Introduction
The increased global mortality rate due to liver
cancer may be attributed to the aggressive nature of the disease
(1). Liver cancer accounts for >1
million newly diagnosed cases annually, making it a global health
care problem with increasing incidence (2). The incidence of liver cancer is markedly
high in developing countries and is steadily rising in developed
countries (3). Although there is a
continuous rise in the 5-year survival rates for liver and
intrahepatic bile duct cancer, the numbers of new cases and
estimated mortalities are increasing (4,5). Liver
cancer is often not detectable until late in disease progression
(6), and chemotherapy remains the
primary treatment for numerous patients with cancer of advanced
stage. However, chemotherapy has side effects, particularly in
normal tissues with potent proliferative activity (7), and this therapy does not always lead to
a better prognosis. Therefore, finding novel therapeutic entities
against longer cancer cell survival and growth is required.
JTC-801
[N-(4-amino-2-methylquinolin-6-yl)-2-(4-ethylphenoxymethyl)benzamide
monohydrochloride] is a high-affinity and selective opioid
receptor-like 1 (ORL1) receptor antagonist, which belongs to the
family of G-protein-coupled receptors (GPCRs) (8). Studies have demonstrated that signal
transduction through GPCRs can affect numerous aspects of cancer
biology, including invasion, migration and vascular remodelling
(9,10). Furthermore, ORL1 may antagonize
lipopolysaccharide-stimulated proliferation, migration and
inflammatory signalling in human glioblastoma U87 cells (11). In a screening of agents that interact
with GPCR pathways, Song et al (12) revealed that JTC-801 induces
pH-dependent cell death (alkaliptosis) specifically in cancer
cells, including pancreatic ductal adenocarcinoma cells, by
reducing the expression of carbohydrate antigen 9, which is
increased in human cancer tissues. JTC-801 may be used in the
development of treatment for pancreatic cancer. Zheng et al
(13) described the antitumour
effects of JTC-801 on human osteosarcoma cells. Therefore, JTC-801
may be used for the treatment of other cancer types, including
liver cancer.
In the present study, the effect of JTC-801 on the
Hep G2 hepatoblastoma cell line was investigated. First, the Cell
Counting Kit-8 (CCK-8) assay was used to detect the proliferation
of Hep G2 cells treated with JTC-801. As cancer is characterized by
increased migratory/invasive capacity, the migratory and invasive
abilities of Hep G2 cells following JTC-801 treatment were
investigated. Furthermore, the expression of apoptotic proteins was
assayed in the Hep G2 cells following JTC-801 treatment.
Additionally, the phosphatidylinositol 3-kinase (PI3K)/protein
kinase B (AKT) signalling pathway may have a function in inducing
the apoptosis of Hep G2 cells.
Materials and methods
Cell lines and cell culture
The human Hep G2 cell line was purchased from the
Shanghai Cell Bank (Shanghai Institute for Biological Science,
Chinese Academy of Science, Shanghai, China). Cells were cultured
in RPMI-1640 medium (GE Healthcare Lifesciences, Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and 0.1
mg/ml streptomycin (both Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) at 37°C in an incubator with 5% CO2. When the
cells entered the logarithmic growth phase they were washed 3 times
with PBS and digested with 0.25% trypsin/EDTA (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China). Cells were
resuspended in RPMI-1640 medium containing 10% FBS to form a
single-cell suspension. The cells were seeded in a 6-well plate for
subsequent experiments. When the cell density reached ~80%, the
cells were treated with 20 µM JTC-801 (MedChem Express, Monmouth
Junction, NJ, USA), whereas the negative control group was treated
with 0.1% dimethylsulfoxide (DMSO) (Amresco, LLC, Solon, OH, USA),
for 24 h at room temperature.
Western blot analysis
Following the treatment of the cells with DMSO and
JTC-801 for 24 h, protein was extracted with
radioimmunoprecipitation assay lysis buffer with the protease
inhibitor phenylmethylsulfonyl fluoride (CWBio, Beijing, China).
The concentration was determined using the BCA method (CWBio). The
protein was heated at 95°C for 5 min. Overall, ~20 µg protein per
group was added to each well in the vertical electrophoresis tank,
separated by 10% SDS-PAGE and transferred onto a polyvinylidene
fluoride membrane. The membrane was blocked with 5% non-fat milk in
Tris-buffered saline containing 0.1% Tween-20 (TBST; Beijing
Solarbio Science & Technology Co., Ltd.) for 1 h at room
temperature and incubated with primary antibodies at 4°C overnight.
On the second day, the membrane was washed 3 times in TBST for 5
min and incubated with secondary antibodies at room temperature for
1 h. An enhanced chemiluminescence chromogenic substrate
(ProteinTech Group, Inc., Chicago, IL, USA) was added to visualize
the bands, following the washing of the membrane. The grey value
was scanned by Quantity One software (version 4.6.9; Bio-Rad
Laboratories, Inc., Hercules, CA, USA). β-tubulin served as the
internal control. The relative expression of each protein was
calculated against that of β-tubulin. Western blotting was
performed with the following antibodies: Rabbit anti-human AKT
(cat. no. 4691; 1:1,000 dilution), rabbit anti-human p-AKT (cat.
no. 4060; 1:1,000 dilution), rabbit anti-human mechanistic target
of rapamycin (mTOR; cat. no. 2983; 1:1,000 dilution), rabbit
anti-human p-mTOR (cat. no. 2971; 1:1,000 dilution) (all Cell
Signaling Technology, Inc., Danvers, MA, USA), rabbit anti-human
B-cell lymphoma 2 (Bcl-2; cat. no. 12789-1-AP; 1:1,000 dilution),
rabbit anti-human apoptosis regulator BAX (Bax; cat. no.
