Introduction
Osteosarcoma (OS) is one of the most common primary
malignant tumors of bone and the third most frequent cancer in
young adolescents and children (1).
In the last several decades, the treatments for OS have included
surgical resection, radiotherapy and combinatorial chemotherapy,
and, although the survival rates of patients with OS have improved,
aggressive invasion into local tissue and a markedly high
metastatic rate remain the primary causes of tumor-associated
mortality (2,3). Therefore, identification of novel
therapeutic approaches against specific targets to systematically
combat OS is an urgent requirement (4).
MicroRNAs (miRNAs) are a group of small non-coding
RNA molecules between 19 and 25 nucleotides in length that have
highly conserved sequences in plants and animals (5,6). By
binding to the 3′-untranslated region (UTR) of specific genes to
degrade mRNA or inhibit translation, miRNAs act as powerful
post-transcriptional regulators of gene expression and have
important functions in multicellular organisms (7). In various types of cancer, including OS,
miRNAs function as tumor suppressors or oncogenes as identified in
a number of studies (8–11). Among the non-coding RNAs, the Let7
miRNA family was the first known human miRNA, which consists of
Let7a/b/c/d/e/f/g/i and miR-98 (12,13). Let7
is involved in embryonic development and participates in the
proliferation and invasion of cancer cells and cancer stem cells
(14,15).
However, the function of Let-7b in OS is not fully
understood. Preliminary findings (Zhang et al, unpublished
data) suggested that decreased expression of Let-7b was observed in
OS cells; therefore, the aim of the present study was to
investigate the potential functions and mechanisms of Let-7b in OS
cells.
Materials and methods
Cell culture and transfection
The U2OS, MG63 and SAOS-2 human OS cell lines and
the human normal osteoblastic hFOB 1.19 cell line were purchased
from the American Type Culture Collection (Manassas, VA, USA). All
cell lines were cultured in Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 U/ml streptomycin
(Gibco; Thermo Fisher Scientific, Inc.). All cells were incubated
at 37°C in a humidified incubator containing 5% CO2. All
transfections were performed using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol.
Tissue collection
A total of 60 primary OS tissues and their matched
adjacent non-tumor tissues were collected by surgical resection
from patients who presented at the Department of Orthopedics, Linyi
Central Hospital (Linyi, China) between March 2010 and December
2012. The study was approved by the Ethics Committee of Linyi
Central Hospital. All patients included in the present study had
not received any other therapeutic treatment prior to surgery.
Written informed consent was obtained from each patient. The
tissues were immediately snap-frozen in liquid nitrogen following
surgical resection and stored at −80°C in liquid nitrogen until
use. The clinical characteristics of the patients are summarized in
Table I. The Tumor-Node-Metastasis
staging was performed according to Fukuma et al (16).
 | Table I.Clinicopathological characteristics of
the 60 patients with osteosarcoma. |
Table I.
Clinicopathological characteristics of
the 60 patients with osteosarcoma.
|
|
| Let-7b
expression |
|
|---|
|
|
|
|
|
|---|
| Characteristic | n | Low (n=36) | High (n=24) | P-value |
|---|
| Sex |
|
|
| 0.830 |
| Male | 36 | 22 | 14 |
|
|
Female | 24 | 14 | 10 |
|
| Age, years |
|
|
| 0.863 |
|
<18 | 36 | 24 | 12 |
|
| ≥18 | 34 | 22 | 12 |
|
| Tumor size, cm |
|
|
| 0.045 |
| ≥5 | 32 | 23 | 9 |
|
|
<5 | 28 | 13 | 15 |
|
| TNM stage |
|
|
| 0.001 |
| IIA | 29 | 11 | 18 |
|
|
IIB/III | 31 | 25 | 6 |
|
| Lung metastasis |
|
|
| 0.020 |
|
Yes | 31 | 23 | 8 |
|
| No | 29 | 13 | 16 |
|
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA from the tissues and cell lines were
extracted using TRIzol® reagent (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. A
total of 2 µg RNA was reverse-transcribed into cDNA using a
PrimeScript First Strand cDNA Synthesis kit (Takara Bio, Inc.,
Otsu, Japan), according to the manufacturer's protocol. qPCR was
performed on an ABI 7500 Real-time PCR detection system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The reaction
conditions were 95°C for 10 min, and 35 cycles of 95°C for 30 sec
and 60°C for 1 min. GADPH was used as the internal reference gene.
To analyze the levels of miRNA expression, qPCR was performed using
an All-in-One™ miRNA qRT-PCR kit (GeneCopoeia, Inc., Rockville, MD,
USA), according to the manufacturer's protocol. The U6 gene was
used as an internal control. The relative mRNA level was calculated
using the 2−∆∆Cq method (17). The primers used were as follows: Let7b
forward, 5′-TGAGGTAGTAGGTTGTGTGGTT-3′; and reverse,
5′-GCTGTCAACGATACGCTACCTA-3′; GAPDH forward,
5′-CCGTCTAGAAAAACCTGCC-3′; and reverse, 5′-GCCAAATTCGTTGTCATACC-3′;
U6, forward 5′-CTCGCTTCGGCAGCACA-3′; and reverse
5′-AACGCTTCACGAATTTGCGT-3′ (all GeneCopoeia, Inc.).
Western blot analysis
Proteins were extracted by lysing cells using
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) 72 h after transfection. The protein
concentration was quantified using a bicinchoninic acid assay kit
(Pierce; Thermo Fisher Scientific, Inc.). Equal amounts of proteins
(30 µg) were subjected to SDS-PAGE (10% gel) and transferred onto
polyvinylidene difluoride membranes (Thermo Fisher Scientific,
Inc.). The membranes were blocked with 5% non-fat milk powder in
PBS for 1 h at room temperature and was subsequently incubated with
primary rabbit polyclonal anti-IGF1R antibody (1:1,000 dilution;
cat. no. ab39675; Abcam, Cambridge, UK) and mouse monoclonal
anti-human GADPH antibody (1:1,000 dilution; cat. no. sc-59540;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) at 4°C overnight.
