Introduction
Bladder cancer is one of the most common malignant
tumors in urinary surgery, and the incidence rate shows a
continuously increasing trend in the world (1). In China, the morbidity and mortality
rates of bladder cancer rank first in malignant tumors of the male
reproductive system (2). Due to the
lack of early diagnostic means, lymph node metastasis has often
occurred at the diagnosis of disease. At present, surgical
treatment is mainly adopted for bladder cancer in clinical
practice, but such a method is more traumatic to patients. As a
result, the quality of life is not high, and tumor cells are prone
to metastasis (3). In addition to
surgery, conventional basic chemotherapy drugs, such as cisplatin,
are also one of the clinical treatment methods, which can achieve
satisfactory effects within a short period. Due to the longer cycle
of chemotherapy, however, the patient's body is prone to drug
tolerance, and bone marrow suppression and adverse reactions in the
nervous system occur easily (4).
Therefore, searching for an effective and safe drug is currently a
main direction in the clinical treatment of bladder cancer.
Puerarin is an isoflavone substance extracted from
plants such as Pueraria thomsonii, and plays an important
role in tumor apoptosis, proliferation and immunity (5). Studies have shown that puerarin can
induce apoptosis of human cervical cancer cells through inhibiting
Wnt/β-catenin signaling pathway (6).
Numerous studies have confirmed that puerarin has a significant
inhibitory effect on various malignant tumor cells, including
bladder cancer cells (7). However,
there is little research on puerarin in regulating bladder cancer
cell proliferation and apoptosis, and its possible molecular
mechanism remains unclear. Silent information regulator 1 (SIRT1)
is one of the members of the mammalian sirtuin family with the main
role of regulating the substance metabolism and lifespan (8). According to studies, SIRT1 can
deacetylate the lysine residue at position 382 of the tumor
suppressor protein p53, thereby reducing p53 activity and allowing
cells to bypass p53-mediated apoptosis and continue to survive
(9). However, little is known about
the role of SIRT1 in bladder cancer cells. In this study, the
regulatory effects of puerarin on apoptosis-related proteins, SIRT1
and p53 in human bladder cancer T24 cells were observed, and the
effect of puerarin on apoptosis of bladder cancer cells and its
molecular mechanism were preliminarily studied.
Materials and methods
Materials
Cell lines
Bladder cancer T24 cell lines (cat. no. FS-0139)
were purchased from the Cell Bank of the Peking Union Medical
College.
Main reagents
High-glucose Dulbecco's modified Eagle's medium
(DMEM), fetal bovine serum (FBS) and trypsin were purchased from
Dow Corning (Midland, MI, USA). Methyl thiazolyl tetrazolium (MTT)
kit and Annexin V cell apoptosis kit were purchased from Nanjing
KeyGen Biotech Co., Ltd. (Nanjing, China). M-MuLV Reverse
Transcriptase kit and SYBR-Green I real-time fluorescence
quantitative polymerase chain reaction (PCR) kit were purchased
from Thermo Fisher Scientific, Inc. (Waltham, MA, USA). SIRT1
agonist SRT1720 and inhibitor EX527 were from Sigma-Aldrich
(Sigma-Aldrich: Merck KGaA, St. Louis, MO, USA) [dissolved at 1
mmol/l with dimethylsulfoxide (DMSO), and stored at −20°C in the
dark]. Primary mouse anti-human SIRT1, p53, B-cell lymphoma 2
(Bcl-2), Bcl-2-associated X protein (Bax) and β-actin monoclonal
antibodies, and horseradish peroxidase-labeled secondary goat
anti-mouse polyclonal antibody (cat. nos. sc-135792, sc-47698,
sc-509,sc-20067, sc-58673 and sc-2005, were purchased from Santa
Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). SIRT1, p53 and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene primer
sequences were synthesized by Nanjing KeyGen Biotech Co., Ltd.
The study was approved by the Ethics Committee of
the Second Hospital of Shandong University (Jinan, China). Informed
consents were signed by the patients and/or guardians.
