1, paracancerous tissue; 2, T1; 3, T2; 4, T3; 5, T4; **P<0.01.
(B) Snail expression in breast cancer cells: 1, MCF-10A cells; 2,
MCF-7 cells: 3, SK-BR-3 cells; 4, MDA-MB-231 cells. **P<0.01 vs.
negative control. miR, microRNA. |
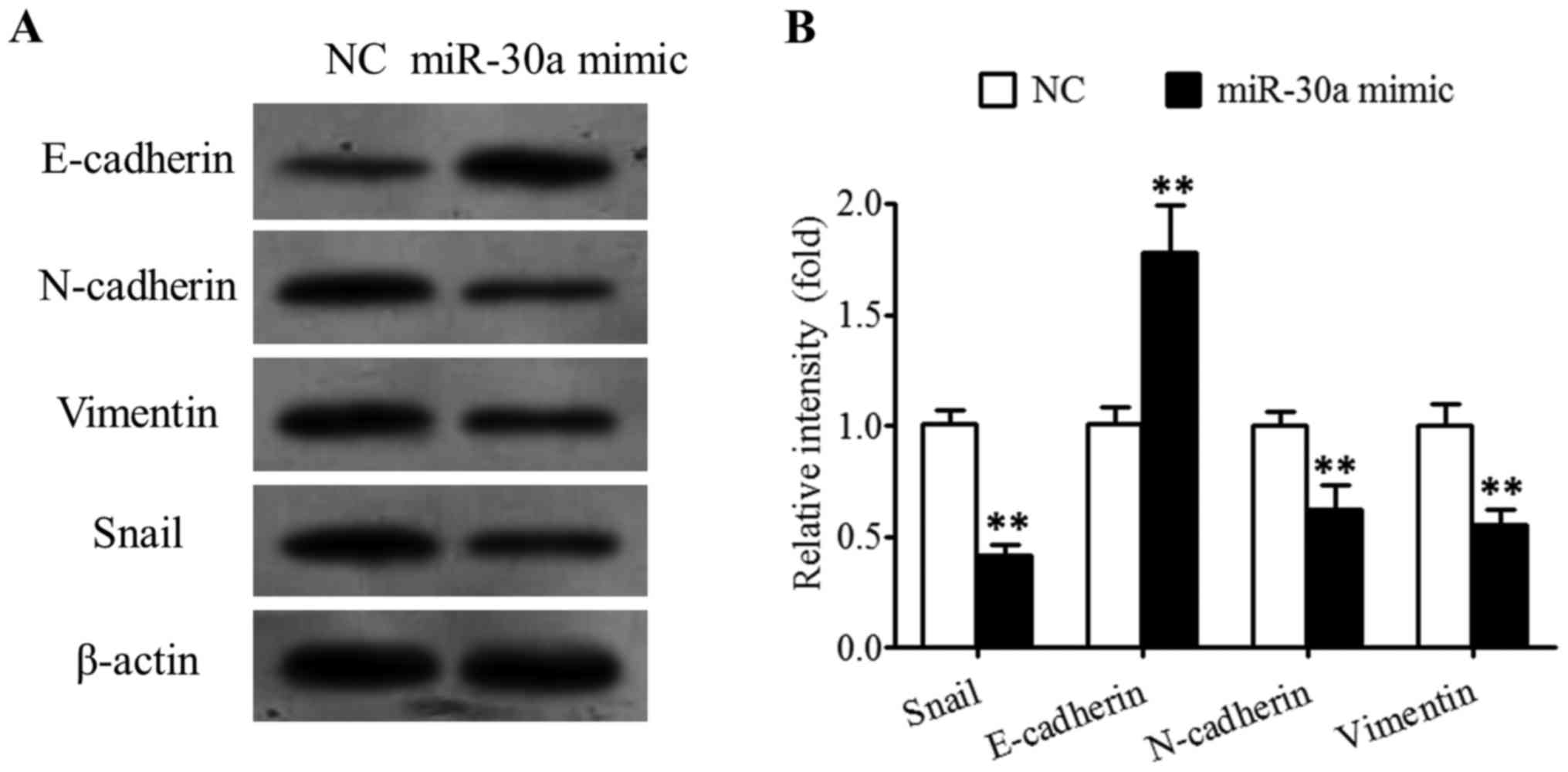
Effects of miR-30a on the expression
levels of Snail, E-cadherin, N-cadherin and Vimentin proteins in
breast cancer cells
As presented in Fig.
6, miR-30a was able to significantly inhibit Snail expression
in SK-BR-3 cells, upregulate the expression of EMT-associated
E-cadherin, and downregulate the expression of EMT-associated
N-cadherin and Vimentin.
