Introduction
Osteosarcoma originates from mesenchymal cells and
is the most common primary malignant bone tumor affecting children
and adolescents (1,2), accounting for ~5% of all pediatric
tumors (3,4). The 5-year survival rate (60–70%) of
patients with osteosarcoma has significantly improved in their
10-year follow-up, due to the implementation of combined treatment
with surgery and multi-agent chemotherapy (5,6). Cisplatin
is the most commonly used antitumor drug, but its therapeutic value
is uncertain due to chemoresistance (7). Therefore, further investigation on how
to reduce cisplatin resistance and improve the therapeutic effect
of cisplatin in osteosarcoma is required (8,9).
MicroRNAs (miRNAs, miRs) are short (~22 nucleotides)
endogenous, non-coding, single-stranded RNAs that
post-transcriptionally regulate the expression of their target
genes (10). miRNAs directly affect
the stability of their target mRNA or inhibit its translation by
binding to its 3′untranslated region (3′-UTR) (11,12).
miRNAs are involved in a variety of biological processes, including
cell survival, differentiation, proliferation, apoptosis,
autophagy, motility and metabolism (13,14).
Abnormal miRNA expression may exhibit tumor suppressor or oncogenic
effects, which influence the occurrence and progression of
malignant tumors (15,16). Several miRNAs have been confirmed to
be involved in osteosarcoma chemoresistance. Meng et al
(17) revealed that miR-140-5p
regulated autophagy-mediated osteosarcoma chemoresistance by
targeting high mobility group nucleosome binding domain 5.
Furthermore, Vanas et al (18)
demonstrated that miR-21 facilitated osteosarcoma cell
proliferation and decreased cisplatin sensitivity by targeting
sprouty RTK signaling antagonist 2. Additionally, Liu et al
(19) determined that miR-200c
suppressed cell proliferation and enhanced cisplatin sensitivity in
osteosarcoma cells by targeting serine/threonine kinase 2. These
studies provide evidence for the use of certain miRNAs as effective
predictive markers for cisplatin resistance in osteosarcoma.
p53 was the first tumor suppressor gene to be
identified and is mutated in ~50% of osteosarcomas (20). The absence of normal p53 function
serves an important role in tumor occurrence and progression, as
p53 protein induces cell cycle arrest, apoptosis or the senescence
of damaged or mutant cells to prevent their proliferation, which
may otherwise promote tumor occurrence and progression (21–23). Zhao
et al (24) demonstrated that
p53 overexpression increased chemosensitivity in
multidrug-resistant osteosarcoma cell lines and Wu et al
(25) revealed that p53 expression
was a useful prognostic biomarker for the prediction of survival in
patients with osteosarcoma. Previous studies have demonstrated that
specific miRNAs are involved in an additional p53-associated
mechanism of osteosarcoma suppression (26,27). He
et al (28) determined that
miR-34 suppressed osteosarcoma cell proliferation and invasion by
targeting p53, whilst Zhang et al (29) determined that miR-29 induced
osteosarcoma cell apoptosis via the activation of p53.
miR-504 has been associated with several types of
malignant tumor, particularly in association with cell
proliferation and apoptosis, with a previous study demonstrating
that miR-504 is overexpressed in osteosarcoma (30). However, to the best of our knowledge,
the specific role and mechanism of miR-504 in modulating cisplatin
resistance in osteosarcoma cells is yet to be elucidated. The
current study therefore aimed to clarify the role and mechanism of
miR-504 in the modulation of cisplatin resistance in human
osteosarcoma cells. The results of the present study verified that
miR-504 promoted cell proliferation and contributed to
cisplatin-induced apoptosis and cell cycle arrest in MG63
osteosarcoma cells, by targeting p53. These results indicate that
miR-504 may be a novel target for the reduction of cisplatin
resistance.
Materials and methods
Tissue samples, cell culture,
lentivirus infection and cell treatment
Osteosarcoma tissues and adjacent normal tissues
(n=10 pairs; 2–5 cm apart) were collected between September 2016
and May 2017 during routine therapeutic surgery at the Department
of Orthopaedics at the First Affiliated Hospital of Wenzhou Medical
University (Wenzhou, China). The human osteosarcoma tissues and
pair-matched adjacent normal tissues were subsequently used to
compare the expression of miR-504 by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR). The
role of miR-504 in osteosarcoma progression was subsequently
analyzed in vitro by using MG63 cells. A total of 10
patients (range, 12–22 years of age), 4 male and 6 female,
participated in the present study. Inclusion criteria were as
follows: Patients with a pathological diagnosis of osteosarcoma,
original site of osteosarcoma was the long bone of limbs, patients
receiving surgical treatment and follow-up time ≥12 months. The
exclusion criteria were as follows: Pathological diagnosis of
non-osteosarcoma, original site of osteosarcoma was not the long
bone of limbs, patient did not receive surgical treatment and
follow-up time was <12 months.) Immediately following surgery,
tumor tissues were stored at −80°C until further use.
The human osteosarcoma cell line MG63 and human
fetal osteoblastic cell line hFOB1.19 were obtained from ZQXZ
Biotech co., Ltd. (Shanghai, China) and cultured in high-glucose
Dulbecco's Modified Eagle's medium (DMEM-HG) and DMEM Nutrient
Mixture F-12 medium (DMEM-F12; both Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), respectively. MG63 and
hFOB1.19 cells were cultured for ~36 h at 37°C in a humidified
incubator supplemented with 5% CO2 and harvested using
0.25% trypsin/0.02% EDTA solution (Gibco; Thermo Fisher Scientific,
Inc.) once the adherent cells had reached a confluence of 80%.
pGCMV-hsa-miR-504-up (miR-504) and negative control (NC or miR-NC;
green fluorescent protein-labeled empty vector) lentiviruses were
provided by Shanghai GeneChem Co., Ltd. (Shanghai, China).
Osteosarcoma cells were then seeded in 6-well plates
(4×104/well, 200 µl/well), grown to a confluence of
30–50% (~5×104/well) and then infected with miR-504 or
the miR-NC lentivirus, where each sample contained 1 µl of
polybrene (5 µg/ml) and 25 µl of lentivirus (1×108
Tube/ml) at a final multiplicity of infection of 50 (based on a
preliminary study) for 96 h (31,32). The
efficiency of miR-504 was detected using a RT-qPCR assay as
subsequently performed. The osteosarcoma cells were then divided
into three groups: A normal group cultured without additional
handling, an miR-NC group transfected with the NC lentivirus and an
miR-504 group transfected with the miR-504 lentivirus.
RNA isolation and RT-qPCR
Total RNA was extracted from the two cells, MG63 and
hFOB1.19 using TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.).
mRNA was reverse transcribed at 25°C for 30 min, 42°C for 30 min
and 85°C for 5 min. RT-qPCR was performed using a Hairpin-it™ miRNA
RT-PCR Quantification kit (Shanghai GenePharma Co., Ltd., shanghai,
China), according to the manufacturer's protocols. The kit of RT
contains 5× MMLV RT buffer, dNTP (10 mM), miR-RT primers (1 µM),
RNasin (40 U/µl), MMLV Reverse Transcriptase (200 U/µl), and RNase
Free H2O. The kit of qPCR includes 2XReal-time PCR
Master Mix, miR-504 specific Primer set (10 µM), miR-504 specific
Probe (10 µM), Taq DNA polymerase (5 U/µl) and sterilized
H2O. Thermocycling conditions of miR-504 qPCR were as
follows: 1 cycle at 95°C for 3 min (pre-degeneration), 40 cycles at
95°C for 12 sec, and fluorescent signal acquisition at 62°C for 40
sec. miR-504 and U6 small nuclear (sn)RNA PCR reverse primers were
synthesized by Shanghai GenePharma Co., Ltd. The forward and
reverse primer sequences were as follows: miR-504 forward,
5′-CCAGCAAGACCCTGGTCTG-3′ and reverse, 5′-CAGAGCAGGGTCCGAGGTA-3′;
U6 snRNA forward, 5′-ATTGGAACGATACAGAGAAGATT-3′ and reverse,
5′-GTTTAAGCACTTCGCAAGG-3′. miR-504 expression data were normalized
to U6 snRNA. Specific products were detected and analyzed using a
Roche LightCycler 480 Detection System (Roche Diagnostics, Basel,
Switzerland). The 2−ΔΔCq method (33) was used to calculate the relative
expression of miR-504.
Cell proliferation assay
Cell proliferation was evaluated via an MTT assay.
MG63 cells (4×104/well) infected with miR-504 and miR-NC
were seeded in 96-well plates and cell proliferation was measured
at 6, 12, 24, 48, 72 and 96 h. 150 µl of MTT was added to each well
and the plate was incubated at 37°C for a further 4 h. Dimethyl
sulfoxide was then added to dissolve the sediment. Absorbance was
measured at 490 nm using a Spectra Max Plus 384 microplate reader
(Molecular Devices LLC, Sunnyvale, CA, USA). Cell proliferation was
also detected via an EdU assay, using an EdU cell proliferation
detection kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China).