50599-2-Ig; 1:1,000 dilution), rabbit anti-human active caspase-3
(cat. no. 19677-1-AP; 1:1,000 dilution), rabbit anti-human cyclin
D1 (cat. no. 60186-1-Ig; 1:1,000 dilution), rabbit anti-human p70S6
kinase (p70S6K; cat. no. 14485-1-AP; 1:1,000 dilution), rabbit
anti-human tubulin (cat. no. 10068-1-AP; 1:5,000 dilution) and the
secondary horseradish peroxidase-labelled goat anti-rabbit/goat
anti-mouse antibody (cat. no. 10545-2-AP; 1:5,000 dilution) (all
ProteinTech Group, Inc.).
Cell Counting Kit-8 (CCK-8)
proliferation test
Hep G2 cells from the conventional culture were
digested with 0.25% trypsin-EDTA solution for 2 min at room
temperature and counted in preparation of the cell suspension.
Subsequently, 100 µl of the cell suspension was seeded onto a
96-well plate with 1,000 cells per well and 0.1% DMSO was added to
the negative control group, while 0.02, 0.2, 2, 20 or 200 µM
JTC-801 was added to the experimental groups. The cells were
cultured in a 5% CO2 incubator at room temperature to
detect cell viability once every 24 h. Prior to detection, 10 µl
CCK-8 solution (CWBio) was added to each well and incubated at 37°C
for 1.5 h. The optical density (OD) was measured with a microplate
reader at 450 nm and the growth curve was plotted.
Cell invasion and migration assessed
by Transwell assay
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA)
was dissolved overnight (serum-free RPMI-1640 medium, 1:6 dilution)
and 100 µl was added to the upper chamber of the 24-well Transwell
insert (Merck KGaA). Once the Matrigel was evenly distributed, it
was placed into a CO2 incubator and cultivated for 4–6 h
at 37°C until gel formation. Subsequent to the culture medium
drying, 500 µl serum-free medium was added to the bottom of the
wells to hydrate the basement membrane for 30 min. The cell
suspension, treated with JTC-801 for 24 h at room temperature, was
prepared using serum-free RPMI-1640 medium. A total of 100 µl cell
suspension (1×105 cells) was loaded into the upper
chamber and 500 µl complete culture medium including 10% FBS was
added to the bottom of the Transwell insert for incubation
overnight at room temperature. The following day, the Transwell
insert was removed and cells remaining on the upper chamber were
removed with a cotton swab. Following the washing of the chamber
with PBS, the cells that adhered to the membrane were fixed in 4%
paraformaldehyde for 30 min, stained with 0.1% crystal violet for
20 min, both at room temperature, and washed with PBS. Finally, 5
visual fields were selected randomly using a BX51 inverted
microscope (Olympus Corporation, Tokyo, Japan) at ×100
magnification, and images were captured for cell counts.
The migration assay procedure was similar to that of
the invasion assay except that Matrigel was not placed in the
Transwell chamber and 5×103 cells were used.
Cell apoptosis assay
The Hep G2 cells were treated with JTC-801 for 24 h,
as aforementioned, and the medium was removed and replaced with 500
µl serum-free medium. Following starvation for 24 h, the cells were
harvested, digested with trypsin without EDTA, collected in a
centrifuge tube, centrifuged at 200 × g for 5 min at room
temperature, resuspended in pre-cooled PBS at 4°C and centrifuged
again as aforementioned, and then the supernatant was carefully
aspirated. A total of 200 µl Annexin V-FITC binding solution
(contained within the kit) was added to resuspend the cells and the
cell density was adjusted to 1–5×106 cells/ml. Overall,
~100 µl cell suspension was transferred into a 5-ml flow tube
following staining with 5 µl Annexin V-fluorescein isothiocyanate
(FITC) (FXP018-100; 4A Biotech Co., Ltd., Beijing, China) for 5 min
at room temperature and kept in the dark. The cells were stained
with 10 µl propidium iodide (PI) in 400 µl PBS for 10 min at room
temperature and kept in the dark, prior to collection and detection
using flow cytometry. The results were analysed with FlowJo
software (version 7.6.5; Tree Star, Inc., Ashland, OR, USA).
Statistical analysis
The experimental data were analysed by SPSS
statistical analysis software (version 18.0; SPSS Inc., Chicago,
IL, USA). The results were expressed as the mean ± standard
deviation. The comparison of 2 groups was performed using the
Student's t-test and >2 groups were compared using one-way
analysis of variance. Multiple comparisons between the groups were
performed using a Student-Newman-Keuls test. P<0.05 was
considered to indicate a statistically significant difference.