Following washing with Tris-buffered saline containing Tween-20
(0.5%) five times for 5 min each at room temperature, the membranes
were incubated with the corresponding horseradish
peroxidase-conjugated secondary antibodies goat anti-mouse IgG
(cat. no. sc-2005) and anti-rabbit IgG (cat. no. sc-2004) (both
1:3,000; Santa Cruz Biotechnology, Inc.) at room temperature for 1
h. The signals detected were visualized using an enhanced
chemiluminescence kit (Pierce; Thermo Fisher Scientific, Inc.).
Densitometric evaluation of the proteins was performed using ImageJ
(version 1.38; National Institutes of Health, Bethesda, MD,
USA).
Flow cytometry assay
U2OS or SAOS-2 cells were seeded at 3×105
per well in 6-well plates and cultured to 85% confluence to analyze
the cell cycle. Each group of cells was washed five times with PBS
at room temperature, then collected by centrifugation at 1,000 × g
for 5 min. All cells were resuspended in 500 µl PBS, and 5 µl
Annexin V-FITC and 10 µl PI (both Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) were added at room temperature for 15 min. The
flow cytometry assay was performed and analyzed by the FACSAria
cell sorter within 1 h of incubation using the FACSuite software
version 1.0 (both BD Biosciences, San Jose, CA, USA).
Cell proliferation assay
MTT assays (Sigma-Aldrich; Merck KGaA) were
performed to determine the proliferation of cells, according to the
manufacturer's protocol. To assess the proliferation rates,
1×106 cells in each group were seeded in 24-well plates
and transfected with Let-7b miRNA mimic alongside a negative
control (NC) (Let-7b mimic sequence, 5′-UGAGGUAGUAGGUUGUGUGGUU-3′;
NC sequence 5′-UUCUCCGAACGUGUCACGUTT-3′) at a final concentration
of 20 nM. After 24 h of transfection at 37°C, the cells were
collected and 3,000 cells/well were seeded in 96-well plates and
cultured for 0, 24, 48, 72 or 96 h. A total of 20 µl MTT solution
(5 mg/ml) was added to each well and the cells were incubated for a
further 4 h at 37°C. Next, the supernatant was removed and 200 µl
dimethylsulfoxide was added to each well for 10 min at 37°C to
dissolve the formazan product. The absorbance was determined using
a Model 680 microplate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) at a wavelength of 570 nm.
Invasion assay
Transwell assays were used to evaluate the invasive
ability of OS cells. The Transwell chambers (8 µm pore size;
Costar; Corning Incorporated, Corning, NY, USA) were coated with 60
µl Matrigel (BD Biosciences, San Jose, CA, USA), according to the
manufacturer's protocol. The cells were collected 48 h after
transfection and 2.5×105 cells per well were resuspended
in 200 µl DMEM without FBS and then added into the upper chambers.
The lower chambers were filled with 800 µl DMEM supplemented with
20% FBS. Following incubation for 24 h at 37°C, cells on the upper
surface of the membrane were carefully removed with cotton swabs.
The invading cells on the bottom of the membrane were fixed with
100% methanol for 10 min and stained with 0.5% crystal violet for
20 min at room temperature. The cells in five randomly selected
fields were enumerated using an inverted microscope (CKX41; Olympus
Corporation, Tokyo, Japan). All experiments were performed at least
three times.
Bioinformatics analysis
Targetscan software (version 5.1; www.targetscan.org) was used to predicate the putative
targets of Let-7b, according to the manufacturer's protocol.
Dual-luciferase reporter assay
Luciferase activities were determined using a
Dual-Luciferase Reporter assay system (Promega Corporation,
Madison, WI, USA), according to the manufacturer's protocol.
PGL3-IGF1R-3′UTR wild-type (WT) and PGL3-IGF1R-3′UTR mutant (Mut)
were obtained from Shanghai GenePharma Co., Ltd. (Shanghai, China).
U2OS or Saos-2 cells were seeded in 96-well plates, co-transfected
with the WT-IGF1R-3′UTR or Mut-IGF1R-3′UTR luciferase reporter
plasmid and transfected with the Let-7b mimic or NC mimic
respectively. After 48 h, luciferase activity was determined, and
firefly luciferase activity was normalized to Renilla
luciferase activity. Luciferase assays were performed in three
independent experiments.
Statistical analysis
All statistical analysis was performed using SPSS
(version 19.0; IBM SPSS, Inc., Armonk, NY, USA). Results are
expressed as the mean ± standard deviation of three independent
experiments. Student's t-test was used to analyze the difference
between two groups. One-way analysis of variance followed by
Tukey's test was used to analyze the differences between multiple
groups. Spearman's correlation analysis was used to investigate the
correlation between IGF1R mRNA and Let-7b in T1-T2 stage
osteosarcoma. P<0.05 was considered to indicate a statistically
significant difference. The associations between the expression of
Let-7b and the clinical characteristics of patients with OS were
analyzed using a χ2 test.
