Introduction
Colorectal cancer (CRC) is responsible for
~7×105 mortalities annually, which makes it the fourth
leading cause of cancer-associated mortality for men and women
worldwide (1). Its incidence has
increased significantly in the majority of developing countries
(2). Sporadic CRC, which usually
presents as an isolated colonic or rectal lesion, is the most
common type of CRC that occurs in older individuals without any
family history of the disease. Genetic and epigenetic changes,
dietary patterns (high fat consumption) and environmental factors
are common risk factors for CRC (3,4). A
number of genetic factors have been identified as predictors for
the prognosis of CRC; however, the precise mechanism of CRC remains
unknown (5–7).
MicroRNAs (miRNAs/miRs), RNAs with a length of ~22
nucleotides, bind to the ‘seed region’ of between 2 and 8
nucleotides at the 3′ untranslated region (UTR) to regulate
targeted gene expression. The perfectly complementary base pairing
between the miRNA and its target mRNA sequence may induce RNA
silencing which results in decreased protein expression levels
(8–11). A number of single nucleotide
polymorphisms (SNPs) in the 3′ UTR of targeted genes had been
identified for their association with an individual's risk of
cancer by regulating targeted gene expression (12,13). As
a histone H4 Lys20 monomethyltransferase implicated in
normal cell cycle progression, SET8 (also known as PR-Set7 or
KMT5a) is regulated by miR-502 though the binding site in the 3′
UTR of SET8 mRNA (14–16).
Inappropriate SET8 expression induces S-phase defects and increased
DNA damage; SET8 also interacts directly with the DNA replication
factor proliferating-cell nuclear antigen and exhibits specific
effects at origins of replication (17–20).
During DNA double-strand break responses, SET8 activation has been
identified to be essential for p53-binding protein 1 (p53BP1)
recruitment (21). It has been
identified that SET8 could increase the metastatic capacity of
breast cancer cells by promoting epithelial-mesenchymal transition
and conferring TWIST dual transcriptional activities (22).
The SNP rs16917496 was identified previously to be
associated with risk of epithelial ovarian cancer and outcome of
hepatocellular carcinoma, small cell lung cancer and non-Hodgkin's
lymphomas (23–26). In the present study, this SNP was
genotyped in patients with CRC to assess its association with
cancer risk and outcome.
Materials and methods
Blood collection and DNA
extraction
Genomic DNA was extracted from blood samples (0.2
ml) of 109 patients with CRC who underwent CRC resection at The
Fourth Hospital of Hebei University (Shijiazhuang, China) between
March 2006 and December 2008 using a Wizard Genomic DNA extraction
kit (Promega Corporation, Madison, WI, USA). Blood samples were
also collected from 142 age and gender matched healthy controls at
the same hospital between April and December 2008. All procedures
were supervised and approved by the Hospital's Human Tissue
Research Committee. Written informed consent was obtained from all
patients enrolled in the present study.
Polymerase chain reaction (PCR)
amplification and sequence analysis
The DNA fragments flanking rs16917496 in the
SET8 3′ UTR were amplified using forward primer
5′-TCACGACGGTGCTACCTAAG-3′ and reverse primer
5′-CATGCTGGTGTGACACAGTC-3′ designed according to the National
Center for Biotechnology Information database (www.ncbi.nlm.nih.gov/snp) using a PCR Master mix
kit (Promega Corporation). The cycling conditions were one cycle of
denaturation at 95°C for 3 min, followed by 35 cycles of
denaturation at 95°C for 30 sec, annealing at 55°C for 30 sec,
extension at 72°C for 30 sec and fluorescence acquisition at 72°C
for 3 min. Cycle sequencing was performed using a Dye Terminator
Cycle Sequencing Ready Reaction kit (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and analyzed using an ABI Prism Genetic
Analyzer 3100 instrument (Thermo Fisher Scientific, Inc.).
Polymorphisms were confirmed by repeating the analysis on the two
DNA strands.