The cells (4×104/well) were seeded in 6-well plates and
cultured at 37°C for 48 h, and then incubated with DMEM containing
EdU (Nanjing KeyGen Biotech Co., Ltd.). They were subsequently
fixed in 4% paraformaldehyde at 4°C for 30 min, washed twice with
PBS, reacted with Apollo 643 and dissolved in Apollo reaction
buffer (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China). The
cells were subsequently stained with DAPI at 37°C for 3 min and
observed under an inverted phase contrast fluorescence microscope
(magnification, ×400; Carl Zeiss AG, Oberkochen, Germany). Cells
stained red were considered EdU-positive.
Cell viability assay
Cell viability was also evaluated via an MTT assay.
MG63 cells from the normal group (4×104/well) were
seeded in 96-well plates and pretreated with different
concentrations (0, 2.5, 5, 7.5, 10, 12.5, 15, 17.5 and 20 µg/ml) of
cisplatin (Qilu Pharmaceutical Co., Ltd., Jinan, China) for 24 and
48 h. MTT solution was then added to each well and the plate was
incubated at 37°C for 4 h. Dimethyl sulfoxide was added to dissolve
the sediment. Absorbance was measured at 490 nm using a microplate
reader. An appropriate concentration of cisplatin (~10 µg/ml;
IC50, half maximal inhibitory concentration) was then
selected to treat the different groups for 24, 48 and 72 h,
respectively, using the aforementioned method.
Cell apoptosis detection
Cell apoptosis was assessed using an Annexin V-APC/7
aminoactinomycin D (7AAD) apoptosis detection kit (Nanjing KeyGen
Biotech Co., Ltd.). MG63 cells were harvested, washed twice with
PBS, resuspended in binding buffer contained in the aforementioned
kit, stained with Annexin V-APC and 7AAD () at 37°C for 10 min, and
analyzed using a flow cytometer FACSCalibur system and CellQuest
5.1 software (BD Biosciences, San Jose, CA, USA). Annexin
V-APC+/7AAD− cells were considered as early
apoptotic cells, while Annexin V-APC+/7AAD+
cells were considered as late apoptotic or necrotic cells.
Morphology of apoptotic cells
Apoptotic MG63 cell morphology was observed directly
using an inverted phase contrast (fluorescence; magnification, ×100
and ×400). Cells (6×104/well) were cultured in 6- and
24-well plates at 37°C for 24 h, and apoptosis was induced by
cisplatin (10 µg/ml). Cells in the 6-well plate were observed
directly under an inverted phase contrast microscope
(magnification, ×100). Cells in 24-well plate were washed twice
with PBS, fixed in 4% paraformaldehyde at 4°C for 30 min, stained
with Hoechst 33258 staining solution (Beyotime Institute of
Biotechnology, Shanghai, China) at 37°C for 10 min and washed twice
with PBS. A total of 20 µl of Anti-fading solution (Beyotime
Institute of Biotechnology) was then dropped onto the cells and
they were imaged under an inverted phase contrast fluorescence
microscope (magnification, ×400).
Cell cycle analysis
The cell cycle was analyzed using a DNA content
quantitation (cell cycle) detection kit (Nanjing KeyGen Biotech
Co., Ltd.). MG63 cells were harvested, washed twice with PBS and
fixed in 70% ethanol overnight at 4°C. The following day, the cells
were centrifuged (800 × g, 4°C, 5 min), washed twice with PBS,
stained with propidium iodide and RNase A (Nanjing KeyGen Biotech
Co., Ltd.), and analyzed using a flow cytometer FACSCalibur system
and CellQuest 5.1 software (BD Biosciences).
Target mRNA gene prediction
The TargetScanHuman database (Agarwal V et
al.; http://www.targetScan.org;
TargetScanHuman Release 7.1 software) (34) was used to predict the potential target
gene of miR-504.
Dual luciferase reporter assay
Briefly, 293 cells (1×104/well) (35) were seeded in 96-well plates and
luciferase activities were measured using a dual luciferase
reporter assay, the Firefly and Renilla luciferase assay kit,
(Biotium, Inc., Freemont, CA, USA), according to the manufacturer's
protocols (36). Cells were
co-transfected with pGL3-miR-504 (miR-NC)-3′-UTR-wild type (wt)-p53
or pGL3-miR-504(miR-NC)-3′-UTR-mutant (mut)-p53 plasmid (Promega
Coorporation, Madison, WI, USA) using Lipofectamine®
2000. Following transfection for 48 h, Firefly and Renilla
luciferase activities were measured using a dual luciferase
reporter assay system (Biotium, Inc., Freemont, CA, USA).
Renilla luciferase activity was used as an internal control
for the evaluation of transfection efficiency.
Western blot analysis
Protein expression was evaluated via western
blotting. MG63 cells were lysed in radioimmunoprecipitation assay
lysis buffer (Beyotime Institute of Biotechnology). Protein
concentrations were measured by bincinchoninic acid assay (CW
Biotechnology, Beijing, China). Total protein (30 µg) was separated
using sodium dodecyl sulfate-polyacrylamide gel electrophoresis on
a 12% running gel and 5% stacking gel, and then transferred to
polyvinylidene fluoride membranes (0.45 µm; Merck KGaA, Darmstadt,
Germany). Membranes were subsequently blocked with 5% non-fat milk
in Tris-buffered saline with Tween-20 (TBS-T), containing 5% bovine
serum albumin (BD Biosciences) at 4°C overnight and incubated with
the following primary antibodies: Rabbit antibodies against p53
(dilution, 1:1,000; cat. no. 2527); Bcl-2-associated X (Bax;
dilution, 1:1,000; cat. no. 5023); B cell lymphoma-2 (Bcl-2;
dilution, 1:1,000; cat. no. 4223); caspase-3 (dilution, 1:1,000;
cat. no. 9665); p21 (dilution, 1:1,000; cat. no. 2947); cyclin D1
(dilution, 1:1,000; cat. no. 2978) and GAPDH (dilution, 1:1,000;
cat. no. 5174); mouse antibody to β-actin (dilution, 1:1,000; cat.
no. 3700) all purchased from Cell Signaling Technology, Inc.,
(Danvers, MA, USA). They were subsequently washed three times with
TBS-T and incubated with horseradish peroxidase-conjugated
secondary antibodies (goat-anti-rabbit; dilution, 1:5,000; cat. no.
CW0234; and goat-anti-mouse; dilution, 1:5,000; cat. no. CW0108);
CW Biotechnology) in TBS-T at room temperature for 2 h. All
proteins were visualized using an enhanced chemiluminescence system
(Thermo Fisher Scientific, Inc.).
Statistical analysis
Significant differences between groups were
evaluated using one-way analysis of variance, followed by
Student-Newman-Keuls test. All of the statistical analyses were
carried out using SPSS 18.0 software (SPSS, Inc., Chicago, IL,
USA). Graphs were created using GraphPad Prism 6 software (GraphPad
Software, Inc., La Jolla, CA, USA). All data were presented as mean
± standard error of the mean and three independent experiments were
analyzed. P<0.05 was considered to indicate a statistically
significant difference.