Results
JTC-801 inhibits Hep G2 cell
proliferation
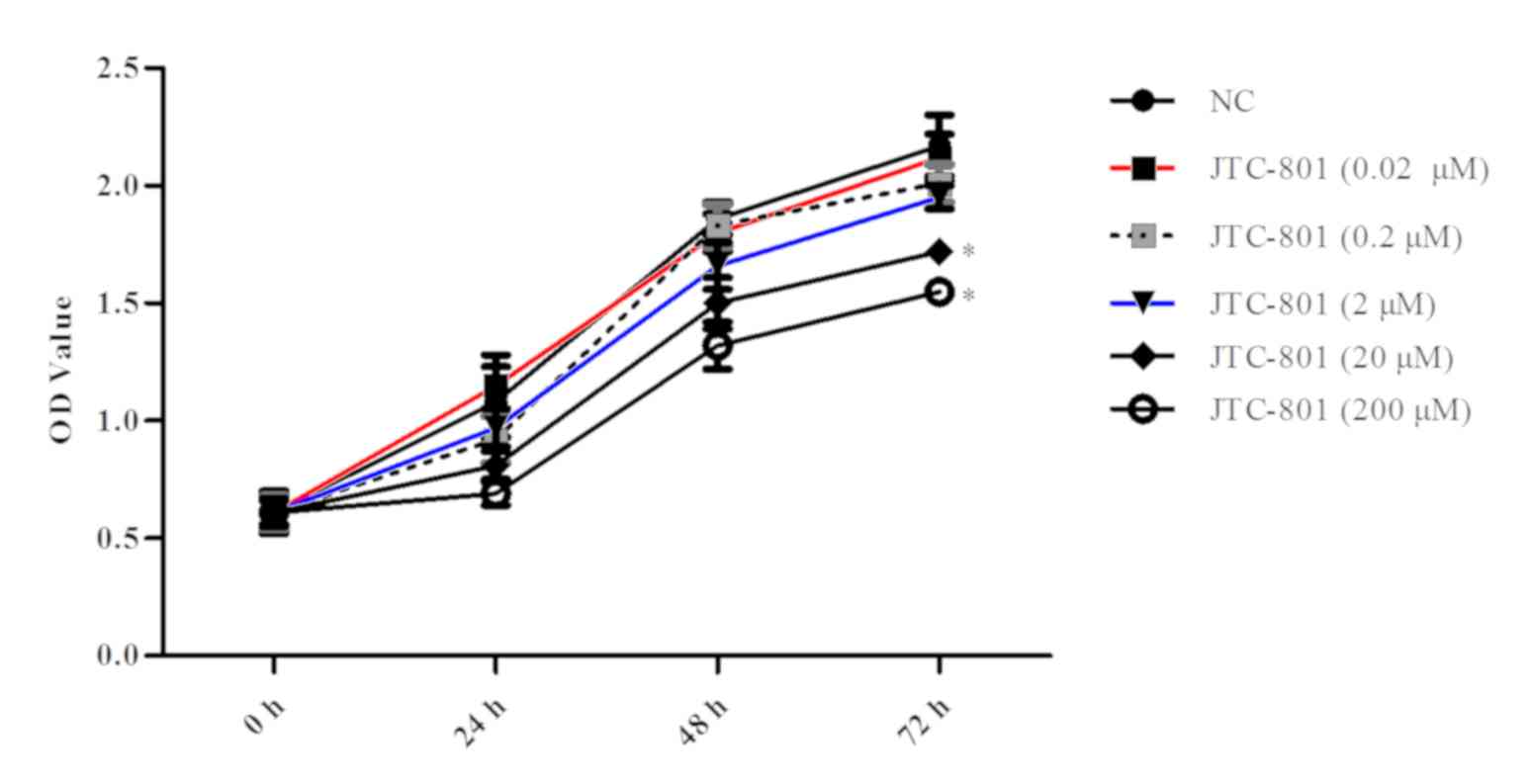
To evaluate the effect of JTC-801 on liver cancer,
the Hep G2 cell line was selected and treated with various doses of
JTC-801 (0.02, 0.2, 2, 20 and 200 µM) for 24, 48 and 72 h to detect
cell viability. As presented in Fig.
1, higher concentrations of JTC-801 (≥20 µM) led to a
statistically significant decrease in the proliferation of the Hep
G2 cells compared with the control (P<0.05). In the following
experiment, 20 µM JTC-801 was used. Furthermore, the OD value
decreased significantly (P<0.05; Fig.
1) at 72 h. This suggests that JTC-801 may effectively inhibit
liver cancer Hep G2 cell proliferation.