Results
Let-7b is downregulated in OS cell
lines and tissues
To reveal the function of Let-7b in OS progression,
RT-qPCR analysis was used to explore the expression of Let-7b in
the human OS cell lines U2OS, MG-63 and SAOS-2, and the human
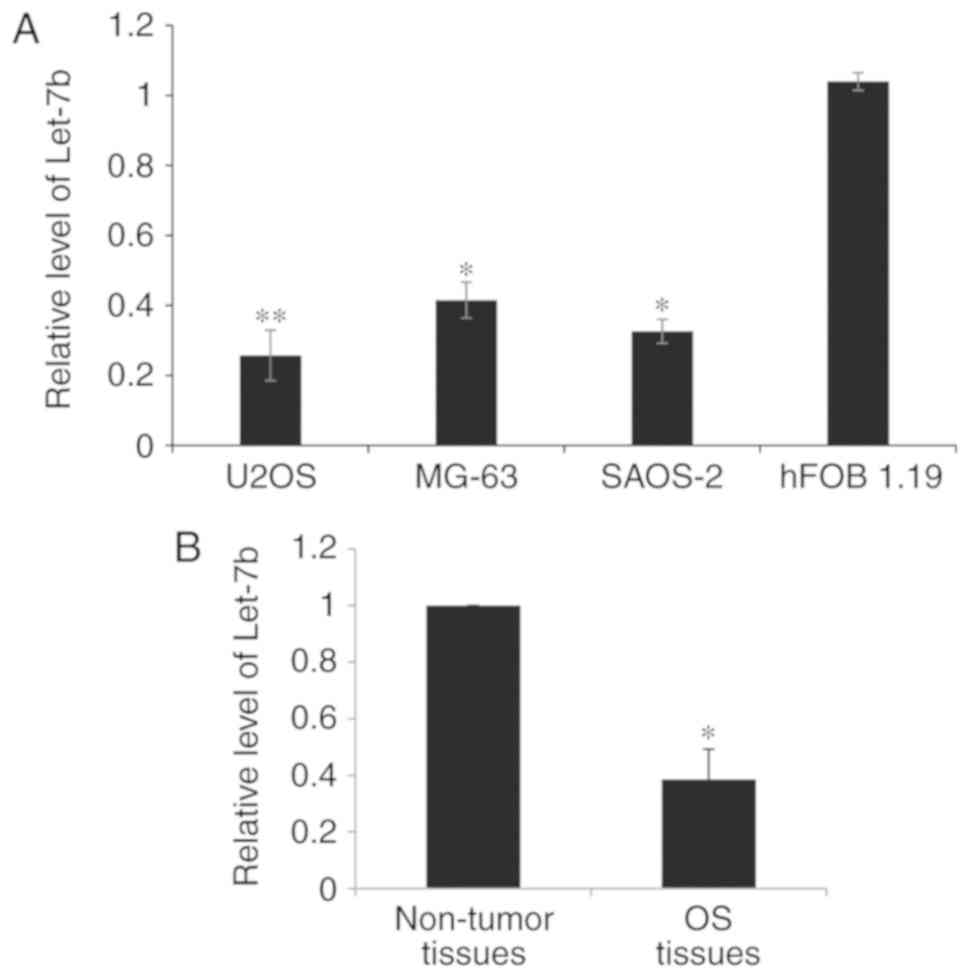
normal osteoblastic hFOB 1.19 cell line. As presented in Fig. 1A, the expression of Let-7b was
significantly decreased in the OS cell lines. The clinical
significance of Let-7b expression in OS was investigated in a total
of 60 paired OS tissues and adjacent non-tumor tissues. It was
observed that the Let-7b level was significantly lower in the OS
tissues compared with the adjacent non-tumor tissues (Fig. 1B). On the basis of the mean value of
Let-7b expression in patients with OS, tissues were divided into
high and low Let-7b expression groups. As presented in Table I, low expression of Let-7b was
significantly associated with increased tumor size, advanced
Tumor-Node-Metastasis stage (16) and
lung metastasis (P<0.05; Table I).
However, there was no significant difference between Let-7b
expression according to age or sex (Table
I). These results indicated that Let-7b is downregulated in OS
cells and tissues and therefore that Let-7b may serve a function in
inhibiting the development of OS in humans.
Let-7b inhibits the proliferation of
OS cells
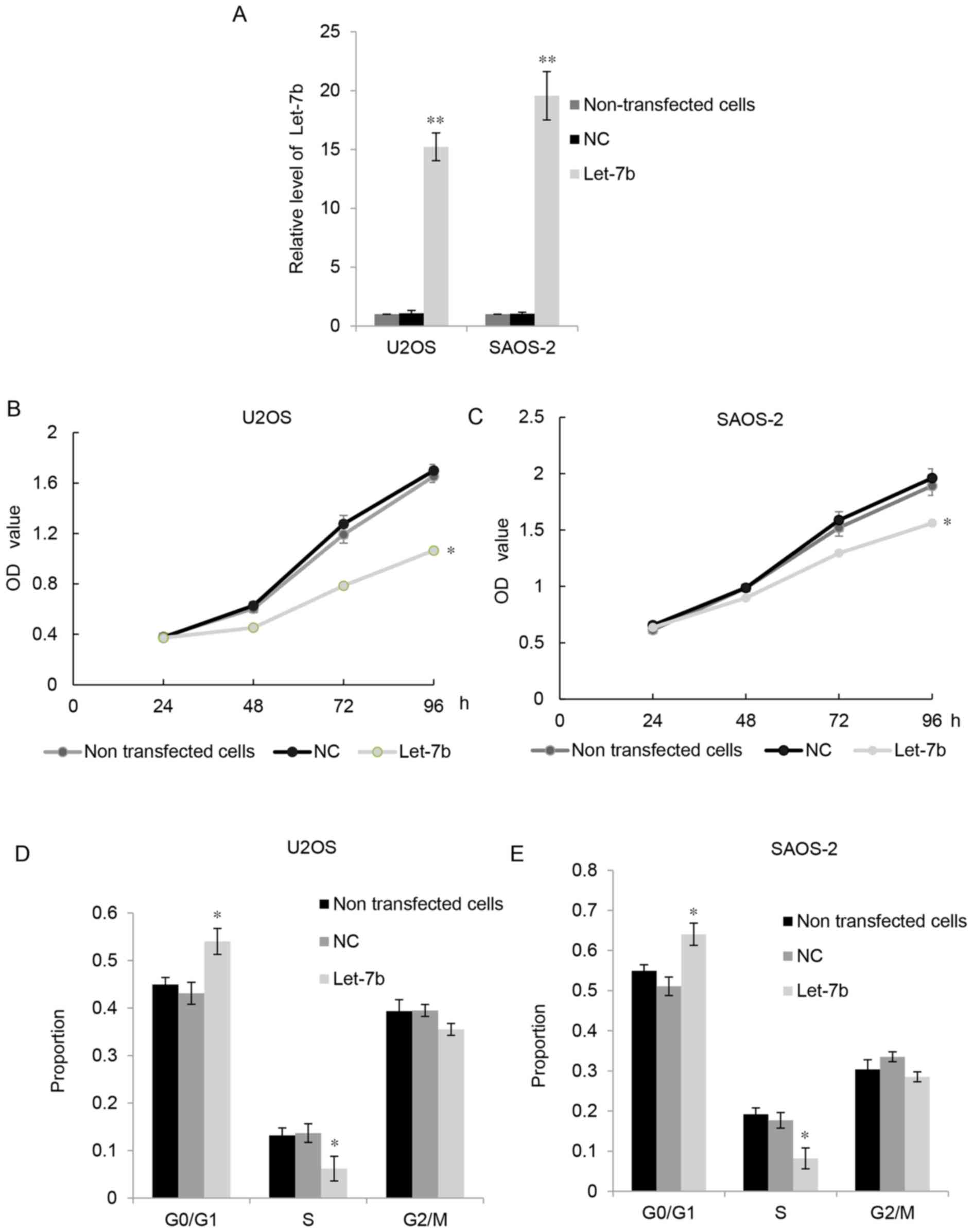