Results
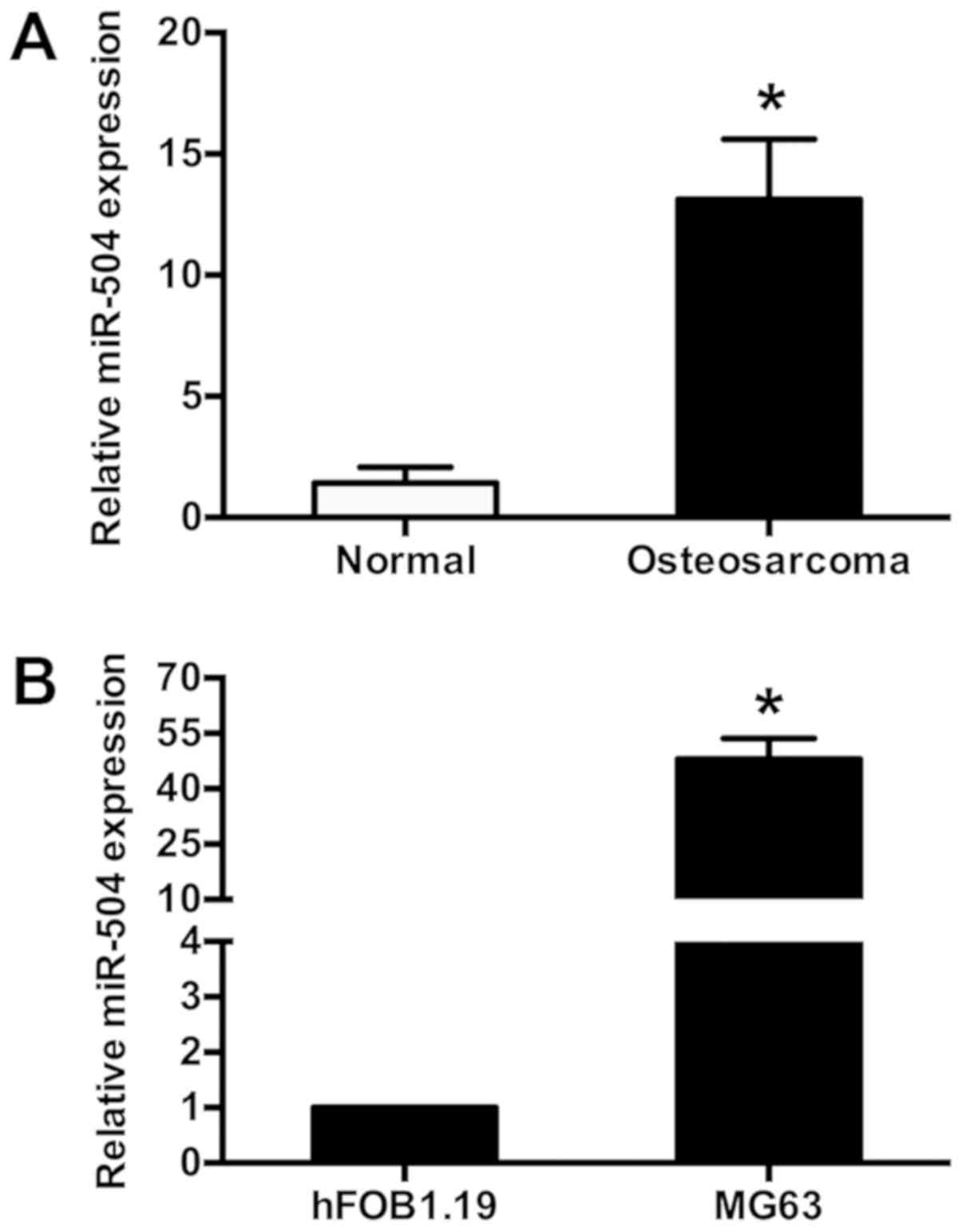
miR-504 expression is increased in
human osteosarcoma tissues and MG63 osteosarcoma cells
Aberrant miR-504 expression has been reported in
certain types of human tumor (20).
The current study assessed miR-504 expression in human osteosarcoma
tissues and pair-matched adjacent normal tissues (10 pairs) using
RT-qPCR. miR-504 expression was significantly increased in
osteosarcoma tissues compared with normal adjacent tissues
(P<0.05; Fig. 1A). miR-504
expression was also determined in MG63 osteosarcoma cells using
RT-qPCR, with hFOB1.19 osteoblastic cells as controls. It was
demonstrated that miR-504 expression was significantly increased in
MG63 cells compared with hFOB1.19 cells (P<0.05; Fig. 1B). These results indicated that
miR-504 expression levels were increased in human osteosarcoma
tissues and osteosarcoma cells.
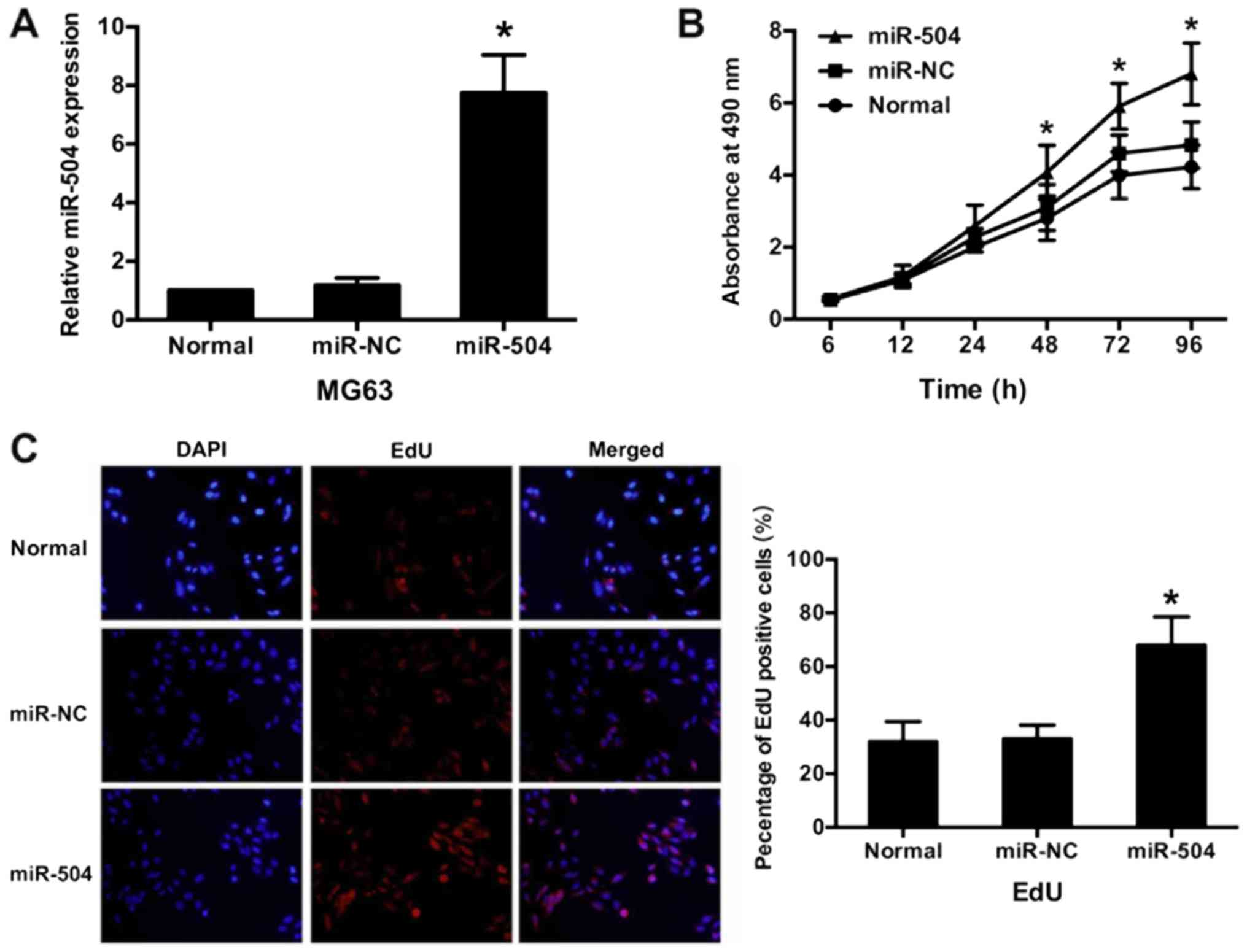
miR-504 lentivirus successfully
infects MG63 cells
To clarify the potential role of miR-504 in
osteosarcoma progression, miR-504 was overexpressed in MG63 cells
via infection with a miR-504 lentivirus. The results demonstrated
that miR-504 expression was 7.75±1.29-fold higher in the infected
cells compared with the normal cells, as determined by RT-qPCR
(P<0.05; Fig. 2A). There was no
significant difference in miR-504 expression between the miR-NC and
normal group. These results indicated that the miR-504 lentivirus
was successfully infected into MG63 cells.
miR-504 promotes cell
proliferation
The role of miR-504 in MG63 cell proliferation was
assessed via an MTT assay. Cell proliferation was compared between
the three groups from 6 to 96 h. The results demonstrated that
miR-504 significantly increased MG63 cell proliferation (from 48 h)
compared with the normal group (P<0.05; Fig. 2B). The greatest increase in
proliferation rate was determined at 96 h in the miR-504 group,
when compared with the normal group (1.61±0.18). However, no
significant differences were identified between the miR-NC and the
normal group (Fig. 2B). These results
were confirmed via an EdU assay, which demonstrated that
EdU-incorporation was significantly increased in the miR-504 group
compared with the normal group (P<0.05; Fig. 2C). However, there were no significant
differences between the miR-NC and normal group. These results
confirmed that miR-504 promotes MG63 cell proliferation.
Cisplatin induces cell apoptosis and
suppresses miR-504 expression in MG63 cells
While Cisplatin has been identified as an effective
chemotherapeutic drug for osteosarcoma (37), resistance to the drug remains a major
challenge. The current study therefore assessed the
growth-inhibitory effects of cisplatin in MG63 cells via an MTT
assay. MG63 cells were treated with different concentrations of
cisplatin for 24 and 48 h. It was determined that cell viability
was negatively associated with cisplatin concentration (from 2.5–20
µg/ml), with an IC50 value of ~10 µg/ml at 48 h
(P<0.05 and P<0.01 vs. normal group; Fig. 3A). A cisplatin concentration of 10
µg/ml was therefore selected for subsequent experiments. The effect
of cisplatin on miR-504 expression in MG63 cells was assessed using
RT-qPCR. Exposure of MG63 cells to 10 µg/ml cisplatin significantly
suppressed miR-504 expression compared with the normal group
(P<0.05; Fig. 3B). These results
indicated that cisplatin induced cell apoptosis and suppresses
miR-504 expression in MG63 cells.