JTC-801 inhibits Hep G2 cell invasion
and migration
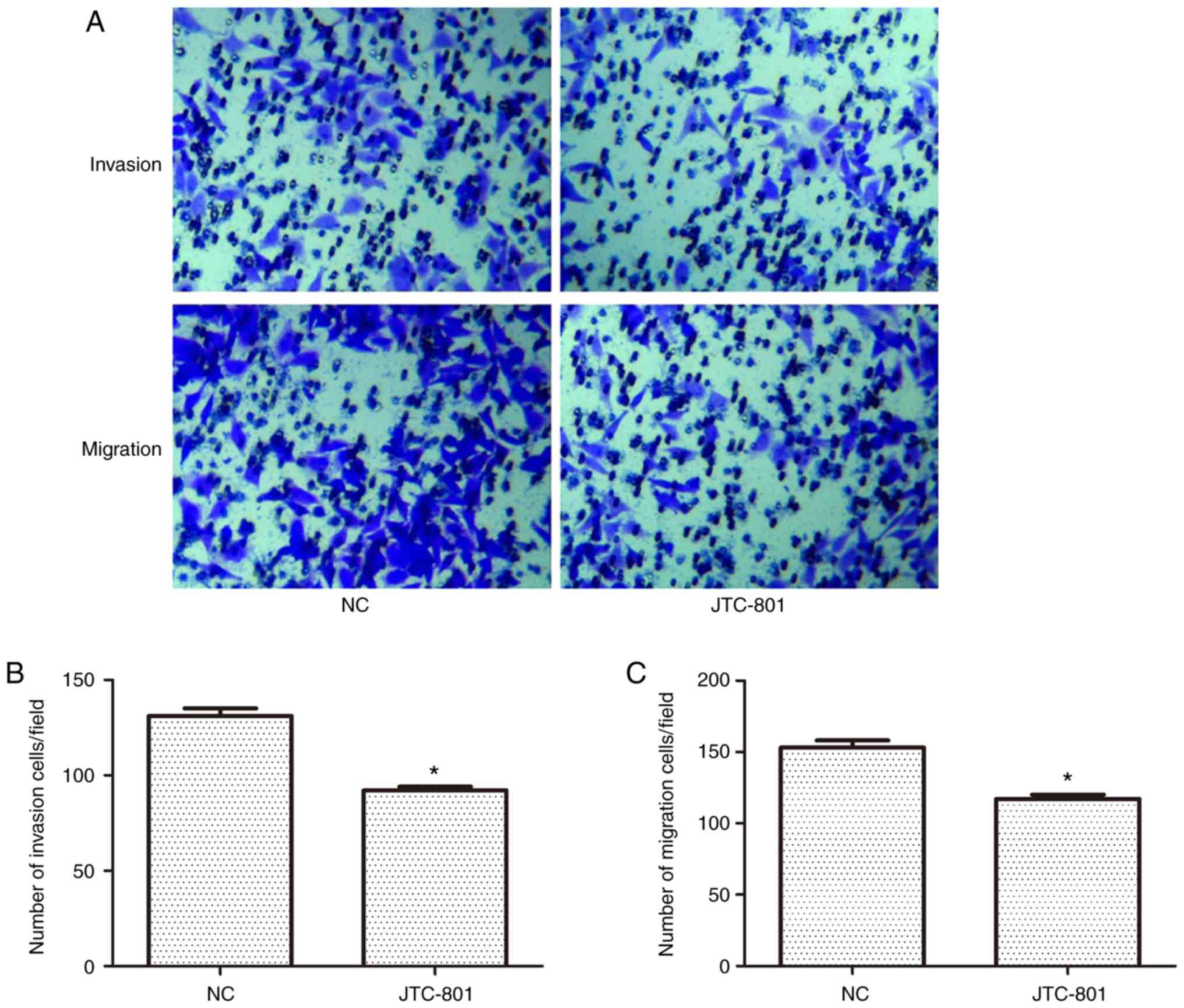
The effects of JTC-801 on the invasion and migration
ability of the Hep G2 cells were investigated using a Transwell
assay (Fig. 2A). In the invasion
experiment, the number of crystal violet-stained cells was
decreased following the JTC-801 treatment. Similarly, in the
migration experiment, cell migration was also decreased. These
results suggest that JTC-801 may inhibit the metastasis of Hep G2
cells. There were fewer invaded cells in the JTC-801-treated group
than in the control group (92±2 vs. 131±4 cells, respectively),
indicating a negative effect of JTC-801 on the invasion ability of
the Hep G2 cells (P<0.05; Fig.
2B). The capacity for migration was also inhibited in the
JTC-801-treated cells as compared with that in the negative control
group (117±3 vs. 153±5 cells, respectively), and the results were
significantly different (P<0.05; Fig.
2C).
