Introduction
As the largest organ, skin interacts with various
environmental factors (1), shields
the organisms from radiation, protects it from mechanical pressure
and creates one of the main barriers against pathogens (2). Among the main environment aggressors,
the UV radiation (UVA and UVB) from the sun induces several
biological effects, high DNA damage and reactive oxygen species
(ROS) generation. These alterations can induce inflammation and can
further initiate tumorigenesis (3,4). ROS and
damaged DNA can activate intracellular protein complexes such as
inflammasomes (5). Stimulated
keratinocytes, the main skin cellular population, along with
melanocytes and resident dendritic cells (DCs) and Langerhans cells
(LCs), secrete cytokines with pro-inflammatory action (6), and these molecules modulate innate and
adaptive immune responses (7). All
the immune-related molecules, cytokines, chemokines and non-immune
molecules, such as growth factors have both paracrine and autocrine
effects upon the microenvironment and design the local millieu that
initiates and then regulates local inflammation or can lose
control, consequently favoring the process of tumorigenesis
(8). Inflammation has acute and
chronic stages, but its link to tumorigenesis is carried out by
chronic inflammation. While acute inflammation is governed by
T-helper (Th)1-polarized T lymphocytes attracted by innate immune
cells, secreting mainly antitumor immune molecules such as
interleukin (IL)-2 and interferon (IFN)-γ, chronic inflammation is
controlled by regulatory T cells (Tregs), Th2 cells, that secrete
pro-tumorigenic factors [e.g., IL-4, IL-6, IL-10, IL-13,
transforming growth factor (TGF)-β]. T cell populations that
attract and activate B cells thus favor tumorigenesis and anergize
cytotoxic T lymphocytes (CTLs) (9).
In this chronic inflammatory milieu, cells and
molecules interact and pro-tumoral microenvironment is sustained
(10,11). In skin, as this organ is subjected to
a myriad of environmental factors, this chronic inflammatory
condition can trigger various processes underlying tumorigenesis
affecting the cell components of its structure. We will focus
herein on the inflammatory portrait of non-melanoma skin cancer
(NMSC) and melanoma cancer, as complex molecular networks that can
be both triggers of tumorigenesis and therapeutic targets.
Inflammasomes
There are several non-immune cells that participate
in the portrait of inflammation. Various agents can activate the
main cellular component of the skin, the keratinocytes. Subsequent
to stimulation, these cells secrete pro-inflammatory cytokines
(6), regulating both innate and
adaptive immunity (7). Keratinocytes,
and in addition to these cells and lesser in number, melanocytes
and skin resident DCs (LCs), secrete immune-related molecules
(e.g., cytokines, and growth factors) that create a local
microenvironment depicting inflammation and further tumorigenesis
(12). Keratinocytes can play a dual
role in T cell activation. Hence, by the secretion of IL-1,
granulocyte-macrophage colony stimulating factor (GM-CSF), tumor
necrosis factor (TNF)-α, IL-6, IL-7, IL-12, IL-15, IL-18, an
upregulation of T cell functions can be achieved. IL-1Rα, IL-10,
α-melanocyte stimulating hormone (α-MSH), chemokine (C-X-C motif)
ligand 10 (CXCL10), contra IL-1, prostaglandin E2 (PGE2) secreted
by keratinocytes can downregulate T cell functions. In contast, T
cells can produce IFN-α, IL-17 and IL-4 that affect the functions
of keratinocytes (13). Fig. 1 shows the keratinocyte-T lymphocyte
interaction in the skin, interaction mediated by cytokines and
chemokines.
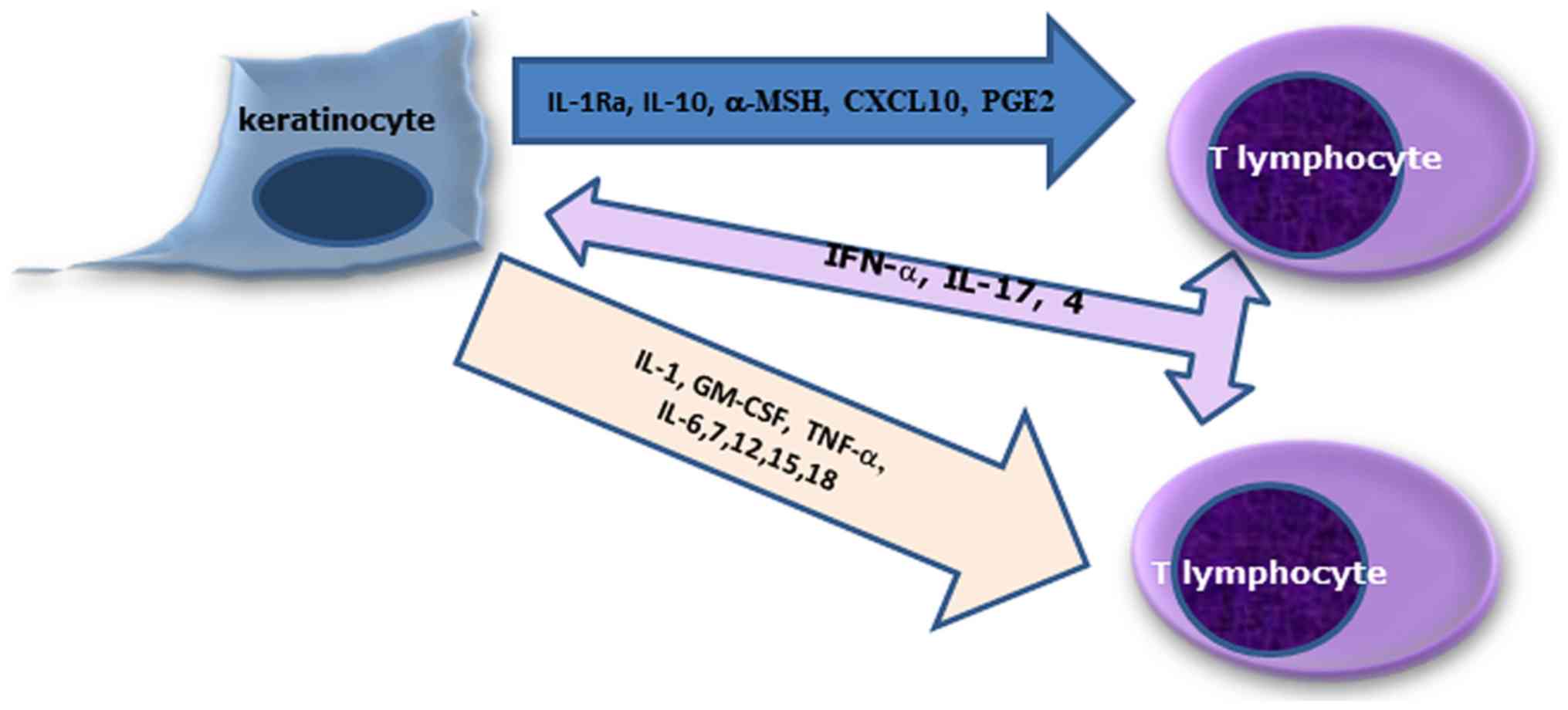 | Figure 1.Relation between keratinocytes and T
lymphocytes. Keratinocytes secrete upregulatory cytokines and
chemokines IL-1, GM-CSF, TNF-α, IL-6, IL-7, IL-12, IL-15 and IL-18,
and downregulatory ones, IL-1Rα, IL-10, α-MSH, CXCL10 and PGE2. T
cells produce IFN-α, IL-17 and IL-4 that influence keratinocyte
functions. Keratinocyte chemoattractant cytokines influence T-cell
trafficking: IL-1, IL-8, CCL27, CCL5, CCL17, CXCL10, MIG, IP9,
CCL20. |
Another cell that is regarded as a by-stander cell
in the skin, the sebocyte, was recently reported as being involved
in maintaining the skin's inflammatory milieu. These cells
encompass the pilosebaceous unit and secrete skin moisturizing
lipids. Yet, they also secrete chemokines and cytokines and act in
response to several pro-inflammatory stimuli and bacteria. Recently
it was reported that CD4+IL-17+ T cells are
in contact with sebocytes in acne lesions. Sebocyte secrete
chemokines (e.g., CXCL8) that call upon neutrophils, monocytes and
T lymphocytes. Cytokines (e.g., IL-6, TGF-β, IL-1β) secreted by
sebocytes induce the differentiation of
CD4+CD45RA+ naive T cells into Th17 cells
without affecting memory T cells. Practically, Mattii et al
presented the first report that proves human sebocytes are actively
involved in the skin's inflammatory processes. Furthermore, as the
main regulated cell is Th17 and, because it is known that the loss
of this regulatory T lymphocyte is linked to chronic inflammation,
sebocyte activity can be associated with pro-tumorigenesis
processes (14).
UVA and UVB radiation directly affects skin, and
this injury leads to DNA damage and ROS production. Furthermore,
these elements induce an inflammatory response and, if regulatory
mechanisms are surpassed, they can trigger pro-tumorigenesis
mechanisms (3,15). Moreover in this process, complex
protein platforms designated as inflammasomes are activated
(16) and the link between chronic
inflammation and distorted inflammasome activity is associated with
skin disorders including cancer (8,17).
Inflammasomes are complexes formed from a cytoplasmic sensor, an
adaptor known as apoptosis-associated speck-like protein (ASC) and
pro-caspase-1 (5) (Fig. 2). Actually inflammasomes are a large
family that is characterized by their sensors [e.g., absent in
melanoma 2 (AIM2), pyrin, NOD-like receptor with pyrin (NLRP)1,
NLRP3, NOD-like receptor with caspase recruitment domain (NLRC)4]
(18).
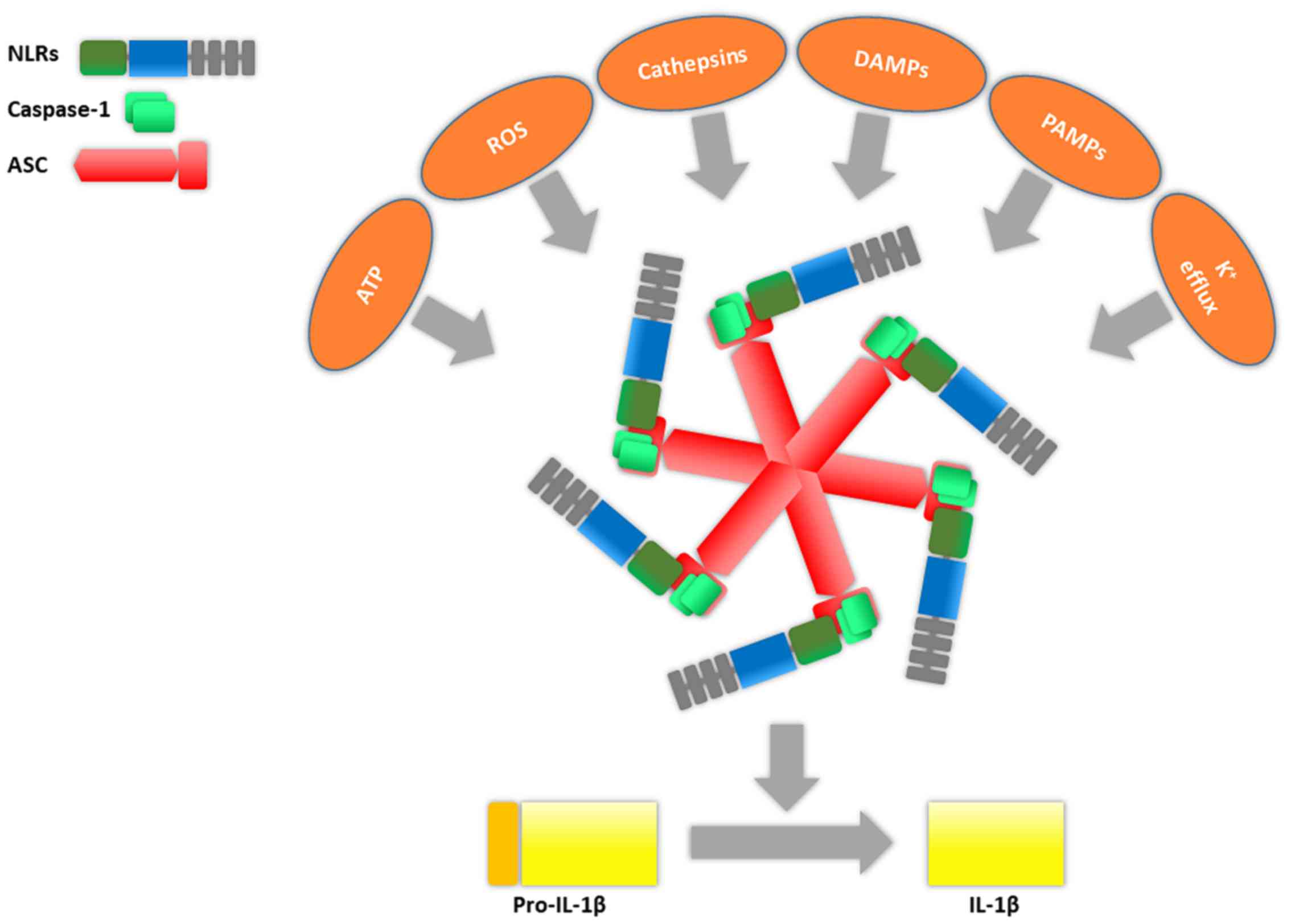 | Figure 2.Inflammasome basic structure consists
of caspase-1, NLRs and ASC. The specific composition of an
inflammasome is dependent on the activator, e.g., ATP, ROS,
cathepsins, DAMPs, PAMPs, K+ efflux. The main action of
the assembled inflammasome is to induce the conversion of pro-IL-1β
in IL-1β. NLRs, nucleotide-binding oligomerization domain and
leucine-rich repeat-containing receptors; ASC, apoptosis-associated
speck-like protein; ROS, reactive oxygen species. |
When UV radiation hits the skin, within sebaceous
lipids, squalene is oxidized and initiates inflammatory processes
(19) thus acting as inflammasome
activating danger signal (20). The
most frequent type of inflammasome, NLRP3, is increased in human
BCC along with higher IL-1β levels and caspase-1 activation
compared to normal skin (21).
Pannexin-1 channels involved in keratinocyte differentiation that
can induce inflammasome activation (22), were also identified in melanocytes and
were found to be upregulated throughout melanoma progression
(23). NLRP1 is strongly expressed in
human skin as compared to other NLRs (24). At the genetic level, polymorphisms
detected in NLRP1/NLRP3 (25) and in
inflammasome-related genes [e.g., caspase recruitment domain
(CARD)8, IL-1β and IL-18) were found to be associated with skin
melanoma, related to both susceptibility and progression (26). The inflammasome involvement was
demonstrated to be correlated with melanoma stages. Hence, cells
isolated from late stage human melanomas generate spontaneously
IL-1β without stimulation, showing auto-inflammatory
characteristics, namely gain-of-function mutations in NLRP3. This
study has pointed out that, IL-1β constitutive secretion can be
clearly linked to the aggressiveness of melanoma (27). The ASC component of the inflammasome
is involved in tumorigenesis in metastatic melanomas, while in
primary melanoma, it inhibits cancer cell growth (28). Of note, the role of ASC is different
in various cells involved in the link between inflammation and
tumorigenesis. ASC exhibits a tumor-suppressor function in
keratinocytes, while in myeloid cells are pro-tumorigenic (29). Collectively, recent information
suggests that among all NLRPs, NLRP1 is the main inflammasome
sensor in human skin (24,30). In skin models it was shown that
cytokine stimulation using IL-1α, IL-1β and IL-18 induces epidermal
hyperplasias (24), an adjacent
characteristic of tumor tissue.