As Let-7b was downregulated in OS cell lines, U2OS
and SAOS-2 cells were transfected with Let-7b. RT-qPCR indicated
that, compared with the control groups, transfection with plasmids
expressing Let-7b significantly upregulated Let-7b levels in U2OS
and MG63 cells (Fig. 2A). MTT assays
were performed to examine the proliferation of OS cells. As
presented in Fig. 2B and C, the
overexpression of Let-7b significantly suppressed the proliferation
of U2OS and SAOS-2 cells compared with that of the control groups.
Furthermore, flow cytometry was used to examine the cell cycle
distribution of the U2OS and SAOS-2 cells. The results indicated
that overexpression of Let-7b led to markedly high levels of cell
cycle arrest at G1 stage (Fig.
2D and E). These results suggested that Let-7b exhibited
suppressive effects on the proliferation of OS cells.
Let-7b suppresses the invasion of OS
cells in vitro
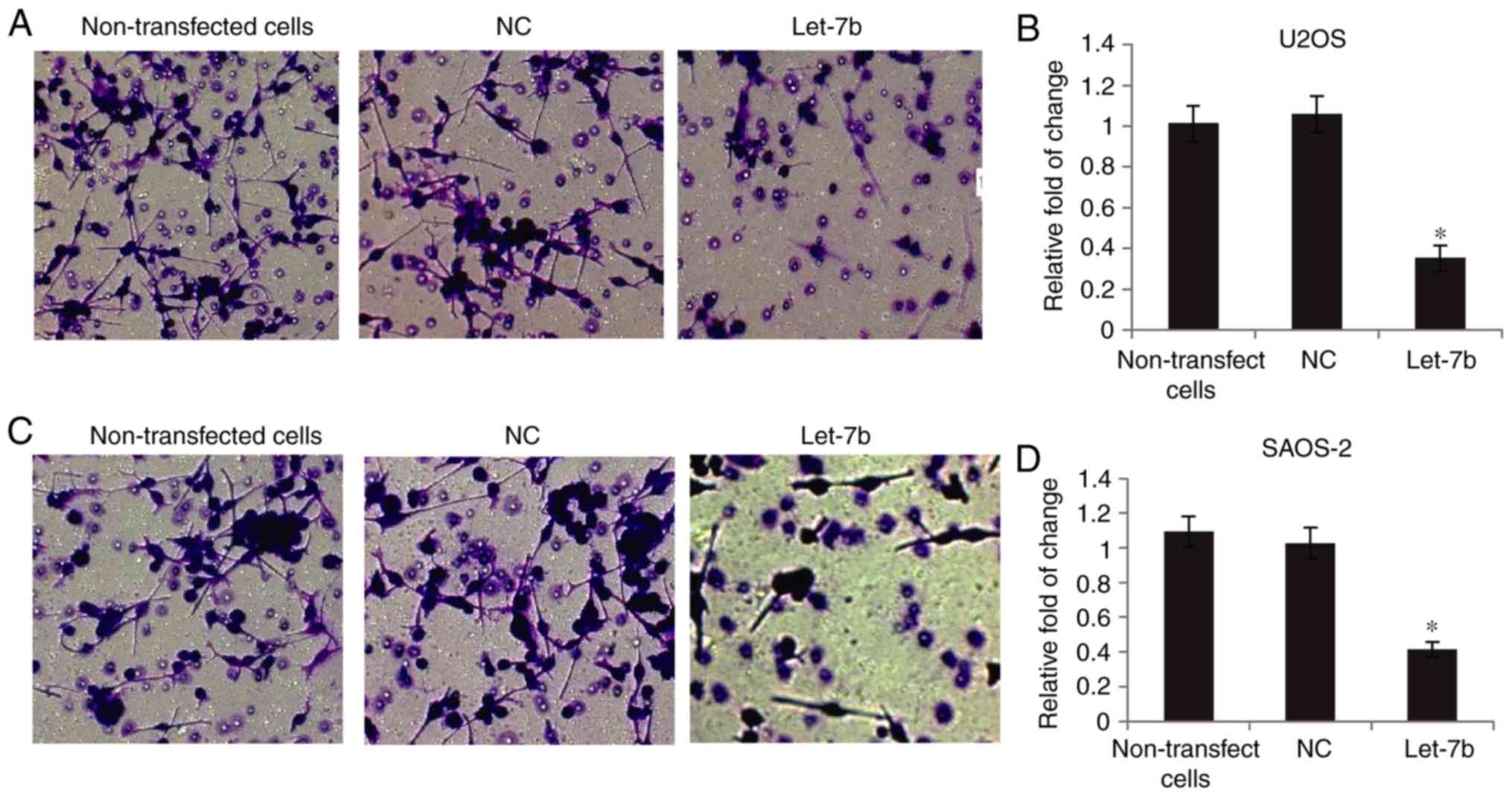
The invasive capability of OS cells was determined
using Transwell assays. As presented in Fig. 3A and B, overexpression of Let-7b
markedly suppressed U2OS invasion, compared with the
non-transfected cells, whereas transfection with NC did not affect
their invasion. Furthermore, SAOS-2 cell invasion was markedly
decreased following overexpression of Let-7b (Fig. 3C and D). Together, these results
suggested that Let-7b significantly inhibited the invasive ability
of OS cells.