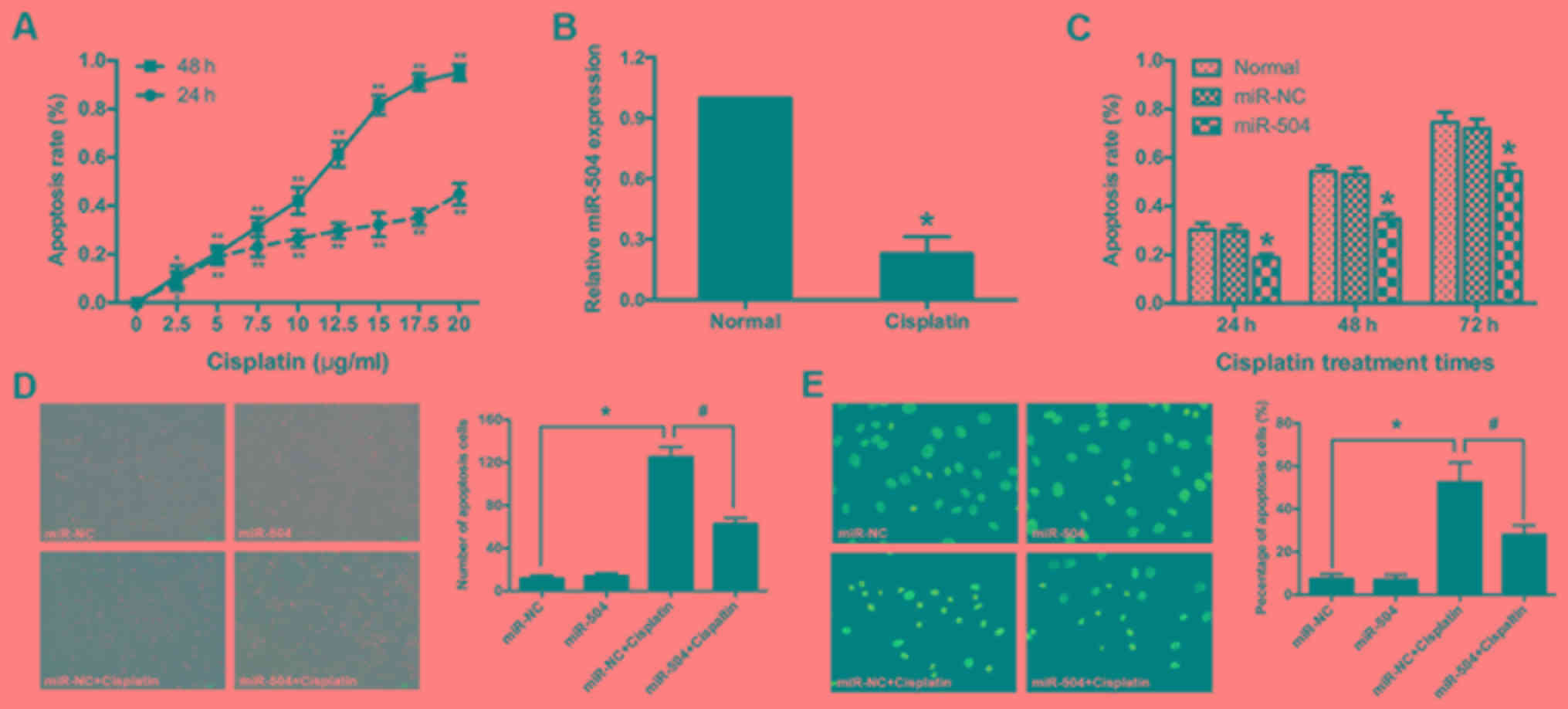 | Figure 3.Cisplatin promoted MG63 cell
apoptosis by decreasing miR-504. (A) MG63 cells were exposed to
different concentrations (0, 2.5, 5, 7.5, 10, 12.5, 15, 17.5 and 20
µl) of cisplatin for 24 or 48 h and an MTT assay was performed to
determine apoptosis rate. At a concentration of 10 µg/ml ~50% of
cells died following 48 h. *P<0.05 and **P<0.01 vs. the
untreated MG63 cells, at 24 and 48 h, respectively. (B) Cisplatin
treatment significantly suppressed miR-504 expression in MG63
cells, as determined using a reverse transcription-quantitative
polymerase chain reaction assay. *P<0.05 vs. the normal group.
(C) MG63 cells were exposed to 10 µg/ml cisplatin for 24, 48 or 72
h and an MTT assay was performed to assess apoptosis rate.
*P<0.05 vs. the normal group. (D) The morphological appearance
of MG63 cells exposed to 10 µg/ml cisplatin for 48 h was observed
under an inverted phase-contrast microscope (magnification, ×100).
*P<0.05 vs. the miR-NC group; #P<0.05 vs. the
miR-NC+cisplatin group. (E) The nuclear morphology of MG63 cells
exposed to 10 µg/ml cisplatin for 48 h was observed by Hoechst
33258 staining and inverted phase contrast microscopy
(magnification, ×400). Each, n=3; *P<0.05 vs. the miR-NC group;
#P<0.05 vs. the miR-NC+cisplatin group. miR, micro
RNA; NC, negative control. |
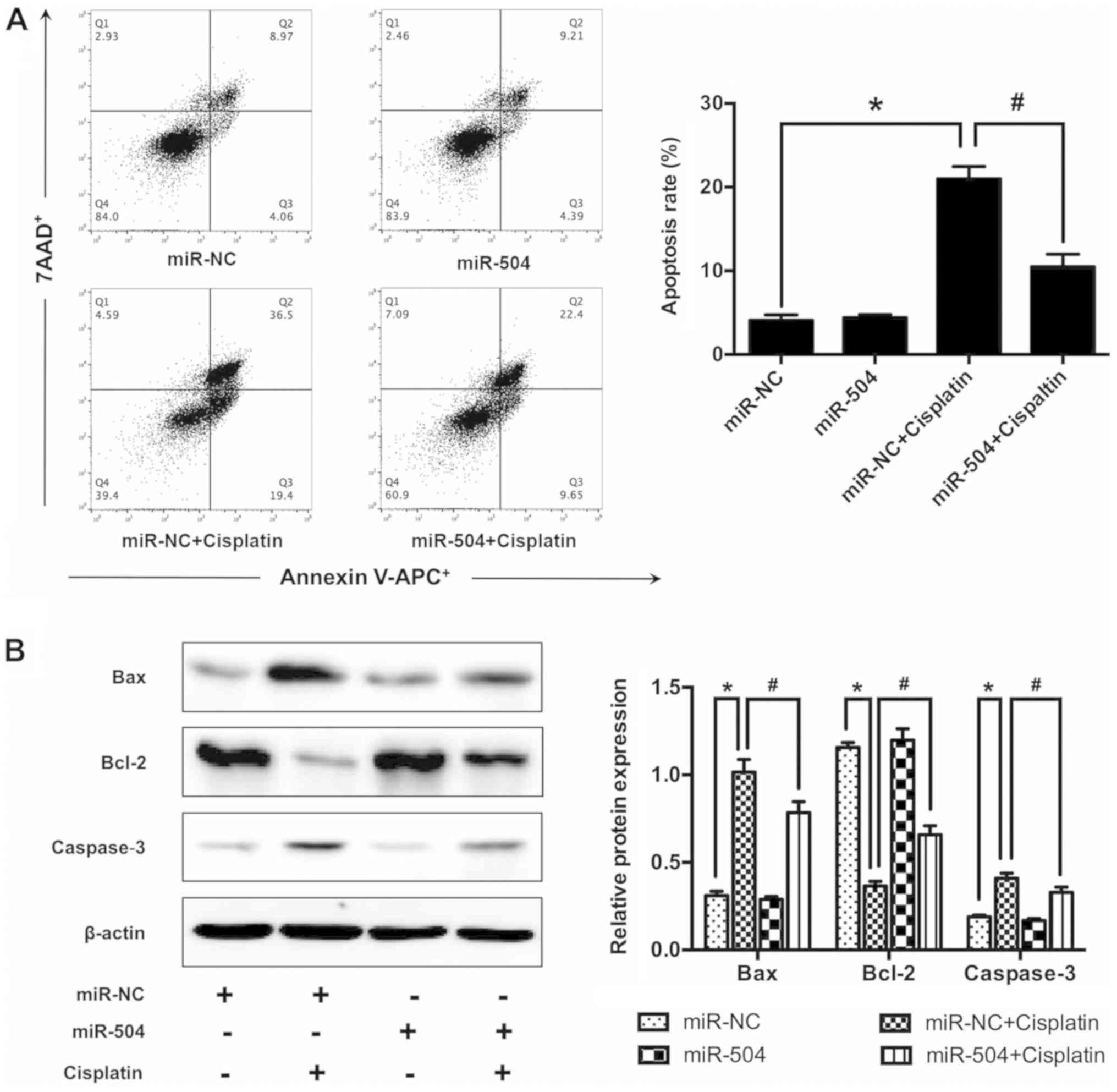
miR-504 suppresses cisplatin-induced
cell apoptosis in MG63 cells
The role of miR-504 in cisplatin-induced MG63 cell
apoptosis was assessed. Apoptosis rate was determined in each group
treated with 10 µg/ml cisplatin for 24, 48 or 72 h. According to
the MTT assay, miR-504 significantly decreased cisplatin-induced
cell apoptosis compared with the normal group at all time points
(P<0.05; Fig. 3C). No significant
differences were identified between the normal and miR-NC groups.