Inflammation leading to
tumorigenesis
In normal homeostasis of the skin, inflammation is
self-limiting, while tumorigenesis has a constitutive activation
pathway (31). Long-term accumulation
of inflammatory factors in the skin tissue (e.g.,
cytokines/chemokines) may finally lead to an immunosuppressive
microenvironment that favors tumorigenesis. Cytokines are produced
in the skin by resident cells, namely keratinocytes, LCs,
melanocytes, mast cells (MCs) and macrophages, whereas recruited
cells, such as neutrophils, eosinophils and lymphocytes add
secreted cytokines to the tissue microenvironment (32). Upon cellular activation, cytokines are
rapidly secreted acting within the tissue, in both paracrine and
autocrine manner. In a prolonged inflammatory status, cytokine
synthesis and production are constantly enhanced, the
auto-regulatory loop is hindered and acts upon both neighboring
cells as well as upon distant cell populations. The action of
cytokines is vast as cytokine receptors can be homologous; namely
various cytokines lead to multidirectional effects. Moreover,
various cytokines can address the same receptor having a
synergistic effect on one cell type, while acting antagonistically
on another cell type. The cytokine cascade is different for acute
and chronic inflammation (Fig. 2) and
while one has antitumoral characteristics, the other one sustains
tumorigenesis (33), tumor cell
migration and cancer metastasis mechanisms (34).
There are various cell populations that intimately
interact (epithelial, mesenchymal and immune cells) for the
inflammatory process to develop and if chronically induced, further
triggers tumorigenesis. Epithelial-mesenchymal transition (EMT) is
a process involved in tumorigenesis mechanisms. It is actually a
process through which epithelial cells lose cell polarity and
cell-cell adhesion, and are able to migrate and invade other
tissues becoming mesenchymal stem cells (35). While in physiological wound healing,
EMT induction (36) is beneficial, as
epidermal keratinocytes acquire migratory phenotypes for wound
re-epithelialization (37), in
tumors, EMT is uncontrolled and epithelial cells acquire oncogenic
mutations (38).
Another cell population that can contribute to the
link between inflammation and tumorigenesis in the skin is the
fibroblast. During wound healing fibroblasts deposit collagen in
excess (fibrosis), fibrotic connective tissue being actually a
pro-tumorigenesis microenvironment (39).
All the recent reports state that there is a clear
relationship between chronic tissue damage, inflammation and
cancer. At chronic inflammation sites, triggered by various
intrinsic and extrinsic factors, tumors can develop. A hindered
regulatory loop for an efficient inflammation process would lead
toward tumorigenesis. Once the program for tumorigenesis is
installed, continuous inflammation supports metastatic progression
(31).
Non-melanoma tumors: squamous cell and basal
cell skin carcinomas
Worldwide, NMSC is the most frequent type of skin
cancer in Caucasian populations, registering an increased incidence
in the last 40 years. This rapid increase is based on UV radiation
exposure accounting for 90% of NMSC cases. Although the incidence
of other malignancies has stabilized or even declined, the
incidence of NMSC has increased constantly with a younger patient
age at diagnosis, representing a main public health concern
(40).
Although NMSC is not a main life-threatening
disease, the direct social costs involved in such a widely
spreading disease are high. Studies concerning the early diagnosis
along with prevention and therapy are the main domains that are
constantly evolving in NMSC. Etiopathogenic mechanisms are
intensively studied and, within, a special focus has been given to
the mechanisms that link inflammation and skin tumorigenesis. Basal
cell carcinoma (BCC) and squamous cell carcinoma (SCC) are the most
frequent NMSC, registering the highest world incidence in Auckland
where rates were 425 for SCC and 1,177 for BCC per 100,000
individuals per year (41).
Histological and immunohistological evaluation of SCC and BCC has
shown that peritumoral inflammatory reaction is diverse in
intensity and distribution, proving the complexity of the immune
cells and tumor cells. In SCCs, the inflammatory reaction is
increased in comparison to BCC. Inflammatory immune cells such as
T-lymphocytes, macrophages and MCs are in direct contact with tumor
cells, sustaining intercellular interaction mechanisms (42).
In addition to immune cells, pro-inflammatory and
tumor-promoting molecules have been reported in BCC and SCC.
Recently, high-mobility group box-1 (HMGB1) protein and the
receptor for advanced glycation end products (RAGE) have been
studied in NMSC. RAGE was reported as being upregulated in SCC
along with advanced stage cutaneous melanoma with poor prognosis.
In BCC and SCC, extracellular expression of HMGB1 was reported to
be released by necrotic tumor cells but further studies are
warranted (43).
Other new associated inflammatory proteins are the
galectin family involved in various pathologies related to skin
through their intracellular and extracellular mechanisms. Skin
structural cells such as keratinocytes, melanocytes, fibroblasts
and endothelial cells express galectins. Immune cells resident in
the skin, such as DCs, lymphocytes and macrophages express these
proteins as well. Non-malignant skin pathologies with high
inflammatory background, such as atopic dermatitis, psoriasis,
contact dermatitis and wound healing are associated with an
increased galectin expression. But recently, skin cancers have been
demonstrated to express these known inflammatory proteins involved
in regulatory mechanisms (44).
Patterns of galectin-3 expression in BCC and SCC were evaluated in
relation to cellular differentiation. The study revealed that there
is a specific pattern where decreased nuclear galectin-3 expression
and cytoplasmic immunoreactivity can be factors involved in SCC
tumor aggressiveness (45).
Specific inflammatory actors, such as IL-17 and
IL-22 were recently studied in SCC and BCC. In both NMSCs, T
lymphocytes that secrete IL-17 and IL-22 are abundant. In BCC and
SCC cell lines, proliferation and migration abilities were
significantly increased by in vitro IL-17 and IL-22.
Furthermore, IL-17 alone or combined with TNF-α, induced the
synthesis and production of two known pro-tumor cytokines, IL-6 and
IL-8. In animal models IL-17 and IL-22 increased tumor growth
proving once more that inflammatory cytokines such as IL-22 and
IL-17 in NMSC promote a tumorigenesis microenvironment (46).
As skin carcinogenesis induced by UV irradiation is
a constant research domain, it was shown that UV activates
oncogenes while inactivating tumor-suppressor genes. Inflammatory
milieu created by infiltrating immune cells contributes to the
chronic inflammation and to the progression of skin tumors
(40). In the link between UV
irradiation and skin inflammation, a special focus has been
developed in the research of vitamin D suggesting that vitamin D
receptor (VDR) gene polymorphisms can favor BCC and SCC. During an
11-year follow-up, a recent study has shown that patients with
rs2228570, rs927650 and rs1544410 dominance while rs7975232 and
rs739837 recessive genotypes were linked to a lower risk to develop
BCC. This is one of the few studies that links a genetic VDR
specificity to the risk of developing NMSC (47).
In addition to UV irradiation, other factors that
induce inflammation and further tumor development have been
studied. Hence in a prospective study reported in 2017, QSkin,
involving over 40,000 patients, it was shown that for smokers the
risk for developing SCC was high without a link with the duration
and/or intensity of smoking (48).
In a mouse model of UV carcinogenesis the
anti-inflammatory naproxen was tested to evaluate its
anti-proliferative action. Naproxen inhibited UVB-induced BCC and
SCC reducing both tumor number and volume. The overall
anti-proliferative effect was associated with reduced proliferating
cell nuclear antigen (PCNA) and cyclin D1 expression and increased
apoptosis. All markers that are usually associated with
inflammation [e.g., inducible nitric oxide synthase (iNOS),
cyclooxygenase-2 (COX-2), nuclear factor (NF)-κBp65] were
decreased. Tumors from treated animals had lower invasiveness with
increased E-cadherin expression and reduced expression of EMT
markers (e.g., N-cadherin, vimentin, Snail, Twist). In BCC and SCC
cells, it was shown that naproxen reduced UVB-induced skin
carcinogenesis through reducing endoplasmic reticulum (ER) stress
pathways (49).
Another molecule involved in inflammation pathways,
Toll-like receptor 4 (TLR4), is involved also in
photo-immunosuppression and chemically-induced carcinogenesis.
During the switch from normal skin to actinic keratosis (AK), TLR4
appears in keratinocytes and it is enhanced once more when the
keratinocyte progresses to SCC. In vitro silencing of TLR4
within keratinocytes blocks UV stress. A TLR4 antagonist,
resatorvid, blocks several inflammatory pathways such as NF-κB and
MAP kinase/AP-1 and hinders cytokine expression, including IL-6,
IL-8 and IL-10. This effect was reproduced also in animal models
(50).
Inflammation in NMSC is a hallmark of tumorigenesis
and several deregulated pathway conjoin in initiating neoplastic
transformation (51).
Inflammation in SCCs
Keratinocytes are the main cells of the epidermis
whose deregulated proliferation could initiate NMSCs. Inflammatory
pattern associated with carcinogenesis can furbish new therapeutic
targets or new inflammatory markers for diagnosis and prognosis
(52,53). Proteome profiling of keratinocytes
identified 50 proteins related to this type of cell, some of them
associated with the immune system [e.g., α-2 macroglobulin-like
protein-1, α-2 macroglobulin-like protein 2 and IFN regulatory
factor-6 (IRF-6)], others involved in differentiation (e.g.,
dermokine and calmodulin like protein 5) and others involved in
motility (e.g., integrin β4) (54).
However the mentioned proteins are deeply regulated by the
inflammatory status, hence when keratinocytes were stimulated with
IL-1β, α-2 macroglobulin like protein-1 and integrin β4 were found
to be reduced. IL-1β stimulation increased the NF-κB pathway,
highly involved in angiogenic and pro-tumorigenic processes. In
epidermoid carcinoma cells all of these alterations were also found
to support this link between inflammation and carcinogenesis
(54).
SCC is the most frequent metastatic NMSC and an
inflammation pattern would indicate cancer progression. As stated
above, inflammasome proteins are a new expanding research domain.
In SCC cell lines upregulation of the AIM2 sensor at both the
protein and mRNA levels has been reported. In human SCC samples,
tumor-specific expression of AIM2 was noted with low or absent
expression in AK and normal skin. Following knockdown of AIM2 in
specific tumor cell lines, cellular viability was decreased and
apoptosis was increased. Moreover this knockdown decreased the
invasive capacity of SCC cell lines associated with the reduction
in matrix metalloproteinases (MMPs), MMP1 and MMP13. In vivo
inoculated knockdown xenografts displayed a lower rate of tumor
growth and vascularization. These experimental results point toward
a component of the inflammasome machinery that can be a future
therapeutic target (55). Besides MMP
supra-expression (MMP1, MMP3, and MMP-9) in human samples it was
reported that keratins 6, 16, and 17 are overexpressed in
keratinocytes. When collagen type I synthesis is reduced upon UV
irradiation, this triggers the TGF-β pathway, a cytokine highly
involved in inflammatory processes (56–58).
Related to the activity of MMP proteins, adherence to the
extracellular matrix (ECM) is important in the migratory process of
tumor cells. A study published in 2018 focused on the aggressive
characteristics of SCC and a series of ECM traits. Using proteomic
and histologic tests for human primary SCC tumor tissue, three
types of samples were analyzed: non-recurring, non-metastasizing,
metastasizing SCC and SCC from patients with recessive dystrophic
epidermolysis bullosa (RDEB). Patients with the RDEB genetic
disorder harbor mutations that hinder both the function and the
amount of type VII collagen and they have a high risk of developing
SCC. The most deregulated samples were proven for RDEB and SCC
samples that displayed the highest mutational rates with important
inflammation and consequent dermal ECM remodeling as tumor set-off
factors. High-risk SCC also may display an enhanced bacterial
challenge as inflammatory activating factors. This report discloses
ECM remodeling as a clear inflammatory trait related to SCC that
sustains the increased risk for a worse prognosis of the disease
(59). In an RDEB mouse model it was
shown that the persistence of chronic wounds leads to SCC
development. These non-healing wounds have high TGF-β1 expression,
enhanced fibrogenesis thus creating a pro-tumorigenic
microenvironment (60).
As mentioned in the inflammasome section, ASC
mediates the secretion of pro-tumorigenic cytokines. ASC expression
in human SCC was evaluated in non-metastatic and metastatic SCC.
After silencing ASC it was demonstrated that alteration in the
activation of innate immune cells can be linked to keratinocyte
activity. Restoring ASC induced AIM2 and NLRP3 activation. Thus it
was demonstrated that pro-tumorigenic inflammation is actually
induced in the tumor cell (61).
NF-κB signaling is extremely important for the
maintenance of immune equilibrium in epithelial tissues. In a mouse
model with IKK2 deletion in GFAP-expressing cells of the epidermis
increased expression of TNF was also found in the SCC-type lesions
(62).
In another mouse model, overexpression of
MMTV-FLAG-hPAD2 (PAD2OE) also induced SCC-type lesions. Skin
tumorigenesis was associated with inflammation in this mouse model.
PAD2OE lesions presented with high inflammatory cell infiltrates
and increased nuclear phosphorylated signal transducer and
activator of transcription 3 (STAT3). This report proves that
benign papillomas can be transformed to SCC lesions when inducing
an inflammatory microenvironment (63).
In addition to keratinocytes, a report published in
2017 showed that LCs, resident in the epidermis and in the
pilosebaceous structure, are involved in maintaining the physiology
of the skin. When an injury occurs, LCs process antigens and
circulate to the local lymph nodes and activate T cells. Thus, LCs
can activate immune effector cells for an anti-tumoral immune
response. Moreover LCs cooperate with NK lymphocytes controlling
the development of SCC. In contrast LCs can also have
pro-tumorigenic activity when involved in the activation of T
suppressor lymphocytes, allowing malignant transformation of
keratinocytes within SCC (64).
Glycosylphosphatidylinositol-anchored glycoprotein
(CD109) is expressed by immune cells, such as T lymphocytes,
activated T lymphoblasts, or non-immune cells including endothelial
cells and activated platelets (65),
but it is expressed also by human cancers, predominantly SCC.
Sunagawa et al demonstrated that CD109-deficient mice
display chronic skin inflammation and epidermal hyperplasia.
Recently, they showed that in CD109-deficient mice the dermis had a
higher level of TGF-β protein expression. In keratinocytes, SMAD
family member 2 (Smad2) phosphorylation and NF-E2-related factor 2
(Nrf2) expression were enhanced in primary keratinocytes along with
reduced apoptosis and DNA damage and reduced H-ras gene mutation
frequency. All these data suggest that CD109 deficiency suppresses
skin tumorigenesis by enhancing TGF-β/Smad/Nrf2 pathway activity
and decreasing the mutation frequency of the H-ras gene (66).