Discussion
Previous studies revealed that a variety of miRNAs,
including miR-224, miR-21 and miR-320, were abnormally expressed in
breast cancer (20–22). The expression levels of miR-224 in
breast cancer tissues and breast cancer cell lines were decreased,
and their decreased level was associated with the degree of
malignancy of the breast cancer. miR-224 has been demonstrated to
regulate the proliferative, migratory and invasive abilities of
breast cancer cells by directly targeting fizzled 5 (20). The expression level of miR-21 in
breast cancer tissue was observed to be abnormal (23). Following overexpression of miR-21, the
proliferative and invasive abilities of breast cancer cells
significantly increased (24).
Furthermore, a study confirmed that miR-21-3p functioned primarily
by targeting multiple downstream target genes, including programed
cell death protein 4, Fas Ligand, phosphatase and tensin homolog,
Ras homolog family member B, Maspin, tissue inhibitor of
metalloproteinases 3 and reversion inducing cysteine rich protein
with kazal motifs (21). MiR-320 was
able to regulate the invasive ability of breast cancer cells by
regulating the expression of the downstream target gene metadherin
and inhibiting breast cancer (22).
In summary, a number of miRNAs may affect the occurrence,
development, invasion and metastasis of breast cancer via targeted
regulation of downstream genes. A number of studies revealed that
miR-30a has a regulatory effect in ovarian cancer, lung cancer,
liver cancer and other tumors (12–14,17,18).
Therefore, the present study analyzed the expression of miR-30a in
breast cancer and explored the potential molecular mechanism by
which it regulates the proliferation and invasion of breast cancer
cells.
The aforementioned miRNAs are closely associated
with the occurrence and development of breast cancer. The present
study focused on the role of miR-30a in the proliferation and
invasion of breast cancer, and explored the mechanism by which it
regulates the proliferation and invasion of breast cancer cells. In
the present study, miR-30a expression in breast cancer tissues and
breast cancer cells was first detected using RT-PCR. The results
revealed that with as the pathological grade and degree of
malignancy of breast cancer cells increased, the expression level
of miR-30a decreased. The effect of miR-30a on the proliferation
and invasion of breast cancer cell lines was further evaluated. The
results revealed that transfection with miR-30a was able to
significantly decrease the colony formation and invasive ability of
SK-BR-3 cells, while transfection with anti-miR-30a significantly
increased the colony formation and invasive ability of SK-BR-3
cells. This suggests that miR-30a may inhibit breast cancer by
inhibiting the proliferation and invasion of breast cancer cells.
Additional data gathered in the present study identified that
miR-30a was able to inhibit the proliferation and invasion of
MDA-MB-231 human breast cancer cells, which have high invasive and
metastatic abilities, while inhibiting miR-30a expression promoted
the proliferation and invasion of MCF-7 breast cancer cells which
have a low invasive ability. RT-PCR revealed that the expression
level of miR-30a in breast cancer tissue was significantly
decreased compared with the normal control tissues. The authors
hypothesize that the decrease in miR-30a in breast cancer tissue
may increase the proliferative and invasive abilities of breast
cancer cells, thus leading to the occurrence and development of
breast cancer.
miRNAs serve a biological function by regulating
downstream target genes. Different functions of downstream target
genes may lead to different functions of the same miRNA in
different tumor tissues (25). This
study confirmed that miR-30a had an inhibitiory effect on the
occurrence of breast cancer. In order to evaluate the downstream
target genes of miR-30a which are able to inhibit tumor development
in breast cancer tissue, the target gene of miR-30a was screened
for using the target gene prediction software TargetScans. Among
the numerous potential target genes, due to its function in the
development of human cancer (26–29), Snail
was selected as the target gene of interest. Snail is a regulatory
factor in the process of EMT in tumor cells (30). It promotes the occurrence of EMT in
tumor cells by upregulating Vimentin and downregulating the
expression of E-cadherin. Its abnormal expression is closely
associated with the invasion, metastasis and recurrence of tumor
cells (31). In the present study,
the results of a luciferase reporter gene assay confirmed that
following cotransfection with wild-type pMIR-Snail13-UTR′ vector
and miR-30a, the luciferase activity was significantly reduced,
while following cotransfection with mutant pMIR-Snail13-UTR′ vector
and miR-30a, luciferase activity did not significantly change. The
luciferase activity did not significantly change (P>0.05)
following cotransfection with wild type pMIR-Snai1 3′-UTR vector
and miR-30b. The data indicates a direct interaction between
miR-30a and Snail. Snail may be a target gene regulated by miR-30a.