JTC-801 promotes Hep G2 cell
apoptosis
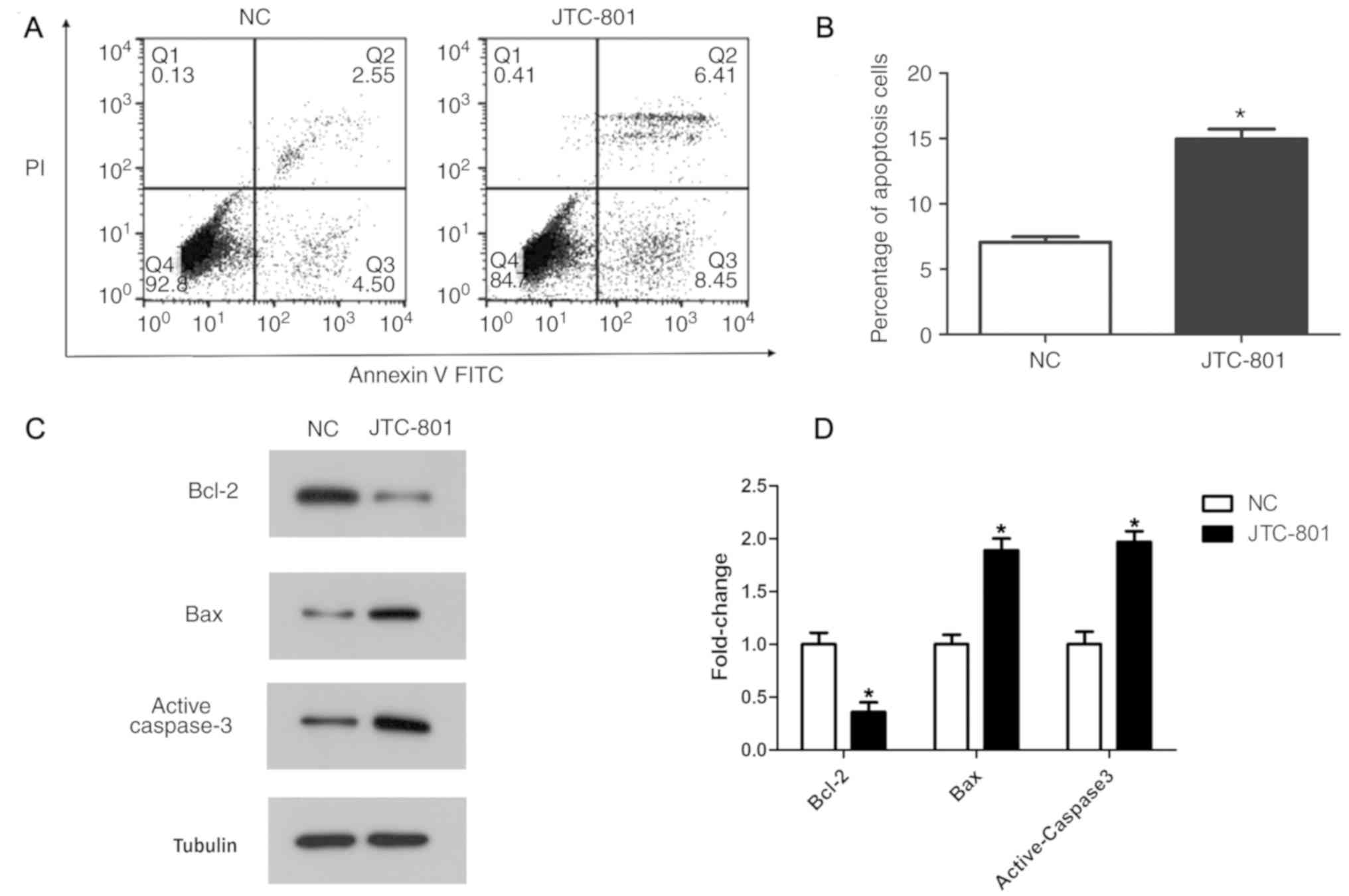
The impact of JTC-801 on Hep G2 cell apoptosis was
determined by an Annexin V-FITC and PI double-staining assay. The
apoptosis rate in the JTC-801-treated group was 14.86%, while the
apoptosis rate in the control group was 7.05%. The statistical
analysis suggested that the apoptosis rate in the JTC-801-treated
group was significantly increased compared with that in the control
group (P<0.05; Fig. 3B).
Furthermore, the apoptosis regulators, including anti-apoptotic
protein Bcl-2, and pro-apoptotic proteins active caspase-3 and Bax,
were analysed by western blotting (Fig.
3C). In concordance with the flow cytometry analysis, the
western blotting revealed that the expression of the anti-apoptotic
protein Bcl-2 was decreased, and the expression of the
pro-apoptotic proteins active caspase-3 and Bax was increased in
the JTC-801-treated group compared with that in the control group
(P<0.05; Fig. 3D). These results
indicate that JTC-801 promotes Hep G2 cell apoptosis.
JTC-801 suppresses the PI3K/AKT
pathway in Hep G2 cells
The PI3K/AKT signalling pathway is important in
tumour progression. Proteins mTOR, p70S6K and cyclin D1 were
selected as indicators to evaluate the activity of the PI3K/AKT
signalling pathway following the JTC-801 treatment. The results of
the western blotting revealed that the phosphorylation levels of
AKT and mTOR were significantly decreased in the JTC-801-treated
Hep G2 cells, as were the expression levels of p70S6K and cyclin D1
(P<0.05; Fig. 4). These results
suggest that JTC-801-induced Hep G2 cell growth inhibition
functions via the PI3K/AKT pathway.