Mast cells (MCs) have been reported as components of
cancer microenvironment in melanoma, BCC, SCC, primary cutaneous
lymphomas, haemangiomas and Merkel cell carcinoma. Their role seems
to be dual. In several studies they were reported as having
pro-tumorigenesis action while in others an anti-tumorigenesis
effect. Tumor-associated MCs should be thoroughly investigated in
the future for establishing their clear role in skin cancers
(67).
As described above, the EMT process can be
pro-tumorigenesis, but the occurrence is different in SCC of
different origins. SCCs emerging from inter-follicular epidermis
(IFE) are well differentiated, while SCC that originates from hair
follicle (HF) stem cell-derived frequently exhibit EMT, with an
increased metastatic capacity. Therefore, IFE and HF
tumor-initiating cells have different gene networks associated with
different biological behavior (68).
In the stem cell niches, inflammatory factors can drive
pro-tumorigenesis processes. IL-27 can have both pro- and
anti-inflammatory properties, and using a K15-KRASG12D mouse model
IL-27 promoted papilloma incidence. IL-27 induced CD11b cells with
endothelin A receptor (ETAR)-positive phenotype. Thus, in SCC
patients, Dibra et al showed that IL-27RA-positive cells in
the tumor stroma are correlated with tumor de-differentiation
(69).
Inflammatory traits in basal cell
carcinomas
As approximately 2.8 million new patients are
diagnosed with BCC only in the USA each year, it remains a major
health issue (70). Several
inflammatory-related pathways were found in BCC. Hence, one
characteristic of BCC is the continuous activation of the Hedgehog
pathway due to mutations in the tumor-suppressor gene patch (Ptch)
that induces inactivation or due to mutation in Smoothened that
leads to activation. These mutations were considered as good
therapeutic target candidates combined with direct
anti-inflammatory approaches. There have been clinical trials
addressing drugs such as difluoromethylornithine, thymidine
dinucleotide, retinoids, non-steroidal anti-inflammatory drugs,
vitamin D3, and silibinin or even green and black tea components
(71).
Another inflammatory-related pathway studied in BCC
is the non-canonical NF-κB pathway. This pathway is dependent on
IκB kinase α (IKKα), and Jia et al demonstrated that nuclear
IKKα binds to the promoters of inflammatory factors. Moreover, it
seems that it binds to a stem cell marker, leucine-rich
repeat-containing G-protein coupled receptor 5 (LGR5), which
activates the STAT3 signaling pathway during tumorigenesis. By
knockdown of IKKα, tumor growth and the EMT process are inhibited
proving that IKKα is an oncogenic transformation factor through
stemness and inflammatory related gene activation; thus BCC
progression is directly linked to the inflammatory microenvironment
(72).
Another factor that promotes stemness in BCC is the
transcription factor, SRY-related HMG-box family of transcription
factors (Sox9). In a mouse model of BCC it was shown that Sox9 is
expressed at tumor initiation and that its expression is
Wnt/β-catenin-dependent. In this genetic model, Sox9 deletion and
constitutive activation of the Hedgehog signaling pathway abolished
BCC initiation highlighting Sox9 involvement in stemness, ECM
remodeling and de-differentiation within tumor development and
metastasis (73).
In another mouse experimental model using SENCAR
mice, inflammation was induced by UVB, and skin tumor initiation
was studied. It was reported that protein tyrosine kinase 6 (PTK6)
expression was increased upon UVB action. In Ptk6+/+ and
Ptk6−/− SENCAR mice exposed to UVB it was shown that in
wild-type PTK6 (Ptk6+/+) UVB induced increased
inflammation and increased PTK6 expression in basal epithelial
cells. This action was correlated with higher tumor frequency and
tumor load compared to Ptk6−/− mice. In human SCC the
activation of PTK6 was also highlighted. It seems that PTK6
contributes to UVB-dependent inflammation further increasing
tumorigenesis in skin (74).
In BCC patients there is a constant search of
inflammatory-related immune cells that can prognosticate disease
evolution and the focus falls on neutrophils, monocytes and
lymphocytes. In a recent retrospective study in <550 patients
white blood cells (WBCs), neutrophil and monocyte values were found
decreased in the BCC group compared to controls.
Neutrophil:lymphocyte ratios were found to be 3.24 in BCC and 3.59
in SCC, as compared to 5.06 in control group (75).
As NMSC is highly associated with UV excessive
irradiation there are intensive studies regarding the association
of vitamin D and skin cancer risk. The endocrine system and vitamin
D are highly involved in inflammation, cell growth and
differentiation (76). Vitamin D acts
through binding to the vitamin D receptor (VDR). In a huge study
reported by Lin et al, involving over 17,000 BCC cases
compared to over 250,000 controls, 2 single-nucleotide
polymorphisms (SNPs) at new loci were found related to BCC risk.
The study pointed out that inherited common variations in VDR are
linked to BCC development (77).
Another study performed by Kaukinen et al, also in 2017,
using an animal skin model, showed that MCs expressing VDR are
involved in UV-mediated immunosuppression. VDR enhanced expression
of CYP24A1 (a hydroxylase) that inactivates vitamin D3 metabolites.
In normal skin, up to 2.9% of the MCs were CYP24A1+,
with a high percentage of CYP24A1+ MCs in AK, SCC and
BCC. The finding that CYP24A1+ MCs in
keratinocyte-derived skin cancers is increased warrants further
study (78). Similarly in other
organs, increased expression of CYP24A1 in skin could be correlated
in murine models with inflammation and progressive fibrosis
(79).
The process of tumorigenesis includes several
additional processes such as neo-vascularization, tissue invasion,
and metastasis. All of these processes rely on tissue remodeling
where the urokinase system is highly involved. Rubina et al
demonstrated in 2017 that as BCC is associated with keratinocyte
hyper-proliferation, inflammatory cell migration, and
angiogenesis-processes, increased urokinase plasminogen activator
receptor (uPAR) expression was found in the tumor surrounding
stroma in BCC. Hence the uPA system is a molecular network that
sustains aggressive proliferation and tumor cell invasion (80). Another molecular system that favors
inflammation and tissue remodeling upon tumorigenesis are MMPs.
Their activation is involved in the degradation of the basement
membrane in processes such as inflammation, wound healing,
angiogenesis and carcinogenesis. In BCC, MMP1 and MMP9 expression
was found to be associated with disease progression. Thus, low
levels were detected in AK foci, while intense expression was found
in different types of BCC (81).
Several anti-inflammatory compounds support the
proof for the link of inflammation with tumorigenesis. Hence,
naproxen, a known anti-inflammatory compound, has also
anti-proliferative and pro-apoptotic action. Chaudhary et al
used a mouse model of UVB-induced skin tumorigenesis where naproxen
significantly inhibited both BCC and SCC. The inhibition was
reflected on the lesion number and volume and the main reduction
was for BCC-type tumors. The effects were associated with decreased
PCNA and cyclin D1 expression, increased apoptosis and
inflammation-related molecules (e.g., iNOS, COX-2 and nuclear
NF-κBp65). Even remaining tumors after naproxen therapy displayed a
lower aggressive potential, lower EMT marker expression (e.g.,
N-cadherin, vimentin, Snail and Twist) and enhanced E-cadherin
expression (49).
Imiquimod is a TLR7 agonist, that addresses an
inflammatory-derived receptor, and it has been approved for in
situ SCC which was recently extended to superficial BCC with
positive clinical results. These results show a treatment strategy
that reduces inflammation and reduces tumorigenesis (82).
Classic anti-inflammatory compounds, such as aspirin
or other nonsteroidal anti-inflammatory drugs (NSAIDs) have been
tested in BCC. Collectively, data from 11 clinical studies showed a
10% risk reduction of BCC for patients using NSAIDs while the use
of aspirin had a weaker association with the decreased risk. This
report highlights that in humans NSAIDs can be used in high-risk
populations to reduce BCC initiation (83).
In summary, clinical data related to NMSC show that
anti-inflammatory therapeutic approaches can reduce significantly
UVB-induced skin carcinogenesis.
Inflammatory platform in melanoma: reshaping
discoveries
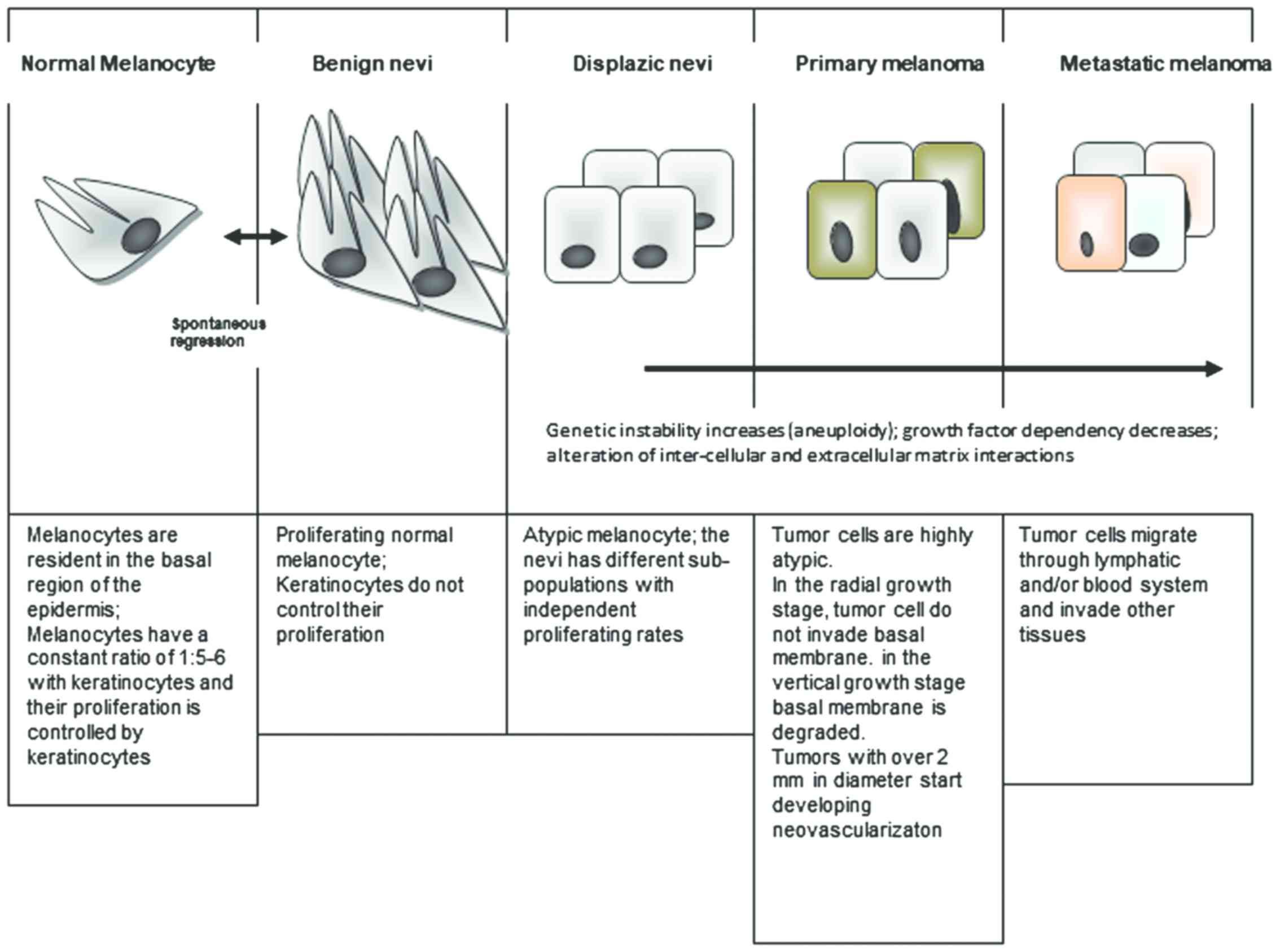
In the cellular flow of transforming a normal
melanocyte into a tumor cell there are several stages that occur
(Fig. 3). From benign nevi to a full
blown tumor cell, genetic instability and a pro-inflammatory milieu
can lead to tumorigenesis and metastasis. In the cellular
microenvironment, immune cells and immune-related molecules have a
definitive role in the inflammatory landscape. Although the typical
studied cellular interface in a tumor is between CTLs and cancer
cells, currently the contribution of other immune cells is widely
recognized. These other immune cells build the complex immune
response in cancer involving both tumor promotion and facilitating
cancer progression (84). In some
unpredictable cases, the clinical evolution of melanoma claim
additional prognostic markers to identify early stage and high risk
melanoma patients thus aiding in improving clinical surveillance
strategies and therapy management (85). One of these additional biomarkers
includes inflammatory immune cell infiltrate that depict the local
antitumor response or could trigger a pro-tumoral path (86).
Cellular profiles of the inflammatory
setting
The immune inflammatory infiltrate (IF) can be
considered as a ‘pro-inflammatory’ phenotype with infiltrating
cells and a cytokine pattern depending on immune activation. The
existence of IF is generally a good prognostic marker, but the
tumor milieu may lack immune cell infiltration as a consequence of
immune system ignorance and therefore tumor resistance occurs
impairing the favorable immune activation (84). For deciphering the correct role of IF,
we must understand its diverse cellular composition comprising
lymphoid cells (CTLs, Tregs, Th and B lymphocytes, NK cells) and
myeloid cells (DCs, myeloid-derived suppressor cells and
macrophages). Each cellular type imprints immunostimulatory or
immunosuppressive effects within the tumor site. Thus, it becomes
essential to also evaluate the functional status of IF within the
tumor environment, in an equal measure with its presence or absence
(84). Nevertheless, there is a large
data panel regarding the prognostic value of IF in cutaneous
melanoma but this subject remains controversial due to the
heterogeneity of patient groups, study methods and tumor
infiltrating lymphocyte (TIL) classification systems. To date, no
cancer staging algorithms integrate immune markers (85).
Myeloid cells in the tumor infiltrate:
Antigen-presenting cells in skin
DCs/LCs
DCs play a critical role residing at the border
between innate and adaptive immunity as specialized
antigen-presenting cells, being important in tailoring immune
reactivity, immune tolerance and activating antitumor immune
response (87).
DCs are one of the immune cells relevant for
melanoma antitumor immunity, having the potential to work as both
targets and delivery agents for immunotherapies. Although recent
immunotherapies do not directly involve DCs (e.g., immune
checkpoint blockade and adoptive cellular transfer), they rely on
DCs ability ‘to shape the quality’ of therapy-associated antitumor
immune response. Moreover, tumor-associated DCs are decisive for
improving melanoma immunotherapies as they are subjected to several
processes specific for melanoma milieu: they are activated by
immunogenic cellular death, could be suppressed by
melanoma-associated factors, could suffer metabolic constraints or
microbiome influences in mediating the anti-melanoma immune
responses (88). Specific DCs in the
skin immune network are represented by LCs (CD1a+,
Langerin+), phenotypically mature, but functionally
defective in melanoma-negative sentinel lymph nodes (SLNs)
(89). In the last few years, the
biology of LCs has considerably changed owing to novel insights in
the developmental origin and functions of these epidermis-specific
immune cells. LCs also have a significant impact on melanoma
pathology by either inducing immune tolerance or mediating
inflammatory processes (90). An
important inference for the antitumor response is that LCs may
activate more efficient naïve CD8+ T lymphocytes than
dermal DCs (91).
