Introduction
Ovarian cancer is the leading cause of
cancer-associated mortality among all gynecological malignancies in
China. Due to the fact that there are few specific or sensitive
biomarkers for ovarian cancer monitoring, the majority of patients
with ovarian cancer are diagnosed at late stages (1,2). The
therapeutic guidelines for patients with ovarian cancer included
aggressive surgical resection followed by adjuvant chemotherapy
(3). Platinum-based chemotherapy has
been the first-line chemotherapeutic treatment for patients with
ovarian cancer. Despite recent improvements in the survival rate of
patients with ovarian cancer, >80% of patients eventually
relapse due in part to the acquisition of chemoresistance (4). Therefore, it is urgent to identify novel
targets for the development of therapeutics for patients with
ovarian cancer.
Several studies have attempted to uncovered the
mechanisms of cisplatin resistance in various human carcinoma cell
lines. Cancerous inhibitor of protein phosphatase 2A (CIP2A), a
novel oncoprotein, is also known as KIAA1524 or p90
tumor-associated antigen (5,6). Overexpression of CIP2A contributed to
cell proliferation and progression of malignancies (7,8). Previous
studies have revealed that CIP2A enhanced the proliferation and
invasion of ovarian cancer cells. A recent study reported that
CIP2A has also been implicated in resistance to chemotherapy
(9). However, it remains unclear
whether CIP2A serves a critical role in cisplatin resistance in
ovarian cancer. Therefore, we hypothesized that CIP2A is associated
with chemoresistance in ovarian cancer. To the best of our
knowledge, the present study was the first to demonstrate that
CIP2A leads to cisplatin resistance in ovarian cancer and that
inhibition of CIP2A may be a promising treatment for patients with
ovarian cancer.
Materials and methods
Patients and specimens
Paraffin-embedded tissue blocks were collected from
36 patients with a median age of 56 years, age range 38–72 years)
with epithelial ovarian cancer at the Affiliated Hospital of Jining
Medical University between January 2008 and December 2014. Paired
adjacent non-cancerous tissues were also taken from the distal
resection margins (>5 cm). None of the patients had undergone
chemotherapy, immunotherapy or radiotherapy prior to specimen
resection. The clinicopathological characteristics of patients were
recorded, including age, clinical stage and tumor differentiation.
Additionally, half the samples available from 36 patients were
snap-frozen in liquid nitrogen immediately following resection and
were stored at −80°C until subsequent analysis. The present study
was approved by the Local Institutional Review Board of the
Affiliated Hospital of Jining Medical University, and written
informed consent was obtained from all patients.
Cell lines
The cisplatin-resistant ovarian carcinoma
SKOV-3CDDP/R cell line and the cisplatin-sensitive SKOV-3 cell line
were purchased from the Cell Bank of the Chinese Academy of
Sciences Institute (Shanghai, China). The two cell lines were
cultured in RPMI-1640 medium (Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA), supplemented with 10% (v/v) fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100
U/ml penicillin and 100 g/ml streptomycin, and cells were
maintained at 37°C in a humidified atmosphere containing 5%
CO2 and 95% air. Cell culture medium was changed every 4
days depending on cell density. For routine passage, when cells
reached 85–90% confluency, they were split at a ratio of 1:4.
Immunohistochemistry (IHC)
For IHC, 4 µm, 10% formalin-fixed (4°C for ~24 h),
paraffin-embedded tissue sections were deparaffinized with xylene,
and rehydrated through a series of decreasing concentrations of
ethanol (100, 95, 90, 80 and 70%). A high-temperature antigen
retrieval method was performed using a citrate buffer solution
(Maixin Bio, Fujian, China), and the slides were immersed in 100 µl
3% hydrogen peroxide for 10 min at 37°C to block endogenous
peroxidase activity. Subsequent to washing with PBS, the sections
were incubated with 5% bovine serum albumin (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) at room temperature for 30 min, followed
by incubation with anti-CIP2A monoclonal antibody (dilution, 1:400;
cat. no. NB110-59722; Novus Biologicals, LLC, Littleton, CO, USA)
at 4°C overnight. Following washing with PBS, the sections were
incubated with a horseradish peroxidase-conjugated rabbit
anti-mouse IgG secondary antibody (1:5,000; cat. no. ab6728; Abcam)
for 30 min at 37°C.
