Introduction
Osteosarcoma (OS), the most frequently occurring
primary malignant bone tumor in children and adolescents, normally
occurs in the metaphysis of the long bones and is characterized by
a high level of relapse and a poor prognosis (1). The estimated worldwide annual incidence
of OS is ~4 million cases, with the highest prevalence occurring in
adolescents between the ages of 15 and 19 years (2). Despite the combination of adjuvant
chemotherapy and advanced surgery, the cumulative 5-year survival
rate of patients is only 55–68, and ~80% of patients eventually
develop metastatic disease and face a poor outcome (3,4).
Therefore, it is imperative to investigate the potential molecular
mechanisms of OS progression and seek an effective molecular
therapy for the treatment of the disease.
MicroRNAs (miRNAs/miRs) are a small class of
endogenous non-coding RNAs that range in length from 18 to 25
nucleotides. miRNAs regulate gene expression at the
post-transcriptional level by binding to partially complementary
sites located in the 3′-untranslated regions (3′-UTR) of target
messenger RNAs (mRNAs) (5–7). In cancer, numerous miRNAs locate within
or close to fragile chromosomal sites that are often lost or
amplified (8). Increasing evidence
has suggested that miRNAs serve a crucial role in specific cellular
processes, including differentiation, morphogenesis and
tumorigenesis. Thus, these key effects demonstrate that miRNAs may
contribute to the regulation of the pathological development of
numerous diseases, particularly in human cancer (9,10).
Furthermore, in OS, aberrant expression of miRNA has been
demonstrated to contribute towards the development and metastasis
of the cancer by promoting the expression of oncogenes or by
inhibiting tumor suppressor genes.
It was recently reported that miR-885-5p is
aberrantly upregulated in the liver metastases of colorectal
cancer, renal cell carcinoma and Hürthle cell thyroid carcinoma
(11–13). By contrast, another study reported
that the expression of miR-885-5p is negatively correlated with the
malignant progression of human hepatocellular carcinoma (HCC)
tissue samples and cell lines via targeting of the catenin β1 gene,
leading to decreased activity of the Wnt/β-catenin signaling
pathway (14). Recently, we searched
for GEO databases regarding miR-885-5p expression and one dataset,
GSE65071, indicated that miR-885-5p was aberrantly expressed in the
plasma of patients with OS compared with that in normal individuals
(15). However, the biological role
of miR-885-5p and its molecular mechanism in OS remains
unknown.
In the present study, the expression of miR-885-5p
in OS cells and tissues was investigated, and the biological
behavior of miR-885-5p in OS cells was observed. In addition,
target genes were predicted by bioinformatics analysis to clarify
the role of miR-885-5p in the development of osteosarcoma.
Materials and methods
Human tissue
Paired human OS tissues (prior to the administration
of neoadjuvant chemotherapy) and matched adjacent normal tissues
were surgically acquired from patients (mean age, 22.19 years) at
the Department of Orthopedics, First Affiliated Hospital of Fujian
Medical University (Fuzhou, Fujian, China) between September 2015
and August 2017. The distance between the normal tissue and the
tumor was ~5 cm. Following surgical removal, all the tissues were
snap-frozen in liquid nitrogen and stored at −80°C until further
use. All clinical samples were collected with written informed
consent from the patients, and the study was approved by the
Ethical Committee of the First Affiliated Hospital of Fujian
Medical University.
Cell culture
The normal human osteoblastic hFOB1.19 cell line,
and the human OS MG-63 and U2OS cell lines (Shanghai Cell Bank,
Chinese Academy of Sciences, Shanghai, China) were cultured in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc. Waltham, MA, USA) with 10% fetal bovine serum
(FBS; Hyclone; GE Healthcare Life Sciences, Logan, UT, USA), 100
U/ml penicillin and 100 mg/ml streptomycin in a humidified
incubator with 5% CO2 at 37°C.
Transfection and generation of stable
cell lines
Recombinant lentivirus containing miR-885-5p mimic
(miR-885-5p-up;
5′-UCCAUUACACUACCCUGCCUCUAGGCAGGGUAGUGUAAUGGAUU-3′), miR-885-5p
inhibitor (miR-885-5p-down; 5-AGAGGCAGGGUAGUGUAAUGGA-3′) and
negative control (NC; 5′-CAGUACUUUGUGUAGUACAA-3′) were synthesized
by GenePharma (Shanghai, China) to generate either upregulated or
downregulated expression of miR-885-5p. The MG-63 cell line was
selected for following experiments as it has previously been used
to confirm a correlation between OS and miRNA, and MG-63 cells are
easily transfected for stable expression of miRNA (16). For transfection, 20 nM miRNA were
transfected into MG-63 cells using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.).
To generate stable cell lines with
miR-885-5p-up/down, a promoter-driven green fluorescent protein
(GFP) reporter was used to detect the transfection efficiency. The
cells were selected with 0.5 µg/ml puromycin for 2 weeks. When the
GFP expression was observed up to 80%, MG-63 cells stably
expressing miR-885-5p-up, miR-885-5p-down and NC were used in the
following cell experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from frozen tissues and
cultured cell lines using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). The concentration and purity of total RNA were
identified by ultraviolet spectrophotometry and
gel-electrophoresis, and reverse transcribed into cDNA using the
PrimeScript reverse transcription kit (Takara Bio, Inc., Otsu,
Japan). RT-qPCR was performed using SYBR Green Supermix (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) and analyzed on an ABI 7500
thermocycler (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The thermocycling conditions were as follows: 95°C for 5 min, 45
cycles of 95°C for 15 sec and 60°C for 45 sec. The relative
expression levels of miR-885-5p (forward,
5′-GTCCATTACACTACCCTGCCTC-3′ and reverse,
5′-CGCGAGCACAGAATTAATACG-3′) were evaluated and normalized to that
of U6 (forward, 5′-CTCGTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′) using the 2−ΔΔCq method
(17).