The active role of LCs should be integrated in
implementing or optimizing melanoma therapies. It was observed that
LCs are remarkably radio-resistant cells in radiotherapy treated
patients, and are highly potent in rapidly repairing DNA lesions
through cyclin-dependent kinase inhibitor (CDKN1A) involvement.
Such LC resistance has to be taken into account in light of new
immunotherapies targeting melanoma, where it is pivotal to
understand the immune cell dynamics within the tumor
microenvironment (92).
LCs might also play a role in EMT in cutaneous
cancers. One of the major regulators for LC activity is TGF-β which
also acts as a master controller of EMT processes in skin cancers
(93). Another regulator for LCs is
bone morphogenetic protein 7 (BMP7) responsible for a rested state
of LCs in the epidermis. On the other hand, BMP7 induces
mesenchymal to epithelial transition, thus acting as a homing
signal to epithelia (94). E-cadherin
expression loss occurring in EMT allows LCs to migrate out of the
epidermis, produces β-catenin stabilization in various skin tumors
and confers tolerogenicity of LCs (95). The migration of LCs from the skin to
the lymph nodes is mediated through Met signaling, a receptor
tyrosine kinase expressed on all DCs (96). In addition, Met signaling intercedes
the enzymatic activity of MMP2 and MMP9 that are vital for LCs to
break the basement membrane when migrating out of the epidermis
(97).
In thin melanomas, the presence of LCs is associated
with a better prognosis, by boosting antitumor defense through
antigen presentation to T CD8+ lymphocytes (98). Maturation phenotype of DCs can be a
prognostic factor in cutaneous melanoma. The density of DCs
expressing CD1a and the maturation marker DC-LAMP in primary tumors
were analyzed in melanoma cell nest infiltrates and in the
surrounding stroma. Thus, infiltration of CD1a+
LAMP+ DCs was inversely correlated with the thickness of
melanomas. High peritumoral density of mature
CD1a+/DC-LAMP+ DCs combined with high number
of CD25+/OX40+ T lymphocytes were associated
with extended survival. A ratio of high mature DCs/high
OX40+ T cells and Breslow index are reported as
independent predictors of good prognosis and indicators of a
functional immune response in primary cutaneous melanoma (99).
Myeloid-derived suppressor cells
Originated from bone marrow, myeloid-derived
suppressor cells (MDSCs) are a subset of immune cells with myeloid
origin and immunosuppressive properties, their development being
modulated by different tumor-derived soluble factors (100). For example, certain factors released
by melanoma cells induce changes in the phenotype of monocytes that
are similar to monocytic (Mo)-MDSCs typical in advanced melanoma
stages (101). Human MDSC
populations can be grouped into Mo-MDSCs
(CD11b+CD14+HLA−DR−CD15−)
or polymorphonuclear PMN-MDSCs
(CD11b+CD14−CD15+) (102). In cancer patients MDSCs are denoted
as CD33+HLA−DR− cells but in many
studies, isolated MDSCs display several phenotypes in relation to
the tumor sites. Thus, the phenotype
(CD14+HLA−DR−) was found to
prevail in melanoma patients and moreover, in patients receiving
ipilimumab there is no particular subset of MDSCs during melanoma
progression (103). The MDSC
phenotype
(CD11b+CD33+CD14+HLA−DR−/low)
was reported to be elevated in the peripheral blood mononuclear
cell (PBMC) population at melanoma onset and remain at comparable
levels throughout disease progression (104). High levels of MDSCs were also
associated with a lack of T lymphocyte clones specific for
melanoma-derived antigens (105),
providing a relevant clinical hint for correlation with patient
survival. Thus, upon analyzing the
CD14+HLA−DR−/low MDSC phenotype in
stage II/III melanoma patients, it was found that low levels of
these cells are associated with a tendency for an improved
disease-free survival (106). In
relation with other immune cell populations crucial for antitumor
defense, it has been reported that in the cancer milieu, PMN-MDSCs
exert suppressive activities on CD8+ T cells leading to
a reduced proliferation and inhibition of IFN-γ and IL-2 release by
T cells (107). Moreover, in
patients with advanced melanoma it was shown that both subsets of
MDSCs suppress CD8+ T cell proliferation (108). The MDSC accumulation in various
tumors has been associated with higher levels of IL-8 and several
cytokines (IL-10, IL-13, IL-6) (104,109).
Both monocytic and granulocytic MDSCs express IL-4Rα that are
associated with the suppressive activity of Mo-MDSCs in melanoma
(110). Anther study focusing on
various inflammatory markers suggested that levels of serum IL-1β,
IFN-γ and CXCL10 were significantly increased in advanced melanoma
and are direct correlated with increased MDSC and Treg populations;
moreover, disease progression was associated with an increased
serum concentration of IL-1β and CXCL10 (111). All these findings providing a strong
foundation in identifying high risk patient groups based on
circulatory profile of MDSCs, and developing therapeutic strategies
relying on MDSC inhibition or even depletion in melanoma
patients.
As in NMSC, in melanoma, inflammasomes have recently
been given special attention as being directly linked to
IL-1β-mediated tumorigenesis. In >200 melanoma tissue samples,
increased expression of NLRP1 was found in the cytoplasm. After
knocking down NLRP1 it was shown that several important molecules
were downregulated, such as caspase-1 activity, IL-1β production
and secretion and nuclear factor-κB activity that pointed out the
NLRP1 inflammasomes role in the metastasis process. Therefore, Zhai
et al showed that NLRP1 promotes melanoma tumorigenesis by
activating inflammasomes and by suppressing apoptotic pathways
(112).
Macrophages
Macrophages are myeloid cells (CD68+),
with an essential role in inflammation and host defense; these
cells are part of the innate immunity through their phagocytic
capacity and also join the adaptive immunity by activating other
immune cells via cytokine release. Macrophages may act in one of
two polarized state, namely classically activated M1 macrophages
and alternatively activated M2 macrophages. The M1 phenotype is
pro-inflammatory (mostly activated by IFN-γ) while M2 macrophages
release anti-inflammatory cytokines (IL-4, IL-10, TGF-β) and lean
towards an anti-inflammatory or immunosuppressive profile (113). Their activity in one or the other
polarized state in the tumor microenvironment affects melanoma
progression and prognosis. Histopathological studies have shown
that the macrophage polarization appears to be more connected to
the presence of lymphocytic infiltrate than to the thickness of the
melanoma lesions (114). A study
performed in 94 cases of stages I–IV skin melanoma with a long
follow-up duration depicts the correlation between M1/M2 phenotype
and disease progression. Thus, by CD68 double immunostaining with
MRP8–14 or iNOS (M1 phenotype) and with CD163 or CD204 (M2
phenotype) it was shown that in early melanoma stages, the M1
population was lower than the M2 population with a progressively
increase in M2 cells during tumor progression. Thus, M1 cells shift
to the M2 phenotype early in melanoma expansion, possibly induced
by an intratumoral increase in iNOS. The intratumoral accumulation
of both M1 and M2 is associated with poor prognostic indicators and
patient survival, favoring neoplastic growth and dissemination
(115).
In addition to polarized activation phenotypes,
macrophages may be classified via their activation status in
relation to the tumor. The inflammatory pattern of melanoma, with
important diagnostic, prognostic and therapeutic biomarker power,
resides in cell clones resembling tumor-associated macrophages. The
inflammatory phenotype of macrophages is linked to Melan A
expression loss from melanoma cells, a specific melanocyte marker
(116). Lack of specific Melan A
expression makes melanoma difficult to discriminate from tumors of
mesenchymal origin. Morphological changes leading to mesenchymal
outline and cellular de-differentiation are correlated with EMT and
tumor dissemination. Events that govern EMT are driven by the
inflammatory process and CD163+ macrophages which induce
E-cadherin and cell-to-cell adhesion loss as the final step of EMT
completion. Melan A-negative clones in tumor tissues are correlated
significantly with an increased inflammatory response elicited by
tumor-infiltrating CD163+ macrophages, the complete loss
of E-cadherin expression and a spindle-shaped morphology of tumor
cells, altogether as possible markers of poor differentiation and
tumor invasiveness (116).
Lymphoid cells in tumor infiltrate T
lymphocytes (cytotoxic, regulatory, helper)
In evaluating the circulatory cellular immune
pattern for a melanoma patient, a first step to be taken is testing
the absolute count of lymphocytes which further provides specific
data regarding circulating subpopulations. T lymphocyte subsets
represent the most extensively studied immune cell populations in
melanoma. These cells are both regulators and effectors of the
antitumor immune response, and CD8+ T cells are often
associated with a favorable clinical outcome (117). The pool of total CD3+ T
lymphocytes is a parameter that will change throughout the patient
follow-up only in advanced melanoma therefore calculating the
CD4+/CD8+ T cells ratio will indicate the
evolution of disease and will prognosticate the patient overall
survival (OS) irrespective to melanoma stage and the received
therapy. Other circulatory immune cells are found to be increased
only in stage III when the CD4+CD69+
phenotype indicates a lymph node-related antitumoral response;
there are reports that claim that the proportion of circulating
CD3+CD4+CD69+ cells evaluated
before therapy administration can be an independent prognostic
factor for OS (118). The influence
of different phenotypes upon survival could be explained by the
abundance of diverse T lymphocyte subsets, especially within
CD4+ Th lymphocytes. The different profiles of T cells
imprint different clinical responses according to their cytokine
repertoire. Thus, cytokines related to the Th1 subset are strongly
linked to positive clinical responses while the Treg
(CD4+CD25highCD127−Foxp3+)
cytokine panel is usually an indicator of a poor prognosis
(119,120). In metastatic melanoma a suppression
of Th1 growth and a Th2 driven chronic inflammatory state,
expressed in an increased Th2/Th1 ratio was reported. Moreover, if
the Th1 subtype is dominant in patients with completely resected
melanoma, in those with metastatic melanoma Th2 cells are the
dominant subset sustained by tumor-derived VEGF. High levels of
Th2-related cytokines (IL-4, IL-10, IL-13) and chemokines (CCL5,
CXCL10) have been detected in plasma of metastatic melanoma
patients (121–124).
As for the Treg phenotype, the proportion of
peripheral Tregs were found to increase with disease stage but no
correlation could be established with metastasis degree (125).
In less than 100 analyzed melanoma patients, it was
found that the IF from regressed and non-regressed tumor area has a
different distribution of inflammatory cells (126), mainly comprising T lymphocytes
(CD3+) (127,128). A significant association between
high pT level, CD3+ T cell frequency and ulceration was
identified. Non-ulcerated tumors have similar distributions of
CD3+ cells irrespective of pT level; ulceration cases
present frequent CD3+ cells in association with high pT
levels. Considering the overall favorable prognosis associated with
active tumor infiltrating leukocytes (TILs) (129,130),
the presence of abundant TILs within thick ulcerated tumors
represent a normal increasing of the IF as a physiologic reaction
to ulceration (126).
T CD8+ cells co-localize with major
histocompatibility complex (MHC) class I expressed on the tumor
cell surface in a pro-inflammatory setting, and with programmed
death-ligand 1 (PD-L1), a critical immune checkpoint that exhibits
an unfavorable prognostic impact in metastatic melanoma. In
addition, PD-L1 expression on circulating T cells predicts a worse
survival (131,132). By blocking these critical molecules,
a significant development in metastatic melanoma treatment has been
achieved. Monitoring the efficacy of such an endeavor is based also
on evaluating cellular immune population reinforced by the
revolutionary therapy. For instance, in advanced melanoma,
immunotherapy by pharmacologically blocking cytotoxic
T-lymphocyte-associated antigen (CTLA)-4 on Tregs, can be monitored
by an increase in circulating CD4+ and CD8+ T
cell lymphocytes (103,133). Immunotherapy endows the effector,
killing functions of T cells for the patient benefit. Thus,
CD8+ T cells exert tumoricidal functions through the
expression of granzyme B, and the activating markers CD25 and OX40,
especially if T cells are present in peritumoral areas or primary
tumor site when it associates with a better outcome (117).
The antitumoral activity of CD4+ T cells
was investigated by analysis of membrane CD134 expression (OX40)
and it was found that CD134 expression has been linked to a
favorable outcome. Moreover the level of CD134 expression on
CD4+ T cells in LNs append to primary melanomas declined
with more advanced stage and LN involvement suggesting an
immunosuppressive effect from tumor to LN location (134).
B lymphocytes
B cells frequently infiltrate the human tumor
milieu and the higher numbers of CD20+ tumor B cells
(TAB) are usually associated with a favorable prognosis. In human
cutaneous primary melanomas, this interrelation is still
controversial. Thus, in a recent study, the authors analyzed the
association of TAB numbers and OS assessing CD20
immunohistochemistry on archival non-metastasized and metastasized
primary melanoma tissues from 2 independent patient cohorts;
survival association was validated with RNA data from a third
independent cohort. The results of the study revealed the TAB
number as a prognostic biomarker in patients with tumors of >1
mm Breslow depth. Moreover, higher CD20/CD19 tumor mRNA levels were
found to be associated with a significantly better OS (135). This report is in line with previous
data that sustain the direct relation of infiltrating B cells and
better prognosis (136). Another B
cell phenotype, namely CD138+ plasmocytes are frequently
reported in areas of regressed melanoma but their expression was
regardless of regression or ulceration type (126). A subset of B-lymphocytes (B1 cells)
with in vivo pro-metastatic properties was also identified
and their presence was directly correlated with MCAM melanoma cell
adhesion molecule (MUC18) (also known as CD146) expression in
melanoma cells. Moreover, MUC18 expression can be therapeutically
triggered in human melanoma, hampering the tumor invasion process
(137). Reports related to advanced
melanoma stages show statistically higher circulating
CD19+ B lymphocytes with no increase in plasma level of
total or immunoglobulin (Ig) subclasses. There is a negative
correlation between the circulating B lymphocyte level and NK cells
in melanoma patients (126),
launching new insights in analysis of prognostic and predictive
significance of lymphoid immune cell interrelation in cutaneous
melanoma.
NK cells
NK cells are phenotypically defined as
CD3−CD56+ expressing the surface receptor
NKp46 (CD335) distinctive for this cell population. In addition,
human NK cells are subdivided into
CD16+CD56dim (prevailing in blood), and
CD16−CD56bright subtypes (138). The role of NK cells in melanoma
tumor inflammatory infiltration are currently not fully elucidated,
representing an actual research topic (139). Different NK cell phenotypes are
involved in organ specific susceptibility to melanoma metastasis.
In an experimental murine model it was reported that immature
CD27+CD11b− NK cells protect liver from
metastasis through a perforin-dependent cytotoxic mechanism against
tumor, while at the pulmonary level, mature subsets
CD27−CD11b− and
CD27−CD11b− are responsible for reducing
tumor burden (140).