The slides were stained with diaminobenzidine (as
the color reagent) for 5 min at 37°C and hematoxylin (as a
counterstain for the nuclei) for 2 min at 37°C. PBS was used as a
negative control for the staining reactions. All slides were
dehydrated in 70% ethanol (5 min), 75% ethanol (5 min), 80% ethanol
(5 min), 90% ethanol (5 min), 95% ethanol (5 min) and 100% ethanol
(10 min). Finally, all sections were mounted with neutral gum. IHC
scores (between 0 and 8) were calculated by adding the intensity
score (0–4) and the percentage score (0–4), with a maximum score of
8. A score of ≥4 was classified as high expression of CIP2A, while
a score of ≤3 was classified as low expression of CIP2A. The
Olympus CKX41SF (Olympus Corporation, Tokyo, Japan) light
microscope was used at a magnification of ×200.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from frozen tissues and cell
lines using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) for 30 min at room temperature. Complementary DNA
(cDNA) was synthesized using a SuperScript VILO cDNA Synthesis kit
(Invitrogen; Thermo Fisher Scientific, Inc.). The reaction was
incubated in a MyCycler Thermocycler (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) for 10 min at 25°C, 60 min at 42°C and 5 min at
85°C. cDNA samples were stored at −20°C prior to RT-qPCR
amplification. RT-qPCR was performed using the SYBRGreen PCR master
mix (Applied Biosystems; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) in a total volume of 20 µl on a 7900HT fast real-time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.) as
follows: 95°C for 30 sec, 40 cycles at 95°C for 5 sec, 60°C for 30
sec. A dissociation step was performed to generate a melting curve
to confirm the specificity of the amplification. β-actin was used
as the reference gene. The relative levels of gene expression were
represented as ΔCq=Cq gene-Cq reference, and the fold change of
gene expression was calculated using the 2−ΔΔCq method
(10). Experiments were repeated in
triplicate. The primer sequences used were as follows: CIP2A
forward, 5′-ATACTTCAGGACCCACGTTTGAT-3′, CIP2A reverse,
5′-TCTCCAAGTACTAAAGCAGGAAAATCT-3′; β-actin forward,
5′-ATAGCACAGCCTGGATAGCAACGTAC-3′, β-actin reverse,
5′-CACCTTCTACAATGAGCTGCGTGTG-3′.
Western blot analysis
The SKOV-3 and SKOV-3CDDP/R ovarian carcinoma cells
were quickly washed twice with pre-cooled PBS quickly and suspended
in radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Haimen, China). Proteins were quantified using a
Bradford assay. Subsequently, 50 mg total protein extracts were
fractionated by 10% SDS-PAGE, prior to being transferred onto
polyvinylidene difluoride membranes (GE Healthcare Life Sciences,
Little Chalfont, UK). The membranes were blocked with 5% milk at
room temperature for 2 h. The membranes were subsequently incubated
with the following primary antibodies: Mouse anti-CIP2A (dilution,
1:2,000; No: NB110-59722; Novus Biologicals, LLC, Littleton, CO,
USA), mouse anti-β-actin (dilution, 1:2,000, cat no. ab8226;
Abcam), rabbit anti-human p-Akt (Ser 473; dilution, 1:200; cat no.
sc-7985-R, Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and
rabbit anti-human Akt antibody (dilution, 1:1,000; cat no: 4691,
Cell Signaling Technology, Inc.) for 12 h at 4°C. Membranes were
then washed three times with TBS-T and were incubated with
horseradish peroxidase-conjugated secondary antibodies (goat
anti-rabbit IgG, 1:2,000; cat. no. sc-2030, Santa Cruz
Biotechnology, Inc., Dallas, TX, USA; goat anti-mouse IgG-B,
1:2,000; cat. no. sc-2039, Santa Cruz Biotechnology, Inc., Dallas,
TX, USA) for 1 h at room temperature. Membranes were then washed
again three times for 10 min each time with TBS-T. Target protein
bands were visualized using the enhanced chemiluminescence method
(Pierce; Thermo Fisher Scientific, Inc.). The intensity of the
bands was quantified using the Tanon GIS system (Tanon 2500R;
Quantity One software; Tanon Science and Technology Co., Ltd.,
Shanghai, China) and the data were normalized to the β-actin
loading controls. All western immunoblot analyses were performed
three times.
Small interfering RNA (siRNA) and
plasmid transfection and co-transfection
siRNA duplexes were prepared by Guangzhou RiboBio
Co., Ltd. (Guangzhou, China). The sequences of the siRNAs were as
follows: CIP2A siRNA (siCIP2A)1, 5′-CUGUGGUUGUGUUUGCACUTT-3′;
siCIP2A2, 5′-ACCAUUGAUAUCCUUAGAATT-3′; and negative control (NC)
siRNA, 5′-UAACAAUGAGAGCACGGCTT-3′. Transfection or co-transfection
with siRNA and plasmids were performed using Lipofectamine reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol.
Cell viability assay
Cell viability was measured using a Cell Counting
kit-8 assay (CCK8; Dojindo Molecular Technologies, Inc., Shanghai,
China). In brief, cells were plated at a density of
5×103 cells/well onto 96-well plates and were
transfected with the control and siCIP2A for 48 h. Following 48 h
incubation at 37°C in a humidified atmosphere containing 5%
CO2, the samples were incubated for another 2 h with
CCK8 reagent. The absorbance was measured at 450 nm using an FLx800
Fluorescence Microplate Reader (Biotek Instruments, Inc., Winooski,
VT, USA).