3-(4,5-dimethyl-2-thiazol)-2,5-diphenyl-2-H-tetrazolium bro-mide
(MTT) and colony formation assays
The MTT assay was used to evaluate cell
proliferation, according to the manufacturer's protocols.
Transfected MG-63 cells (five replicates) were plated at a density
of 5×103 cells per well in 96-well culture plates for 24
h. Subsequent to 24, 48 or 72 h of incubation, 20 µl MTT (5 mg/ml;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added to each
well for an additional incubation period of 4 h. Subsequently, the
MTT solution was discarded and 150 µl dimethyl sulfoxide
(Sigma-Aldrich; Merck KGaA) was added into each well and the plate
was agitated gently for 10 min. The absorbance was measured using a
microplate reader SpectraMax i3× (Molecular Devices, LLC,
Sunnyvale, CA, USA) at a wavelength of 490 nm.
For the colony formation assay, cells were seeded in
a 12-well plate at 500 cells per well (three replicates).
Subsequent to 2 weeks of incubation at 37°C the colonies were fixed
with methanol for 30 min at room temperature and stained with 0.5%
crystal violet for 30 min at room temperature. Each experiment was
repeated five times.
Transwell migration assay
Migration assays were performed in triplicate using
a 24-well Transwell chamber (Costar 3422; Corning Inc., Corning,
NY, USA). MG-63 cells transfected with miR-885-5p-up,
miR-885-5p-down and NC, respectively, were seeded in the upper
chamber at 2×104 cells per well in serum-free DMEM. DMEM
supplemented with 10% FBS was added to the bottom well at a volume
of 0.6 ml. Following incubation for 24 h, the cells on the top
filter were gently removed with cotton swabs and cells that had
migrated to the lower surface were fixed with methanol for 30 min
at room temperature and stained with 0.5% crystal violet for 30 min
at room temperature. The values for migration were obtained by
counting three fields under a light microscope. The experiment was
repeated three times over multiple days.
Luciferase activity assay
MG-63 cells were seeded in 24-well plates 1 day
prior to transfection, followed by co-transfection with constructed
pGL3 vectors (GenePharma) containing the wild-type 3′-UTR of CDC73
or its 3′-UTR mutant, and miR-885-5p mimic or the miRNA NC using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Cells were harvested for 36 h after transfection
and luciferase activity was measured by the Dual-Luciferase assay
(Promega Corporation, Madison, WI, USA), according to the
manufacturer's protocol. The firefly luciferase activity was then
normalized to the Renilla luciferase activity. Each
experiment was performed in duplicate three times.
Western blotting
Total protein was isolated from cells using RIPA
lysis buffer (Beyotime Institute of Biotechnology, Haimen, China)
and the protein concentrations were measured using the
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology). A total of 20 µg of protein were resolved with 10%
SDS-PAGE gel, transferred to polyvinylidene difluoride membranes
and blocked in 5% skimmed dry milk in Tris-buffered saline (pH
7.4), containing 0.05% Tween-20. Subsequently, the membrane was
incubated with anti-CDC73 antibody (catalog no. PB0587; 1:1,000;
Boster Biological Technology, Wuhan, China) overnight at 4°C and
then with anti-GAPDH (catalog no. BM3876; 1:3,000; Boster
Biological Technology) for 2 h at room temperature as a loading
control. Signals were detected by secondary antibodies labeled with
horseradish peroxidase (HRP) and were visualized using an enhanced
chemiluminescent kit (Beyotime Institute of Biotechnology).
Statistical analysis
Data are presented as the mean ± standard deviation
for at least three independent experiments. One-way analysis of
variance or an unpaired Student's t-test was used for the
statistical analysis. The association between clinicopathological
features and miR-885-5p was estimated by Fisher's exact test. The
association between CDC73 and miR-885p expression was explored by
Pearson's correlation coefficient. P<0.05 was considered to be
statistically significant. All analyses were performed using SPSS
version 23.0 (IBM, Corp., Armonk, NY, USA).
Results
miR-885-5p is significantly
upregulated in human OS tissues and cell lines
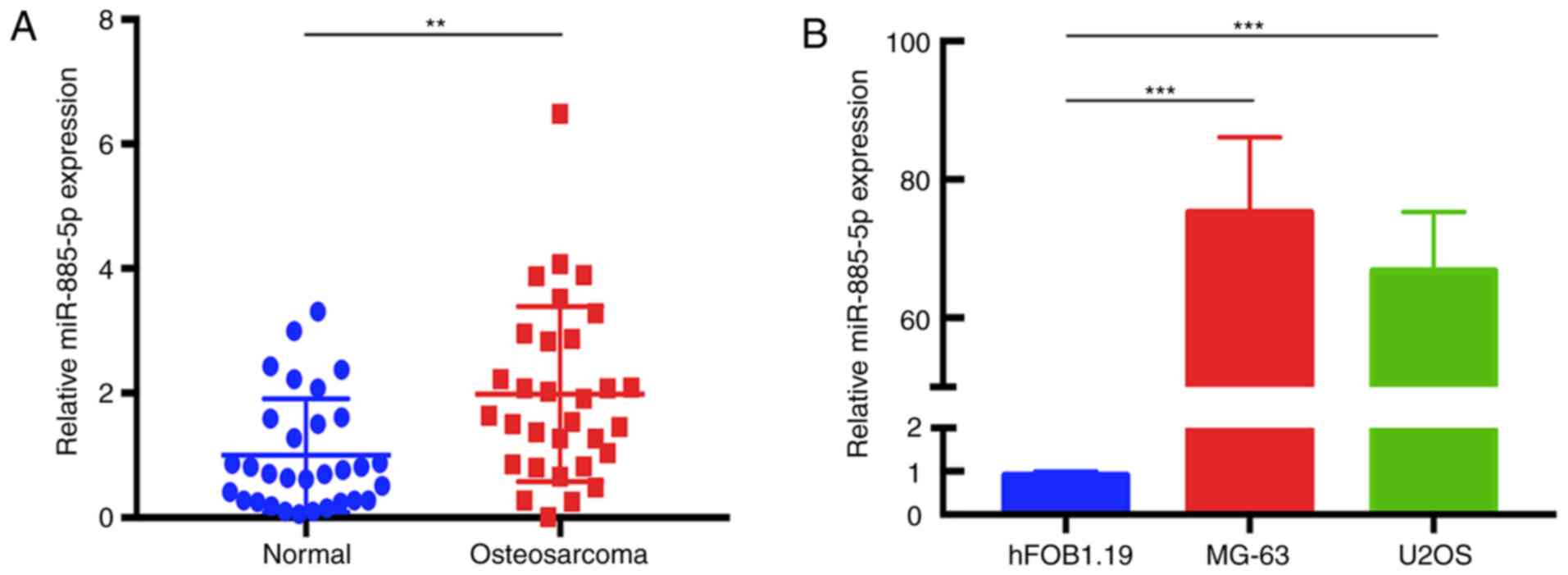
To determine the expression levels of miR-885-5p in
human OS tissues, RT-qPCR was performed in 31 pairs of OS tissues
compared with adjacent normal tissues, and in the human OS MG-63
and U2OS cell lines compared with the normal osteoblast hFOB1.19
cell line. The data revealed that miR-885-5p was significantly
upregulated in human OS tissues compared with that in adjacent
normal tissues (Fig. 1A). In
addition, similar results were observed between MG-63 and hFOB1.19,
and between U2OS and hFOB1.19 (Fig.