IGF1R is the direct target of
Let-7b
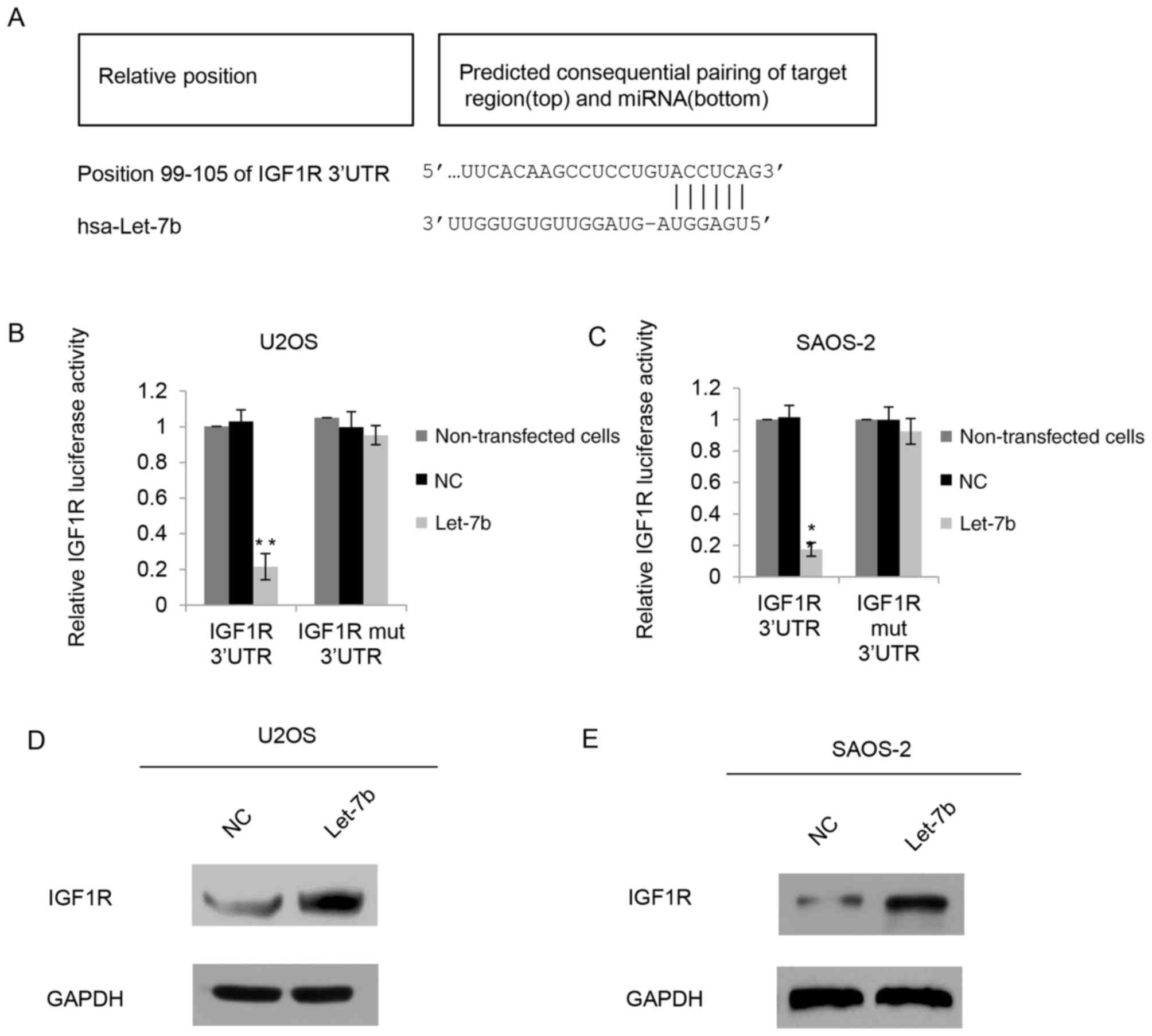
The function of miRNAs is to inhibit the expression
of their target genes; therefore, the potential targets of Let-7b
were investigated using Targetscan software. IGF1R was identified
to be a putative target of Let-7b (Fig.
4A). To further identify the mechanisms of Let-7b affecting the
biological behavior in OS, a luciferase activity assay was
performed. A luciferase reporter vector containing the IGF1R 3′-UTR
or a IGF1R mutant 3′-UTR was constructed, and the constructs were
co-transfected with Let-7b or NC into U2OS cells. As presented in
Fig. 4B, Let-7b significantly
inhibited the luciferase activity of the IGF1R 3′-UTR reporter
compared with that of the control groups, whereas the IGF1R mutant
3′-UTR eliminated the effect of Let-7b on luciferase activity, as
there was no significant difference between the effects of the
mutation and the control groups. In the SAOS-2 cells, a similar
tendency was exhibited (Fig. 4C).
Since miRNAs generally inhibit the expression of target genes at
the post-transcriptional level, IGF1R protein expression was also
determined in U2OS cells following Let-7b overexpression. The
results from the western blot analysis identified that
overexpression of Let-7b decreased the expression of endogenous
IGF1R (Fig. 4D). Similarly, in SAOS-2
cells, IGF1R protein levels were decreased in the Let-7b
transfection groups (Fig. 4E).