Annexin V assay of apoptosis
At 48 h after siRNA transfection, the cells were
exposed to cisplatin (1.5 mg/ml) for 24 h. Next, cells were
harvested and washed twice with ice-cold PBS, suspended in Annexin
V-binding buffer (BD Pharmingen; BD Biosciences, Franklin Lakes,
NJ, USA), and propidium iodide and Annexin V-FITC (BD Pharmingen;
BD Biosciences, Franklin Lakes, NJ, USA) was added. Following
incubation for 20 min at room temperature in the dark, fluorescence
was measured on a flow cytometer and analyzed with FlowJo 7.6 (BD
Biosciences, Franklin Lakes, NJ, USA).
Statistical analysis
Data analyses were performed using SPSS statistical
package 15.0 (SPSS Inc., Chicago, IL, USA). Patient characteristics
are expressed as the mean ± standard deviation for continuous
variables, and as the count and percentage for discrete variables.
All experiments were performed in triplicate. Comparisons between
two groups were performed using Student's t-test, while comparisons
among ≥3 groups were performed using one-way analysis of variance
and post-hoc Student-Newman-Keuls test. P<0.05 was considered to
indicate a statistically significant difference.
Results
CIP2A is upregulated in ovarian cancer
tissues
In order to investigate the role of CIP2A in ovarian
cancer, the expression of CIP2A in 36 ovarian cancer tissue
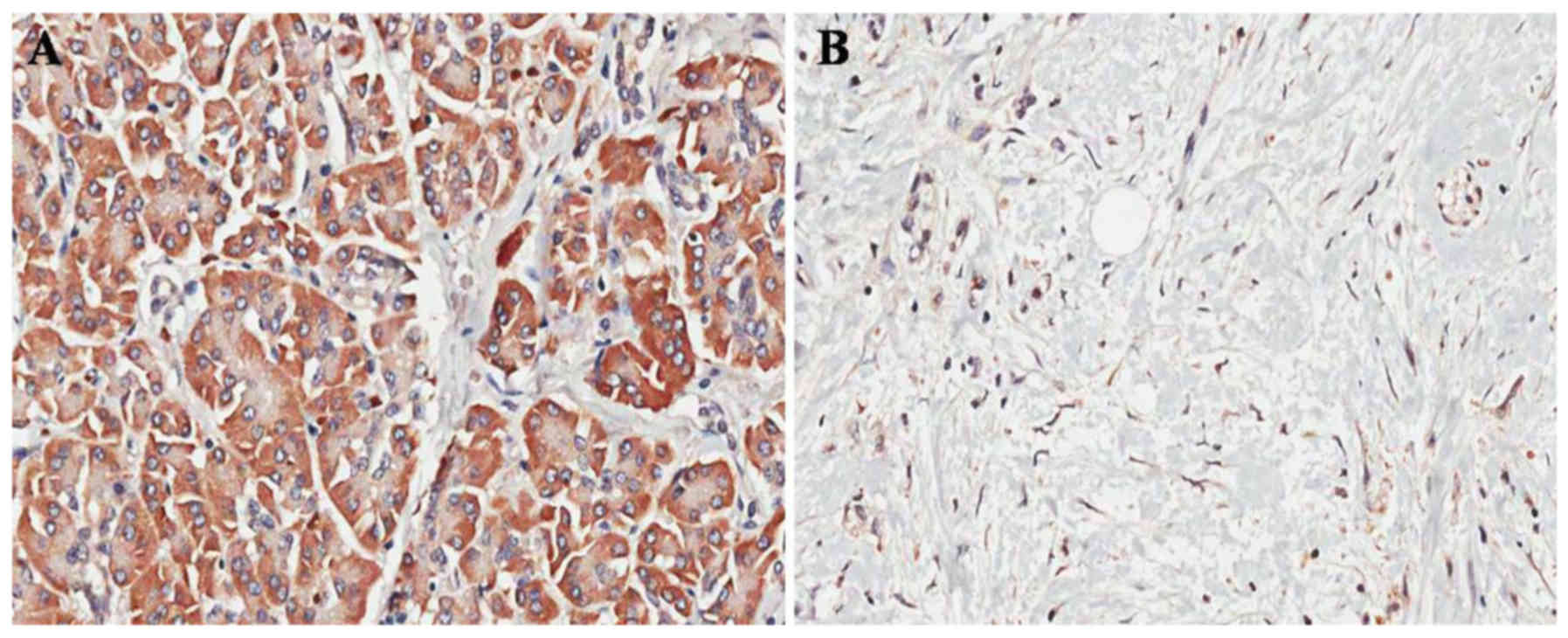
specimens was examined by IHC. Positive staining for CIP2A was
observed in 83.33% (30/36) of the ovarian cancer tissues vs. 13.89%
(5/36) of the paired adjacent non-cancerous tissues (P<0.001;
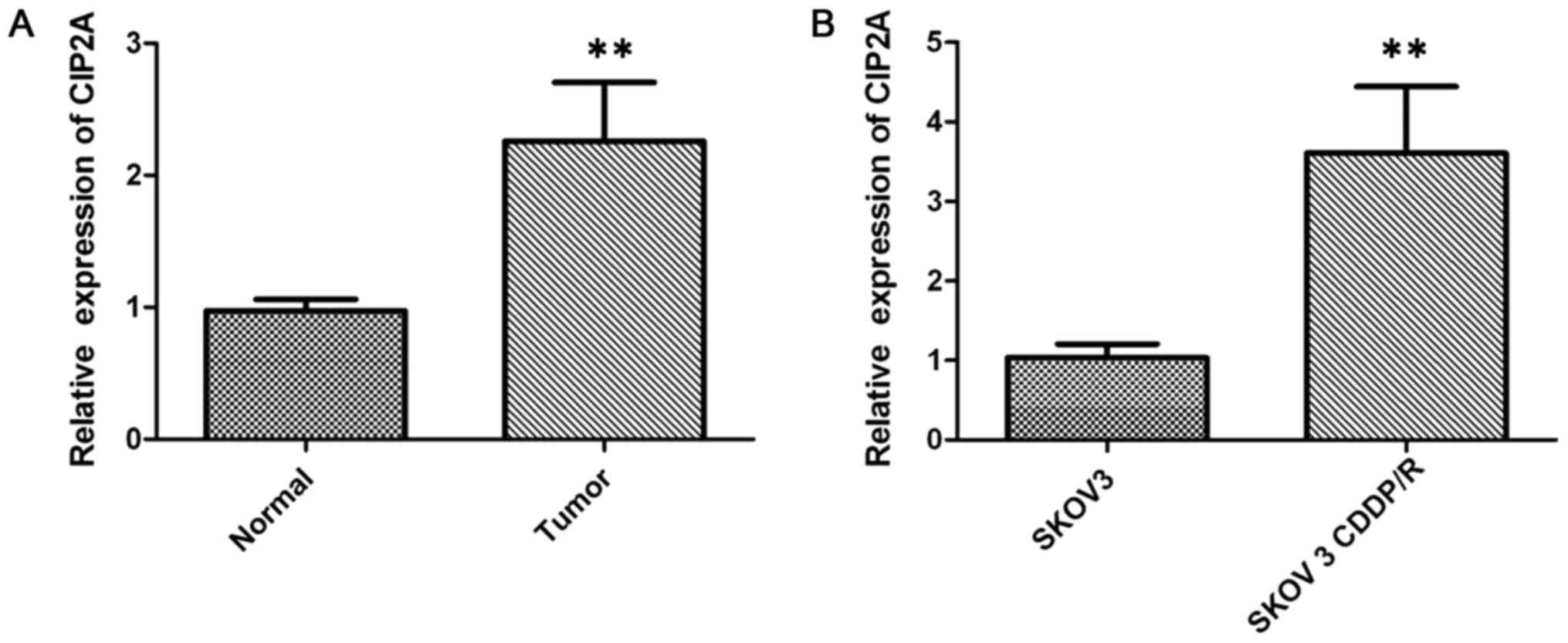
Fig. 1A and B). In addition, CIP2A
mRNA expression was also examined by RT-qPCR in cancer tissues. The
CIP2A mRNA expression was significantly higher in ovarian cancer
tissues than in paired adjacent non-cancerous tissues (P<0.01;
Fig. 2A).
Cisplatin-resistant ovarian cancer
cells exhibited higher CIP2A expression