1B).
Upregulation of miR-885-5p is
associated with advanced clinicopathological features of OS
To evaluate the clinical relevance of miR-885-5p in
OS samples, the high and low expression of miR-885-5p was defined
based on the median expression levels. Measuring tumor size on
computed tomography, 5 cm was defined as the cut-off line (18). OS stage was determined by Enneking
staging system (19). A fusion of
stage IIB and stage III was to indicate the higher malignance than
stage IIA (20). As shown in Table I, high miR-885-5p expression was
significantly associated with tumor size (P=0.017) and clinical
stage (P=0.003). No significant difference was found between the
expression of miR-885-5p and patient age (P=0.458), gender
(P=0.677), anatomic location (P=0.174) and metastasis (P=0.999).
The Enneking stage of each sample was judged by two experienced
surgeons and further confirmed by pathological results.
 | Table I.Association of miR-885-5p expression
with the clinicopathological features of osteosarcoma cases
(n=31). |
Table I.
Association of miR-885-5p expression
with the clinicopathological features of osteosarcoma cases
(n=31).
|
|
| miR-885-5p
expression |
|
|---|
|
|
|
|
|
|---|
| Characteristics | Number of cases | High, n | Low, n | P-value |
|---|
| Age, years |
|
|
| 0.458 |
|
<18 | 15 | 9 | 6 |
|
|
≥18 | 16 | 12 | 4 |
|
| Gender |
|
|
| 0.677 |
|
Male | 21 | 14 | 7 |