Determination of SET8 expression
levels in CRC tissue
CRC tissues collected from the same 109 patients,
from which blood samples were obtained, were fixed in formalin
(10%) for 24 h at room temperature immediately following resection,
dehydrated in absolute ethanol, embedded in paraffin and serial
sections (4-µm thick). CRC tissue was immunostained using an
anti-SET8 antibody (catalog no. ab3798; Abcam, Cambridge, UK) at a
dilution of 1:100 at 4°C overnight, followed by incubation with a
biotinylated secondary anti-mouse immunoglobulin G antibody
(pre-diluted; catalog no. PV600; Zhongshan, Inc., Guangzhou, China)
at room temperature for 1 h. Following incubation at room
temperature for 5 min with horseradish peroxidase-conjugated
streptavidin, the staining of CRC tissue was developed with
3,3-diaminobenzidine.
The stained slides were semi-quantified by two
pathologists who were blinded to the sequencing data using HScore
(25). Briefly, the percentage of
positively stained CRC cells in each of five samples was graded (0,
1+, 2+, 3+ and 4+). The HScore was calculated as follows:
HScore=(i+1)x, where i=1, 2, 3 and 4, and
x varied between 0 and 100%. High expression is defined as a
score of >100 and low expression is defined as a score of
<100.
Statistical analysis
The distribution of expression grades for each
SET8 genotype was compared using a χ2 test.
Survival curves were created using the Kaplan-Meier method with a
log-rank test and multivariate survival analysis was performed
using a Cox proportional hazards model. Statistical analyses were
performed using the SPSS software package (version 18.0; SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
SET8 genotype is associated with CRC
survival
A total of 109 patients with CRC were enrolled in
the present study. The post-operative survival of these patients
according to their clinical characteristics was analyzed using the
Kaplan-Meier method and a log-rank test. The clinical stage and
tumor length were investigated for their association with survival
times of patients with CRC using univariate analysis (Table I).
 | Table I.Univariate analysis of clinical
characteristics associated with post-operative survival in patients
with colorectal cancer. |
Table I.