The expression level of CIP2A was examined by
RT-qPCR to study its regulatory role in chemotherapy resistance in
SKOV-3 (cisplatin-sensitive) and SKOV-3CDDP/R (cisplatin-resistant)
ovarian carcinoma cells, with the IC50 value to
cisplatin of the latter cell line being significantly higher than
that of the SKOV-3 cells. The results demonstrated that the
expression of CIP2A in the SKOV-3CDDP/R cells was nearly 3-fold
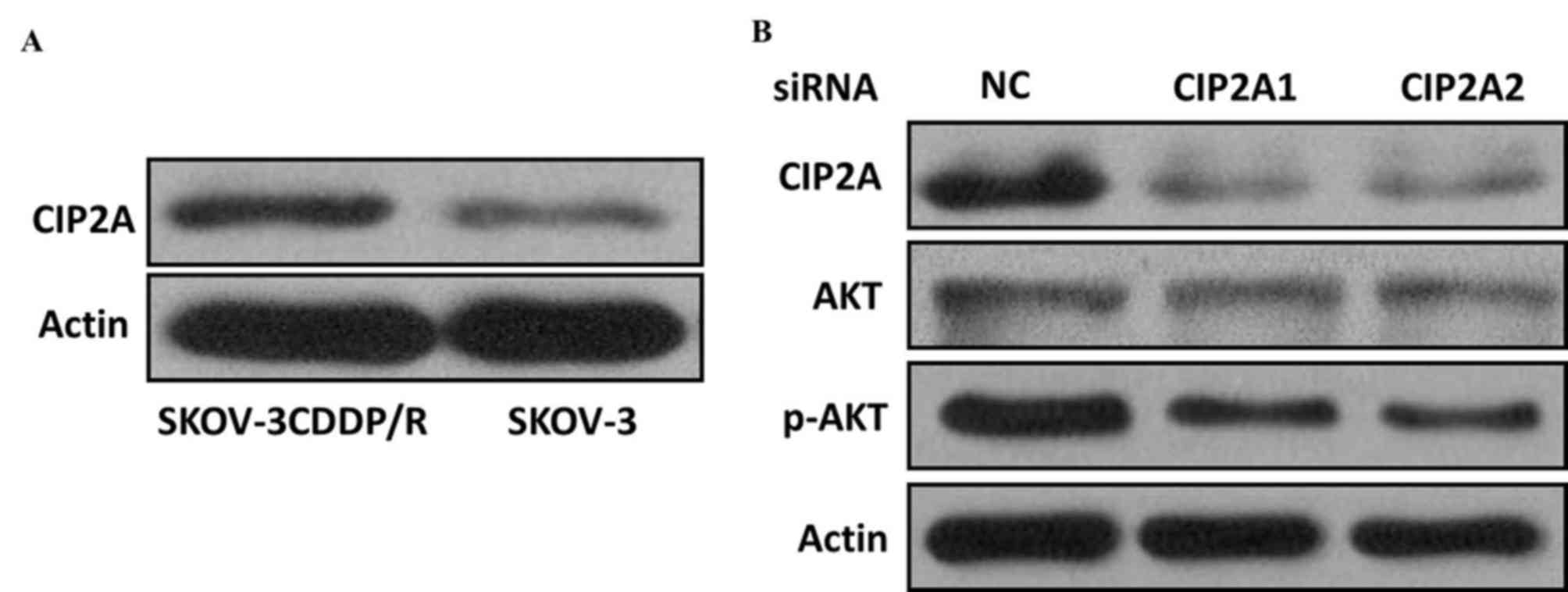
higher than that in the SKOV-3 cells (P<0.001; Fig. 2B). Western blot analysis further
confirmed that the CIP2A protein expression was elevated in
cisplatin-resistant cells but decreased in cisplatin-sensitive
cells (Fig. 3A), suggesting that
CIP2A may serve an important role in inducing the cisplatin
resistance of ovarian cancer cells.
CIP2A induces cisplatin resistance and
proliferation in vitro
In order to further investigate the role of CIP2A in
the cisplatin resistance of ovarian cancer cells, two siRNA