|
|
Female | 10 | 8 | 2 |
|
| Tumor size, cm |
|
|
| 0.017a |
|
<5 | 5 | 2 | 4 |
|
| ≥5 | 26 | 21 | 5 |
|
| Anatomical
location |
|
|
| 0.174 |
|
Femur | 19 | 11 | 8 |
|
|
Tibia | 7 | 6 | 1 |
|
|
Humerus | 5 | 4 | 0 |
|
| Clinical stage
(Enneking) |
|
|
| 0.003a |
|
IIA | 8 | 2 | 6 |
|
|
IIB/III | 23 | 20 | 3 |
|
| Metastasis |
|
|
| 0.999 |
|
Yes | 11 | 8 | 3 |
|
| No | 20 | 14 | 6 |
|
Transfection efficiency of the
generated stable cell lines
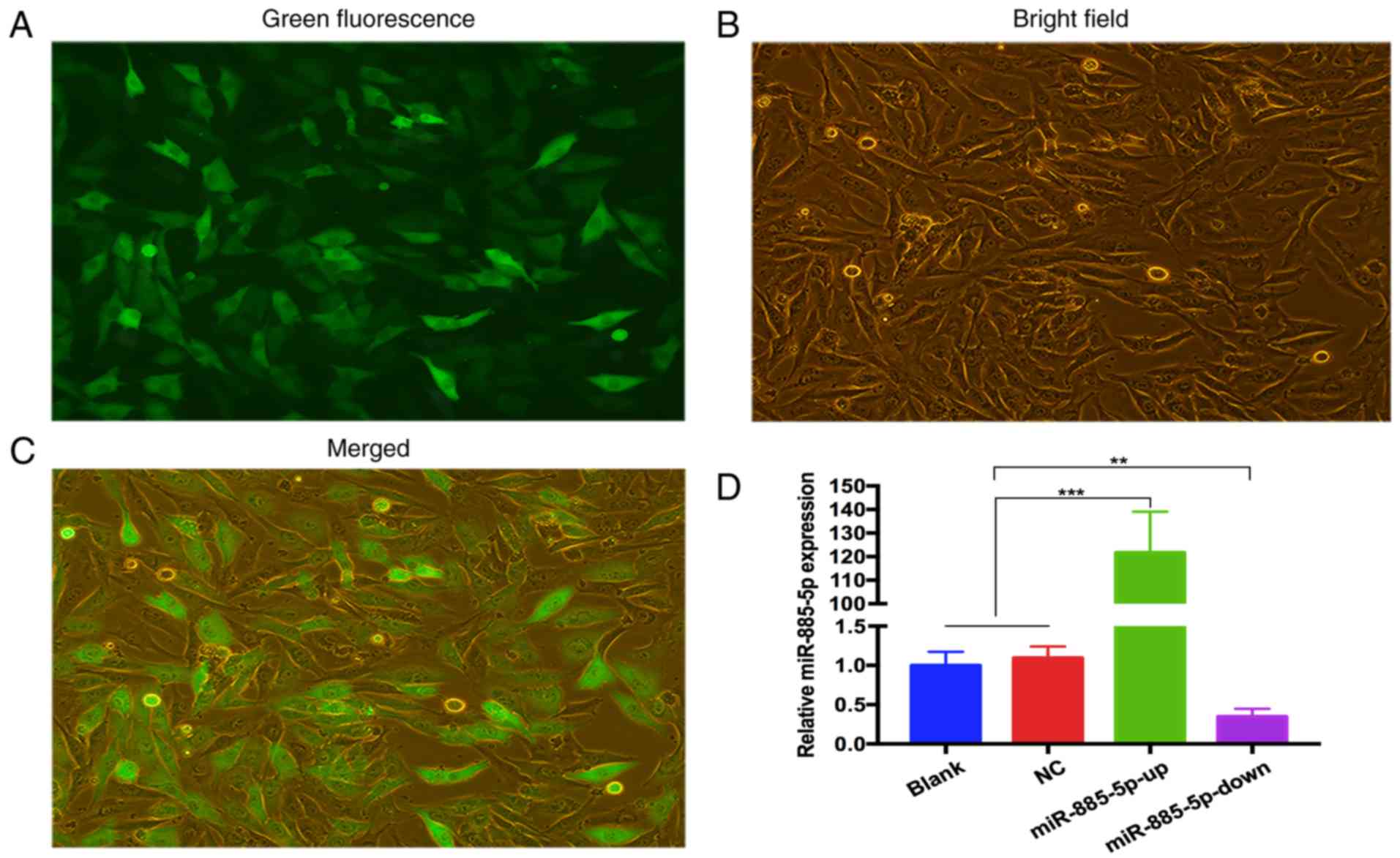
Following transfection and selection of cells, the
expression of GFP-labeled oligonucleotides was detected using
fluorescence microscopy (Fig. 2A-C).
RT-qPCR was used to validate the transfection efficiency. The
results revealed that miR-885-5p expression levels were
significantly higher in the miR-885-5p-up group and lower in the
miR-885-5p-down group compared with the blank and NC groups. There
was no difference between the blank group and the NC groups. The
data indicated that lentivirus containing miR-885-5p-up/down was
successfully transfected in MG-63 cells (Fig. 2D).
miR-885-5p promotes proliferation of
OS cells
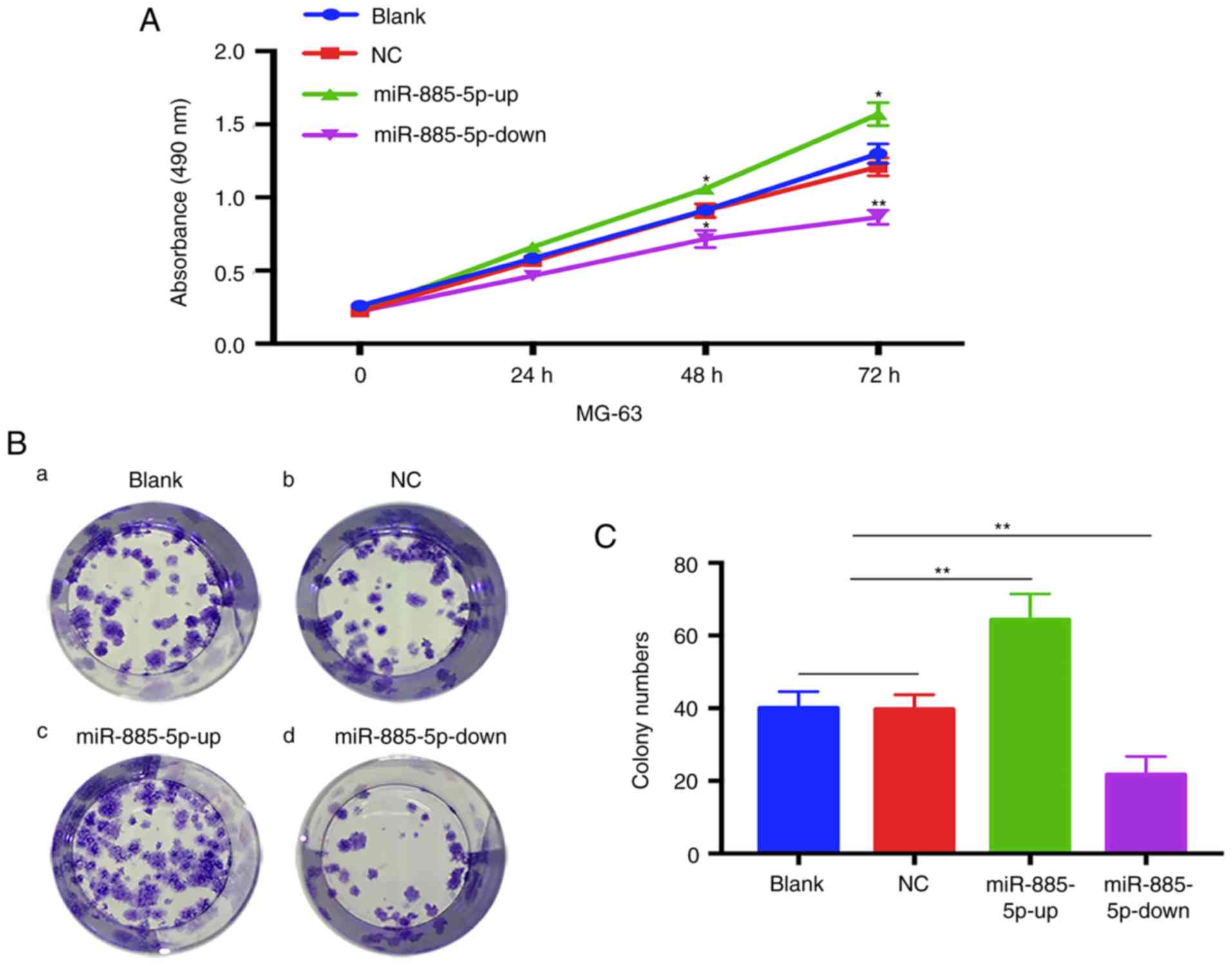
The MTT assay showed that MG-63 cells in the
miR-885-5p-up group exhibited a significant increase in
proliferation compared with the other three groups, while the cells
in the miR-885-5p-down group exhibited a decrease in proliferation.