The morphological changes associated with MG63 cell
cisplatin-induced apoptosis were assessed using phase contrast
microscopy. Apoptotic rate (non-adherent cells) were significantly
increased in the miR-NC+cisplatin group compared with the miR-NC
group (P<0.05) and were significantly decreased in the
miR-504+cisplatin group compared with the miR-NC+cisplatin group
(P<0.05). There was no significant difference between the miR-NC
and miR-504 groups (Fig. 3D). Similar
trends were also observed via the Hoechst 33258 staining assay
(Fig. 3E). The involvement of miR-504
in cisplatin-induced cell apoptosis was confirmed via flow
cytometry, which revealed that cisplatin treatment significantly
increased cell apoptosis in the miR-NC+cisplatin group compared
with the miR-NC group (P<0.05; Fig.
4A), the same result was also revealed between miR-504 group
and miR-504+cisplatin group. Furthermore, miR-504 significantly
decreased cell apoptosis in the miR-504+cisplatin group compared
with the miR-NC+cisplatin group (P<0.05). No significant
differences were identified between the miR-NC and miR-504 groups.
To further assess the molecular mechanisms of miR-504, western
blotting was performed. The results demonstrated that cisplatin
treatment significantly increased Bax and caspase-3 (pro-apoptotic
protein) expression in the miR-NC+cisplatin group compared with the
miR-NC group (P<0.05). Furthermore, miR-504 significantly
decreased the expression of Bax and caspase-3 in the
miR-504+cisplatin group compared with the miR-NC+cisplatin group
(P<0.05). No significant differences were identified between the
miR-NC and miR-504 groups. The anti-apoptotic protein Bcl-2
demonstrated the opposite trends not only in the miR-NC+cisplatin
group compared with the miR-NC group (P<0.05), but also in the
miR-504+cisplatin group compared with the miR-NC+cisplatin group
(P<0.05; Fig. 4B). These results
demonstrated that miR-504 suppressed cisplatin-induced MG63 cell
apoptosis.
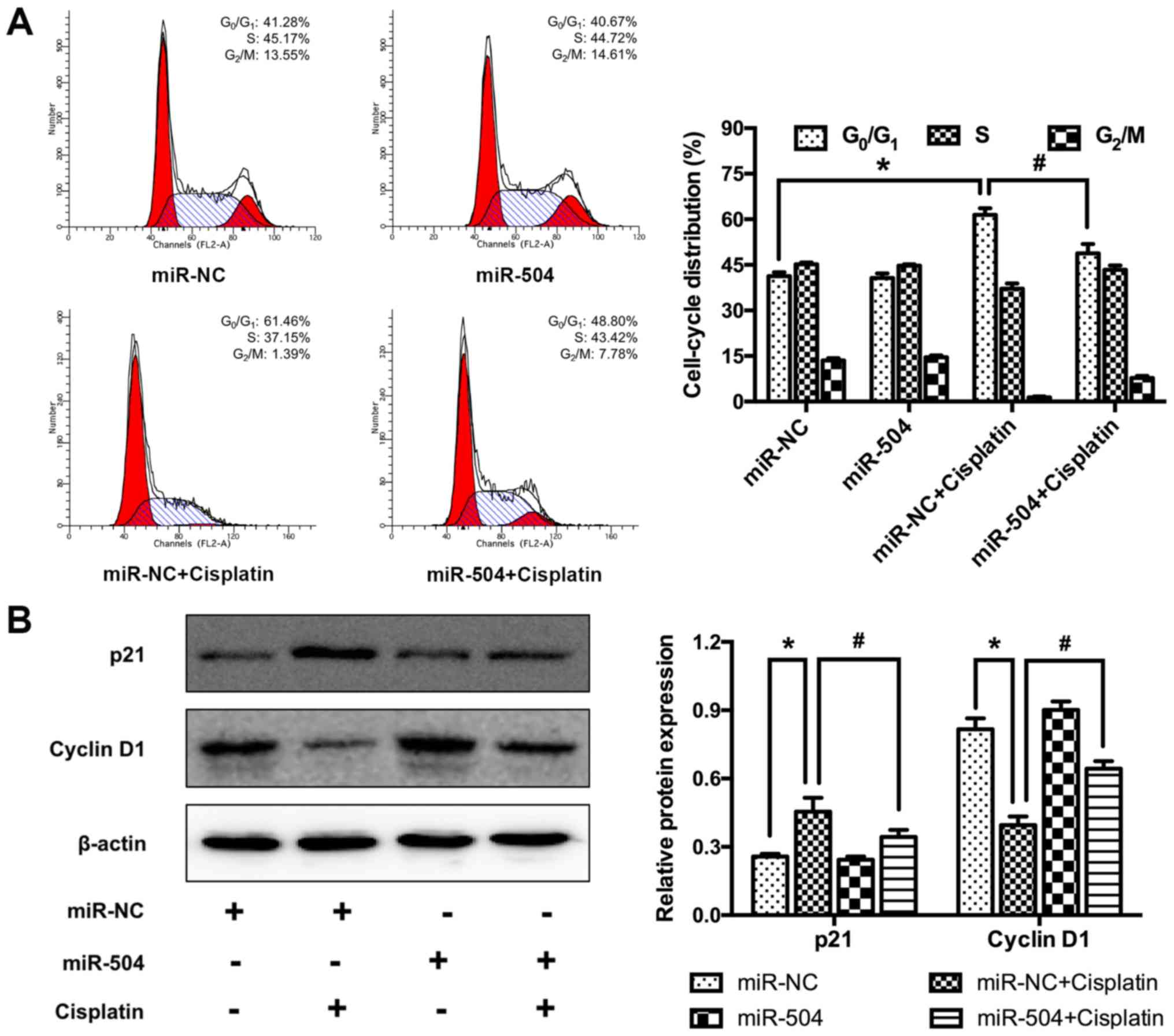
miR-504 suppresses cisplatin-induced
cell cycle arrest in MG63 cells
The involvement of miR-504 in cisplatin-induced MG63
cell cycle arrest was determined using flow cytometry. Cisplatin
treatment significantly increased the percentage of
G0/G1 phase cells in the miR-NC+cisplatin
group compared with the miR-NC group (P<0.05; Fig. 5A). However, miR-504 significantly
decreased the percentage of cells in G0/G1
phase in the miR-504+cisplatin group compared with the
miR-NC+cisplatin group (P<0.05). No significant differences were
identified between the miR-NC and miR-504 groups. The underlying
molecular mechanism was assessed using western blotting, which
revealed that cisplatin significantly increased the expression of
p21 (a non-specific suppressor of cell cycle progression) in the
miR-NC+cisplatin group compared with the miR-NC group (P<0.05).
Additionally, miR-504 treatment significantly decreased the
expression of p21 in the miR-504+cisplatin group compared with the
miR-NC+cisplatin group (P<0.05). There were no significant
differences between the miR-NC and the miR-504 groups.
Additionally, cisplatin treatment significantly decreased the
expression of cyclin D1 (a specific promotor of G1 to S
phase) in the miR-NC+cisplatin group compared with the miR-NC group
(P<0.05). Additionally, it was determined that miR-504
significantly increased cyclin D1 expression in the
miR-504+cisplatin group compared with the miR-NC+cisplatin group
(P<0.05; Fig. 5B). However, no
significant differences were identified between the miR-NC and
miR-504 groups. These results indicate that miR-504 suppressed
cisplatin-induced G0/G1 arrest in MG63
cells.