constructs, siCIP2A1 and siCIP2A2, were utilized to interfere with
CIP2A expression in ovarian cancer cells. SKOV-3 and SKOV-3 CDDP/R
cells were transiently transfected with control and siCIP2A for 48
h. The two different CIP2A siRNAs acquired a knockdown efficiency
of ~50%, which was confirmed by RT-qPCR. Furthermore, the CIP2A
protein was inhibited using siCIP2A in SKOV-3 CDDP/R cells
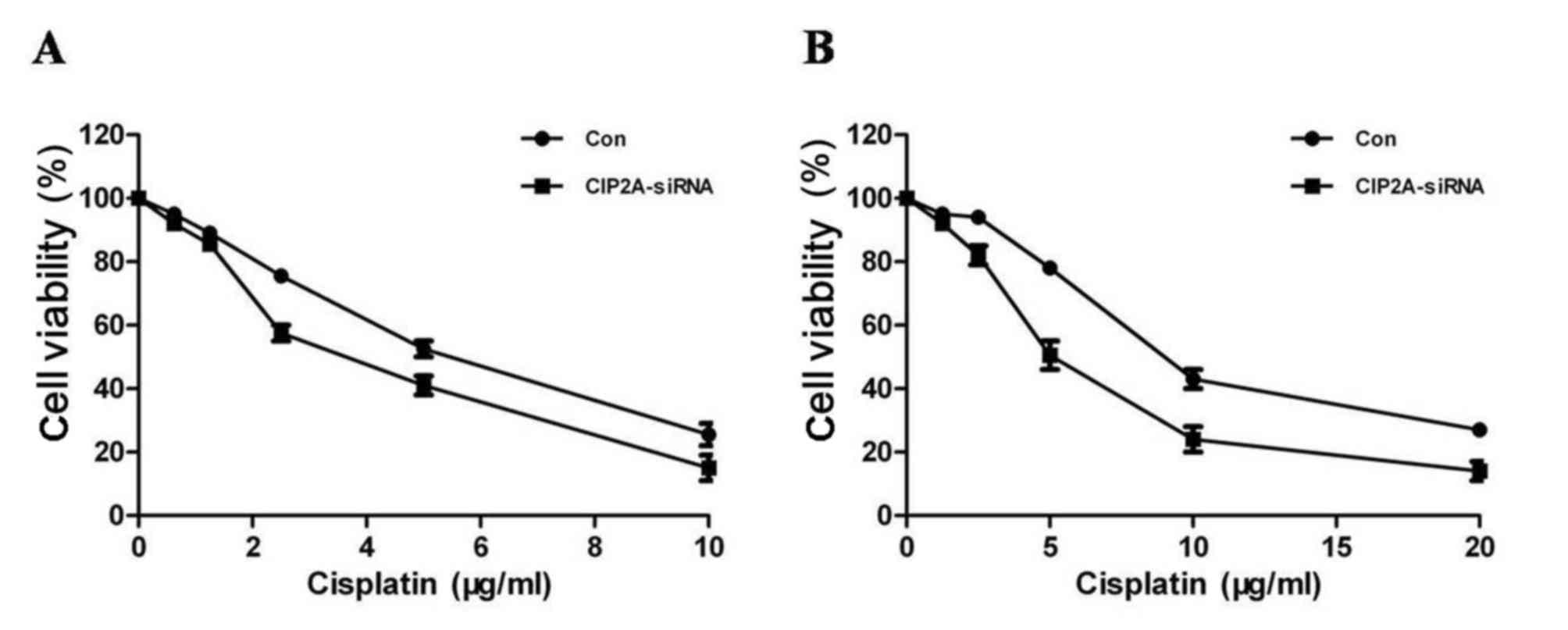
(Fig. 3B). The CCK8 assay revealed
that CIP2A silencing decreased SKOV-3 cell proliferation (Fig. 4A), suggesting that CIP2A may
participate in the development of ovarian cancer. In addition,
SKOV-3 CDDP/R cells were more sensitive to cisplatin and exhibited
a decreased proliferation rate following CIP2A suppression
(Fig. 4B).
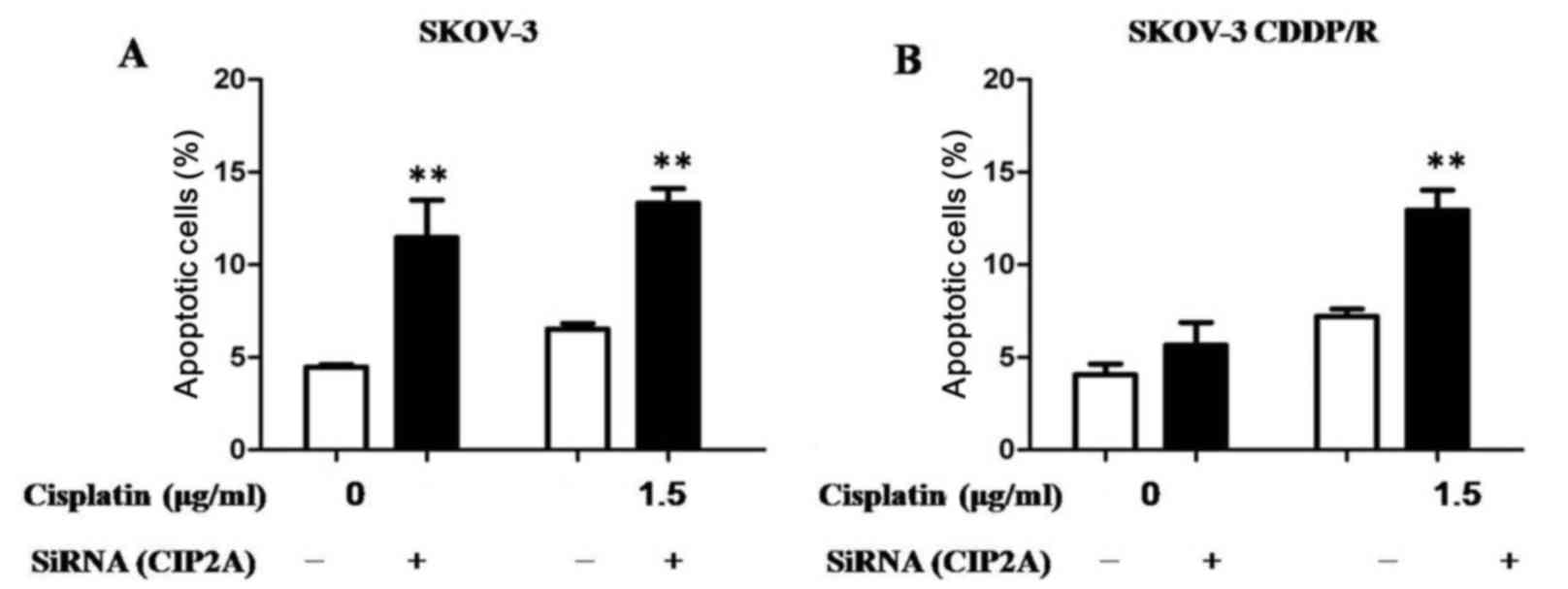
Depletion of CIP2A induces
apoptosis
The primary antitumor mechanism for cisplatin
results from the ability of cisplatin to induce apoptosis.