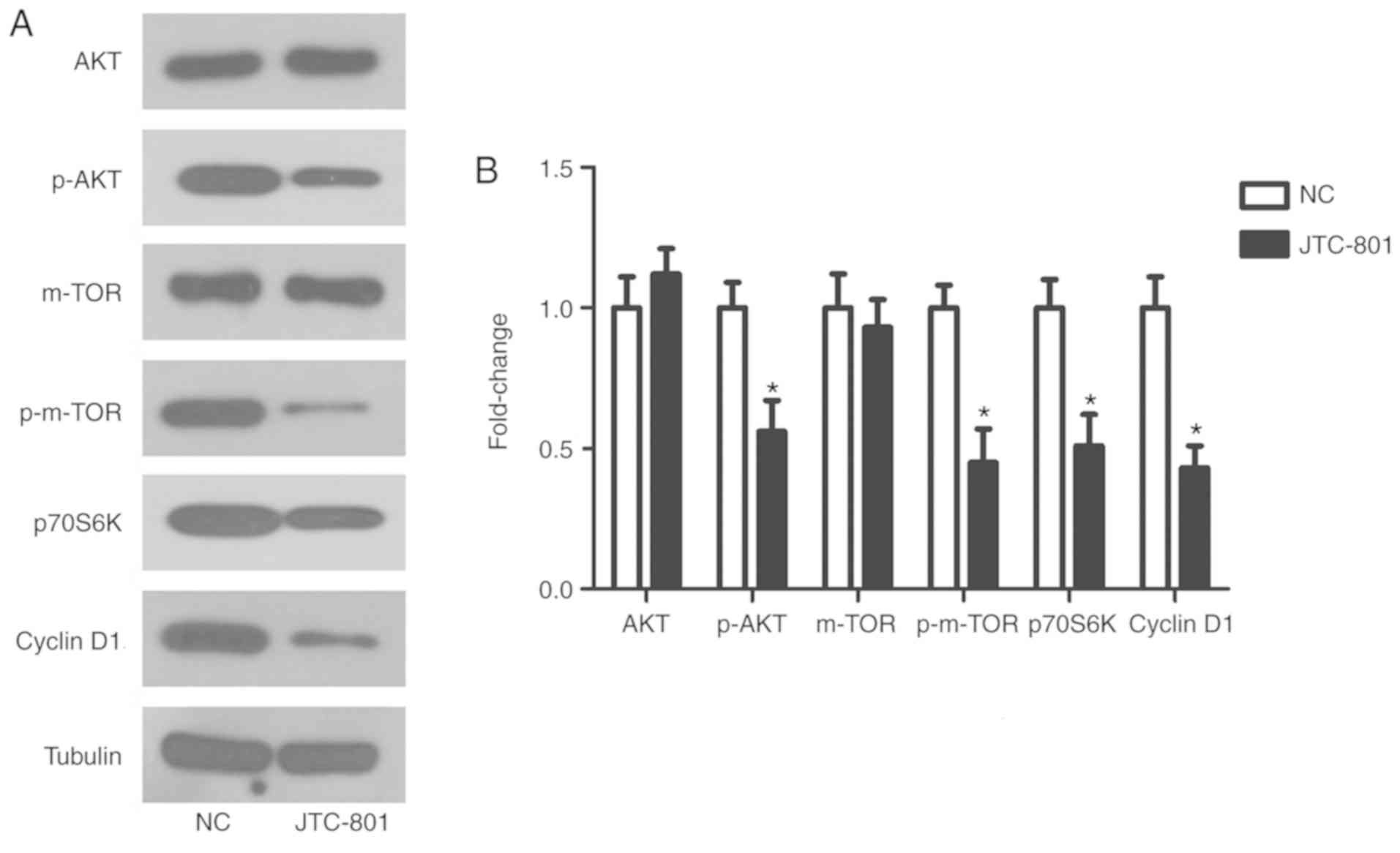 | Figure 4.Effects of JTC-801 on the
phosphatidylinositol 3-kinase/AKT signalling pathway in Hep G2
cells. (A) Expression levels of AKT, p-AKT, mTOR, p-mTOR, p70S6K
and cyclin D1 were measured in the Hep G2 cells treated with
JTC-801 using western blot analysis. (B) The relative protein
levels of AKT, p-AKT, mTOR, p-mTOR, p70S6K and cyclin D1 compared
with the control group. *P<0.05 compared with the NC group. AKT,
protein kinase B; p-AKT, phosphorylated AKT; mTOR, mechanistic
target of rapamycin; p-mTOR, phosphorylated mTOR; P70S6K, P70S6
kinase; NC, negative control. |
Discussion
In the present study, the inhibition of
hepatoblastoma Hep G2 cell proliferation, invasion and migration by
the JTC-801 antagonist was demonstrated, along with the promotion
of cell apoptosis. Finally, the effects of JTC-801 on Hep G2 cells
were indicated to be associated with the inhibition of PI3K/AKT
signalling.
JTC-801 is an opioid analgesic drug used in
scientific research. However, the antitumour activity of JTC-801 is
not dependent on its known analgesic function (14). JTC-801 contributes to this process by
inducing a unique pH-dependent form of regulated cell death known
as alkaliptosis (14,15). In U20S osteosarcoma cells, JTC-801
inhibited cell growth by promoting apoptosis via the PI3K/AKT
signalling pathway (13).
Furthermore, JTC-801 demonstrated its anticancer effect in the
ovarian cancer SKOV3 cell line, particularly on cell growth and
metastasis (16). Song et al
(12) reported that JTC-801 may be
used for the treatment of pancreatic cancer. Similarly, the present
study revealed the inhibitory effect of the drug on Hep G2
hepatoblastoma cells. The anticancer effect of JTC-801 only
occurred at a concentration ≥20 µM. The high concentration of
JTC-801 (>100 µM) acts on a wide range of receptors, including
GPCRs, ion channel receptors and nociception opioid peptide
receptors (17). Therefore, the
mechanism by which JTC-801 functions through receptors requires
further experimental validation.