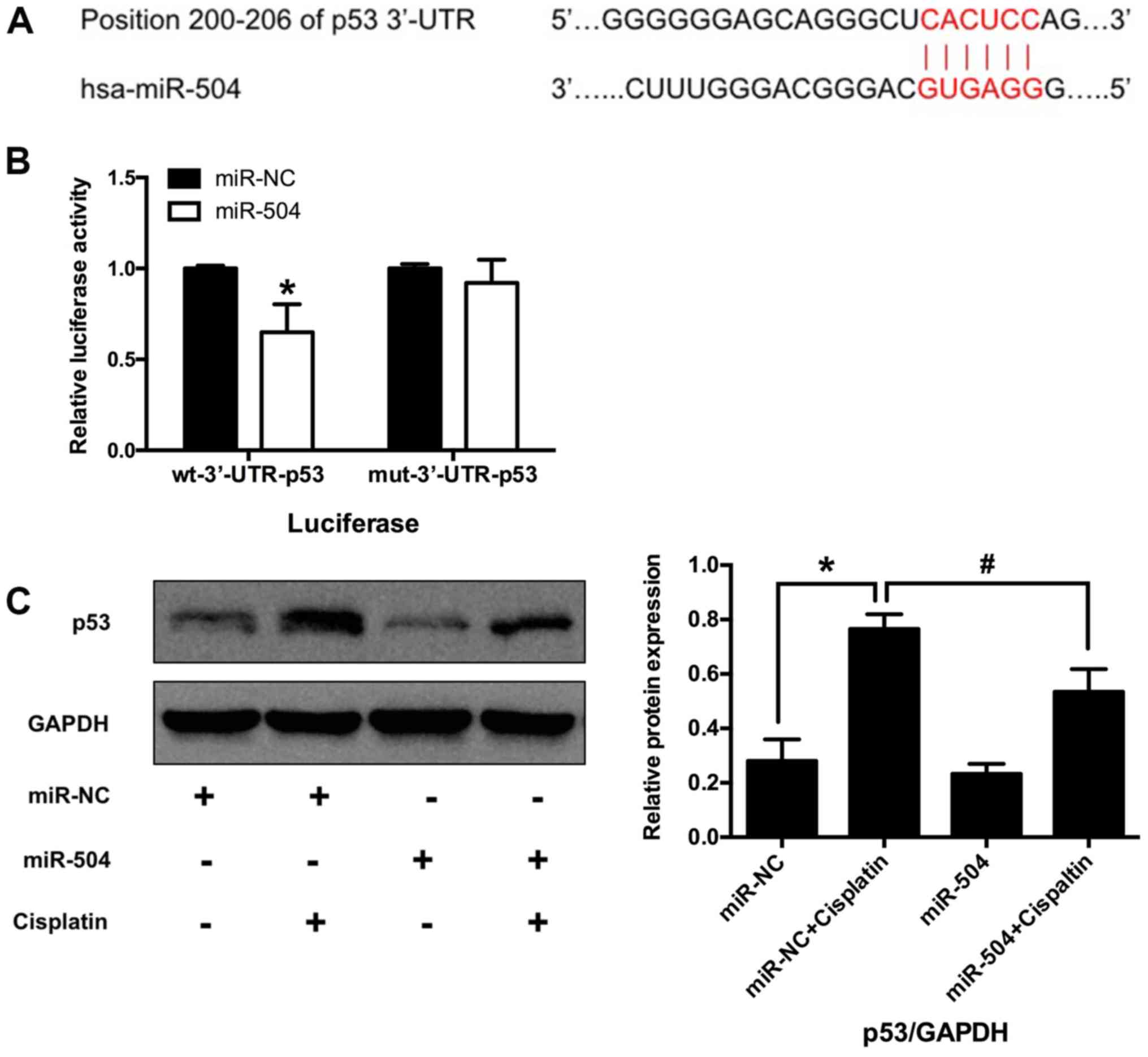
p53 is a direct target of miR-504 in
MG63 cells
p53-mediated apoptosis is a primary mechanism by
which p53 effects tumor suppression (38). Hu et al (20) demonstrated that miR-504 negatively
regulates the expression of p53 in various types of cell by binding
to the 3′-UTR of p53 mRNA. The current study therefore hypothesized
that p53 may be a target gene of miR-504 in osteosarcoma cells. The
TargetScanHuman database (Release 7.1, http://www.targetScan.org) identified a putative
region (at position 200–206) in the 3′-UTR of p53 mRNA that may
bind to miR-504 (Fig. 6A). Whether
miR-504 directly targeted p53 was then assessed using a luciferase
reporter assay. The results demonstrated that luciferase activity
was significantly decreased in wt miR-504 compared with wt miR-NC
(P<0.05; Fig. 6B). No significant
differences were identified between mut miR-504 and mut miR-NC
groups. These results indicate that p53 is a direct target of
miR-504 in MG63 cells.
p53 participates in miR-504-mediated
cell apoptosis and cell cycle arrest in MG63 cells
The role of p53 in miR-504-reduced MG63 cell
apoptosis was assessed via western blotting. The results
demonstrated that cisplatin significantly increased p53 expression
in the miR-NC+cisplatin group compared with the miR-NC group
(P<0.05; Fig. 6C). Furthermore,
miR-504 significantly decreased the expression of p53 in the
miR-504+cisplatin group compared with the miR-NC+cisplatin group
(P<0.05). However, no significant differences were identified
between the miR-NC and miR-504 groups. These results indicated that
p53 may be involved in miR-504-mediated cell apoptosis and cell
cycle arrest in MG63 cells.
Discussion
Osteosarcoma is the most common human primary
malignant bone tumor that is primarily characterized by local pain
and early metastasis (38). Although
osteosarcoma may be treated via surgery and chemotherapy, its
clinical prognosis remains poor. Osteosarcoma has been a recent
focus of research at the molecular level and studies have indicated
that miRNAs serve important roles in the development of
osteosarcoma (39,40). Several miRNAs have exhibited abnormal
expressions in osteosarcoma occurrence. For example, miR-93,
miR-181a and miR-191 are identified to be upregulated and miR-34a,
miR-145 and miR-199a-3p are downregulated (41). The current study revealed that miR-504
promoted proliferation and suppressed apoptosis, indicating that it
may serve an oncogenic role in osteosarcoma.
Cisplatin is the most commonly used
anti-osteosarcoma drug due to its unique therapeutic advantages,
which include a high efficiency, mild side effects and easy
administration. However, cisplatin resistance is frequently
reported, meaning that the enhancement of cisplatin sensitivity is
important for chemotherapy (42).
Song et al (43) demonstrated
that lysophosphatidic acid acytltransferase β silencing decreased
cisplatin resistance in osteosarcoma cells by activating the
phosphoinositide 3-kinase/protein kinase B/mammalian target of
rapamycin signaling pathway. Furthermore, Kim et al
(44) revealed that GDNF receptor
alpha 1 could overcome cisplatin resistance in osteosarcoma by
inhibiting AMP-activated protein activated kinase-dependent
autophagy. Additionally, Zheng et al (45) demonstrated that MAX dimerization
protein 1-mediated hypoxia-induced cisplatin resistance in
osteosarcoma cells by suppressing the expression of phosphatase and
tensin homolog. miRs have also been identified as novel modulators
that regulate the effect of cisplatin in osteosarcoma. For example,
miR-133b, miR-21 and miR-214 were revealed to be involved in the
induction of cisplatin resistance in osteosarcoma cells (46–48),
whilst miR-125b, miR-138 and miR-199a-5p enhanced osteosarcoma cell
cisplatin sensitivity (49–51).
However, previous studies have revealed that miR-504
expression differs among different types of human tumor cell;
miR-504 expression was increased in oral squamous carcinoma
(52), pancreatic ductal
adenocarcinoma (53) and gastric
cancer cells (54). Furthermore,
miR-504 overexpression was demonstrated to promote tumor
proliferation and reduce tumor sensitivity to radiotherapy and
chemotherapy. However, miR-504 expression was decreased in
hypopharyngeal squamous cell carcinoma (55) and glioma (56). The present study demonstrated that
miR-504 was overexpressed in osteosarcoma tissues and MG63 cells,
which lead to the hypothesis that it may be involved in
osteosarcoma occurrence and progression. The lentivirus-mediated
expression of miR-504 in MG63 cells indicated that miR-504 promoted
cell proliferation and reduced the cisplatin sensitivity by
suppressing MG63 cell apoptosis. This suggests that miR-504 may be
a sensitive index for the evaluation of cisplatin's therapeutic
effect. However, further studies are required to clarify the
mechanisms involved in this process.
The tumor suppressor protein, p53, activates DNA
repair proteins following DNA damage and initiates apoptosis in
cells with irreparable DNA damage to avoid the division of abnormal
genetic information (57). The p53
protein distinguishes DNA damage by inducing cell cycle arrest on
G1/S phase (58). DNA
damage induces the phosphorylation of p53, allowing it to
dissociate from E3 ubiquitin-protein ligase mdm2 and cause
p53-mediated tumor suppression via cell cycle arrest or apoptosis
(59). The p53 gene is mutated in
>50% of all types of malignant tumor and its mutations have been
demonstrated to be involved in osteosarcoma tumorigenesis (60). Furthermore, Li-Fraumeni syndrome is a
hereditary condition caused by the lack of the tumor suppressor p53
gene, which leads to the development and progression of multiple
types of malignant tumor, including osteosarcoma (61). The current study demonstrated that
miR-504 suppresses cell apoptosis and reduced G1 arrest
by negatively regulating p53, implying that miR-504 serves as an
oncogene in osteosarcoma.
Potential miRNA targets can be predicted using
bioinformatics software, including TargetScan. However, miRNAs
possess cell-specific target genes and functions, meaning these
predictions need to be confirmed experimentally. It has been
revealed that miR-504 mediates the expression of p53 in mammary
tumors (62) and in gastric carcinoma
(63). The current study therefore
hypothesized that p53 may also be a target gene of miR-504 in
osteosarcoma, which was confirmed by the results obtained.