Main methods
Cell culture and grouping
Human bladder cancer T24 cell lines were cultured in
high-glucose DMEM containing 10% FBS. After meeting the
experimental conditions, cells in the logarithmic growth phase were
taken for experiments. After centrifugation at 860 × g for 5 min at
4°C, cell sediment was taken, and cells were resuspended and
inoculated into culture plates in different specifications
according to the experimental requirements. After cells fully
adhered to the wall, puerarin in a given concentration (0–200
µmol/l), SIRT1 agonist SRT1720 (1 µmol/l) or SIRT1 inhibitor EX527
(1 µmol/l) was added for routine culture under 5% CO2 at
37°C for a predetermined time, followed by subsequent
experiment.
Detection of T24-cell proliferation
via MTT assay
Experimental subjects were divided into blank
control, control and experimental groups. Human bladder cancer T24
cells were inoculated into a 96-well plate, and the cell density
was controlled at 5×103 cells/well, and six repeated
wells were set in each group. After cells adhered to the wall, 1%
bovine serum albumin containing no puerarin (control group) and
containing 20, 50, 100 and 200 µmol/l puerarin (experimental
groups) were added, respectively, for incubation in an incubator
with 5% CO2 at 37°C for 24, 48 and 72 h. Then, 200 µl
DMSO was added into each well and vibrated at a low speed on a
shaking table for 10 min to completely dissolve the crystals. The
protocol was repeated 3 times. The absorbance value of each well
was measured at a wavelength of 490 nm by using a microplate reader
(Bio-Rad 680; Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
blank control group (without cells and puerarin) was used for zero
setting. Cell inhibition rate = 1 - (absorbance
valueexperimental group - absorbance valueblank
control group)/(absorbance valuecontrol group -
absorbance valueblank control group) ×100%.
Detection of T24-cell apoptosis via
flow cytometry
T24 cells in the logarithmic growth phase were
inoculated into the petri dish, and treated with puerarin (100
µmol/l), SRT1720 (1 µmol/l) and EX527 (1 µmol/l) for 24 h according
to the experimental requirements. After digestion with trypsin,
adherent cells were collected, counted and washed with
phosphate-buffered saline (PBS) twice. Then, the cells in each
group were resuspended in 500 µl binding buffer, and 5 µl Annexin V
and 5 µl propidium iodide were added, followed by incubation in the
dark at room temperature for 15 min. Finally, the apoptosis rate
was detected via flow cytometry (Thermo Fisher Scientific,
Inc.).
Detection of protein expression levels
via western blotting
T24 cells in the logarithmic growth phase were
inoculated into the petri dish, and treated with puerarin (100
µmol/l), SRT1720 (1 µmol/l) and EX527 (1 µmol/l) for 24 h according
to the experimental requirements. Cells were added with the
radioimmunoprecipitation assay (RIPA) lysis buffer at 4°C and
incubated for 30 min. The total protein was extracted, and the
protein concentration was detected by using the bicinchoninic acid
(BCA) kit, and the protein curve was drawn. Then, 60 µg total
protein was taken for sample spotting, and isolated via 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE),
after which the protein was transferred onto a polyvinylidene
fluoride (PVDF) membrane under 100 V for 150 min, blocked with 5%
skim milk powder at room temperature for 1 h, and incubated with
primary antibody (1:500) at 4°C overnight, with β-actin (1:600) as
the internal reference. After being washed with PBS, the protein
was added with the secondary antibody (1:800) for incubation at
37°C for 2 h. Finally, the band image was collected after
electrochemiluminescence development (Bio-Rad Laboratories, Inc.)
and the optical density value of protein band was measured by using
the gel imaging analysis system (Bio-Rad Laboratories, Inc.). The
gray value of the target protein band was scanned by Quantity One
software, and the expression intensity of each group was indicated
by the ratio of the gray value of the target protein band to
β-actin band.