No statistical differences were found between the blank and NC
groups (Fig. 3A). The results of the
colony formation assay also presented a similar trend in which the
colony formation was significantly decreased in the miR-885-5p-down
group of MG-63 cells compared with the blank and NC groups
(Fig. 3B and C). By contrast, cells
transfected with miR-885-5p-up exhibited significantly increased
colony formation compared with the other groups.
miR-885-5p promotes migration of OS
cells
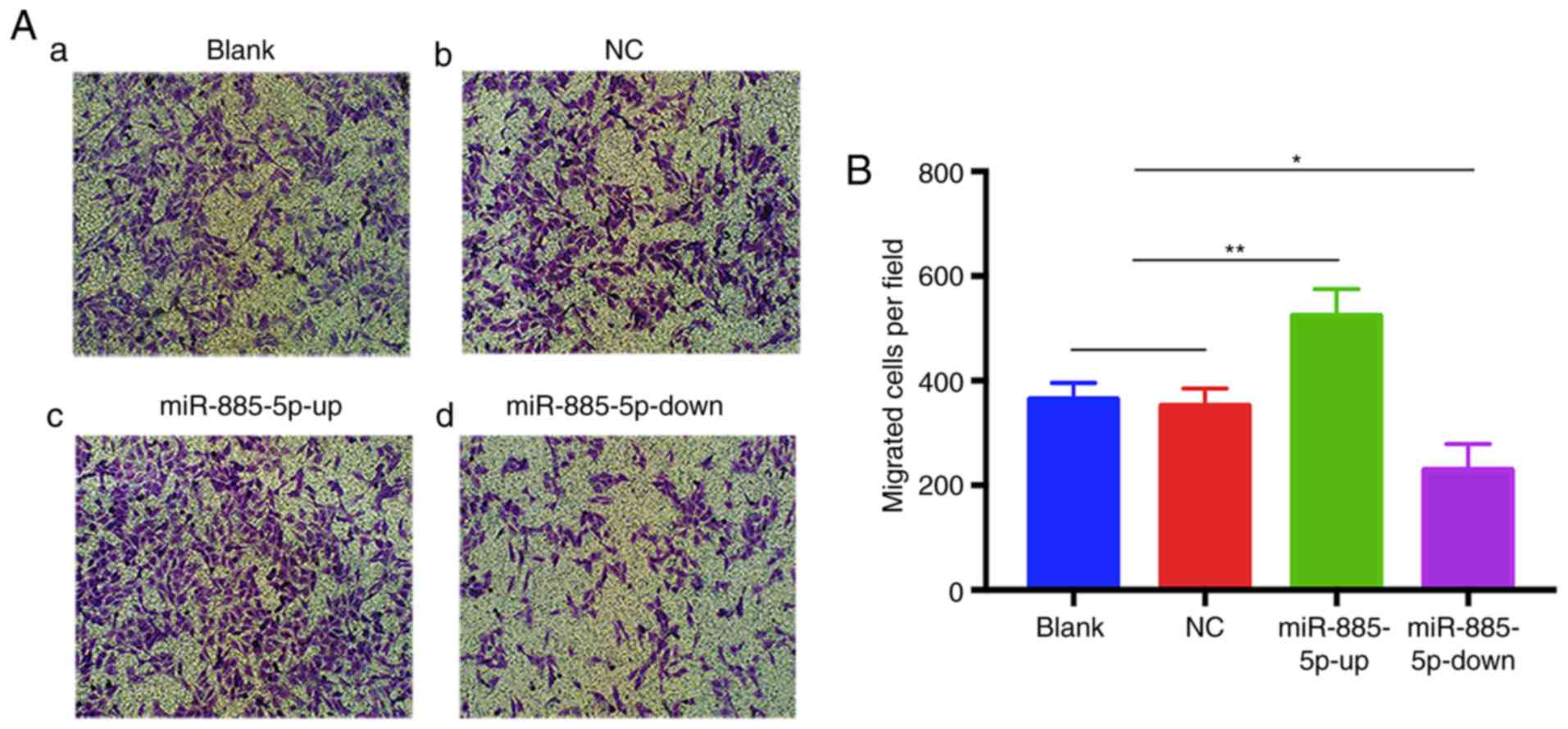
To further corroborate the effect of miR-885-5p on
MG-63 cell migration, a Transwell migration assay was performed.
The results revealed that the numbers of migrated cells in the
blank and NC groups were not significantly different, while the
miR-885-5p-up group exhibited significantly increased migration
compared with the other groups. Conversely, the number of migrated
cells transfected with miR-885-5p-down was significantly suppressed
(Fig. 4A and B). These results
indicated that miR-885-5p promotes the migration of OS cells in
vitro.