Consistent with the luciferase reporter data, these results
indicated that Let-7b bound directly to the 3-UTR of IGF1R, and
therefore negatively regulated the expression of IGF1R.
IGF1R is involved in Let-7b-mediated
proliferation and invasion of OS cells
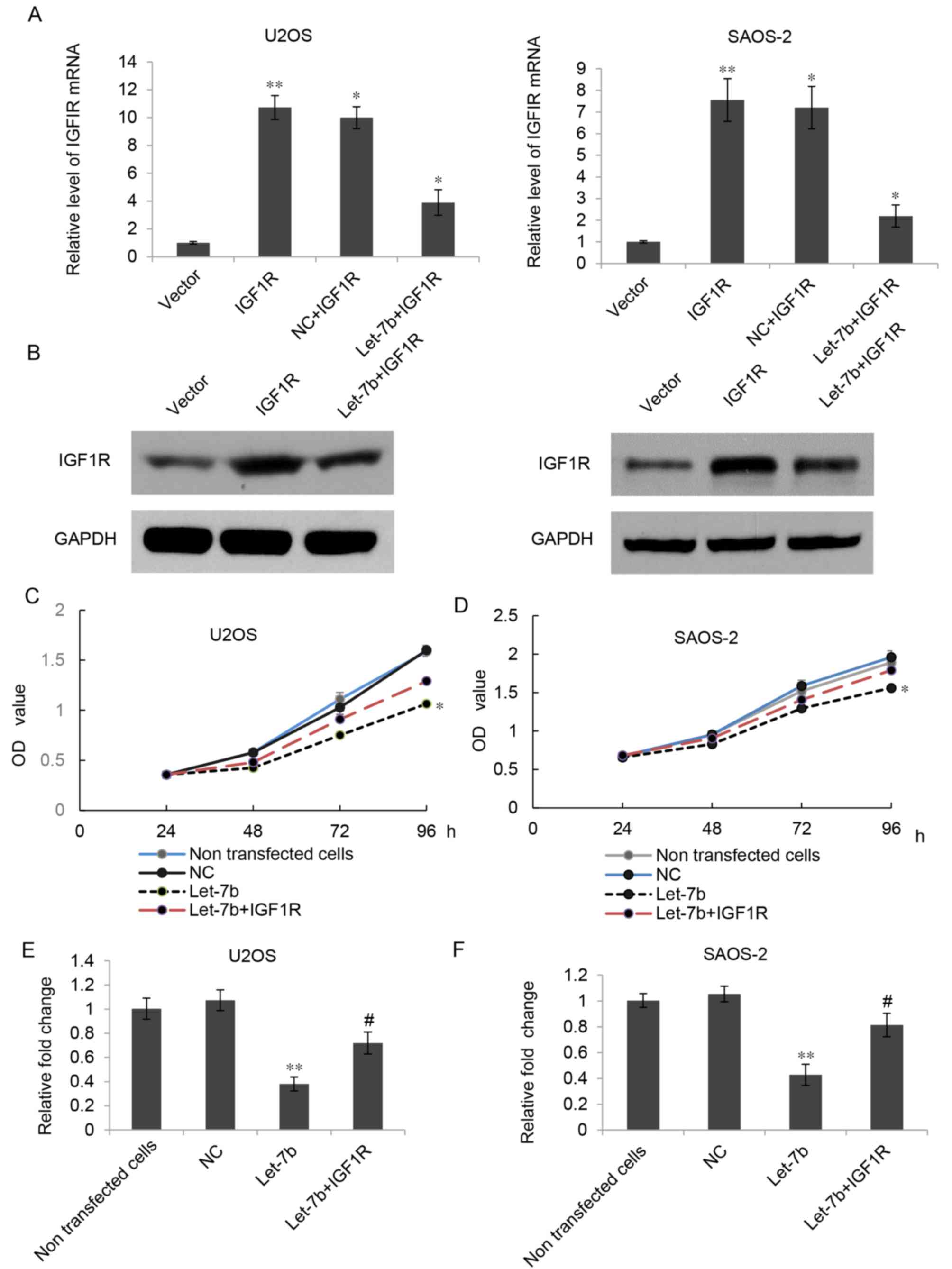
As IGF1R has been identified to be an oncogene in
OS, and overexpression of Let-7b led to downregulation of IGF1R, it
was hypothesized that Let-7b may suppress the proliferation and
invasion of OS cells by regulating IGF1R. To verify this
hypothesis, Let-7b-overexpressing U2OS and SAOS-2 cells were
further transfected with a pcDNA3.1-IGF1R plasmid or a pcDNA3.1
vector. As presented in Fig. 5A and
B, the mRNA and protein expression of IGF1R was markedly
increased in the Let-7b+IGF1R group compared with that in the
Let-7b group; however, transfection with the pcDNA3.1 vector did
not affect the expression of IGF1R. The results of the MTT assay
further indicated that the proliferation of U2OS and SAOS-2 cells
was significantly increased in the Let-7b+IGF1R group when compared
with that in the Let-7b group (Fig. 5C
and D). A Transwell assay indicated that the overexpression of
IGF1R could partially reverse the effects of Let-7b on the invasion
of U2OS and SAOS-2 cells (Fig. 5E and
F). Accordingly, these results suggested that IGF1R was indeed
involved in the Let-7b-mediated suppressive effects in the
proliferation and invasion of U2OS and SAOS-2 cells.