Detection of messenger ribonucleic
acid (mRNA) expression via reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
T24 cells in the logarithmic growth phase were taken
and treated with puerarin (100 µmol/l), SRT1720 (1 µmol/l) and
EX527 (1 µmol/l) for 24 h according to the experimental
requirements. RNA was extracted from cells by using TRIzol reagent
via one-step method, reverse transcribed into complementary
deoxyribonucleic acid (cDNA) and then amplified into DNA in strict
accordance with the manufacturers instructions of the reverse
transcription and SYBR-Green I real-time fluorescence quantitative
polymerase chain reaction (PCR) kit, followed by quantitative
analysis of the target gene. Primer sequences of SIRT1: forward,
5′-CTGCCTGGATCCCCTTAGTTTTG-3′ and reverse,
5′-GGGCCTGTTGCTCTCCTCATTAA-3′. Primer sequences of p53: forward,
5′-CCGGCGCACAGAGGAAGAGA-3′ and reverse,
5′-TGGGGAGAGGAGCTGGTGTTGT-3′. Primer sequences of GAPDH as an
internal reference: forward, 5′-CATGGGGTGTGAACCATGAGA-3′ and
reverse, 5′-GTCTTCTGGGTGGCAGTGAT-3′. The 2−ΔΔCq method
was used for quantification (10).
The thermocycling conditions were as follows: Predenaturation at
95°C for 10 min, then denaturation at 95°C for 30 sec, annealing at
50°C for 30 sec, at 95°C, extension at 70°C for 1 min, 40 cycles,
and finally extending 10 min at 70 °C. The experiment was repeated
3 times.
Statistical analysis
Statistical Product and Service Solutions (SPSS)
22.0 software (IBM Corp., Armonk, NY, USA) was used for data entry,
and data were collected and statistically analyzed. t-test was used
for the comparison of measurement data between the two groups, and
one-way analysis of variance was used for the data comparison among
groups and the post hoc test was the Least Significant Difference
test. P<0.05 indicated that the difference was statistically
significant.
Results
Inhibitory effect of puerarin on
proliferation of bladder cancer T24 cells (Fig. 1)
Results of MTT assay revealed that the inhibition
rate of bladder cancer T24 cells was significantly increased after
treatment with puerarin at different concentrations (0, 20, 50, 100
and 200 µg/ml), and it was elevated with the increase of treatment
time and dose, displaying an obvious dose-effect relationship. A
total of 20 and 50 µg/ml puerarin could inhibit the proliferation
of T24 cells, and there was no statistically significant difference
compared with the normal control group (p>0.05). The cell
inhibition rates under 100 µg/ml puerarin at 24, 48 and 72 h had
statistically significant differences compared with those in the
normal control group (p<0.05). Besides, there were also
statistically significant differences in the cell inhibition rates
under 200 µg/ml puerarin at 24, 48 and 72 h compared with those in
the normal control group (p<0.01).
Puerarin induces apoptosis of bladder
cancer T24 cells
Results of flow cytometry manifested that compared
with control group, 100 µg/ml puerarin could significantly increase
the proportion of apoptosis in bladder cancer T24 cells
(p<0.05). However, compared with that in the puerarin group, the
proportion of apoptotic T24 cells was significantly decreased after
pre-treatment with SIRT1 agonist SRT1720 for 1 h and then treatment
with 100 µg/ml puerarin for 24 h (p<0.05). Compared with the
control group, the administration of SRT1720 alone did not increase
the proportion of apoptotic T24 cells, and the difference was not
statistically significant (p>0.05). However, the administration
of SIRT1 inhibitor EX527 alone for 24 h could simulate the effect
of puerarin to induce T24-cell apoptosis (p<0.05) (Fig. 2A and B). According to the results of
western blotting, compared with the control group, 100 µg/ml
puerarin could obviously increase the Bax protein expression and
decrease the Bcl-2 protein expression in T24 cells, so the
Bax/Bcl-2 ratio was increased significantly (p<0.05). Moreover,
compared with those in the puerarin group, expression of the Bax
and Bcl-2 protein in T24 cells were reversed, and the Bax/Bcl-2
ratio was decreased after administration of SIRT1 agonist SRT1720
(p<0.05). Compared with those in the control group, the Bax
protein expression was not increased and the Bcl-2 protein
expression was not decreased after administration of SRT1720 alone,
and there were no statistically significant differences
(p>0.05). However, the administration of SIRT1 inhibitor EX527
alone for 24 h could simulate the effect of puerarin to increase
the Bax protein expression and Bax/Bcl-2 ratio and decrease the
Bcl-2 protein expression (p<0.05) (Fig. 2C-F).