The results of the western blot analysis indicated
that JTC-801 may affect the expression of apoptosis-related
proteins in liver cancer. Apoptosis is an important antitumour
pathway and several antitumour drugs serve an important role in
cancer by inducing apoptosis (18,19). The
activation of apoptosis is regulated by multiple signalling
pathways, of which the PI3K/AKT pathway is one of the most
important (20). The PI3K/AKT pathway
regulates a number of malignant phenotypes, including
anti-apoptotic, cell growth and proliferation phenotypes (21,22). In
this pathway, AKT activation inhibits cell cycle arrest and
angiogenesis, and promotes tumour invasion and metastasis via
phosphorylation of the protein kinase mTOR (23,24).
Furthermore, P70S6K, which is closely associated with cell
proliferation, is located downstream of the PI3K/AKT/mTOR pathway
(25). Cyclin D1 has also been
reported to be a direct downstream target of the PI3K/AKT
signalling pathway (26). The
expression of cyclin D1 is increased in different types of tumour
tissues. The present western blot results demonstrated that the
phosphorylation levels of AKT and mTOR were decreased significantly
in JTC-801-treated Hep G2 cells. Similarly, the expression levels
of p70S6K and cyclin D1 were reduced following JTC-801 treatment.
These results suggest that JTC-801 may have an antitumour effect
via the regulation of apoptosis through the PI3K/AKT pathway.
In summary, the present study demonstrated that
JTC-801 inhibited cell proliferation, invasion and migration, and
promoted cell apoptosis through the PI3K/AKT signalling pathway in
hepatoblastoma Hep G2 cells. This study provides a basis for
further clinical research on JTC-801 in liver cancer treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets analysed during the present study are
available from the corresponding author on reasonable request.
Authors' contributions
BFZ designed the study and was involved in data
collection. TH analysed and interpreted the experimental data. Both
authors prepared figures, wrote the initial manuscript and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
GPCRs
|
G-protein-coupled receptors
|
|
PI3K
|
phosphatidylinositol 3-kinase
|
|
AKT
|
protein kinase B
|
|
mTOR
|
mechanistic target of rapamycin
|
|
Bcl-2
|
B-cell lymphoma 2
|
|
Bax
|
apoptosis regulator BAX
|
|
P70S6K
|
P70S6 kinase
|
|
CCK-8
|
Cell Counting Kit-8
|
|
FITC
|
fluorescein isothiocyanate
|
|
PI
|
propidium iodide
|
References
|
1
|
Pang Y, Kartsonaki C, Turnbull I, Guo Y,
Clarke R, Chen Y, Bragg F, Yang L, Bian Z, Millwood IY, et al:
Diabetes, plasma glucose and incidence of fatty liver, cirrhosis
and liver cancer: A prospective study of 0.5 million people.
Hepatology. 68:1308–1318. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lozano R, Naghavi M, Foreman K, Lim S,
Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, et
al: Global and regional mortality from 235 causes of death for 20
age groups in 1990 and 2010: A systematic analysis for the Global
Burden of Disease Study 2010. Lancet. 380:2095–2128. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Costentin CE, Layese R, Bourcier V, Cagnot
C, Marcellin P, Guyader D, Pol S, Larrey D, De Lédinghen V, Ouzan
D, et al: Compliance with hepatocellular carcinoma surveillance
guidelines associated with increased lead-time adjusted survival of
patients with compensated viral cirrhosis. Gastroenterology.
155:431–442. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boot A, Huang MN, Ng AWT, Ho SC, Lim JQ,
Kawakami Y, Chayama K, Teh BT, Nakagawa H and Rozen SG: In-depth
characterization of the cisplatin mutational signature in human
cell lines and in esophageal and liver tumors. Genome Res.
28:654–665. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hama N, Totoki Y, Miura F, Tatsuno K,
Saito-Adachi M, Nakamura H, Arai Y, Hosoda F, Urushidate T, Ohashi
S, et al: Epigenetic landscape influences the liver cancer genome
architecture. Nat Commun. 9:16432018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Collino A, Termanini A, Nicoli P, Diaferia
G, Polletti S, Recordati C, Castiglioni V, Caruso D, Mitro N,
Natoli G and Ghisletti S: Sustained activation of detoxification
pathways promotes liver carcinogenesis in response to chronic bile
acid-mediated damage. PLoS Genet. 14:e10073802018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li G, Liang Y, Sun C, Peng X, Hao N, Liu
M, Gao W, Wu H and He B: Effective combination therapy of
percutaneous ethanol injection and chemotherapy based on injectable
low molecular weight gels. Artif Cells Nanomed Biotechnol. 9:1–11.