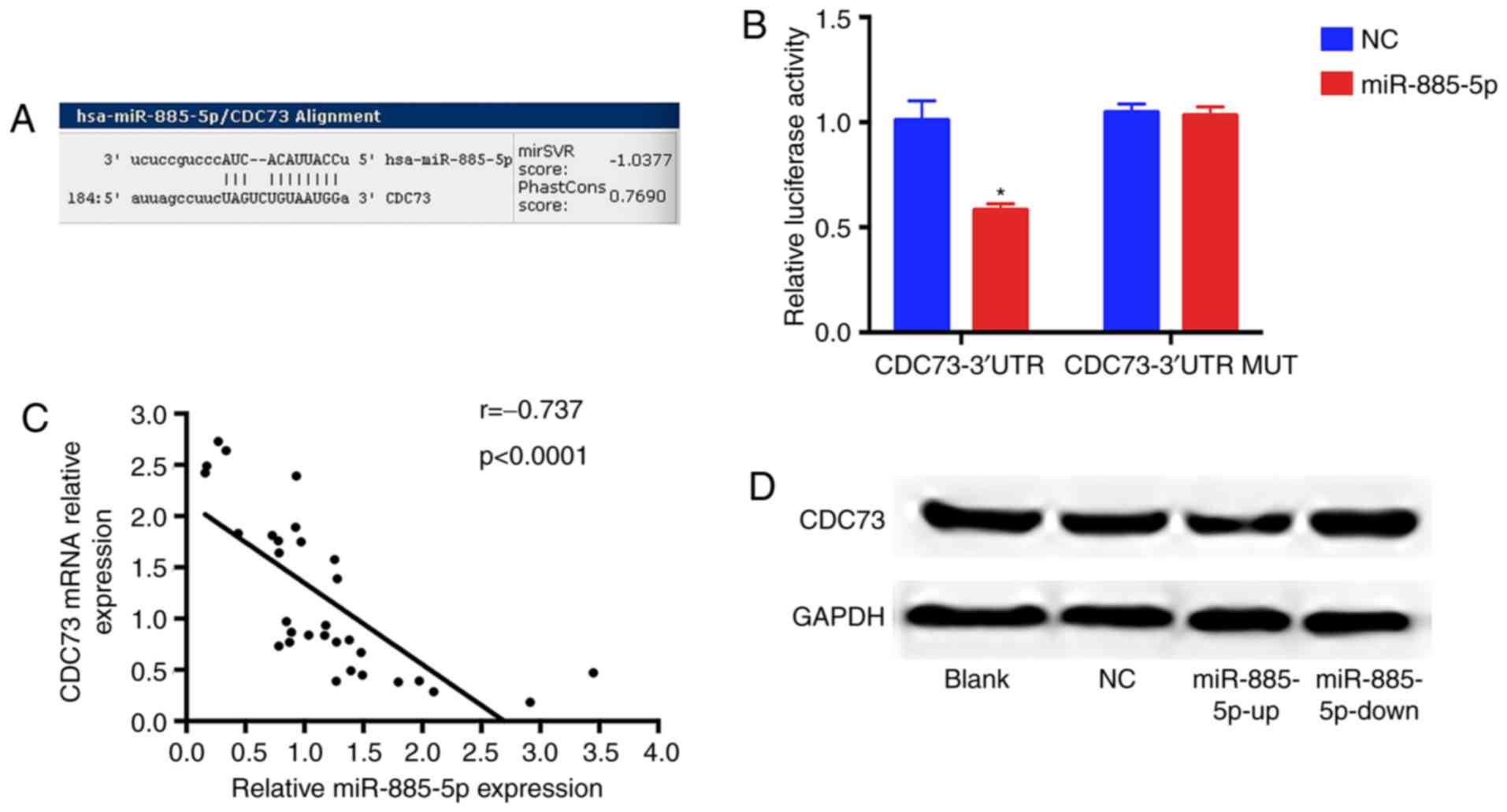
miR-885-5p directly targets and
inhibits CDC73 protein expression
To elucidate the underlying miR-885-5p-mediated
mechanisms, TargetScan (targetscan.org), miRanda (microrna.org)
and miRBaseTargets (mirbase.org)
were used to find potential target genes of miR-885-5p. Based on
bioinformatics algorithms, a highly-conserved miR-885-5p targeting
sequence was observed in the CDC73 3′-UTR, which suggested that
CDC73 could be a candidate target of miR-885-5p. Dual luciferase
reporter assay results showed that luciferase activity was markedly
reduced in MG-63 cells when CDC73 3′-UTR wild-type construct was
co-transfected with miR-885-5p-up compared with the mutant group.
Furthermore, miR-885-5p and CDC73 expression levels were examined
in 31 pairs of OS tissues and the association between miR-885-5p
and CDC73 was investigated. As demonstrated in Fig. 5, mRNA and protein levels of CDC73 were
significantly decreased and increased in response to miR-885-5p-up
and miR-885-5p-down groups, respectively. These data support the
hypothesis that miR-885-5p directly targets the 3′-UTR of CDC73
mRNA, leading to mRNA degradation and inhibiting its
expression.
Discussion
OS is the most common primary solid tumor of the
bone. The malignancy is characterized by malignant mesenchymal
cells, which generate osteoid and/or immature bone (21). One previous study demonstrated that
the dysregulation of miRNAs contributes to proliferation and
migration of numerous types of human cancer (22). Moreover, the expression of several
miRNAs, including miR-9, miR-21, miR-93 and miR-603, was shown to
be increased in OS tissues and cells (23–26).
Several studies have confirmed that miR-885-5p is aberrantly
expressed in various types of cancer. Furthermore, miR-885-5p
functions as a negative regulator of osteogenic differentiation of
bone marrow-derived mesenchymal stem cells, leading to terminal
differentiation failure. OS development may also be associated with
defects in osteogenic differentiation (27,28). Thus,
for a better understanding of the underlying gene network
orchestrated by miRNAs involved in OS tumor initiation, development
and progression, further studies are required to find effective
biomarkers for the diagnosis and treatment of OS.