Furthermore, previous studies have reported that miR-34a (64), miR-125b (65), miR-192 and miR-215 (66) negatively regulate p53 function,
particularly p53-mediated cell apoptosis and cell cycle arrest.
However, to the best of our knowledge, the current study provides
the first evidence to verify that miR-504 promotes cell
proliferation and suppresses the apoptosis of osteosarcoma cells by
targeting p53. The present study further assessed the changes in
apoptosis- and cell cycle-associated proteins at a molecular level.
The results revealed that miR-504 overexpression suppresses
apoptosis and induces the G1 arrest of MG63 cells by
regulating these proteins. However, further studies assessing the
in vitro knockdown of miR-504 are required to confirm the
results of the current study. Furthermore, subsequent studies
performed by the present authors will include in vivo
studies that assess patient clinicopathological characteristics,
which will further validate these results.
In conclusion, the current study revealed that
miR-504 promotes proliferation and suppresses cisplatin-induced
osteosarcoma cell apoptosis by targeting p53. However, further
studies are required to clarify the association between miR-504 and
p53, and the molecular mechanisms that are involved. The present
results indicate that miR-504 may be an effective marker for the
prediction of osteosarcoma occurrence and progression, and its
sensitivity to cisplatin chemotherapy.
Acknowledgements
The authors would like to thank Dr. Junying Sun for
his review of the manuscript and the Faculty of the Translational
Medicine Laboratory of the First Affiliated Hospital of Wenzhou
Medical University for supplying the laboratory to finish this
study.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JS designed the study to predict microRNA-504's role
in modulating osteosarcoma cell chemoresistance to cisplatin. CL,
ZH and SY analyzed and interpreted the patient data regarding
osteosarcoma. XC, XZ and WL performed the histological collection
of human osteosarcoma tissues and adjacent normal tissues. CL and
LW performed the in vitro study, which including MTT, cell
apoptosis assay, cell cycle analysis, dual luciferase reporter
assay and western blot analysis. XC, CL and JS were major
contributors in writing the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
institutional and/or national research committee and with the 1964
Helsinki declaration and its later amendments or comparable ethical
standards. The present study was approved by the Clinical Ethics
Committee of the First Affiliated Hospital of Wenzhou Medical
University. All participants provided written informed consent and
supported the study.
Patient consent for publication
Patients provided consent for the publication of the
present study and respective associated publications.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Endo-Munoz L, Evdokiou A and Saunders NA:
The role of osteoclasts and tumour-associated macrophages in
osteosarcoma metastasis. Biochim Biophys Acta. 1826:434–442.
2012.PubMed/NCBI
|
|
2
|
Poletajew S, Fus L and Wasiutynski A:
Current concepts on pathogenesis and biology of metastatic
osteosarcoma tumors. Ortop Traumatol Rehabil. 13:537–545. 2011.(In
English, Polish). View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lewis VO: What's new in musculoskeletal
oncology. J Bone Joint Surg Am. 91:1546–1556. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cho Y, Jung GH, Chung SH, Kim JY, Choi Y
and Kim JD: Long-term survivals of stage IIb osteosarcoma: A
20-year experience in a single institution. Clin Orthop Surg.
3:48–54. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bölling T, Schüller P, Distelmaier B,
Schuck A, Ernst I, Gosheger G, Winkelmann W, Dirksen U, Jürgens H,
Kronholz HL, et al: Perioperative high-dose rate brachytherapy
using a bendy applicator (flab): Treatment results of 74 patients.
Anticancer Res. 28:3885–3890. 2008.PubMed/NCBI
|
|
6
|
Faisham WI, Saad Mat AZ, Alsaigh LN, Azman
Nor MZ, Imran Kamarul M, Biswal BM, Bhavaraju VM, Salzihan MS,
Hasnan J, Ezane AM, et al: Prognostic factors and survival rate of
osteosarcoma: A single-institution study. Asia Pac J Clin Oncol.
13:e104–e110. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bacci G, Bertoni F, Longhi A, Ferrari S,
Forni C, Biagini R, Bacchini P, Donati D, Manfrini M, Bernini G, et
al: Neoadjuvant chemotherapy for high-grade central osteosarcoma of
the extremity. Histologic response to preoperative chemotherapy
correlates with histologic subtype of the tumor. Cancer.
97:3068–3075. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ferrari S and Serra M: An update on
chemotherapy for osteosarcoma. Expert Opin Pharmacother.
16:2727–2736. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shukla GC, Singh J and Barik S: MicroRNAs:
Processing, maturation, target recognition and regulatory
functions. Mol Cell Pharmacol. 3:83–92. 2011.PubMed/NCBI
|
|
11
|
Farh KK, Grimson A, Jan C, Lewis BP,
Johnston WK, Lim LP, Burge CB and Bartel DP: The widespread impact
of mammalian MicroRNAs on mRNA repression and evolution. Science.
310:1817–1821. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pasquinelli AE, Hunter S and Bracht J:
MicroRNAs: Adeveloping story. Curr Opin Genet Dev. 15:200–205.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Baer C, Claus R and Plass C: Genome-wide
epigenetic regulation of miRNAs in cancer. Cancer Res. 73:473–477.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin S, Pan L, Guo S, Wu J, Jin L, Wang JC
and Wang S: Prognostic role of microRNA-181a/b in hematological
malignancies: A meta-analysis. PLoS One. 8:e595322013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meng Y, Gao R, Ma J, Zhao J, Xu E, Wang C
and Zhou X: MicroRNA-140-5p regulates osteosarcoma chemoresistance
by targeting HMGN5 and autophagy. Sci Rep. 7:4162017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vanas V, Haigl B, Stockhammer V and
Sutterlüty-Fall H: MicroRNA-21 increases proliferation and
cisplatin sensitivity of osteosarcoma-derived cells. PLoS One.
11:e01610232016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y, Zhu ST, Wang X, Deng J, Li WH,
Zhang P and Liu BS: MiR-200c regulates tumor growth and
chemosensitivity to cisplatin in osteosarcoma by targeting AKT2.
Sci Rep. 7:135982017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu W, Chan CS, Wu R, Zhang C, Sun Y, Song
JS, Tang LH, Levine AJ and Feng Z: Negative regulation of tumor
suppressor p53 by microRNA miR-504. Mol Cell. 38:689–699. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bedi A and Mookerjee B: Biological
significance and molecular mechanisms of p53-induced apoptosis.
Apoptosis. 3:237–244. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Velletri T, Xie N, Wang Y, Huang Y, Yang
Q, Chen X, Chen Q, Shou P, Gan Y, Cao G, et al: P53 functional
abnormality in mesenchymal stem cells promotes osteosarcoma
development. Cell Death Dis. 7:e20152016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie C, Wu B, Chen B, Shi Q, Guo J, Fan Z
and Huang Y: Histone deacetylase inhibitor sodium butyrate
suppresses proliferation and promotes apoptosis in osteosarcoma
cells by regulation of the MDM2-p53 signaling. Onco Targets Ther.
9:4005–4013. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao YX, Wang YS, Cai QQ, Wang JQ and Yao
WT: Up-regulation of HDAC9 promotes cell proliferation through
suppressing p53 transcription in osteosarcoma. Int J Clin Exp Med.
8:11818–11823. 2015.PubMed/NCBI
|
|
25
|
Wu J, Guo A, Li Q and Wang D:
Meta-analysis of clinical significance of p53 protein expression in
patients with osteosarcoma. Future Oncol. 13:1883–1891. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
van der Deen M, Taipaleenmäki H, Zhang Y,
Teplyuk NM, Gupta A, Cinghu S, Shogren K, Maran A, Yaszemski MJ,
Ling L, et al: MicroRNA-34c inversely couples the biological
functions of the runt-related transcription factor RUNX2 and the
tumor suppressor p53 in osteosarcoma. J Biol Chem. 288:21307–21319.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang J, Ma B, Li X, Jin W, Han C, Wang L
and Wang H: MiR-1281, a p53-responsive MicroRNA, impairs the
survival of human osteosarcoma cells upon ER stress via targeting
USP39. Am J Cancer Res. 8:1764–1774. 2018.PubMed/NCBI
|
|
28
|
He C, Xiong J, Xu X, Lu W, Liu L, Xiao D
and Wang D: Functional elucidation of MiR-34 in osteosarcoma cells
and primary tumor samples. Biochem Biophys Res Commun. 388:35–40.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang W, Qian JX, Yi HL, Yang ZD, Wang CF,
Chen JY, Wei XZ, Fu Q and Ma H: The microRNA-29 plays a central
role in osteosarcoma pathogenesis and progression. Mol Biol (Mosk).
46:622–627. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cai Q, Zeng S, Dai X, Wu J and Ma W:
miR-504 promotes tumour hrowth and metastasis in human osteosarcoma
by targeting TP53INP1. Oncol Rep. 38:2993–3000. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lv C, Hao Y and Tu G: MicroRNA-21 promotes
proliferation, invasion and suppresses apoptosis in human
osteosarcoma line MG63 through PTEN/Akt pathway. Tumour Biol.