CD56dimCD57+ activated cells exert their
functions in spite of Treg cell presence. During melanoma
progression, the
CD56dimCD57+/CD56brightCD57+
cell ratio increases and could be used as a prognostic marker
(141).
The NK cells discriminate between normal and
transformed cells sensing the insufficient level of MHC class I
molecules expressed on latest ones (142). There is a bidirectional interaction
between melanoma cells and NK cells as target recognition by NK
depends on the interplay between killer activating (KAR) and killer
inhibitory (KIR) receptors expressed on NK cells, and further by
signals delivered to the tumor cell. Activated NK cells secrete
factors (perforines, granzymes), express death mediating
biomolecules (FasL/CD95 and TRAIL) and produce various cytokines
(e.g., IFN-γ) that destroy the tumor target and also recruit other
immune cells to the tumor site (139,143).
An altered/decreased MHC class I expression on
tumor cells is an escape mechanism by which melanomas avoid
CD8+ T cell attack but facilitate NK cell-mediated
killing. This is the reason why melanoma is considered a model for
the study of NK cell-mediated tumor killing (144). Recent studies report that a high
percentage of melanoma cells hold ligands for NK activating
receptors (e.g., NKG2D and DNAM1), and ligands for natural
cytotoxicity receptors such as NKp30 (138). There is a process denominated as
‘melanoma immunoediting’ that leads to tumor escape from NK cell
attack by multiple mechanisms such as increased expression of
MHC-I, or downregulation of NK ligands especially in metastatic
sites; the same inhibitory action upon NK cells is exerted by
indole amine 2,3-dioxygenase (IDO) and PGE2 secreted by melanoma
cells (145). Melanoma cells
regulate NK lymphocytes via different cytokine/chemokine
repertoire; thus, it was reported that IL-18 secreted by tumor
cells upregulates PD-1 expression on NK cells (146). Also, it seems that melanoma
metastatic evolution is associated with an increased frequency of
peripheral NK cells expressing receptors for CXCL8, corroborated
with CXCL8 released by tumor cells (141,147).
Moreover, it has been proposed that anti-IL-18 antibodies in
combination with anti-PD-1 mAb (nivolumab) may avoid NK cell
inhibition by PD-1 (148).
Recently, it was shown that melanoma-infiltrated
lymph nodes contains twice as many NK cells compared with
tumor-free nodes, and a population of highly cytotoxic NK cells
(CD56dimKIR+CCR7+) with potential
prognostic value was identified in melanoma (141).
Existing cancer immunotherapies largely focus on
CD8+ T lymphocyte empowering, although NK cells are also
cytotoxic and effector cells in antitumor defense. Personalized
cancer therapies should integrate both CD8+ T cells in
acquired immunity and NK cells in innate immunity as a strong
weapon for precision targeting of tumors (149). NK cells are strongly accountable for
enlarging the immunotherapeutic arsenal in melanoma. Besides the
much studied immune checkpoint blockade of CTLA4, NK cells could be
tackled from this point of view as one of the major checkpoints in
NK cell activation is mediated by MHC class I specific KIR
receptors. Presently, two antibodies directed to NK cell checkpoint
blockade are under clinical development for melanoma therapy,
namely, lirilumab (anti-KIR in combination with ipilimumab) and
IPH2201 (anti-NKG2A) (139).
Some outlines regarding circulatory immune cells in
the melanoma inflammatory setting could be drawn and a first
conclusion is that there is no perfect match between circulating
immune cells and tumor-associated ones, a still non-deciphered
inconsistency (98). Furthermore,
circulating immune cell-specific phenotypes are finely linked to
the diagnosed stage of the melanoma; therefore, a single immune
cell population cannot depict accurately the disease evolution.
Hence circulatory immune cell subsets, displaying an activated
and/or a suppressor phenotype would give the physician a more
focused immune status of the patient for future personalized
disease management.
Inflammatory-related molecules in melanoma:
the pattern of inflammatory molecules in melanoma tissue
There is a clear immune suppressive environment
developed at the tumor site where several inflammatory cells and
molecules affect tumor development and invasiveness (32). Macrophages secrete IDO, an
immunosuppressive enzyme, that induces inhibition of T cell
proliferation due to tryptophan depletion and, moreover, IDO
recruits more Tregs into the tumor area. As a consequence,
TGF-β-secreting Tregs will induce suppression on the effector
couple CD4+/CD8+ diminishing the control on
tumor development. Tumor cells also secrete TGF-β, IL-10, VEGF and
PGE2 that induce DCs to release more TGF-β contributing to the
conversion of CD4+ T cells to a Treg phenotype and thus
augmenting the cellular immune suppression. By a concerted action,
a favorable microenvironment is created resulting in Treg
proliferation that hinders the cooperation of
CD4+/CD8+ T cells and obliterates the
antitumoral activity of cytotoxic cells. In addition, IDO is
proposed as a prognostic and follow-up marker in melanoma. Thus, in
a recent study IDO, lactate dehydrogenase (LDH) and S100B levels
were measured in 186 serum samples from patients in all melanoma
stages, at diagnosis and twice a year afterward. At diagnosis,
serum IDO levels were significantly higher in stages IB, II, III
and IV, whereas S100B levels were significantly higher in stages
III and IV; as expected, LDH levels were higher only in stage IV.
In relapsed patients, all three tested markers were found to be
significantly increased. Finally, OS was significantly longer in
patients with IDO levels below a certain cut-off value at diagnosis
(1.65 µM) than in those with higher IDO levels (91.3 vs. 71.0% at
36 months). These data indicate IDO as a potential useful serum
prognostic biomarker for melanoma (150).
Circulatory inflammatory marker
pattern in melanoma
The immune tolerance could rely on the initiation
of the chronic inflammatory phase; thus, tumor cells could avoid
the immune system because the pro-inflammatory status is diminished
and switched to immunosuppression. This inflammatory status
conversion is triggered by a wide variety of mediators. Thus, high
levels of circulating biomolecules associated with poor prognosis
in melanoma (TNFR2, TGF-α, TIMP1, CRP) were recently identified by
multiplex ELISA sandwich and proposed as being part of a valuable
formulation for prediction of OS (151,152).
Another study performed on stage II and III melanoma patients
receiving IFN-α2b treatment point that the combination of serum
TNF-α, soluble IL-2 receptor and β2 microglobulin could be robust
predictive markers of melanoma relapse in relation to treatment.
Increased serum levels of TNF-α seem to have a protective role
before and despite high toxicity after IFN-α2b treatment (153) but the prognostic value of TNF-α is
still a matter of debate (154). In
recent years new insights were gained regarding
pro-inflammatory/antitumor vs. anti-inflammatory/protumoral effects
of TNF-α. It seems that membrane bound TNF-α, rather than soluble
TNF-α, can activate MDSCs as an active part of tumor-related IF;
stimulated MDSCs will release a whole cascade of mediators (ARG1,
iNOS, NO, ROS, IL-10, TGF-β) that finally leads to a suppressed
immune response against tumor (155). In this orchestrated action beside
the soluble form of TNF it seems that membrane-bound TNFR2 is also
involved (156). Moreover, TNFR2 is
also expressed on a subset of Tregs sustaining the
anti-inflammatory condition and tumor tolerance (157).
Although groundbreaking progress has been made in
the last few years in terms of immunotherapies, the panel of
circulatory reliable/validated markers for monitoring melanoma
prognosis or staging still remains limited. Thus, LDH is the first
serum biomarker included in 2001 by AJCC to be used for staging,
prognosis and overall survival evaluation in melanoma stage IV
patients (158). Moreover, LDH
remains a clinically significant marker associated with response,
progression-free survival, melanoma-specific survival (MSS) and OS
in the new era of targeted and immunotherapies. At the 8th AJCC
edition (2017), between several key changes it was included that an
elevated LDH level no longer independently defines M1c disease
‘with or without M1a or M1b sites of disease’ (159). The serum S100 calcium binding
protein B (S100B) marker is also in process of validation and a
recent study on a large cohort of non-resectable stage IV melanoma
patients suggest S100B to be a better independent marker than LDH
for long-term survival prediction. This could be explained by the
non-specificity of the largely expressed LDH marker, released in
the systemic circulation in many inflammatory disorders associated
with cell lysis, as opposed to the more specific S100B, that is
secreted by cells originated from the neural crest, including
melanoma cells (160). Other recent
data claim that S100B serum levels correlate with tumor load,
response to treatment and might identify patients with increased
risk of disease relapse; very important, S100B may predict
prognosis independent to LDH, and could act as an early biomarker
of tumor recurrence (161). S100B
was also connected to inflammation as it interacts with the
activated leukocyte cell adhesion molecule (ALCAM) and mediates
NF-κB signaling (162).
There is a strong correlation between S100B and
melanoma inhibitory activity (MIA), a protein secreted by
chondrocytes and melanoma cells, association that matches an
unfavorable clinical evolution. Significant MIA increases were
found to occur as early as stage II, with a better specificity and
sensitivity when used together with S100B, correlating with immune
parameters and having the potential of being biomarkers for
prognosis and therapy monitoring (163). The increased MIA level in patients
with poor prognosis could be a potential indicator of a
pro-inflammatory status switching to a more anti-inflammatory and
immunosuppressive phase of the disease (164).
Acute phase reactant proteins (APRPs) are usually
produced by hepatocytes upon cytokine stimulation, and in a wide
variety of diseases, including melanoma, a prolonged inflammation
status leads to the persistence of APRP level changes. MALDI-TOF
mass spectrometric analysis identified serum amyloid A as a
valuable prognostic marker for all melanoma stages, with an
increased specificity and sensitivity for early stages in
combination with C reactive protein (CRP). These two acute phase
proteins may have great clinical significance in melanoma
considering also the cost efficiency of their in tandem testing
(165).
The extensive diversity of soluble mediators and
cells involved in the complex switch between acute and chronic
inflammation could provide novel early indicators regarding tumor
immunosuppressive status, induced by prolonged inflammation, and
offer new insights in early diagnosis, prognosis evaluation and
melanoma therapy monitoring.
Conclusion
Inflammation has a physiological important role as
its final goal is tissue damage healing. When this process develops
as chronic inflammation it triggers molecular and cellular networks
that generate an immunosuppressive millieu, that can drive skin
tumorigenesis. Various intrinsic and extrinsic factors can trigger
the chronic status of inflammation, but the major initiating
trigger in NMSC and melanoma skin cancers is photoaging. Disturbing
factors alter the normal interactions between resident skin cells
and immune cells that further alter tissue homeostasis.
It is extremely important in skin cancers to detect
the molecular pathways that switch from acute to chronic
inflammation because these pathways can be used as both predictors
and markers as future therapy targets. The inflammatory status of
the patients, whether related to the tissue and/or circulatory
markers can aid the overall prognosis of the patient.
Anti-inflammatory approaches, as already proven in NMSC are
bringing new therapeutical tools to classical therapies.
Unveiling chronic inflammation patterns related to
tumorigenesis can further direct/redirect the therapy choice and
furnish identification of new target molecules (166). There are still questions to be
answered, such as the link between sex steroid hormones and
inflammation and the involvement of inflammation pattern in
immunosuppressive mechanisms, but future research would further
elucidate the inflammatory complex networks that should be driven
towards antitumorigenic processes.
Acknowledgements
Not applicable.
Funding
This manuscript was supported by grant no.
PN-III-P1-1.2-PCCDI-2017-0341 and PN 18.21.02.02.
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
MN, CCo, CCa, CD, MS and SZ were responsible for
data gathering, analysis and contributed to writing the manuscript
and revising it critically for important intellectual content. All
authors read and approved the final version of manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ROS
|
reactive oxygen species
|
|
Th
|
T helper
|
|
Tregs
|
regulatory T cells
|
|
IL
|
interleukin
|
|
CTLs
|
cytotoxic T lymphocytes
|
|
LCs
|
Langerhans cells
|
|
GM-CSF
|
granulocyte-macrophage
colony-stimulating factor
|
|
TNF
|
tumor necrosis factor
|
|
MSH
|
melanocyte stimulating hormone
|
|
CXCL
|
chemokine (C-X-C motif) ligand
|
|
PGE2
|
prostaglandin E2
|
|
IFN
|
interferon
|
|
TGF
|
transforming growth factor
|
|
ASC
|
apoptosis-associated speck-like
protein
|
|
AIM2
|
absent in melanoma 2
|
|
NLR
|
NOD-like receptor
|
|
NLRP
|
NOD-like receptor with pyrin
|
|
NLRC
|
NOD-like receptor with caspase
recruitment domain
|
|
BCC
|
basal cell carcinoma
|
|
SCC
|
squamous cell carcinoma
|
|
CARD
|
caspase recruitment domain
|
|
MCs
|
mast cells
|
|
EMT
|
epithelial-mesenchymal transition
|
|
NMSC
|
non-melanoma skin cancer
|
|
HMGB1
|
high-mobility group box-1
|
|
RAGE
|
receptor for advanced glycation end
products
|
|
VDR
|
vitamin D receptor
|
|
PCNA
|
proliferating cell nuclear
antigen
|
|
iNOS
|
inducible nitric oxide synthase
|
|
COX-2
|
cyclooxygenase-2
|
|
NF-κB
|
nuclear factor-κB
|
|
ER
|
endoplasmic reticulum
|
|
TLR
|
Toll-like receptor
|
|
MAP kinase
|
mitogen-activated protein kinase
|
|
AP-1
|
activator protein-1
|
|
IRF
|
interferon regulatory factor
|
|
AK
|
actinic keratosis
|
|
MMPs
|
matrix metalloproteinases
|
|
ECM
|
extracellular matrix
|
|
RDEB
|
recessive dystrophic epidermolysis
bullosa
|
|
IKK2, IKKα
|
inhibitor of NF-κB kinase subunit
β
|
|
GFAP
|
glial fibrillary acidic protein
|
|
PAD2OE
|
MMTV-FLAG-hPAD2
|
|
STAT3
|
signal transducer and activator of
transcription 3
|
|
NK
|
natural killer
|
|
Smad2
|
SMAD family member 2
|
|
SMA
|
small worm phenotype
|
|
MAD
|
mothers against decapentaplegic
|
|
Nrf2
|
NF-E2-related factor 2
|
|
H-ras
|
V-Ha-ras Harvey rat sarcoma viral
oncogene homolog
|
|
IFE
|
inter-follicular epidermis
|
|
HF
|
hair follicle
|
|
K15-KRASG12D
|
mouse model with mutations in
oncogene KrasG12D (activation) and in Smad4 (deletion), in stem
cells expressing keratin 15 (K15+)
|
|
ETAR
|
endothelin A receptor
|
|
LGR5
|
leucine-rich repeat-containing
G-protein coupled receptor 5
|
|
Sox9
|
SRY-related HMG-box family of
transcription factors
|
|
Wnt
|
wingless-related integration site
|
|
PTK6
|
protein tyrosine kinase 6
|
|
WBCs
|
white blood cells
|
|
VDR
|
vitamin D receptor
|
|
SNP
|
single-nucleotide polymorphism
|
|
CYP24A1
|
cytochrome P450 family 24 subfamily A
member 1
|
|
uPAR
|
urokinase plasminogen activator
receptor
|
|
NSAID
|
non-steroidal anti-inflammatory
drugs
|
|
IF
|
immune infiltrate
|
|
TILs
|
tumor infiltrating lymphocytes
|
|
DCs
|
dendritic cells
|
|
SLN
|
sentinel lymph node
|
|
CDKN1A
|
cyclin-dependent kinase inhibitor
|
|
BMP7
|
bone morphogenetic protein 7
|
|
MDSCs
|
myeloid-derived suppressor cells
|
|
Mo-MDSCs
|
monocytic MDSCs
|
|
PMN-MDSCs
|
polymorphonuclear MDSCs
|
|
Met
|
HGF receptor
|
|
PBMCs
|
peripheral blood mononuclear
cells
|
|
CCL
|
chemokine C-C motif ligand
|
|
MHC
|
major histocompatibility complex
|
|
PD-L1
|
programmed death-ligand 1
|
|
CTLA
|
cytotoxic T lymphocyte antigen
|
|
TAB
|
tumor associated B cells
|
|
MUC18
|
MCAM melanoma cell adhesion molecule
or CD146
|
|
Ig
|
immunoglobulin
|
|
KARs
|
killer activating receptors
|
|
KIRs
|
killer inhibitory receptors
|
|
FasL
|
first apoptosis signal receptor
ligand
|
|
TRAIL
|
TNF-related apoptosis-inducing
ligand
|
|
IDO
|
indole amine 2,3-dioxygenase
|
|
VEGF
|
vascular endothelial growth
factor
|
|
LDH
|
lactate dehydrogenase
|
|
S100B
|
S100 calcium binding protein B
|
|
TIMP1
|
tissue inhibitor of metalloproteinase
1
|
|
CRP
|
C-reactive protein
|
|
OS
|
overall survival
|
|
MSS
|
melanoma-specific survival
|
|
M1c
|
metastasis to non-CNS visceral
sites
|
|
M1a
|
metastasis to skin, soft tissue
including muscle, and/or non-regional lymph node
|
|
M1b
|
metastasis to lung with or without
M1a sites of disease
|
|
ALCAM
|
activated leukocyte cell adhesion
molecule
|
|
MIA
|
melanoma inhibitory activity
|
|
APRP
|
acute phase reactant protein
|
|
MALDI-TOF
|
matrix-assisted laser
desorption/ionization-time of flight
|
References
|
1
|
Νeagu M: The immune system: A hidden
treasure for biomarker discovery in cutaneous melanoma. Adv Clin
Chem. 58:89–140. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Boda D, Docea AO, Calina D, Ilie MA,
Caruntu C, Zurac S, Neagu M, Constantin C, Branisteanu DE,
Voiculescu V, et al: Human papilloma virus: Apprehending the link
with carcinogenesis and unveiling new research avenues (Review).