The present study first investigated the effect of
miR-885-5p on OS tumor growth and migration. Consistent with
previous studies that demonstrated a role for miR-885-5p in certain
cancer types, including colorectal cancer, renal cell carcinoma and
Hurthle cell carcinoma (11–13) the present study observed that the
expression of miR-885-5p was significantly higher in OS tissues
compared with that in corresponding adjacent normal tissues. In
addition, OS cells lines (MG-63 and U2OS) were compared with a
normal human osteoblastic cell line, hFOB1.19, and it was found
that the expression of miR-885-5p was significantly upregulated.
The upregulated miR-885-5p expression was strongly associated with
malignant clinicopathological features of OS patients, including
tumor size and clinical stage. It was observed that the
overexpression of miR-885-5p promoted the proliferation and
migration of MG-63 cells, while the downregulation of miR-885-5p
expression showed the opposite results.
CDC73, also known as hyperparathyroidism 2, encodes
a tumor suppressor gene that is involved in regulating
transcriptional and post-transcriptional control pathways.
Mutations in this gene are associated with hyperparathyroidism-jaw
tumor syndrome and parathyroid carcinomas (29,30). A
previous study reported that loss of heterozygosity of the
wild-type CDC73 allele can be observed in sporadic human renal
tumors, including, papillary, clear cell and chromophobe renal cell
carcinomas, Wilms' tumors and oncocytomas (31). Overexpression of CDC73 inhibits the
formation of colonies and the proliferation of cells, and induces
arrest of the cell cycle in the G1 phase, suggesting
that it has a key role in cell growth and proliferation (32). Moreover, it was reported that Bruton's
tyrosine kinase increased the abundance of CDC73 when stimulation
was not present and that CDC73 functioned as a repressor of
β-catenin-mediated transcription in human colorectal cancer cells
and B cells (33). These findings
indicate that CDC73 serves a critical role in the development and
progression of certain types of malignant tumors. However, the
potential function of CDC73 and associated miRNAs in OS remain to
be established. In the present study, bioinformatics analysis was
used to identify CDC73 as a putative target of miR-885-5p.
miR-885-5p negatively modulates CDC73 mRNA and protein expression
in MG-63 cells. In the luciferase activity assay, it was observed
that overexpression of miR-885-5p markedly reduced the luciferase
activity of wild-type CDC73 3′-UTR. These data demonstrated that
miR-885-5p directly targets CDC73 by interacting with its 3′-UTR.
Although this does not fully explain how miR-885-5p induces OS cell
proliferation and migration, the TargetScan program predicted
hundreds of potential targets of miR-885-5p. Indeed, a single miRNA
can target multiple mRNAs to regulate gene expression (34). It is possible that other potential
targets may be involved in the proliferation and migration of OS
cells. Therefore, further research should be conducted to
comprehensively understand the role of miR-885-5p in OS
tumorigenesis.
Taken together, the results of the present study
demonstrated that miR-885-5p is highly upregulated in OS tissues
and cell lines, and that its increased expression is strongly
associated with malignant clinicopathological features. Moreover,
we speculate that the overexpression of miR-885-5p acts as an
oncogene that promotes cell proliferation and migration. It was
also confirmed that miR-885-5p directly targets CDC73 and inversely
regulates its expression. Collectively, it may be concluded that
the ectopic expression of miR-885-5p may serve a critical role in
the progression of OS, and may act as a novel diagnostic biomarker
and potential therapeutic target for OS.
Acknowledgements
The authors would like to thank Professor Lurong
Zhang, Dr Ruiqing Chen, Dr Fei Huang, Dr Ruilong Lan, Dr Junying
Chen and Dr Jinrong Chen (Central Laboratory, The First Affiliated
Hospital of Fujian Medical University, Fuzhou, China) for providing
excellent technical assistance and useful discussions.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (no. 31571292) and the Science
Foundation for Youths of Province (no. 2016J05182).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FY and ZW conducted the experiment and drafted the
manuscript. XW and SW contributed to statistical analysis and
manuscript writing. XL and QH participated in cell experiments and
clinical data collection. JL conceived the present study and helped
revise the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of The First Affiliated Hospital of Fujian Medical
University (Fuzhou, China) and the protocols conformed to the
ethical guidelines of the Declaration of Helsinki. All participants
involved in this study provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kobayashi E, Hornicek FJ and Duan Z:
MicroRNA involvement in osteosarcoma. Sarcoma. 2012:3597392012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the surveillance, epidemiology, and end results program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marina N, Gebhardt M, Teot L and Gorlick
R: Biology and therapeutic advances for pediatric osteosarcoma.
Oncologist. 9:422–441. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zamore PD and Haley B: Ribo-gnome: The big
world of small RNAs. Science. 309:1519–1524. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shin C, Nam JW, Farh KK, Chiang HR,
Shkumatava A and Bartel DP: Expanding the microRNA targeting code:
Functional sites with centered pairing. Mol Cell. 38:789–802. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qin S, Ai F, Ji WF, Rao W, Zhang HC and
Yao WJ: miR-19a promotes cell growth and tumorigenesis through
targeting SOCS1 in gastric cancer. Asian Pac J Cancer Prev.
14:835–840. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jones KB, Salah Z, Del Mare S, Galasso M,
Gaudio E, Nuovo GJ, Lovat F, LeBlanc K, Palatini J, Randall RL, et
al: miRNA signatures associate with pathogenesis and progression of
osteosarcoma. Cancer Res. 72:1865–1877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lam CS, Ng L, Chow AK, Wan TM, Yau S,
Cheng NS, Wong SK, Man JH, Lo OS, Foo DC, et al: Identification of
microRNA 885-5p as a novel regulator of tumor metastasis by
targeting CPEB2 in colorectal cancer. Oncotarget. 8:26858–26870.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoshino H, Yonemori M, Miyamoto K,
Tatarano S, Kofuji S, Nohata N, Nakagawa M and Enokida H:
microRNA-210-3p depletion by CRISPR/Cas9 promoted tumorigenesis
through revival of TWIST1 in renal cell carcinoma. Oncotarget.