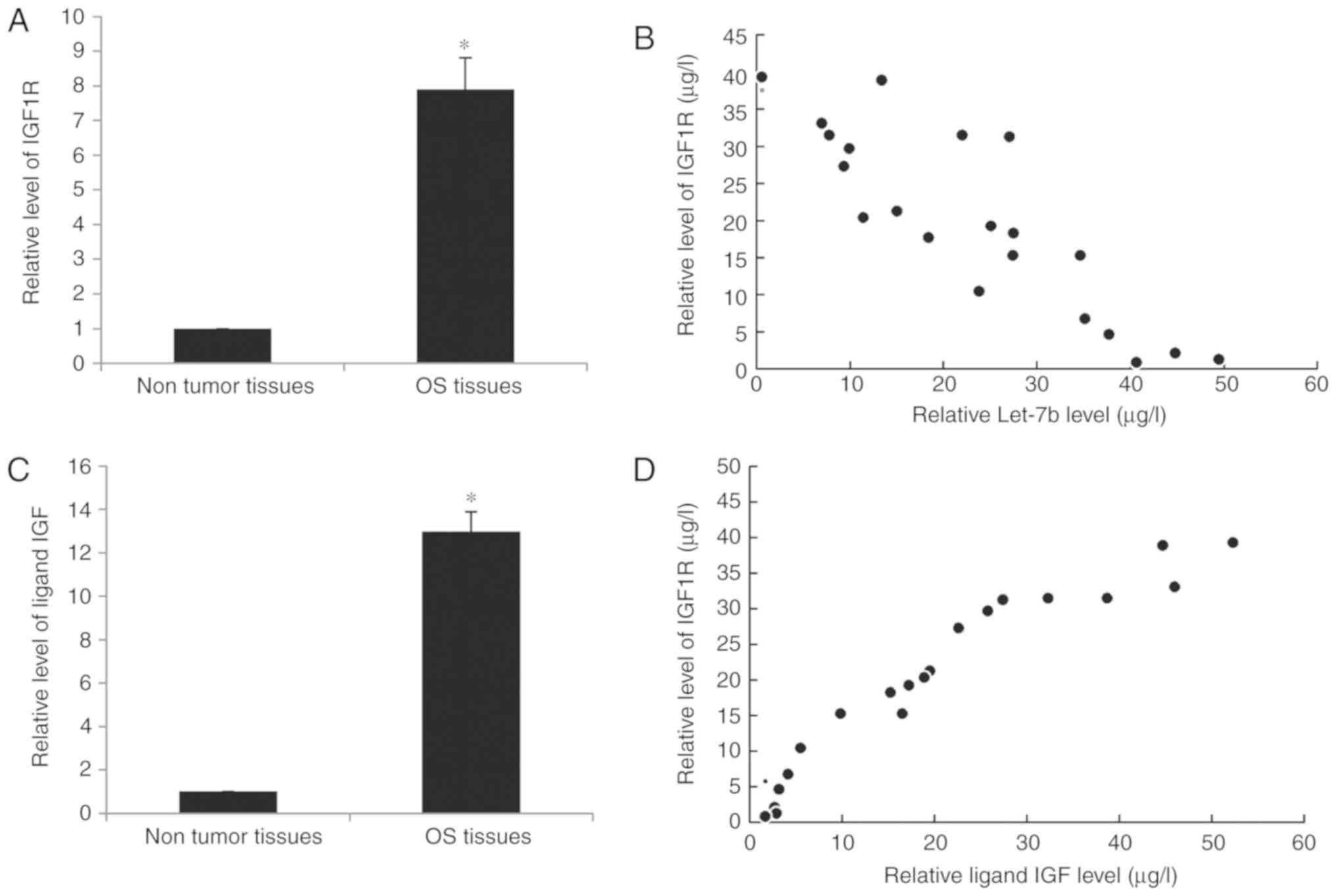
Increased IGF1R levels are inversely
associated with Let-7b in OS tissues
Finally, the expression of IGF1R in human OS tissues
was investigated. RT-qPCR results indicated that IGF1R was
significantly increased in OS tissues compared with the adjacent
non-tumorous tissues (Fig. 6A).
Furthermore, a significant inverse correlation was identified
between the expression of Let-7b and IGF1R in OS at T1-T2 stage
(Fig. 6B), suggesting that the
increased expression of IGF1R may be due to the downregulation of
Let-7b. Furthermore, the expression of ligand insulin-like growth
factor (IGF) in the serum of patients with OS was also upregulated
(Fig. 6C) and exhibited a positive
correlation with IGF1R (Fig. 6D),
which represented an increase in IGF signal transduction.
Discussion
It has been identified previously that a number of
oncogenes and tumor suppressors serve key functions in the
progression of OS (18). Therefore,
understanding the underlying molecular mechanisms of OS may enable
the development of effective therapeutic strategies (19).
IGF1R is a widely expressed cell membrane receptor
and it is involved in different cancer types (20,21). As
identified previously, increased expression of IGF1R in OS is
associated with the development of tumor metastasis and indicates a
poor prognosis, and such molecules usually represent an attractive
therapeutic target (22,23). These results are consistent with the
results of the present study that IGF1R is significantly increased
in OS tissues compared with the adjacent non-tumorous tissues. By
targeting cadherin 2 and IGF1R, miR-194 suppressed OS cell
proliferation and metastasis in vitro and in vivo
(24). miR-133a inhibited OS cellular
proliferation and invasion via targeting IGF1R (25). miR-16 inhibited OS cell proliferation
by targeting IGF1R (9). In the
present study, it was identified that Let-7b was another regulator
for IGF1R in OS. Let-7b bound directly to the 3-UTR of IGF1R, and
therefore negatively regulated the expression of IGF1R.
A single miRNA is able to silence the expression of
multiple genes simultaneously. It is reasonable to propose the
hypothesis that, in addition to IGF1R, additional target genes are
involved in suppression of OS by Let-7b. Using TargetScan, putative
targets of Let-7b were identified, of which several were associated
with tumor growth or metastasis. These predictions were the
experimental basis for the present study of the regulatory
mechanism of Let-7b.