Int J Oncol. 52:637–655. 2018.PubMed/NCBI
|
|
3
|
Calleja-Agius J, Brincat M and Borg M:
Skin connective tissue and ageing. Best Pract Res Clin Obstet
Gynaecol. 27:727–740. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fisher GJ, Kang S, Varani J, Bata-Csorgo
Z, Wan Y, Datta S and Voorhees JJ: Mechanisms of photoaging and
chronological skin aging. Arch Dermatol. 138:1462–1470. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martinon F, Burns K and Tschopp J: The
inflammasome: A molecular platform triggering activation of
inflammatory caspases and processing of proIL-β. Mol Cell.
10:417–426. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sollberger G, Strittmatter GE, Grossi S,
Garstkiewicz M, Auf dem Keller U, French LE and Beer HD: Caspase-1
activity is required for UVB-induced apoptosis of human
keratinocytes. J Invest Dermatol. 135:1395–1404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ortiz ML, Kumar V, Martner A, Mony S,
Donthireddy L, Condamine T, Seykora J, Knight SC, Malietzis G, Lee
GH, et al: Immature myeloid cells directly contribute to skin tumor
development by recruiting IL-17-producing CD4+ T cells.
J Exp Med. 212:351–367. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Awad F, Assrawi E, Louvrier C, Jumeau C,
Giurgea I, Amselem S and Karabina SA: Photoaging and skin cancer:
Is the inflammasome the missing link? Mech Ageing Dev. 172:131–137.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
DeNardo DG and Coussens LM: Inflammation
and breast cancer. Balancing immune response: Crosstalk between
adaptive and innate immune cells during breast cancer progression.
Breast Cancer Res. 9:2122007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Neagu M, Constantin C, Dumitrascu G, Lupu
A, Caruntu C, Boda D and Zurac S: Inflammation markers in cutaneous
melanoma-edgy biomarkers for prognosis. Discoveries (Craiova).
3:e382015. View Article : Google Scholar
|
|
12
|
Lin WW and Karin M: A cytokine-mediated
link between innate immunity, inflammation, and cancer. J Clin
Invest. 117:1175–1183. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Neagu M, Constantin C, Manda G and
Margaritescu I: Biomarkers of metastatic melanoma. Biomarkers Med.
3:71–89. 2009. View Article : Google Scholar
|
|
14
|
Mattii M, Lovászi M, Garzorz N, Atenhan A,
Quaranta M, Lauffer F, Konstantinow A, Küpper M, Zouboulis CC,
Kemeny L, et al: Sebocytes contribute to skin inflammation by
promoting the differentiation of T helper 17 cells. Br J Dermatol.
178:722–730. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Neagu M, Caruntu C, Constantin C, Boda D,
Zurac S, Spandidos DA and Tsatsakis AM: Chemically induced skin
carcinogenesis: Updates in experimental models (Review). Oncol Rep.
35:2516–2528. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Martinon F: Dangerous liaisons:
Mitochondrial DNA meets the NLRP3 inflammasome. Immunity.
36:313–315. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feldmeyer L, Keller M, Niklaus G, Hohl D,
Werner S and Beer HD: The inflammasome mediates UVB-induced
activation and secretion of interleukin-1beta by keratinocytes.
Curr Biol. 17:1140–1145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Broz P and Dixit VM: Inflammasomes:
Mechanism of assembly, regulation and signalling. Nat Rev Immunol.
16:407–420. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kostyuk V, Potapovich A, Stancato A, De
Luca C, Lulli D, Pastore S and Korkina L: Photo-oxidation products
of skin surface squalene mediate metabolic and inflammatory
responses to solar UV in human keratinocytes. PLoS One.
7:e444722012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oyewole AO and Birch-Machin MA: Sebum,
inflammasomes and the skin: Current concepts and future
perspective. Exp Dermatol. 24:651–654. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ahmad I, Muneer KM, Chang ME, Nasr HM,
Clay JM, Huang CC and Yusuf N: Ultraviolet radiation-induced
downregulation of SERCA2 mediates activation of NLRP3 inflammasome
in basal cell carcinoma. Photochem Photobiol. 93:1025–1033. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Latz E, Xiao TS and Stutz A: Activation
and regulation of the inflammasomes. Nat Rev Immunol. 13:397–411.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Penuela S, Gyenis L, Ablack A, Churko JM,
Berger AC, Litchfield DW, Lewis JD and Laird DW: Loss of pannexin 1
attenuates melanoma progression by reversion to a melanocytic
phenotype. J Biol Chem. 287:29184–29193. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhong FL, Mamaï O, Sborgi L, Boussofara L,
Hopkins R, Robinson K, Szeverényi I, Takeichi T, Balaji R, Lau A,
et al: Germline NLRP1 mutations cause skin inflammatory and
cancersusceptibility syndromes via inflammasome activation. Cell.
167:187–202. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Verma D, Bivik C, Farahani E, Synnerstad
I, Fredrikson M, Enerbäck C, Rosdahl I and Söderkvist P:
Inflammasome polymorphisms confer susceptibility to sporadic
malignant melanoma. Pigment Cell Melanoma Res. 25:506–513. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Da Silva WC, Oshiro TM, de Sá DC, Franco
DD, Festa Neto C and Pontillo A: Genotyping and differential
expression analysis of inflammasome genes in sporadic malignant
melanoma reveal novel contribution of CARD8, IL1B and IL18 in
melanoma susceptibility and progression. Cancer Genet. 209:474–480.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Okamoto M, Liu W, Luo Y, Tanaka A, Cai X,
Norris DA, Dinarello CA and Fujita M: Constitutively active
inflammasome in human melanoma cells mediating autoinflammation via
caspase-1 processing and secretion of interleukin-1β. J Biol Chem.
285:6477–6488. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu W, Luo Y, Dunn JH, Norris DA,
Dinarello CA and Fujita M: Dual role of apoptosis-associated
speck-like protein containing a CARD (ASC) in tumorigenesis of
human melanoma. J Invest Dermatol. 133:518–527. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Drexler SK, Bonsignore L, Masin M,
Tardivel A, Jackstadt R, Hermeking H, Schneider P, Gross O, Tschopp
J and Yazdi AS: Tissue-specific opposing functions of the
inflammasome adaptor ASC in the regulation of epithelial skin
carcinogenesis. Proc Natl Acad Sci USA. 109:18384–18389. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Uhlén M, Fagerberg L, Hallström BM,
Lindskog C, Oksvold P, Mardinoglu A, Sivertsson Å, Kampf C,
Sjöstedt E, Asplund A, et al: Proteomics. Tissue-based map of the
human proteome. Science. 347:12604192015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gonda TA, Tu S and Wang TC: Chronic
inflammation, the tumor microenvironment and carcinogenesis. Cell
Cycle. 8:2005–2013. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Neagu M, Constantin C and Tanase C:
Immune-related biomarkers for diagnosis/prognosis and therapy
monitoring of cutaneous melanoma. Expert Rev Mol Diagn. 10:897–919.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nedoszytko B, Sokołowska-Wojdyło M,
Ruckemann- Dziurdzińska K, Roszkiewicz J and Nowicki RJ: Chemokines
and cytokines network in the pathogenesis of the inflammatory skin
diseases: Atopic dermatitis, psoriasis and skin mastocytosis.
Postepy Dermatol Alergol. 31:84–91. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Justus CR, Leffler N, Ruiz-Echevarria M
and Yang LV: In vitro cell migration and invasion assays. J Vis
Exp. 1:882014.
|
|
35
|
Kong D, Li Y, Wang Z and Sarkar FH: Cancer
stem cells and epithelial-to-mesenchymal transition
(EMT)-phenotypic cells: Are they cousins or twins? Cancers (Basel).
3:716–729. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Plikus MV, Guerrero-Juarez CF, Treffeisen
E and Gay DL: Epigenetic control of skin and hair regeneration
after wounding. Exp Dermatol. 24:167–170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yan C, Grimm WA, Garner WL, Qin L, Travis
T, Tan N and Han YP: Epithelial to mesenchymal transition in human
skin wound healing is induced by tumor necrosis factor-α through
bone morphogenic protein-2. Am J Pathol. 176:2247–2258. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Leopold PL, Vincent J and Wang H: A
comparison of epithelial-to-mesenchymal transition and
re-epithelialization. Semin Cancer Biol. 22:471–483. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Egeblad M, Nakasone ES and Werb Z: Tumors
as organs: Complex tissues that interface with the entire organism.
Dev Cell. 18:884–901. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim Y and He YY: Ultraviolet
radiation-induced non-melanoma skin cancer: Regulation of DNA
damage repair and inflammation. Genes Dis. 1:188–198. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pondicherry A, Martin R, Meredith I, Rolfe
J, Emanuel P and Elwood M: The burden of non-melanoma skin cancers
in Auckland, New Zealand. Australas J Dermatol. 59:210–213. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Avrămoiu I, Petrescu IO, Ciurea ME, Bold
A, Siloşi I, ŢânŢu MM, Niculescu M, Anghel Savciu RE and Mogoantă
SŞ: Peritumoral inflammatory reaction in non-melanoma skin
cancers-histological and immunohistochemical study. Rom J Morphol
Embryol. 57:943–950. 2016.PubMed/NCBI
|
|
43
|
Nguyen AH, Detty SQ and Agrawal DK:
Clinical implications of high-mobility group box-1 (HMGB1) and the
receptor for advanced glycation end-products (RAGE) in cutaneous
malignancy: A systematic review. Anticancer Res. 37:1–7. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wu NL and Liu FT: The expression and
function of galectins in skin physiology and pathology. Exp
Dermatol. 27:217–226. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kapucuoglu N, Basak PY, Bircan S, Sert S
and Akkaya VB: Immunohistochemical galectin-3 expression in
non-melanoma skin cancers. Pathol Res Pract. 205:97–103. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nardinocchi L, Sonego G, Passarelli F,
Avitabile S, Scarponi C, Failla CM, Simoni S, Albanesi C and Cavani
A: Interleukin-17 and interleukin-22 promote tumor progression in
human nonmelanoma skin cancer. Eur J Immunol. 45:922–931. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Von Schuckmann LA, Law MH, Montgomery GW,
Green AC and Van Der Pols JC: Vitamin D pathway gene polymorphisms
and keratinocyte cancers: A nested case-control study and
meta-analysis. Anticancer Res. 36:2145–2152. 2016.PubMed/NCBI
|
|
48
|
Dusingize JC, Olsen CM, Pandeya NP,
Subramaniam P, Thompson BS, Neale RE, Green AC and Whiteman DC;
QSkin Study, : Cigarette smoking and the risks of basal cell
carcinoma and squamous cell carcinoma. J Invest Dermatol.
137:1700–1708. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chaudhary SC, Waseem M, Rana M, Xu H,
Kopelovich L, Elmets CA and Athar M: Naproxen inhibits UVB-induced
basal cell and squamous cell carcinoma development in
Ptch1+/−/SKH-1 hairless mice. Photochem Photobiol.
93:1016–1024. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Janda J, Burkett NB, Blohm-Mangone K,
Huang V, Curiel-Lewandrowski C, Alberts DS, Petricoin EF III,
Calvert VS, Einspahr J, Dong Z, et al: Resatorvid-based
pharmacological antagonism of cutaneous TLR4 blocks UV-induced
NF-κB and AP-1 signaling in keratinocytes and mouse skin. Photochem
Photobiol. 92:816–825. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Matei C, Tampa M, Caruntu C, Ion RM,
Georgescu SR, Dumitrascu GR, Constantin C and Neagu M: Protein
microarray for complex apoptosis monitoring of dysplastic oral
keratinocytes in experimental photodynamic therapy. Biol Res.
47:33–41. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Voiculescu V, Calenic B, Ghita M, Lupu M,
Caruntu A, Moraru L, Voiculescu S, Ion A, Greabu M, Ishkitiev N, et
al: From normal skin to squamous cell carcinoma: A quest for novel
biomarkers. Dis Markers. 2016:45174922016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Voiculescu VM, Caruntu C, Solomon I, Lupu
M, Ilie MA, Boda D, Constantin C and Neagu M: Squamous cell
carcinoma: Biomarkers and potential therapeutic targets. Human Skin
Cancers - Pathways, Mechanisms, Targets and Treatments. Blumenberg
M: IntechOpen; London: pp. 135–159. 2018
|
|
54
|
Paulitschke V, Gerner C, Hofstätter E,
Mohr T, Mayer RL, Pehamberger H and Kunstfeld R: Proteome profiling
of keratinocytes transforming to malignancy. Electrophoresis.