Therefore, miR-30a may serve a function inhibiting breast cancer by
regulating the expression of Snail. Therefore further study was
undertaken. First, the expression of Snail in breast cancer tissues
and breast cancer cells was detected. The results revealed that
with the increase in pathological grade and malignant degree of
breast cancer cells, Snail protein expression level gradually
increased (P<0.01). The effects of miR-30a on the expression
levels of Snail, E-cadherin, N-cadherin and Vimentin proteins in
breast cancer cells were further evaluated. The results revealed
that miR-30a was able to significantly inhibit the expression of
Snail in SK-BR-3 cells, upregulate the expression of EMT-associated
E-cadherin, and downregulate the expression of EMT-associated
N-cadherin and Vimentin. These results suggest that Snail is a
downstream target gene of miR-30a, and that miR-30a may inhibit
breast cancer by regulating the expression of Snail in breast
cancer tissues.
In summary, the present study has revealed that the
expression of miR-30a in breast cancer tissue was significantly
decreased, and was closely associated with the malignant degree of
breast cancer. miR-30a is able to inhibit the proliferation and
invasion of breast cancer cells. A decrease in miR-30a expression
levels in breast cancer tissue may be associated with the
occurrence and development of breast cancer. Further study revealed
that miR-30a inhibited the proliferation and invasion of breast
cancer cells by regulating the expression of Snail, E-cadherin,
N-cadherin and Vimentin proteins.
Acknowledgements
Not applicable.
Funding
No funding received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BX designed the experiments. XS performed the
bioinformatic analyses. All other experiments were performed by JB.
The figures were prepared by XS, and the manuscript was written by
JB, with input from BX and XS.
Ethics approval and consent to
participate
The present study was approved by the ethics
committee of the No. 254 Hospital of the People's Liberation Army.
Written informed consent was obtained from all patients.
Patient consent for publication
All study participants provided consent for the
publication of data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
DeSantis C, Ma J, Bryan L and Jemal A:
Breast cancer statistics, 2013. CA Cancer J Clin. 64:52–62. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
McGuire S: World Cancer Report 2014.
Geneva, Switzerland: World Health Organization, International
agency for research on cancer, WHO Press, 2015. Adv Nutr.
7:418–419. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fan L, Strasser-Weippl K, Li JJ, St Louis
J, Finkelstein DM, Yu KD, Chen WQ, Shao ZM and Goss PE: Breast
cancer in China. Lancet Oncol. 15:e279–e289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dickens C, Joffe M, Jacobson J, Venter F,
Schüz J, Cubasch H and McCormack V: Stage at breast cancer
diagnosis and distance from diagnostic hospital in a periurban
setting: A South African public hospital case series of over 1,000
women. Int J Cancer. 135:2173–2182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Narod SA, Iqbal J, Giannakeas V, Sopik V
and Sun P: Breast cancer mortality after a diagnosis of ductal
carcinoma in situ. JAMA Oncol. 1:888–896. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xia M, Li H, Wang JJ, Zeng HJ and Wang SH:
MiR-99a suppress proliferation, migration and invasion through
regulating insulin-like growth factor 1 receptor in breast cancer.
Eur Rev Med Pharmacol Sci. 20:1755–1763. 2016.PubMed/NCBI
|
|
7
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rathod SS, Rani SB, Khan M, Muzumdar D and
Shiras A: Tumor suppressive miRNA-34a suppresses cell proliferation
and tumor growth of glioma stem cells by targeting Akt and Wnt
signaling pathways. FEBS Open Bio. 4:485–495. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li XT, Wang HZ, Wu ZW, Yang TQ, Zhao ZH,
Chen GL, Xie XS, Li B, Wei YX, Huang YL, et al: miR-494-3p
regulates cellular proliferation, invasion, migration, and
apoptosis by PTEN/AKT signaling in human glioblastoma cells. Cell
Mol Neurobiol. 35:679–687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ozen M, Creighton CJ, Ozdemir M and
Ittmann M: Widespread deregulation of microRNA expression in human
prostate cancer. Oncogene. 27:1788–1793. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu H, Wang Y, Li X, Zhang YJ, Li J, Zheng
YQ, Liu M, Song X and Li XR: Expression and regulatory function of
miRNA-182 in triple-negative breast cancer cells through its
targeting of profilin 1. Tumour Biol. 34:1713–1722. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sestito R, Cianfrocca R, Rosanò L, Tocci
P, Semprucci E, Di Castro V, Caprara V, Ferrandian G, Sacconi A,
Blandino G and Bagnato A: MiR-30a inhibits endothelin A receptor
and chemoresistance in ovarian carcinoma. Oncotarget. 7:4009–4023.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tang R, Liang L, Luo DZ, Feng ZB, Huang
QX, He RQ, Gan TQ, Yang LH and Chen G: Downregulation of MiR-30a is
associated with poor prognosis in lung cancer. Med Sci Monit.