Subsequently, the CIP2A-mediated chemosensitivity of ovarian cancer
cells was determined using an apoptosis assay. Regardless of
whether the cells were transfected with CIP2A or the NC siRNA,
cisplatin was able to significantly induce the apoptosis of ovarian
cancer SKOV-3 cells, indicating that depletion of CIP2A has no
additive effects to cisplatin in SKOV-3 cells (Fig. 5A). Notably, it was revealed that
cisplatin induced a higher proportion of apoptotic SKOV-3CDDP/R
cells following CIP2A knockdown compared with transfection with NC
siRNA, suggesting that CIP2A silencing may enhance
cisplatin-mediated cell apoptosis (Fig.
5B).
Discussion
Ovarian cancer is one of the leading causes of
mortality of all gynecological tumors worldwide (11). The development of chemoresistance is a
major challenge for patients with ovarian cancer (12). Although certain patients with ovarian
cancer exhibit favorable responses to the initial chemotherapy
treatment, the majority of them eventually succumb to
chemoresistance. In those patients exhibiting chemoresistance, the
5-year overall survival rate is only 31% (13). Therefore, novel approaches to
restoring the chemotherapy sensitivity of ovarian cancer are
urgently required.
Recent studies have indicated that elevated CIP2A
expression in numerous types of cancer is associated with a poor
prognosis (14–18); however, the clinical significance of
CIP2A in ovarian cancer, including chemoresistance, has yet to be
reported. To begin with, the expression levels of CIP2A in ovarian
cancer tissues and the cisplatin-resistant ovarian cancer cell line
were analyzed, in order to determine the influence of CIP2A in the
drug resistance of patients with ovarian cancer. The results of the
present study revealed that the mRNA and protein expression of
CIP2A was significantly upregulated in ovarian cancer tissues
compared with that in the paired adjacent non-cancerous tissues. In
addition, it was revealed that CIP2A expression was significantly
higher in the cisplatin-resistant ovarian cancer SKOV-3CDDP/R cell
line, compared with that in the cisplatin-sensitive ovarian cancer
SKOV-3 cell line. Using siCIP2A, the effect of CIP2A knockdown in
SKOV-3CDDP/R and SKOV-3 cells was analyzed. Transient CIP2A
depletion was able to suppress ovarian cancer cell proliferation
while enhancing the sensitivity of these cells to cisplatin
suggesting that CIP2A is involved in ovarian carcinogenesis and
progression. In cisplatin resistant cells, CIP2A inhibition
significantly sensitized cancer cells to cisplatin compared with
control cells, suggesting that directly targeting CIP2A may be a
promising approach to increase the sensitivity of ovarian cancer
cells to cisplatin.
Cisplatin-induced cancer cell apoptosis has been
previously studied in detail (19).
The decrease in CIP2A expression, as well as the inactivation of
the Akt pathway, may attenuate the proliferation and induce the
apoptosis of numerous types of cancer cells (20). However, whether or not CIP2A is a key
protein that affects cisplatin-induced apoptosis remains unknown.
The results of the present study demonstrated that, compared with
NC siRNA, suppression of CIP2A contributed to a significant
increase in the apoptosis of SKOV-3 cells when treated with
cisplatin (Fig. 3). Notably,
knockdown of CIP2A in the cisplatin resistant SKOV-3 CDDP/R cells
promoted the apoptotic cell death induced by cisplatin and
decreased the expression of phosphorylated Akt. Therefore,
appropriate combination of cisplatin and CIP2A depletion would be a
potential strategy for the treatment of ovarian cancer.
In conclusion, cisplatin resistance is a complicated
molecular process. The results of the present study revealed that
CIP2A is highly expressed in ovarian cancer tissues and
cisplatin-resistant cells. CIP2A knockdown may increase the
chemotherapeutic sensitivity of ovarian cancer cells to cisplatin.
Therefore, CIP2A may be a potential therapeutic target for
improving cisplatin sensitivity in ovarian cancer. Future clinical
trials should be designed to evaluate the usefulness of CIP2A
inhibitors in cisplatin-based chemotherapy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The data that support the findings of the present
study are available from JMU but restrictions apply to the
availability of these data, which were used under license for the
current study, and so are not publicly available. Data are however
available from the authors upon reasonable request and with
permission of JMU.
Authors' contributions
HZ conceived and designed the experiments. WL and HZ
performed the experiments. LY and YW performed the proliferative
ability analysis. WL and HZ performed the cell migration assay. WL
and HZ performed the cell cycle analysis and statistical analysis.
HZ provided oversight for all aspects of the study. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Local
Institutional Review Board of the Affiliated Hospital of Jining
Medical University, and written informed consent was obtained from
all patients.