36:564–576. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Farshchian M, Nissinen L, Siljamäki E,
Riihilä P, Piipponen M, Kivisaari A, Kallajoki M, Grénman R,
Peltonen J, Peltonen S, et al: Tumor cell-specific AIM2 regulates
growth and invasion of cutaneous squamous cell carcinoma.
Oncotarget. 8:45825–45836. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Muller HK and Woods GM: Ultraviolet
radiation effects on the proteome of skin cells. Adv Exp Med Biol.
990:111–119. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
López-Camarillo C, Ocampo EA, Casamichana
ML, Pérez-Plasencia C, Alvarez-Sánchez E and Marchat LA: Protein
kinases and transcription factors activation in response to
UV-radiation of skin: Implications for carcinogenesis. Int J Mol
Sci. 13:142–172. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Bertrand-Vallery V, Boilan E, Ninane N,
Demazy C, Friguet B, Toussaint O, Poumay Y and Debacq-Chainiaux F:
Repeated exposures to UVB induce differentiation rather than
senescence of human keratinocytes lacking p16(INK-4A).
Biogerontology. 11:167–181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Föll MC, Fahrner M, Gretzmeier C, Thoma K,
Biniossek ML, Kiritsi D, Meiss F, Schilling O, Nyström A and Kern
JS: Identification of tissue damage, extracellular matrix
remodeling and bacterial challenge as common mechanisms associated
with high-risk cutaneous squamous cell carcinomas. Matrix Biol.
66:1–21. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Guerra L, Odorisio T, Zambruno G and
Castiglia D: Stromal microenvironment in type VII
collagen-deficient skin: The ground for squamous cell carcinoma
development. Matrix Biol. 63:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Meier K, Drexler SK, Eberle FC, Lefort K
and Yazdi AS: Silencing of ASC in cutaneous squamous cell
carcinoma. PLoS One. 11:e01647422016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kirkley KS, Walton KD, Duncan C and
Tjalkens RB: Spontaneous development of cutaneous squamous cell
carcinoma in mice with cell-specific deletion of inhibitor of κB
kinase 2. Comp Med. 67:407–415. 2017.PubMed/NCBI
|
|
63
|
Mohanan S, Horibata S, Anguish LJ, Mukai
C, Sams K, McElwee JL, McLean D, Yan A and Coonrod SA: PAD2
overexpression in transgenic mice augments malignancy and
tumor-associated inflammation in chemically initiated skin tumors.
Cell Tissue Res. 370:275–283. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Atmatzidis DH, Lambert WC and Lambert MW:
Langerhans cell: Exciting developments in health and disease. J Eur
Acad Dermatol Venereol. 31:1817–1824. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lin M, Sutherland DR, Horsfall W, Totty N,
Yeo E, Nayar R, Wu XF and Schuh AC: Cell surface antigen CD109 is a
novel member of the α(2) macroglobulin/C3, C4, C5 family of
thioester-containing proteins. Blood. 99:1683–1691. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Sunagawa M, Mii S, Enomoto A, Kato T,
Murakumo Y, Shiraki Y, Asai N, Asai M, Nagino M and Takahashi M:
Suppression of skin tumorigenesis in CD109-deficient mice.
Oncotarget. 7:82836–82850. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Varricchi G, Galdiero MR and Marone G,
Granata F, Borriello F and Marone G: Controversial role of mast
cells in skin cancers. Exp Dermatol. 26:11–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Latil M, Nassar D, Beck B, Boumahdi S,
Wang L, Brisebarre A, Dubois C, Nkusi E, Lenglez S, Checinska A, et
al: Cell-type-specific chromatin states differentially prime
squamous cell carcinoma tumor-initiating cells for epithelial to
mesenchymal transition. Cell Stem Cell. 20:191–204. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Dibra D, Mitra A, Newman M, Xia X, Keenan
C, Cutrera JJ, Mathis JM, Wang XJ, Myers J and Li S: IL27 controls
skin tumorigenesis via accumulation of ETAR-positive CD11b cells in
the pre-malignant skin. Oncotarget. 7:77138–77151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Ghita MA, Caruntu C, Rosca AE, Kaleshi H,
Caruntu A, Moraru L, Docea AO, Zurac S, Boda D, Neagu M, et al:
Reflectance confocal microscopy and dermoscopy for in vivo,
non-invasive skin imaging of superficial basal cell carcinoma.
Oncol Lett. 11:3019–3024. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Tilley C, Deep G and Agarwal R:
Chemopreventive opportunities to control basal cell carcinoma:
Current perspectives. Mol Carcinog. 54:688–697. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Jia J, Shi Y, Yan B, Xiao D, Lai W, Pan Y,
Jiang Y, Chen L, Mao C, Zhou J, et al: LGR5 expression is controled
by IKKα in basal cell carcinoma through activating STAT3 signaling
pathway. Oncotarget. 7:27280–27294. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Larsimont JC, Youssef KK, Sánchez-Danés A,
Sukumaran V, Defrance M, Delatte B, Liagre M, Baatsen P, Marine JC,
Lippens S, et al: Sox9 controls self-renewal of oncogene targeted
cells and links tumor initiation and invasion. Cell Stem Cell.
17:60–73. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chastkofsky MI, Bie W, Ball-Kell SM, He YY
and Tyner AL: Protein tyrosine kinase 6 regulates UVB-induced
signaling and tumorigenesis in mouse skin. J Invest Dermatol.
135:2492–2501. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Baykan H, Cihan YB and Ozyurt K: Roles of
white blood cells and subtypes as inflammatory markers in skin
cancer. Asian Pac J Cancer Prev. 16:2303–2306. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lupu M, Caruntu A, Caruntu C, Papagheorghe
LML, Ilie MA, Voiculescu V, Boda D, Constantin C, Tanase C, Sifaki
M, et al: Neuroendocrine factors: The missing link in non melanoma
skin cancer (Review). Oncol Rep. 38:1327–1340. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Lin Y, Chahal HS, Wu W, Cho HG, Ransohoff
KJ, Dai H, Tang JY, Sarin KY and Han J: Association between genetic
variation within vitamin D receptor-DNA binding sites and risk of
basal cell carcinoma. Int J Cancer. 140:2085–2091. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Κaukinen A, Siiskonen H, Pelkonen J and
Harvima IT: Immunoreactivity to CYP24A1, but not vitamin D
receptor, is increased in mast cells of keratinocyte skin cancers.
Eur J Dermatol. 27:590–598. 2017.PubMed/NCBI
|
|
79
|
Weidenbusch M, Rodler S, Song S, Romoli S,
Marschner JA, Kraft F, Holderied A, Kumar S, Mulay SR, Honarpisheh
M, et al: Gene expression profiling of the Notch-AhR-IL22 axis at
homeostasis and in response to tissue injury. Biosci Rep.
37:BSR201700992017.doi: 10.1042/BSR20170099. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Rubina KA, Sysoeva VY, Zagorujko EI,
Tsokolaeva ZI, Kurdina MI, Parfyonova YV and Tkachuk VA: Increased
expression of uPA, uPAR, and PAI-1 in psoriatic skin and in basal
cell carcinomas. Arch Dermatol Res. 309:433–442. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Kuznetsova EV, Snarskaya ES, Zavalishina
LE and Tkachenko SB: Immunohistochemical study of the specific
features of expression of matrix metalloproteinases 1, 9 in the
photoaged skin, the foci of actinic keratosis and basal cell
carcinoma. Arkh Patol. 78:17–22. 2016.(In Russian). View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Papakostas D and Stockfleth E: Topical
treatment of basal cell carcinoma with the immune response modifier
imiquimod. Future Oncol. 11:2985–2990. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Muranushi C, Olsen CM, Green AC and
Pandeya N: Can oral nonsteroidal antiinflammatory drugs play a role
in the prevention of basal cell carcinoma? A systematic review and
metaanalysis. J Am Acad Dermatol. 74:108–119.e1. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Barnes TA and Amir E: HYPE or HOPE: The
prognostic value of infiltrating immune cells in cancer. Br J
Cancer. 117:451–460. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Weiss SA, Han SW, Lui K, Tchack J, Shapiro
R, Berman R, Zhong J, Krogsgaard M, Osman I and Darvishian F:
Immunologic heterogeneity of tumor-infiltrating lymphocyte
composition in primary melanoma. Hum Pathol. 57:116–125. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Zurac S, Negroiu G, Andrei R, Petrescu S,
Tebeica T, Petre M, Neagu M, Constantin C, Chitu V, Salavastru C,
et al: Inflammatory infiltrate in melanoma with regression as
prognostic parameter. Virchows Arch. 463:1272013.
|
|
87
|
Klechevsky E: Functional diversity of
human dendritic cells. Adv Exp Med Biol. 850:43–54. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Hargadon KM: Strategies to improve the
efficacy of dendritic cell-based immunotherapy for melanoma. Front
Immunol. 8:15942017. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Van de Ven R, van den Hout MF, Lindenberg
JJ, Sluijter BJ, Van Leeuwen PA, Lougheed SM, Meijer S, Van den Tol
MP, Scheper RJ and De Gruijl TD: Characterization of four
conventional dendritic cell subsets in human skin-draining lymph
nodes in relation to T-cell activation. Blood. 118:2502–2510. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Deckers J, Hammad H and Hoste E:
Langerhans cells: Sensing the environment in health and disease.
Front Immunol. 9:932018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Klechevsky E, Morita R, Liu M, Cao Y,
Coquery S, Thompson-Snipes L, Briere F, Chaussabel D, Zurawski G,
Palucka AK, et al: Functional specializations of human epidermal
Langerhans cells and CD14+ dermal dendritic cells.
Immunity. 29:497–510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Price JG, Idoyaga J, Salmon H, Hogstad B,
Bigarella CL, Ghaffari S, Leboeuf M and Merad M: CDKN1A regulates
Langerhans cell survival and promotes Treg cell generation upon
exposure to ionizing irradiation. Nat Immunol. 16:1060–1068. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Hieronymus T, Zenke M, Baek JH and Seré K:
The clash of Langerhans cell homeostasis in skin: Should I stay or
should I go? Semin Cell Dev Biol. 41:30–38. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Yasmin N, Bauer T, Modak M, Wagner K,
Schuster C, Köffel R, Seyerl M, Stöckl J, Elbe-Bürger A, Graf D, et
al: Identification of bone morphogenetic protein 7 (BMP7) as an
instructive factor for human epidermal Langerhans cell
differentiation. J Exp Med. 210:2597–2610. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Arwert EN, Hoste E and Watt FM: Epithelial
stem cells, wound healing and cancer. Nat Rev Cancer. 12:170–180.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Baek JH, Birchmeier C, Zenke M and
Hieronymus T: The HGF receptor/Met tyrosine kinase is a key
regulator of dendritic cell migration in skin immunity. J Immunol.
189:1699–1707. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Yen JH, Khayrullina T and Ganea D:
PGE2-induced metalloproteinase-9 is essential for dendritic cell
migration. Blood. 111:260–270. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Haanen JB, Baars A, Gomez R, Weder P,
Smits M, De Gruijl TD, Von Blomberg BM, Bloemena E, Scheper RJ, Van
Ham SM, et al: Melanoma-specific tumor-infiltrating lymphocytes but
not circulating melanoma-specific T cells may predict survival in
resected advanced-stage melanoma patients. Cancer Immunol
Immunother. 55:451–458. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ladányi A, Kiss J, Somlai B, Gilde K,
Fejos Z, Mohos A, Gaudi I and Tímár J: Density of
DC-LAMP+ mature dendritic cells in combination with
activated T lymphocytes infiltrating primary cutaneous melanoma is
a strong independent prognostic factor. Cancer Immunol Immunother.
56:1459–1469. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Dai J, El Gazzar M, Li GY, Moorman JP and
Yao ZQ: Myeloid-derived suppressor cells: Paradoxical roles in
infection and immunity. J Innate Immun. 7:116–126. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Mao Y, Poschke I, Wennerberg E, Pico de
Coaña Y, Egyhazi Brage S, Schultz I, Hansson J, Masucci G,
Lundqvist A and Kiessling R: Melanoma-educated CD14+
cells acquire a myeloid-derived suppressor cell phenotype through
COX-2-dependent mechanisms. Cancer Res. 73:3877–3887. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Bronte V, Brandau S, Chen SH, Colombo MP,
Frey AB, Greten TF, Mandruzzato S, Murray PJ, Ochoa A,
Ostrand-Rosenberg S, et al: Recommendations for myeloid-derived
suppressor cell nomenclature and characterization standards. Nat
Commun. 7:121502016. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Tarhini AA, Edington H, Butterfield LH,
Lin Y, Shuai Y, Tawbi H, Sander C, Yin Y, Holtzman M, Johnson J, et
al: Immune monitoring of the circulation and the tumor
microenvironment in patients with regionally advanced melanoma
receiving neoadjuvant ipilimumab. PLoS One. 9:e877052014.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Rudolph BM, Loquai C, Gerwe A, Bacher N,
Steinbrink K, Grabbe S and Tuettenberg A: Increased frequencies of
CD11b+ CD33+ CD14+
HLA-DRlow myeloid-derived suppressor cells are an early
event in melanoma patients. Exp Dermatol. 23:202–204. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Martens A, Zelba H, Garbe C, Pawelec G and
Weide B: Monocytic myeloid-derived suppressor cells in advanced
melanoma patients: Indirect impact on prognosis through inhibition
of tumor-specific T-cell responses? OncoImmunology. 3:e278452014.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Filipazzi P, Pilla L, Mariani L, Patuzzo
R, Castelli C, Camisaschi C, Maurichi A, Cova A, Rigamonti G,
Giardino F, et al: Limited induction of tumor cross-reactive T
cells without a measurable clinical benefit in early melanoma
patients vaccinated with human leukocyte antigen class I-modified
peptides. Clin Cancer Res. 18:6485–6496. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Brandau S, Trellakis S, Bruderek K,
Schmaltz D, Steller G, Elian M, Suttmann H, Schenck M, Welling J,
Zabel P, et al: Myeloid-derived suppressor cells in the peripheral
blood of cancer patients contain a subset of immature neutrophils
with impaired migratory properties. J Leukoc Biol. 89:311–317.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Schilling B, Sucker A, Griewank K, Zhao F,
Weide B, Görgens A, Giebel B, Schadendorf D and Paschen A:
Vemurafenib reverses immunosuppression by myeloid derived
suppressor cells. Int J Cancer. 133:1653–1663. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Ene CD, Anghel AE, Neagu M and Nicolae I:
25-OH Vitamin D and interleukin-8 emerging biomarkers in cutaneous
melanoma development and progression. Mediators Inflam. 2015:1–8.
2015. View Article : Google Scholar
|
|
110
|
Mandruzzato S, Solito S, Falisi E,
Francescato S, Chiarion-Sileni V, Mocellin S, Zanon A, Rossi CR,
Nitti D, Bronte V, et al: IL4Rα+ myeloid-derived
suppressor cell expansion in cancer patients. J Immunol.