21:2514–2520. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang W, Lin H, Zhou L, Zhu Q, Gao S, Xie
H, Liu Z, Xu Z, Wei J, Huang X and Zheng S: MicroRNA-30a-3p
inhibits tumor proliferation, invasiveness and metastasis and is
downregulated in hepatocellular carcinoma. Eur J Surg Oncol.
40:1586–1594. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang LL, Zhang XH, Zhang X and Chu JK:
MiR-30a increases cisplatin sensitivity of gastric cancer cells
through suppressing epithelial-to-mesenchymal transition (EMT). Eur
Rev Med Pharmacol Sci. 20:1733–1739. 2016.PubMed/NCBI
|
|
17
|
Liu Z, Tu K and Liu Q: Effects of
microRNA-30a on migration, invasion and prognosis of hepatocellular
carcinoma. FEBS Lett. 588:3089–3097. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu Z, Chen L, Zhang X, Xu X, Xing H,
Zhang Y, Li W, Yu H, Zeng J and Jia J: RUNX3 regulates vimentin
expression via miR-30a during epithelial-mesenchymal transition in
gastric cancer cells. J Cell Mol Med. 18:610–623. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Li SJ, Pan J, Che Y, Yin J and
Zhao Q: Specific expression of the human voltage-gated proton
channel Hv1 in highly metastatic breast cancer cells, promotes
tumor progression and metastasis. Biochem Biophys Res Commun.
412:353–359. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu F, Liu Y, Shen J, Zhang G and Han J:
MicroRNA-224 inhibits proliferation and migration of breast cancer
cells by down-regulating fizzled 5 expression. Oncotarget.
7:49130–49142. 2016.PubMed/NCBI
|
|
21
|
Kuang Y and Nie YJ: Exploration of the
regulatory effect of miR-21 on breast cancer cell line
proliferation and invasion as well as the downstream target genes.
Asian Pac J Trop Med. 9:470–473. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu J, Wang JG, Zhang L, Yang HP, Wang L,
Ding D, Chen Q, Yang WL, Ren KH, Zhou DM, et al: MicroRNA-320a
inhibits breast cancer metastasis by targeting metadherin.
Oncotarget. 7:38612–38625. 2016.PubMed/NCBI
|
|
23
|
Yan LX, Huang XF, Shao Q, Huang MY, Deng
L, Wu QL, Zeng YX and Shao JY: MicroRNA miR-21 overexpression in
human breast cancer is associated with advanced clinical stage,
lymph node metastasis and patient poor prognosis. RNA.
14:2348–2360. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yan LX, Wu QN, Zhang Y, Li YY, Liao DZ,
Hou JH, Fu J, Zeng MS, Yun JP, Wu QL, et al: Knockdown of miR-21 in
human breast cancer cell lines inhibits proliferation, in vitro
migration and in vivo tumor growth. Breast Cancer Res. 13:R22011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Crow MS, Lum KK, Sheng X, Song B and
Cristea IM: Diverse mechanisms evolved by DNA viruses to inhibit
early host defenses. Crit Rev Biochem Mol Biol. 51:452–481. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qi J, Li T, Bian H, Li F, Ju Y, Gao S, Su
J, Ren W and Qin C: SNAI1 promotes the development of HCC through
the enhancement of proliferation and inhibition of apoptosis. FEBS
Open Bio. 6:326–337. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang L, Wang YJ, Zheng LY, Jia YM, Chen
YL, Chen L, Liu DG, Li XH, Guo HY, Sun YL, et al: Genetic
polymorphisms of TGFB1, TGFBR1, SNAI1 and TWIST1 are associated
with endometrial cancer susceptibility in Chinese Han women. PLoS
One. 11:e01552702016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Haraguchi M, Sato M and Ozawa M:
CRISPR/Cas9n-mediated deletion of the Snail 1Gene (SNAI1) reveals
its role in regulating cell morphology, cell-cell interactions, and
gene expression in ovarian cancer (RMG-1) cells. PLoS One.
10:e01322602015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bai Z, Sun J, Wang X, Wang H, Pei H and
Zhang Z: MicroRNA-153 is a prognostic marker and inhibits cell
migration and invasion by targeting SNAI1 in human pancreatic
ductal adenocarcinoma. Oncol Rep. 34:595–602. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Y, Shi J, Chai K, Ying X and Zhou BP:
The role of Snail in EMT and tumorigenesis. Curr Cancer Drug
Targets. 13:963–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kaufhold S and Bonavida B: Central role of
Snail1 in the regulation of EMT and resistance in cancer: A target
for therapeutic intervention. J Exp Clin Cancer Res. 33:622014.
View Article : Google Scholar : PubMed/NCBI
|