In conclusion, the results of the present study
indicated that Let-7b suppressed the proliferation and invasion of
OS cells in vitro. Overexpression of Let-7b downregulated
the protein level of IGF-IR, and there was a significant inverse
correlation between the expression of Let-7b and IGF1R in OS at
T1-T2 stage, which suggested the function of Let-7b as a tumor
suppressor in OS progression by repression of the expression of
IGF1R. In the future, investigating further putative target genes
and understanding the molecular mechanisms of the contribution of
Let-7b to OS are required. Let-7b may prove to be a promising gene
therapeutic agent.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
KZ conceived and designed the study. WW and YL
performed transwell and MTT assays. AG analyzed the data. DY
performed tissue collection and wrote the paper. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Linyi Central Hospital (Linyi, China). Written
informed consent was obtained from each patient.
Patient consent for publication
Patients provided consent for the publication of the
data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Valery PC, Laversanne M and Bray F: Bone
cancer incidence by morphological subtype: A global assessment.
Cancer Causes Control. 26:1127–1139. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the surveillance, epidemiology, and end results program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Broadhead ML, Clark JC, Myers DE, Dass CR
and Choong PF: The molecular pathogenesis of osteosarcoma: A
review. Sarcoma. 2011:9592482011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chou AJ, Geller DS and Gorlick R: Therapy
for osteosarcoma: Where do we go from here? Paediatric Drugs.
10:315–327. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Developmental Cell. 11:441–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liang J, Liu C, Qiao A, Cui Y, Zhang H,
Cui A, Zhang S, Yang Y, Xiao X, Chen Y, et al: MicroRNA-29a-c
decrease fasting blood glucose levels by negatively regulating
hepatic gluconeogenesis. J Hepatol. 58:535–542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen L, Wang Q, Wang GD, Wang HS, Huang Y,
Liu XM and Cai XH: miR-16 inhibits cell proliferation by targeting
IGF1R and the Raf1-MEK1/2-ERK1/2 pathway in osteosarcoma. FEBS
Lett. 587:1366–1372. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ji F, Zhang H, Wang Y, Li M, Xu W, Kang Y,
Wang Z, Wang Z, Cheng P, Tong D, et al: MicroRNA-133a,
downregulated in osteosarcoma, suppresses proliferation and
promotes apoptosis by targeting Bcl-xL and Mcl-1. Bone. 56:220–226.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim HR, Shin CH, Lee H, Choi KH, Nam DH,
Ohn T and Kim HH: MicroRNA-1908-5p contributes to the oncogenic
function of the splicing factor SRSF3. Oncotarget. 8:8342–8355.
2017.PubMed/NCBI
|
|
12
|
Reinhart BJ, Slack FJ, Basson M,
Pasquinelli AE, Bettinger JC, Rougvie AE, Horvitz HR and Ruvkun G:
The 21-nucleotide let-7 RNA regulates developmental timing in
Caenorhabditis elegans. Nature. 403:901–906. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pasquinelli AE, Reinhart BJ, Slack F,
Martindale MQ, Kuroda MI, Maller B, Hayward DC, Ball EE, Degnan B,
Müller P, et al: Conservation of the sequence and temporal
expression of let-7 heterochronic regulatory RNA. Nature.
408:86–89. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lan FF, Wang H, Chen YC, Chan C, Ng SS, Li
K, Xie D, He ML, Lin MC and Kung HF: Hsa-let-7g inhibits
proliferation of hepatocellular carcinoma cells by downregulation
of c-Myc and upregulation of p16(INK4A). Int J Cancer. 128:319–331.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao C, Sun G, Li S, Lang MF, Yang S, Li W
and Shi Y: MicroRNA let-7b regulates neural stem cell proliferation
and differentiation by targeting nuclear receptor TLX signaling.
Proc Nati Acad Sci USA. 107:1876–1881. 2010. View Article : Google Scholar
|
|
16
|
Fukuma H, Beppu Y, Yokoyama R and Omiya K:
TNM classification of bone and soft tissue sarcomas. Gan to kagaku
Ryoho (Japanese). 24:1035–1040. 1997.
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Leichter AL, Sullivan MJ, Eccles MR and
Chatterjee A: MicroRNA expression patterns and signalling pathways
in the development and progression of childhood solid tumours. Mol
Cancer. 16:152017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tang N, Song WX, Luo J, Haydon RC and He
TC: Osteosarcoma development and stem cell differentiation. Clin
Orthop Relat Res. 466:2114–2130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li S, Ferber A, Miura M and Baserga R:
Mitogenicity and transforming activity of the insulin-like growth
factor-I receptor with mutations in the tyrosine kinase domain. J
Biol Chem. 269:32558–32564. 1994.PubMed/NCBI
|
|
21
|
Kirschner LS: Inhibition of IGF-1R in
adrenocortical carcinoma. Lancet Oncol. 16:356–357. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maniscalco L, Iussich S, Morello E,
Martano M, Gattino F, Miretti S, Biolatti B, Accornero P,
Martignani E, Sánchez-Céspedes R, et al: Increased expression of
insulin-like growth factor-1 receptor is correlated with worse
survival in canine appendicular osteosarcoma. Vet J. 205:272–280.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cao Y, Roth M, Piperdi S, Montoya K,
Sowers R, Rao P, Geller D, Houghton P, Kolb EA, Gill J and Gorlick
R: Insulin-like growth factor 1 receptor and response to anti-IGF1R
antibody therapy in osteosarcoma. PLoS One. 9:e1062492014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Han K, Zhao T, Chen X, Bian N, Yang T, Ma
Q, Cai C, Fan Q, Zhou Y and Ma B: MicroRNA-194 suppresses
osteosarcoma cell proliferation and metastasis in vitro and in vivo
by targeting CDH2 and IGF1R. Int J Oncol. 45:1437–1449. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen G, Fang T, Huang Z, Qi Y, Du S, Di T,
Lei Z, Zhang X and Yan W: MicroRNA-133a inhibits osteosarcoma cells
proliferation and invasion via targeting IGF-1R. Cell Physiol
Biochem. 38:598–608. 2016. View Article : Google Scholar : PubMed/NCBI
|