182:6562–6568. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Jiang H, Gebhardt C, Umansky L, Beckhove
P, Schulze TJ, Utikal J and Umansky V: Elevated chronic
inflammatory factors and myeloid-derived suppressor cells indicate
poor prognosis in advanced melanoma patients. Int J Cancer.
136:2352–2360. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Zhai Z, Liu W, Kaur M, Luo Y, Domenico J,
Samson JM, Shellman YG, Norris DA, Dinarello CA, Spritz RA, et al:
NLRP1 promotes tumor growth by enhancing inflammasome activation
and suppressing apoptosis in metastatic melanoma. Oncogene.
36:3820–3830. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Lopes RL, Borges TJ, Araújo JF, Pinho NG,
Bergamin LS, Battastini AM, Muraro SP, Souza AP, Zanin RF and
Bonorino C: Extracellular mycobacterial DnaK polarizes macrophages
to the M2-like phenotype. PLoS One. 9:e1134412014. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Scali E, Mignogna C, Di Vito A, Presta I,
Camastra C, Donato G and Bottoni U: Inflammation and macrophage
polarization in cutaneous melanoma: Histopathological and
immunohistochemical study. Int J Immunopathol Pharmacol.
29:715–719. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Falleni M, Savi F, Tosi D, Agape E, Cerri
A, Moneghini L and Bulfamante GP: M1 and M2 macrophages'
clinicopathological significance in cutaneous melanoma. Melanoma
Res. 27:200–210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Bønnelykke-Behrndtz ML, Steiniche T,
Damsgaard TE, Georgsen JB, Danielsen A, Bastholt L, Møller HJ,
Nørgaard PH and Schmidt H: MelanA-negative spindle-cell associated
melanoma, a distinct inflammatory phenotype correlated with dense
infiltration of CD163 macrophages and loss of E-cadherin. Melanoma
Res. 25:113–118. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Jacquelot N, Pitt JM, Enot DP, Roberti MP,
Duong CPM, Rusakiewicz S, Eggermont AM and Zitvogel L: Immune
biomarkers for prognosis and prediction of responses to immune
checkpoint blockade in cutaneous melanoma. OncoImmunology.
6:e12993032017. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Hernberg M, Mattila PS, Rissanen M,
Hansson J, Aamdal S, Bastholt L, Von der Maase H, Schmidt H,
Stierner U and Tarkkanen J: The prognostic role of blood lymphocyte
subset distribution in patients with resected high-risk primary or
regionally metastatic melanoma. J Immunother. 30:773–779. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Fridman WH, Galon J, Pagès F, Tartour E,
Sautès-Fridman C and Kroemer G: Prognostic and predictive impact of
intra- and peritumoral immune infiltrates. Cancer Res.
71:5601–5605. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Jacobs JFM, Nierkens S, Figdor CG, de
Vries IJM and Adema GJ: Regulatory T cells in melanoma: The final
hurdle towards effective immunotherapy? Lancet Oncol. 13:e32–e42.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Nevala WK, Vachon CM, Leontovich AA, Scott
CG, Thompson MA and Markovic SN: Melanoma study group of the Mayo
Clinic Cancer Center: Evidence of systemic Th2-driven chronic
inflammation in patients with metastatic melanoma. Clin Cancer Res.
15:1931–1939. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Umansky V and Sevko A: Melanoma-induced
immunosuppression and its neutralization. Semin Cancer Biol.
22:319–326. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Burkholder B, Huang RY, Burgess R, Luo S,
Jones VS, Zhang W, Lv ZQ, Gao CY, Wang BL, Zhang YM, et al:
Tumor-induced perturbations of cytokines and immune cell networks.
Biochim Biophys Acta. 1845:182–201. 2014.PubMed/NCBI
|
|
124
|
Neagu M, Constantin C and Longo C:
Chemokines in the melanoma metastasis biomarkers portrait. J
Immunoassay Immunochem. 36:559–566. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Wang W, Edington HD, Rao UN, Jukic DM,
Radfar A, Wang H and Kirkwood JM: Effects of high-dose IFNα2b on
regional lymph node metastases of human melanoma: Modulation of
STAT5, FOXP3, and IL-17. Clin Cancer Res. 14:8314–8320. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Neagu M, Constantin C and Zurac S: Immune
parameters in the prognosis and therapy monitoring of cutaneous
melanoma patients: Experience, role, and limitations. BioMed Res
Int. 2013:1079402013. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Nguyen LT, Yen PH, Nie J, Liadis N,
Ghazarian D, Al-Habeeb A, Easson A, Leong W, Lipa J, McCready D, et
al: Expansion and characterization of human melanoma
tumor-infiltrating lymphocytes (TILs). PLoS One. 5:e139402010.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Ηussein MR, Elsers DA, Fadel SA and Omar
AE: Immunohistological characterisation of tumour infiltrating
lymphocytes in melanocytic skin lesions. J Clin Pathol. 59:316–324.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Azimi F, Scolyer RA, Rumcheva P, Moncrieff
M, Murali R, McCarthy SW, Saw RP and Thompson JF:
Tumor-infiltrating lymphocyte grade is an independent predictor of
sentinel lymph node status and survival in patients with cutaneous
melanoma. J Clin Oncol. 30:2678–2683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Burton AL, Roach BA, Mays MP, Chen AF,
Ginter BA, Vierling AM, Scoggins CR, Martin RC, Stromberg AJ,
Hagendoorn L, et al: Prognostic significance of tumor infiltrating
lymphocytes in melanoma. Am Surg. 77:188–192. 2011.PubMed/NCBI
|
|
131
|
Kluger HM, Zito CR, Barr ML, Baine MK,
Chiang VL, Sznol M, Rimm DL, Chen L and Jilaveanu LB:
Characterization of PD-L1 expression and associated T-cell
infiltrates in metastatic melanoma samples from variable anatomic
sites. Clin Cancer Res. 21:3052–3060. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Ancuceanu R and Neagu M: Immune based
therapy for melanoma. Indian J Med Res. 143:135–144. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Ménard C, Ghiringhelli F, Roux S, Chaput
N, Mateus C, Grohmann U, Caillat-Zucman S, Zitvogel L and Robert C:
Ctla-4 blockade confers lymphocyte resistance to regulatory T-cells
in advanced melanoma: Surrogate marker of efficacy of tremelimumab?
Clin Cancer Res. 14:5242–5249. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Sarff M, Edwards D, Dhungel B, Wegmann KW,
Corless C, Weinberg AD and Vetto JT: OX40 (CD134) expression in
sentinel lymph nodes correlates with prognostic features of primary
melanomas. Am J Surg. 195:621–625. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Garg K, Maurer M, Griss J, Brüggen MC,
Wolf IH, Wagner C, Willi N, Mertz KD and Wagner SN:
Tumor-associated B cells in cutaneous primary melanoma and improved
clinical outcome. Hum Pathol. 54:157–164. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Chiou SH, Sheu BC, Chang WC, Huang SC and
Hong-Nerng H: Current concepts of tumor-infiltrating lymphocytes in
human malignancies. J Reprod Immunol. 67:35–50. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Staquicini FI, Tandle A, Libutti SK, Sun
J, Zigler M, Bar-Eli M, Aliperti F, Pérez EC, Gershenwald JE,
Mariano M, et al: A subset of host B-lymphocytes control melanoma
metastasis through a MCAM/MUC18-dependent interaction: Evidence
from mice and humans. Cancer Res. 68:8419–8428. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Sadozai H, Gruber T, Hunger RE and Schenk
M: Recent successes and future directions in immunotherapy of
cutaneous melanoma. Front Immunol. 8:16172017. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Tarazona R, Duran E and Solana R: Natural
killer cell recognition of melanoma: New clues for a more effective
immunotherapy. Front Immunol. 6:6492016. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Ballas ZK, Buchta CM, Rosean TR, Heusel JW
and Shey MR: Role of NK cell subsets in organ-specific murine
melanoma metastasis. PLoS One. 8:e655992013. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Ali TH, Pisanti S, Ciaglia E, Mortarini R,
Anichini A, Garofalo C, Tallerico R, Santinami M, Gulletta E, Ietto
C, et al: Enrichment of
CD56dimKIR+CD57+ highly cytotoxic
NK cells in tumour-infiltrated lymph nodes of melanoma patients.
Nat Commun. 5:56392014. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Nielsen N, Ødum N, Ursø B, Lanier LL and
Spee P: Cytotoxicity of CD56bright NK cells towards
autologous activated CD4+ T cells is mediated through
NKG2D, LFA-1 and TRAIL and dampened via CD94/NKG2A. PLoS One.
7:e319592012. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Morvan MG and Lanier LL: NK cells and
cancer: You can teach innate cells new tricks. Nat Rev Cancer.
16:7–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Mendez R, Aptsiauri N, Del Campo A, Maleno
I, Cabrera T, Ruiz-Cabello F, Garrido F and Garcia-Lora A: HLA and
melanoma: Multiple alterations in HLA class I and II expression in
human melanoma cell lines from ESTDAB cell bank. Cancer Immunol
Immunother. 58:1507–1515. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Baumeister SH, Freeman GJ, Dranoff G and
Sharpe AH: Coinhibitory pathways in immunotherapy for cancer. Annu
Rev Immunol. 34:539–573. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Terme M, Ullrich E, Aymeric L, Meinhardt
K, Desbois M, Delahaye N, Viaud S, Ryffel B, Yagita H, Kaplanski G,
et al: IL-18 induces PD-1-dependent immunosuppression in cancer.
Cancer Res. 71:5393–5399. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Singh S, Singh AP, Sharma B, Owen LB and
Singh RK: CXCL8 and its cognate receptors in melanoma progression
and metastasis. Future Oncol. 6:111–116. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Gras Navarro A, Bjorklund AT and Chekenya
M: Therapeutic potential and challenges of natural killer cells in
treatment of solid tumors. Front Immunol. 6:2022015. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Rosenberg J and Huang J: CD8+ T
cells and NK cells: Parallel and complementary soldiers of
immunotherapy. Curr Opin Chem Eng. 19:9–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
De Lecea MV, Palomares T, Al Kassam D,
Cavia M, Geh JLC, De Llano P, Muñiz P, Armesto D, Martinez-Indart L
and Alonso-Varona A: Indoleamine 2,3 dioxygenase as a prognostic
and follow-up marker in melanoma. A comparative study with LDH and
S100B. J Eur Acad Dermatol Venereol. 31:636–642. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Tarhini AA, Lin Y, Yeku O, LaFramboise WA,
Ashraf M, Sander C, Lee S and Kirkwood JM: A four-marker signature
of TNF-RII, TGF-α, TIMP-1 and CRP is prognostic of worse survival
in high-risk surgically resected melanoma. J Transl Med. 12:192014.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Zurac S, Neagu M, Constantin C, Cioplea M,
Nedelcu R, Bastian A, Popp C, Nichita L, Andrei R, Tebeica T, et
al: Variations in the expression of TIMP1, TIMP2 and TIMP3 in
cutaneous melanoma with regression and their possible function as
prognostic predictors. Oncol Lett. 11:3354–3360. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Hofmann MA, Kiecker F, Küchler I, Kors C
and Trefzer U: Serum TNF-α, B2M and sIL-2R levels are biological
correlates of outcome in adjuvant IFN-α2b treatment of patients
with melanoma. J Cancer Res Clin Oncol. 137:455–462. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Porter GA, Abdalla J, Lu M, Smith S,
Montgomery D, Grimm E, Ross MI, Mansfield PF, Gershenwald JE and
Lee JE: Significance of plasma cytokine levels in melanoma patients
with histologically negative sentinel lymph nodes. Ann Surg Oncol.
8:116–122. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Hu X, Li B, Li X, Zhao X, Wan L, Lin G, Yu
M, Wang J, Jiang X, Feng W, et al: Transmembrane TNF-α promotes
suppressive activities of myeloid-derived suppressor cells via
TNFR2. J Immunol. 192:1320–1331. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Polz J, Remke A, Weber S, Schmidt D,
Weber-Steffens D, Pietryga-Krieger A, Müller N, Ritter U, Mostböck
S and Männel DN: Myeloid suppressor cells require membrane TNFR2
expression for suppressive activity. Immun Inflamm Dis. 2:121–130.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Chen X and Oppenheim JJ: TNF-α: An
activator of CD4+FoxP3+TNFR2+
regulatory T cells. Curr Dir Autoimmun. 11:119–134. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Balch CM, Buzaid AC, Soong SJ, Atkins MB,
Cascinelli N, Coit DG, Fleming ID, Gershenwald JE, Houghton A Jr,
Kirkwood JM, et al: Final version of the American Joint Committee
on cancer staging system for cutaneous melanoma. J Clin Oncol.
19:3635–3648. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Gershenwald JE, Scolyer RA, Hess KR,
Sondak VK, Long GV, Ross MI, Lazar AJ, Faries MB, Kirkwood JM,
McArthur GA, et al: Melanoma staging: Evidence-based changes in the
American Joint Committee on cancer eighth edition cancer staging
manual. CA Cancer J Clin. 67:472–492. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Weide B, Richter S, Büttner P, Leiter U,
Forschner A, Bauer J, Held L, Eigentler TK, Meier F and Garbe C:
Serum S100B, lactate dehydrogenase and brain metastasis are
prognostic factors in patients with distant melanoma metastasis and
systemic therapy. PLoS One. 8:e816242013. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Karonidis A, Mantzourani M, Gogas H and
Tsoutsos D: Serum S100B levels correlate with stage, N status,
mitotic rate and disease outcome in melanoma patients independent
to LDH. J BUON. 22:1296–1302. 2017.PubMed/NCBI
|
|
162
|
Von Bauer R, Oikonomou D, Sulaj A,
Mohammed S, Hotz-Wagenblatt A, Gröne HJ, Arnold B, Falk C, Luethje
D, Erhardt A, et al: CD166/ALCAM mediates proinflammatory effects
of S100B in delayed type hypersensitivity. J Immunol. 191:369–377.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Dumitraşcu G, Constantin C, Manda G,
Hristescu S, Mărgaritescu I, Chiriţă D and Neagu M: Serum markers
in skin melanoma-preliminary study. Roum Arch Microbiol Immunol.
68:125–135. 2009.PubMed/NCBI
|
|
164
|
Schmidt J, Riechers A, Stoll R, Amann T,
Fink F, Spruss T, Gronwald W, König B, Hellerbrand C and Bosserhoff
AK: Targeting melanoma metastasis and immunosuppression with a new
mode of melanoma inhibitory activity (MIA) protein inhibition. PLoS
One. 7:e379412012. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Findeisen P, Zapatka M, Peccerella T,
Matzk H, Neumaier M, Schadendorf D and Ugurel S: Serum amyloid A as
a prognostic marker in melanoma identified by proteomic profiling.
J Clin Oncol. 27:2199–2208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Neagu M and Constantin C: Immune-therapy
in cutaneous melanoma-efficacy immune markers. Advancements in
Tumor Immunotherapy and Cancer Vaccines. Arnouk H: InTech; Rijeka,
Croatia: pp. 83–106. 2012
|