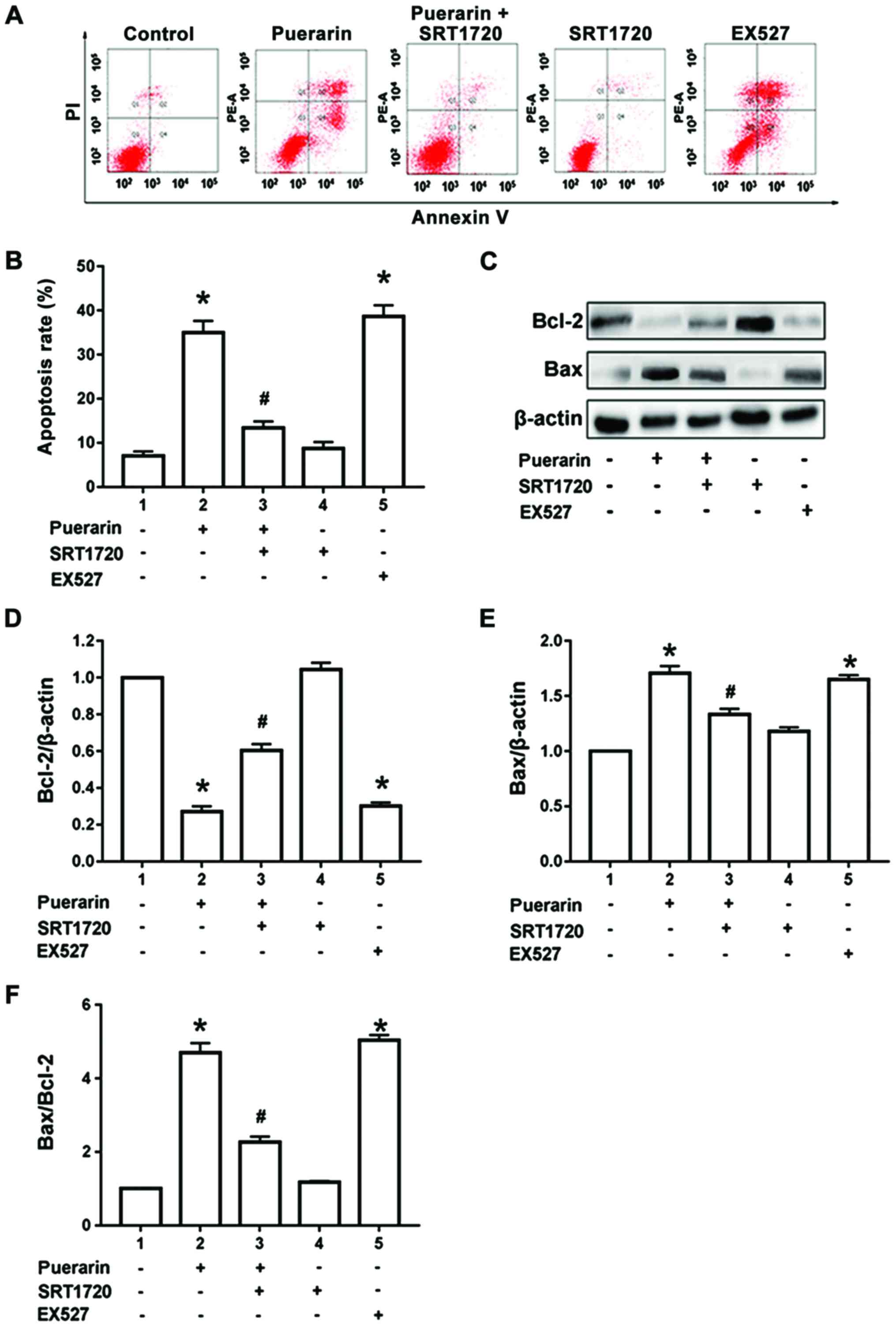 | Figure 2.Puerarin induces apoptosis of bladder
cancer T24 cells. (A and B) Flow cytometry was conducted to
determine apoptosis of T24 cells treated with puerarin, SRT1720,
EX527 and puerarin combined with SRT1720. (C-E) The protein levels
of Bax, Bcl-2 were determined by western blotting in T24 cells
treated with puerarin, SRT1720, EX527 and puerarin combined with
SRT1720. (F) The Bax/Bcl-2 ratio in T24 cells treated with
puerarin, SRT1720, EX527 and puerarin combined with SRT1720.