37:9333–9342. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cui R, Guan Y, Sun C, Bao Y, Chen L, Li G,
Qiu B, Meng X, Pang C and Wang Y: A tumor-suppressive microRNA,
miR-504, inhibits cell proliferation and promotes apoptosis by
targeting FOXP1 in human Glioma. Cancer Lett. 374:1–11. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:2015.doi: 10.7554/eLife.05005. View Article : Google Scholar
|
|
35
|
Stepanenko AA and Dmitrenko VV: HEK293 in
cell biology and cancer research: Phenotype, karyotype,
tumorigenicity, and stress-induced genome-phenotype evolution.
Gene. 569:182–190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nieuwenhuijsen BW, Huang Y, Wang Y,
Ramirez F, Kalgaonkar G and Young KH: A dual luciferase multiplexed
high-throughput screening platform for protein-protein
interactions. J Biomol Screen. 8:676–684. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rosen G, Caparros B, Huvos AG, Kosloff C,
Nirenberg A, Cacavio A, Marcove RC, Lane JM, Mehta B and Urban C:
Preoperative chemotherapy for osteogenic sarcoma: Selection of
postoperative adjuvant chemotherapy based on the response of the
primary tumor to preoperative chemotherapy. Cancer. 49:1221–1230.
1982. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bennett JH, Thomas G, Evans AW and Speight
PM: Osteosarcoma of the jaws: A 30-year retrospective review. Oral
Surg Oral Med Oral Pathol Oral Radiol Endod. 90:323–332. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chang L, Shrestha S, LaChaud G, Scott MA
and James AW: Review of microRNA in osteosarcoma and
chondrosarcoma. Med Oncol. 32:6132015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kumar Ram RM, Boro A and Fuchs B:
Involvement and clinical aspects of MicroRNA in osteosarcoma. Int J
Mol Sci. 17:pii: E877. 2016.
|
|
41
|
Kobayashi E, Hornicek FJ and Duan Z:
MicroRNA involvement in osteosarcoma. Sarcoma. 2012:3597392012.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yang J, Guo W, Wang L, Yu L, Mei H, Fang
S, Ji P, Liu Y, Liu G and Song Q: Cisplatin-resistant osteosarcoma
cells possess cancer stem cell properties in a mouse model. Oncol
Lett. 12:2599–2605. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Song L, Duan P, Gan Y, Li P, Zhao C, Xu J,
Zhang Z and Zhou Q: Silencing LPAATβ inhibits tumor growth of
cisplatin-resistant human osteosarcoma in vivo and in
vitro. Int J Oncol. 50:535–544. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kim M, Jung JY, Choi S, Lee H, Morales LD,
Koh JT, Kim SH, Choi YD, Choi C, Slaga TJ, et al: GFRA1 promotes
cisplatin-induced chemoresistance in osteosarcoma by inducing
autophagy. Autophagy. 13:149–168. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zheng D, Wu W, Dong N, Jiang X, Xu J, Zhan
X, Zhang Z and Hu Z: Mxd1 mediates hypoxia-induced cisplatin
resistance in osteosarcoma cells by repression of the PTEN tumor
suppressor gene. Mol Carcinog. 56:2234–2244. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zou Y, Yang J, Wu J, Luo C and Huang Y:
miR-133b induces chemoresistance of osteosarcoma cells to cisplatin
treatment by promoting cell death, migration and invasion. Oncol
Lett. 15:1097–1102. 2018.PubMed/NCBI
|
|
47
|
Ziyan W and Yang L: MicroRNA-21 regulates
the sensitivity to cisplatin in a human osteosarcoma cell line. Ir
J Med Sci. 185:85–91. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Song YD, Li DD, Guan Y, Wang YL and Zheng
J: miR-214 modulates cisplatin sensitivity of osteosarcoma cells
through regulation of anaerobic glycolysis. Cell Mol Biol
(Noisy-le-grand). 63:75–79. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhu Z, Tang J, Wang J, Duan G, Zhou L and
Zhou X: MiR-138 acts as a tumor suppressor by targeting EZH2 and
enhances cisplatin-induced apoptosis in osteosarcoma cells. PLoS
One. 11:e01500262016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang F, Yu D, Liu Z, Wang R, Xu Y, Cui H
and Zhao T: MiR-125b functions as a tumor suppressor and enhances
chemosensitivity to cisplatin in osteosarcoma. Technol Cancer Res
Treat. 15:NP105–NP112. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li Y, Jiang W, Hu Y, Da Z, Zeng C, Tu M,
Deng Z and Xiao W: MicroRNA-199a-5p inhibits cisplatin-induced drug
resistance via inhibition of autophagy in osteosarcoma cells. Oncol
Lett. 12:4203–4208. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yang MH, Lin BR, Chang CH, Chen ST, Lin
SK, Kuo MY, Jeng YM, Kuo ML and Chang CC: Connective tissue growth
factor modulates oral squamous cell carcinoma invasion by
activating a miR-504/FOXP1 signalling. Oncogene. 31:2401–2411.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jiang B, Gu Y and Chen Y: Identification
of novel predictive markers for the prognosis of pancreatic ductal
adenocarcinoma. Cancer Invest. 32:218–225. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Soutto M, Chen Z, Saleh MA, Katsha A, Zhu
S, Zaika A, Belkhiri A and El-Rifai W: TFF1 activates p53 through
down-regulation of miR-504 in gastric cancer. Oncotarget.
5:5663–5673. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kikkawa N, Kinoshita T, Nohata N, Hanazawa
T, Yamamoto N, Fukumoto I, Chiyomaru T, Enokida H, Nakagawa M,
Okamoto Y and Seki N: microRNA-504 inhibits cancer cell
proliferation via targeting CDK6 in hypopharyngeal squamous cell
carcinoma. Int J Oncol. 44:2085–2092. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Guan Y, Chen L, Bao Y, Pang C, Cui R, Li
G, Liu J and Wang Y: Downregulation of microRNA-504 is associated
with poor prognosis in high-grade glioma. Int J Clin Exp Pathol.
8:727–734. 2015.PubMed/NCBI
|
|
57
|
Mirzayans R, Andrais B, Kumar P and Murray
D: Significance of wild-type p53 signaling in suppressing apoptosis
in response to chemical genotoxic agents: Impact on chemotherapy
outcome. Int J Mol Sci. 18:pii: E928. 2017.
|
|
58
|
Vassilev LT, Vu BT, Graves B, Carvajal D,
Podlaski F, Filipovic Z, Kong N, Kammlott U, Lukacs C, Klein C, et
al: In vivo activation of the p53 pathway by small-molecule
antagonists of MDM2. Science. 303:844–848. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hu W, Feng Z, Ma L, Wagner J, Rice JJ,
Stolovitzky G and Levine AJ: A single nucleotide polymorphism in
the MDM2 gene disrupts the oscillation of p53 and MDM2 levels in
cells. Cancer Res. 67:2757–2765. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Hansen MF: Molecular genetic
considerations in osteosarcoma. Clin Orthop Relat Res. 237–246.
1991.PubMed/NCBI
|
|
61
|
Hauben EI, Arends J, Vandenbroucke JP, van
Asperen CJ, Van Marck E and Hogendoorn PC: Multiple primary
malignancies in osteosarcoma patients. Incidence and predictive
value of osteosarcoma subtype for cancer syndromes related with
osteosarcoma. Eur J Hum Genet. 11:611–618. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ford NA, Dunlap SM, Wheatley KE and
Hursting SD: Obesity, independent of p53 gene dosage, promotes
mammary tumor progression and upregulates the p53 regulator
microRNA-504. PLoS One. 8:e680892013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Fukuda Y, Kurihara N, Imoto I, Yasui K,
Yoshida M, Yanagihara K, Park JG, Nakamura Y and Inazawa J: CD44 is
a potential target of amplification within the 11p13 amplicon
detected in gastric cancer cell lines. Genes Chromosomes Cancer.
29:315–324. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Novello C, Pazzaglia L, Conti A, Quattrini
I, Pollino S, Perego P, Picci P and Benassi MS: p53-dependent
activation of microRNA-34a in response to etoposide-induced DNA
damage in osteosarcoma cell lines not impaired by dominant negative
p53 expression. PLoS One. 9:e1147572014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Le MT, Teh C, Shyh-Chang N, Xie H, Zhou B,
Korzh V, Lodish HF and Lim B: MicroRNA-125b is a novel negative
regulator of p53. Genes Dev. 23:862–876. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Braun CJ, Zhang X, Savelyeva I, Wolff S,
Moll UM, Schepeler T, Ørntoft TF, Andersen CL and Dobbelstein M:
p53-responsive micrornas 192 and 215 are capable of inducing cell
cycle arrest. Cancer Res. 68:10094–10104. 2008. View Article : Google Scholar : PubMed/NCBI
|