2018. View Article : Google Scholar
|
|
8
|
Chaturvedi M, Schilling J, Beautrait A,
Bouvier M, Benovic JL and Shukla AK: Emerging paradigm of
intracellular targeting of G protein-coupled receptors. Trends
Biochem Sci. 43:533–546. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Renard D: Cerebral microbleeds: A magnetic
resonance imaging review of common and less common causes. Eur J
Neurol. 25:441–450. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tan M, Yamaguchi S, Nakamura M and
Nagamune T: Real-time monitoring of pH-dependent intracellular
trafficking of ovarian cancer G protein-coupled receptor 1 in
living leukocytes. J Biosci Bioeng. 126:363–370. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bedini A, Baiula M, Vincelli G, Formaggio
F, Lombardi S, Caprini M and Spampinato S: Nociceptin/orphanin FQ
antagonizes lipopolysaccharide-stimulated proliferation, migration
and inflammatory signaling in human glioblastoma U87 cells. Biochem
Pharmacol. 140:89–104. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song X, Zhu S, Xie Y, Liu J, Sun L, Zeng
D, Wang P, Ma X, Kroemer G, Bartlett DL, et al: JTC801 Induces
pH-dependent death specifically in cancer cells and slows growth of
tumors in mice. Gastroenterology. 154:1480–1493. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zheng CJ, Yang LL, Liu J and Zhong L:
JTC-801 exerts anti-proliferative effects in human osteosarcoma
cells by inducing apoptosis. J Recept Signal Transduct Res.
38:133–140. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Simpson-Durand CD and Standifer
KM: Nociceptin/orphanin FQ peptide receptor antagonist JTC-801
reverses pain and anxiety symptoms in a rat model of post-traumatic
stress disorder. Br J Pharmacol. 172:571–582. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Witkin JM, Statnick MA, Rorick-Kehn LM,
Pintar JE, Ansonoff M, Chen Y, Tucker RC and Ciccocioppo R: The
biology of Nociceptin/Orphanin FQ (N/OFQ) related to obesity,
stress, anxiety, mood, and drug dependence. Pharmacol Ther.
141:283–299. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li JX, Bi YP, Wang J, Yang X, Tian YF and
Sun ZF: JTC-801 inhibits the proliferation and metastasis of
ovarian cancer cell SKOV3 through inhibition of the PI3K-AKT
signaling pathway. Die Pharmazie. 73:283–287. 2018.PubMed/NCBI
|
|
17
|
da Silva JA, Biagioni AF, Almada RO, de
Freitas RL and Coimbra NC: Panicolytic-like effects caused by
substantia nigra pars reticulata pretreatment with low doses of
endomorphin-1 and high doses of CTOP or the NOP receptors
antagonist JTC-801 in male Rattus norvegicus. Psychopharmacology
(Berl). 234:3009–3025. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rybczynska AA, Boersma HH, de Jong S,
Gietema JA, Noordzij W, Dierckx RAJO, Elsinga PH and van Waarde A:
Avenues to molecular imaging of dying cells: Focus on cancer. Med
Res Rev. 38:1713–1768. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ucker DS and Levine JS: Exploitation of
apoptotic regulation in cancer. Front Immunol. 9:2412018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang C, Ma S, Hu R, Wang X, Li M, Tian F,
Jiang W, Zhu L and Bian Z: Effect of CXCR4 on apoptosis in
osteosarcoma cells via the PI3K/Akt/NF-κβ signaling pathway. Cell
Physiol Biochem. 46:2250–2260. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Carroll MJ, Fogg KC, Patel HA, Krause HB,
Mancha AS, Patankar MS, Weisman PS, Barroilhet L and Kreeger PK:
Alternatively activated macrophages upregulate mesothelial
expression of P-selectin to enhance adhesion of ovarian cancer
cells. Cancer Res. 78:3560–3573. 2018.PubMed/NCBI
|
|
22
|
De Santis MC, Porporato PE, Martini M and
Morandi A: Signaling pathways regulating redox balance in cancer
metabolism. Front Oncol. 8:1262018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jondal DA, Thompson SM, Butters KA,
Knudsen BE, Anderson JL, Carter RE, Roberts LR, Callstrom MR and
Woodrum DA: Heat stress and hepatic laser thermal ablation induce
hepatocellular carcinoma growth: Role of PI3K/mTOR/AKT signaling.
Radiology. 288:730–738. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim EJ, Kang GJ, Kang JI, Boo HJ, Hyun JW,
Koh YS, Chang WY, Kim YR, Kwon JM, Maeng YH, et al: Over-activation
of AKT signaling leading to 5-Fluorouracil resistance in
SNU-C5/5-FU cells. Oncotarget. 9:19911–19928. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo Y, Wang F, Li H, Liang H, Li Y, Gao Z
and He X: Metformin protects against spinal cord injury by
regulating autophagy via the mTOR signaling pathway. Neurochem Res.
2018.[Epub ahead of print]. View Article : Google Scholar
|
|
26
|
Yamazaki S, Higuchi Y, Ishibashi M,
Hashimoto H, Yasunaga M, Matsumura Y, Tsuchihara K, Tsuboi M, Goto
K, Ochiai A and Ishii G: Collagen type I induces EGFR-TKI
resistance in EGFR-mutated cancer cells via mTOR activation through
Akt-independent pathway. Cancer Sci. 109:2063–2073. 2018.
View Article : Google Scholar : PubMed/NCBI
|