*P<0.05 vs. control group, #P<0.01 vs. puerarin
group. Bax, Bcl-2-associated X protein; Bcl-2, B-cell lymphoma
2. |
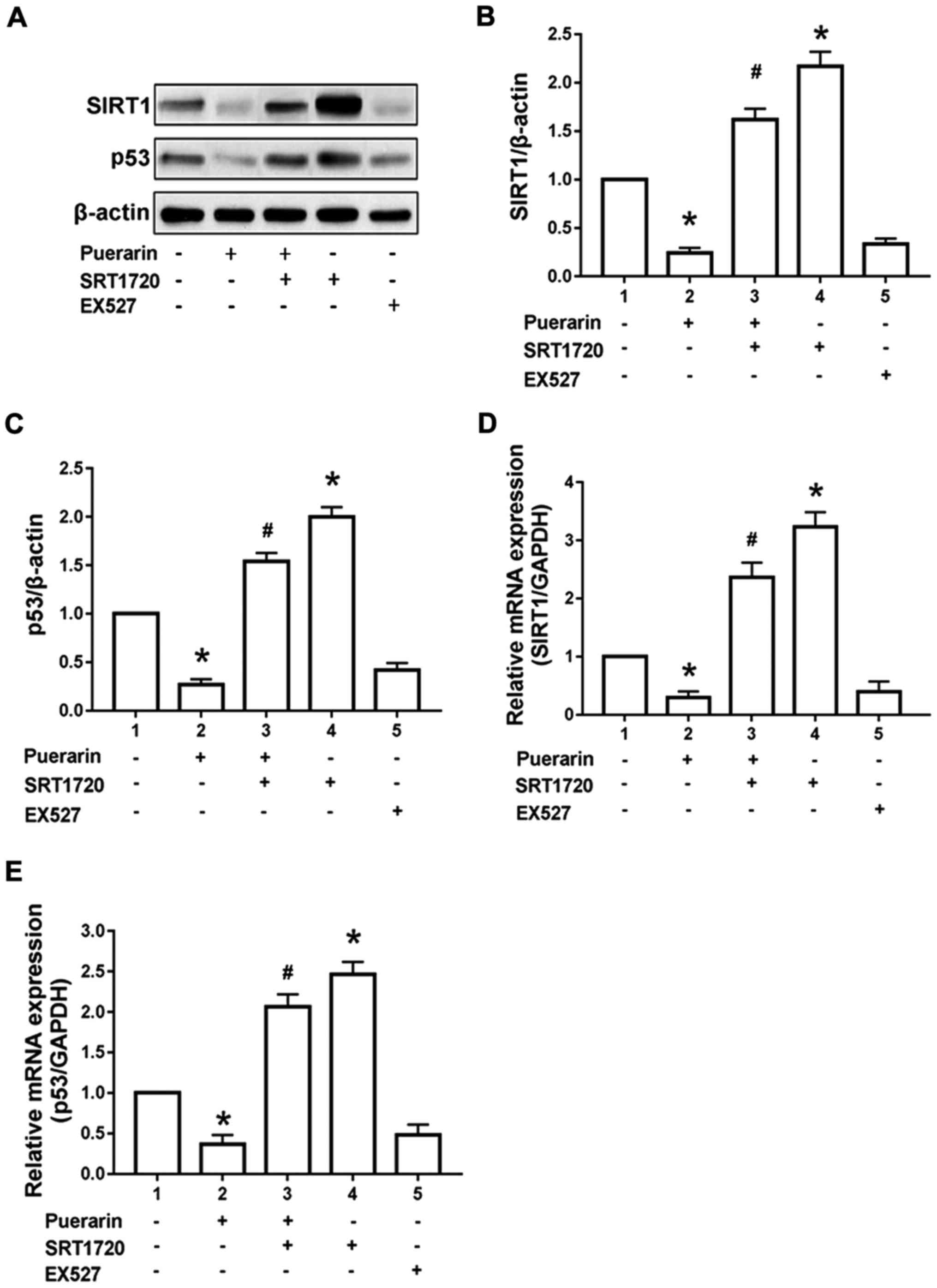
Effect of puerarin on SIRT1/p53
pathway in bladder cancer T24 cells
Results of western blotting manifested that compared
with control group, 100 µg/ml puerarin could remarkably reduce
expression of SIRT1 and p53 protein in T24 cells (p<0.05).