8:20881–20894. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Petric R, Gazic B, Goricar K, Dolzan V,
Dzodic R and Besic N: Expression of miRNA and occurrence of distant
metastases in patients with Hürthle cell carcinoma. Int J
Endocrinol. 2016:89452472016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Z, Yin J, Yang J, Shen W, Zhang C,
Mou W, Luo J, Yan H, Sun P, Luo Y, et al: miR-885-5p suppresses
hepatocellular carcinoma metastasis and inhibits Wnt/β-catenin
signaling pathway. Oncotarget. 7:75038–75051. 2016.PubMed/NCBI
|
|
15
|
Allen-Rhoades W, Kurenbekova L,
Satterfield L, Parikh N, Fuja D, Shuck RL, Rainusso N, Trucco M,
Barkauskas DA, Jo E, et al: Cross-species identification of a
plasma microRNA signature for detection, therapeutic monitoring,
and prognosis in osteosarcoma. Cancer Med. 4:977–988. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao G, Cai C, Yang T, Qiu X, Liao B, Li
W, Ji Z, Zhao J, Zhao H, Guo M, et al: MicroRNA-221 induces cell
survival and cisplatin resistance through PI3K/Akt pathway in human
osteosarcoma. PLoS One. 8:e539062013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi K, Lan RL, Tao X, Wu CY, Hong HF and
Lin JH: Vitronectin significantly influences prognosis in
osteosarcoma. Int J Clin Exp Pathol. 8:11364–11371. 2015.PubMed/NCBI
|
|
19
|
Enneking WF, Spanier SS and Goodman MA: A
system for the surgical staging of musculoskeletal sarcoma. Clin
Orthop Relat Res. 106–120. 1980.PubMed/NCBI
|
|
20
|
Han K, Zhao T, Chen X, Bian N, Yang T, Ma
Q, Cai C, Fan Q, Zhou Y and Ma B: microRNA-194 suppresses
osteosarcoma cell proliferation and metastasis in vitro and in vivo
by targeting CDH2 and IGF1R. Int J Oncol. 45:1437–1449. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Arndt CA, Rose PS, Folpe AL and Laack NN:
Common musculoskeletal tumors of childhood and adolescence. Mayo
Clin Proc. 87:475–487. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nelson KM and Weiss GJ: MicroRNAs and
cancer: Past, present, and potential future. Mol Cancer Ther.
7:3655–3660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu SW, Li JP, Ma XL, Ma JX, Yang Y, Chen
Y and Liu W: miR-9 modulates osteosarcoma cell growth by targeting
the gcip tumor suppressor. Asian Pac J Cancer Prev. 16:4509–4513.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ziyan W, Shuhua Y, Xiufang W and Xiaoyun
L: MicroRNA-21 is involved in osteosarcoma cell invasion and
migration. Med Oncol. 28:1469–1474. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kawano M, Tanaka K, Itonaga I, Ikeda S,
Iwasaki T and Tsumura H: microRNA-93 promotes cell proliferation
via targeting of PTEN in Osteosarcoma cells. J Exp Clin Cancer Res.
34:762015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma C, Zhan C, Yuan H, Cui Y and Zhang Z:
MicroRNA-603 functions as an oncogene by suppressing BRCC2 protein
translation in osteosarcoma. Oncol Rep. 35:3257–3264. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu JF, Yang GH, Pan XH, Zhang SJ, Zhao C,
Qiu BS, Gu HF, Hong JF, Cao L, Chen Y, et al: Altered microRNA
expression profile in exosomes during osteogenic differentiation of
human bone marrow-derived mesenchymal stem cells. PLoS One.
9:e1146272014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wagner ER, Luther G, Zhu G, Luo Q, Shi Q,
Kim SH, Gao JL, Huang E, Gao Y, Yang K, et al: Defective osteogenic
differentiation in the development of osteosarcoma. Sarcoma.
2011:3252382011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Carpten JD, Robbins CM, Villablanca A,
Forsberg L, Presciuttini S, Bailey-Wilson J, Simonds WF, Gillanders
EM, Kennedy AM, Chen JD, et al: HRPT2, encoding parafibromin, is
mutated in hyperparathyroidism-jaw tumor syndrome. Nat Genet.
32:676–680. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shattuck TM, Välimäki S, Obara T, Gaz RD,
Clark OH, Shoback D, Wierman ME, Tojo K, Robbins CM, Carpten JD, et
al: Somatic and germ-line mutations of the HRPT2 gene in sporadic
parathyroid carcinoma. N Engl J Med. 349:1722–1729. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao J, Yart A, Frigerio S, Perren A,
Schraml P, Weisstanner C, Stallmach T, Krek W and Moch H: Sporadic
human renal tumors display frequent allelic imbalances and novel
mutations of the HRPT2 gene. Oncogene. 26:3440–3449. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang C, Kong D, Tan MH, Pappas DL Jr,
Wang PF, Chen J, Farber L, Zhang N, Koo HM, Weinreich M, et al:
Parafibromin inhibits cancer cell growth and causes G1 phase
arrest. Biochem Biophys Res Commun. 350:17–24. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
James RG, Biechele TL, Conrad WH, Camp ND,
Fass DM, Major MB, Sommer K, Yi X, Roberts BS, Cleary MA, et al:
Bruton's tyrosine kinase revealed as a negative regulator of
Wnt-beta-catenin signaling. Sci Signal. 2:ra252009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Selbach M, Schwanhäusser B, Thierfelder N,
Fang Z, Khanin R and Rajewsky N: Widespread changes in protein
synthesis induced by microRNAs. Nature. 455:58–63. 2008. View Article : Google Scholar : PubMed/NCBI
|