Patient consent for publication
The patients provided written informed consent for
the publication of the present study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vaughan S, Coward JI, Bast RC Jr, Berchuck
A, Berek JS, Brenton JD, Coukos G, Crum CC, Drapkin R,
Etemadmoghadam D, et al: Rethinking ovarian cancer: Recommendations
for improving outcomes. Nat Rev Cancer. 11:719–725. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Capoluongo E, Ellison G, López-Guerrero
JA, Penault-Llorca F, Ligtenberg MJL, Banerjee S, Singer C,
Friedman E, Markiefka B, Schirmacher P, et al: Guidance statement
On BRCA1/2 tumor testing in ovarian cancer patients. Semin Oncol.
44:187–197. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vargas-Hernández VM, Moreno-Eutimio MA,
Acosta-Altamirano G and Vargas-Aguilar VM: Management of recurrent
epithelial ovarian cancer. Gland Surg. 3:198–202. 2014.PubMed/NCBI
|
|
5
|
Li W, Ge Z, Liu C, Liu Z, Björkholm M, Jia
J and Xu D: CIP2A is overexpressed in gastric cancer and its
depletion leads to impaired clonogenicity, senescence, or
differentiation of tumor cells. Clin Cancer Res. 14:3722–3728.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Junttila MR, Puustinen P, Niemelä M, Ahola
R, Arnold H, Böttzauw T, Ala-aho R, Nielsen C, Ivaska J, Taya Y, et
al: CIP2A inhibits PP2A in human malignancies. Cell. 130:51–62.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Khanna A, Böckelman C, Hemmes A, Junttila
MR, Wiksten JP, Lundin M, Junnila S, Murphy DJ, Evan GI, Haglund C,
et al: MYC-dependent regulation and prognostic role of CIP2A in
gastric cancer. J Natl Cancer Inst. 101:793–805. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Côme C, Laine A, Chanrion M, Edgren H,
Mattila E, Liu X, Jonkers J, Ivaska J, Isola J, Darbon JM, et al:
CIP2A is associated with human breast cancer aggressivity. Clin
Cancer Res. 15:5092–5100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Choi YA, Park JS, Park MY, Oh KS, Lee MS,
Lim JS, Kim KI, Kim KY, Kwon J, Yoon DY, et al: Increase in CIP2A
expression is associated with doxorubicin resistance. FEBS Lett.
585:755–760. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yin X, Zhang N and Di W: Regulation of
LC3-dependent protective autophagy in ovarian cancer cells by
protein phosphatase 2A. Int J Gynecol Cancer. 23:630–641. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Miller DS, Blessing JA, Krasner CN, Mannel
RS, Hanjani P, Pearl ML, Waggoner SE and Boardman CH: Phase II
evaluation of pemetrexed in the treatment of recurrent or
persistent platinum-resistant ovarian or primary peritoneal
carcinoma: A study of the gynecologic oncology group. J Clin Oncol.
27:2686–2691. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Burger RA: A new model of ovarian
carcinogenesis may influence early detection strategies. Am J
Obstet Gynecol. 198:349–350. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu J, Wang M, Zhang X, Wang Q, Qi M, Hu
J, Zhou Z, Zhang C, Zhang W, Zhao W and Wang X: CIP2A is associated
with multidrug resistance in cervical adenocarcinoma by a
P-glycoprotein pathway. Tumour Biol. 37:2673–2682. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen KF, Liu CY, Lin YC, Yu HC, Liu TH,
Hou DR, Chen PJ and Cheng AL: CIP2A mediates effects of bortezomib
on phospho-Akt and apoptosis in hepatocellular carcinoma cells.
Oncogene. 29:6257–6266. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen KF, Pao KC, Su JC, Chou YC, Liu CY,
Chen HJ, Huang JW, Kim I and Shiau CW: Development of erlotinib
derivatives as CIP2A-ablating agents independent of EGFR activity.
Bioorg Med Chem. 20:6144–6153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tseng LM, Liu CY, Chang KC, Chu PY, Shiau
CW and Chen KF: CIP2A is a target of bortezomib in human triple
negative breast cancer cells. Breast Cancer Res. 14:R682012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu HC, Hou DR, Liu CY, Lin CS, Shiau CW,
Cheng AL and Chen KF: Cancerous inhibitor of protein phosphatase 2A
mediates bortezomib-induced autophagy in hepatocellular carcinoma
independent of proteasome. PLoS One. 8:e557052012. View Article : Google Scholar
|
|
19
|
Chattopadhyay S, Machado-Pinilla R,
Manguan-Garcı'a C, Belda-Iniesta C, Moratilla C, Cejas P,
Fresno-Vara JA, de Castro-Carpeño J, Casado E, Nistal M, et al:
MKP1/CL100 controls tumor growth and sensitivity to cisplatin in
non-small-cell lung cancer. Oncogene. 25:3335–3345. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma L, Wen ZS, Liu Z, Hu Z, Ma J, Chen XQ,
Liu YQ, Pu JX, Xiao WL, Sun HD and Zhou GB: Overexpression and
small molecule-triggered downregulation of CIP2A in lung cancer.
PLoS One. 6:e201592011. View Article : Google Scholar : PubMed/NCBI
|