Compared with those in the puerarin group, expression of SIRT1 and
p53 protein in T24 cells could be reversed after administration of
SIRT1 agonist SRT1720. The admini-stration of SRT1720 alone could
increase the SIRT1 and p53 protein expression compared with the
control group (p<0.05). However, the administration of SIRT1
inhibitor EX5272 alone for 24 h simulated the effect of puerarin to
reduce the SIRT1 and p53 protein in T24 cells (p<0.05) (Fig. 3A-C). According to the results of
RT-qPCR, 100 µg/ml puerarin could obviously reduce the expression
of SIRT1 and p53 mRNA in T24 cells compared with the control group
(p<0.05). Compared with those in the puerarin group, the
expression of SIRT1 and p53 mRNA in T24 cells was reversed after
administration of SIRT1 agonist SRT1720. The administration of
SRT1720 alone could increase the SIRT1 and p53 mRNA expression
compared with the control group (p<0.05). However, the
administration of SIRT1 inhibitor EX5272 alone for 24 h could
simulate the effect of puerarin to reduce the expression of SIRT1
and p53 mRNA in T24 cells (p<0.05) (Fig. 3D and E).
Discussion
Bladder cancer is one of the frequently occurring
tumors in men, as well as an important cause of cancer death in
men. Studies have demonstrated that heavy smoking and drinking will
significantly increase the onset risk of bladder cancer, which are
not conducive to the prognosis and improvement of life quality of
patients (11). However, due to the
high recurrence and metastasis rates of bladder cancer, its
clinical treatment is often restricted, and there is still lack of
reliable and effective treatment means. Therefore, it is of great
importance to find a new treatment method.
There is considerable research showing that
traditional Chinese medicine can play a regulatory role in disease
at a macro-level, and possesses excellent curative effects on
delaying, controlling and preventing the recurrence of disease
(12). Puerarin is an isoflavone
substance extracted from plants such as Pueraria thomsonii.
Numerous studies have revealed that puerarin significantly inhibits
the proliferation of a variety of breast cancer cells in a time-
and dose-dependent manner (13,14).
Besides, puerarin can induce apoptosis through the classic caspase
apoptosis pathway, and arrest cells in the G2/M phase (15). Results of this study manifested that
the proliferation capacity of bladder cancer T24 cells remarkably
declined after treatment with puerarin at different concentrations
(0, 20, 50, 100 and 200 µg/ml) in a time- and dose-dependent
manner, which were consistent with the above views.
Blocked apoptosis is one of the causes of the
infinite proliferation of tumor cells. Bcl-2 family genes and p53
genes are all important in regulating apoptosis. Bcl-2 and Bax, as
the two most representative genes in the Bcl-2 family, exert
effects of inhibiting and promoting apoptosis, respectively. When
Bcl-2 is relatively predominant in cells, Bcl-2 will form a
heterodimer with Bax, thus inhibiting apoptosis. However, when Bax
is predominant in cells, Bax itself will form the homodimer, thus
promoting apoptosis (16,17). Therefore, the Bax/Bcl-2 ratio has
important significance for the occurrence and degree of apoptosis.
In this study, it was found that 100 µg/ml puerarin could
significantly inhibit the proliferation of bladder cancer T24cells,
increase the Bax protein expression, and reduce the Bcl-2 protein
expression, so the Bax/Bcl-2 ratio was obviously increased, thereby
inducing apoptosis. After administration of SIRT1 agonist SRT1720,
the expression of Bax and Bcl-2 protein in T24 cells was reversed,
and the Bax/Bcl-2 ratio was decreased. Besides, the administration
of SIRT1 inhibitor EX5272 alone for 24 h simulated the effect of
puerarin to increase the Bax protein expression and the Bax/Bcl-2
ratio, and decrease the Bcl-2 protein expression, suggesting that
puerarin can be used as a novel drug in the treatment of bladder
cancer. However, the specific mechanism of puerarin in inhibiting
proliferation and inducing apoptosis of bladder cancer cells has
not been clarified yet, and it is pending further study.
In recent years, the major role of SIRT1 gene in the
occurrence and development of tumors has attracted extensive
attention. SIRT1 is an important member of the nicotinamide
(NAD+)-dependent class III histone deacetylase sirtuin
family (18). Previous studies found
that SIRT1 plays a crucial role in regulating the senescence and
substance metabolism of tumor cells (19). Some studies have manifested that the
low expression of SIRT1 can induce apoptosis of ovarian cancer
cells (20,21), but little is known about SIRT1 in
bladder cancer cells. In addition, studies have shown that SIRT1
can downregulate the p53 expression through inhibiting acetylation
of p53 protein, thus promoting cell growth (22). In this study, it was found that SIRT1
and p53 protein and mRNA expression in bladder cancer T24 cells
could be obviously reduced by 100 µg/ml puerarin, but they could be
significantly increased after pre-treatment of T24 cells with
SRT1720 for 1 h. Administration of EX527 alone in the treatment of
T24 cells could stimulate the effect of puerarin to reduce
expression of SIRT1 and p53 protein and mRNA. Therefore, puerarin
may induce apoptosis of bladder cancer cells via inhibiting
SIRT1/p53 pathway, but its specific regulatory mechanism remains
unclear.
In conclusion, this experiment suggested that
puerarin can inhibit proliferation and induce apoptosis of bladder
cancer T24 cells, and proved that SIRT1/p53 signaling pathway is
involved in the pathological process of apoptosis of human bladder
cancer T24 cells, laying a foundation for the application of
puerarin in prevention and treatment of bladder cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
GY designed the study and drafted the manuscript. SK
collected and analyzed the data. JC performed RT-qPCR. XL was
responsible for western blotting. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the Second Hospital of Shandong University (Jinan, China). Informed
consents were signed by the patients and/or guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mowlds DS, Foster CE and Ichii H: Invasive
squamous cell bladder cancer of the ureterovesical junction in a
renal transplant patient: A case report. J Surg Case Rep.
2017:rjx0662017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wood DP: Re: Current clinical practice
gaps in the treatment of intermediate- and high-risk
non-muscle-invasive bladder cancer (NMIBC) with emphasis on the use
of Bacillus Calmette-Guérin (BCG): results of an international
individual patient data survey (IPDS). J Urol. 191:17312014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zahoor H, Elson P, Stephenson A, Haber GP,
Kaouk J, Fergany A, Lee B, Koshkin V, Ornstein M, Gilligan T, et
al: Patient characteristics, treatment patterns and prognostic
factors in squamous cell bladder cancer. Clin Genitourin Cancer.
16:e437–e442. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arruda RP, Mariano MB, Pereira CF, Lima
GC, Lessa TN and Neto MC: Laparoscopic cystoprostatectomy for
bladder cancer in a male patient combined with open ileal conduit
urinary diversion. Int Braz J Urol. 43:169–170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jiang K, Chen H, Tang K, Guan W, Zhou H,
Guo X, Chen Z, Ye Z and Xu H: Puerarin inhibits bladder cancer cell
proliferation through the mTOR/p70S6K signaling pathway. Oncol
Lett. 15:167–174. 2018.PubMed/NCBI
|
|
6
|
Wang PP, Zhu XF, Yang L, Liang H, Feng SW
and Zhang RH: Puerarin stimulates osteoblasts differentiation and
bone formation through estrogen receptor, p38 MAPK, and
Wnt/β-catenin pathways. J Asian Nat Prod Res. 14:897–905. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo XF, Yang ZR, Wang J, Lei XF, Lv XG and
Dong WG: Synergistic antitumor effect of puerarin combined with
5-fluorouracil on gastric carcinoma. Mol Med Rep. 11:2562–2568.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin MH, Lee YH, Cheng HL, Chen HY, Jhuang
FH and Chueh PJ: Capsaicin inhibits multiple bladder cancer cell
phenotypes by inhibiting tumor-associated NADH oxidase (tNOX) and
sirtuin1 (SIRT1). Molecules. 21:8492016. View Article : Google Scholar
|
|
9
|
Vaziri H, Dessain SK, Ng Eaton E, Imai SI,
Frye RA, Pandita TK, Guarente L and Weinberg RA: hSIR2(SIRT1)
functions as an NAD-dependent p53 deacetylase. Cell. 107:149–159.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using realtime quantitative PCR and
the 2(Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Reulen RC, de Vogel S, Zhong W, Zhong Z,
Xie LP, Hu Z, Deng Y, Yang K, Liang Y, Zeng X, et al: Physical
activity and risk of prostate and bladder cancer in China: The
South and East China case-control study on prostate and bladder
cancer. PLoS One. 12:e01786132017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang JL, Liu BY and Ma KW: Traditional
Chinese medicine. Lancet. 372:1938–1940. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen T, Chen H, Wang Y and Zhang J: In
vitro and in vivo antitumour activities of puerarin 6″-O-xyloside
on human lung carcinoma A549 cell line via the induction of the
mitochondria-mediated apoptosis pathway. Pharm Biol. 54:1793–1799.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu X, Zhao W, Wang W, Lin S and Yang L:
Puerarin suppresses LPS-induced breast cancer cell migration,
invasion and adhesion by blockage of NF-κB and Erk pathway. Biomed
Pharmacother. 92:429–436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu Z and Li W: Induction of apoptosis by
puerarin in colon cancer HT-29 cells. Cancer Lett. 238:53–60. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Al-Qathama A, Gibbons S and Prieto JM:
Differential modulation of Bax/Bcl-2 ratio and onset of caspase-3/7
activation induced by derivatives of justicidin B in human melanoma
cells A375. Oncotarget. 8:95999–96012. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang S, Qin F, Yang L, Xian J, Zou Q, Jin
H, Wang L and Zhang L: Nucleophosmin mutations induce
chemosensitivity in THP-1 leukemia cells by suppressing NF-κB
activity and regulating Bax/Bcl-2 expression. J Cancer.
7:2270–2279. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ruan L, Wang L, Wang X, He M and Yao X:
SIRT1 contributes to neuroendocrine differentiation of prostate
cancer. Oncotarget. 9:2002–2016. 2017.PubMed/NCBI
|
|
19
|
Kang YY, Sun FL, Zhang Y and Wang Z: SIRT1
acts as a potential tumor suppressor in oral squamous cell
carcinoma. J Chin Med Assoc. 81:416–422. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shuang T, Wang M, Zhou Y and Shi C:
Over-expression of Sirt1 contributes to chemoresistance and
indicates poor prognosis in serous epithelial ovarian cancer (EOC).
Med Oncol. 32:2602015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang X, Chen J, Sun L and Xu Y: SIRT1
deacetylates KLF4 to activate Claudin-5 transcription in ovarian
cancer cells. J Cell Biochem. 119:2418–2426. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Han L, Liang XH, Chen LX, Bao SM and Yan
ZQ: SIRT1 is highly expressed in brain metastasis tissues of
non-small cell lung cancer (NSCLC) and in positive regulation of
NSCLC cell migration. Int J Clin Exp Pathol. 6:2357–2365.
2013.PubMed/NCBI
|