Univariate analysis of clinical
characteristics associated with post-operative survival in patients
with colorectal cancer.
| Characteristic | n | 5-year survival rate,
% | P-value |
|---|
| Sex |
|
| 0.649 |
| Male | 61 | 55.7 |
|
|
Female | 48 | 58.3 |
|
| Age, years |
|
| 0.664 |
| ≤60 | 53 | 58.5 |
|
|
>60 | 56 | 55.4 |
|
| Tumor length, cm |
|
| 0.034 |
| ≤6 | 69 | 63.8 |
|
|
>6 | 40 | 45.0 |
|
| Tumor location |
|
| 0.161 |
| Right
colon | 57 | 50.9 |
|
|
Transverse colon | 7 | 100.0 |
|
| Left
colon | 43 | 58.1 |
|
|
Rectum | 2 | 50.0 |
|
| Clinical stage |
|
| 0.001 |
|
I+II | 57 | 71.9 |
|
|
III+IV | 52 | 40.4 |
|
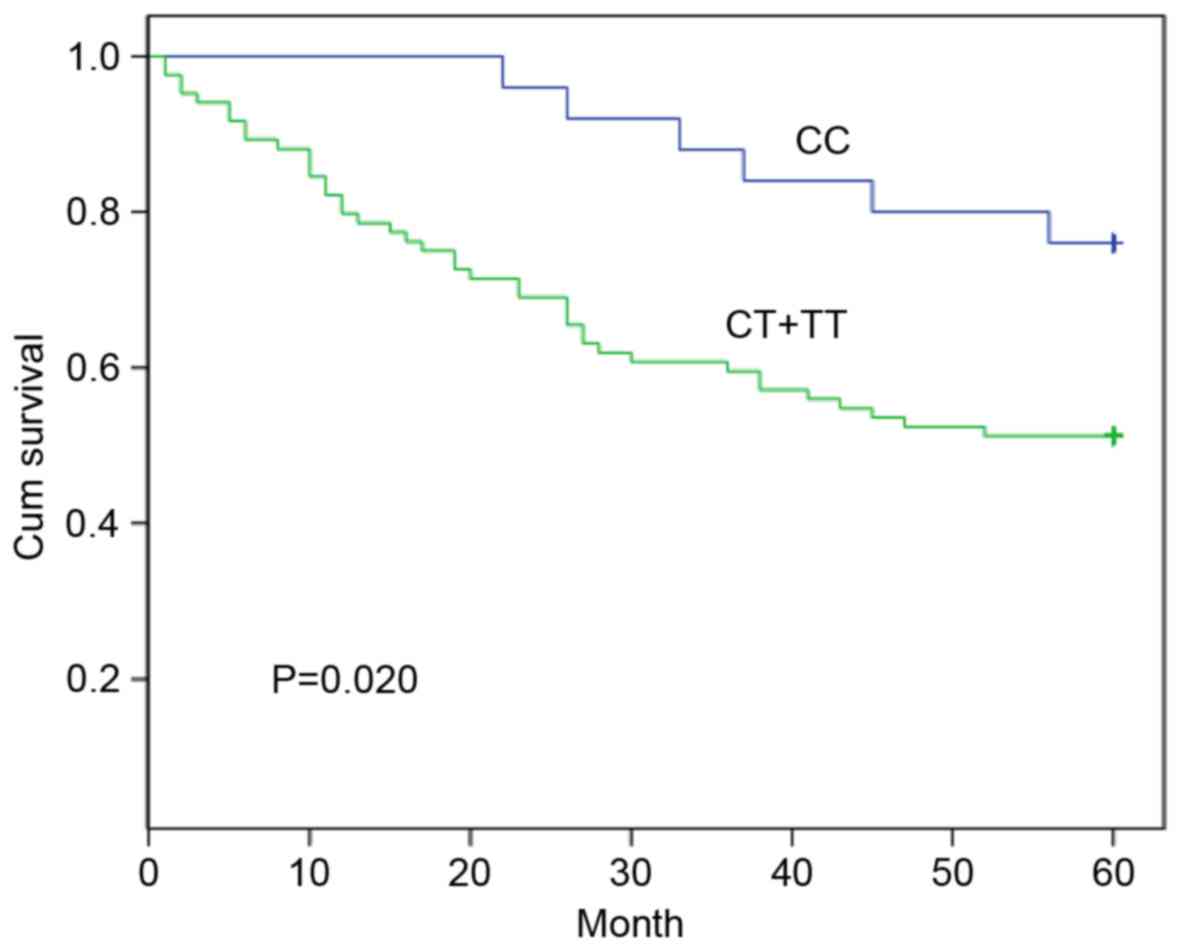
The SNP rs16917496 of SET8 was genotyped (CC,
CT and TT) in 109 patients with CRC and 142 controls; the
rs16917496 distribution followed a Hardy-Weinberg equilibrium, and
no difference in distribution frequency of the SET8 genotype
was identified between patients with CRC and controls (data not
shown). The association of SET8 genotype and post-operative
survival of patients with CRC was assessed using the Kaplan-Meier
method; the 5-year survival rate was 76.0% for patients with the CC
genotype and 51.2% for patients with the CT + TT genotypes
(Fig. 1). The patients carrying the
CC genotype was associated with a significantly longer survival
time compared with that of patients with the CT + TT genotypes
(P=0.020). The multivariate analysis with Cox proportional hazards
model was performed including all CRC survival-associated
predictors (Table II). The
SET8 genotype was identified as an independent predictor for
the prognosis of patients with CRC (relative risk, 2.406; 95%
confidence interval, 1.017–5.691; P=0.046).
| Table II.Multivariate analysis of prognostic
factors associated with post-operative survival in patients with
colorectal cancer with Cox hazard model. |
Table II.
Multivariate analysis of prognostic
factors associated with post-operative survival in patients with
colorectal cancer with Cox hazard model.
| Factor | RR | 95% CI | P-value |
|---|
| rs16917496 | 2.406 | 1.017–5.691 | 0.046 |
| Tumor length | 1.425 | 0.788–2.576 | 0.241 |
| Clinical stage | 2.406 | 1.289–4.491 | 0.006 |
SET8 expression mediated by miR-502 is
associated with CRC outcome
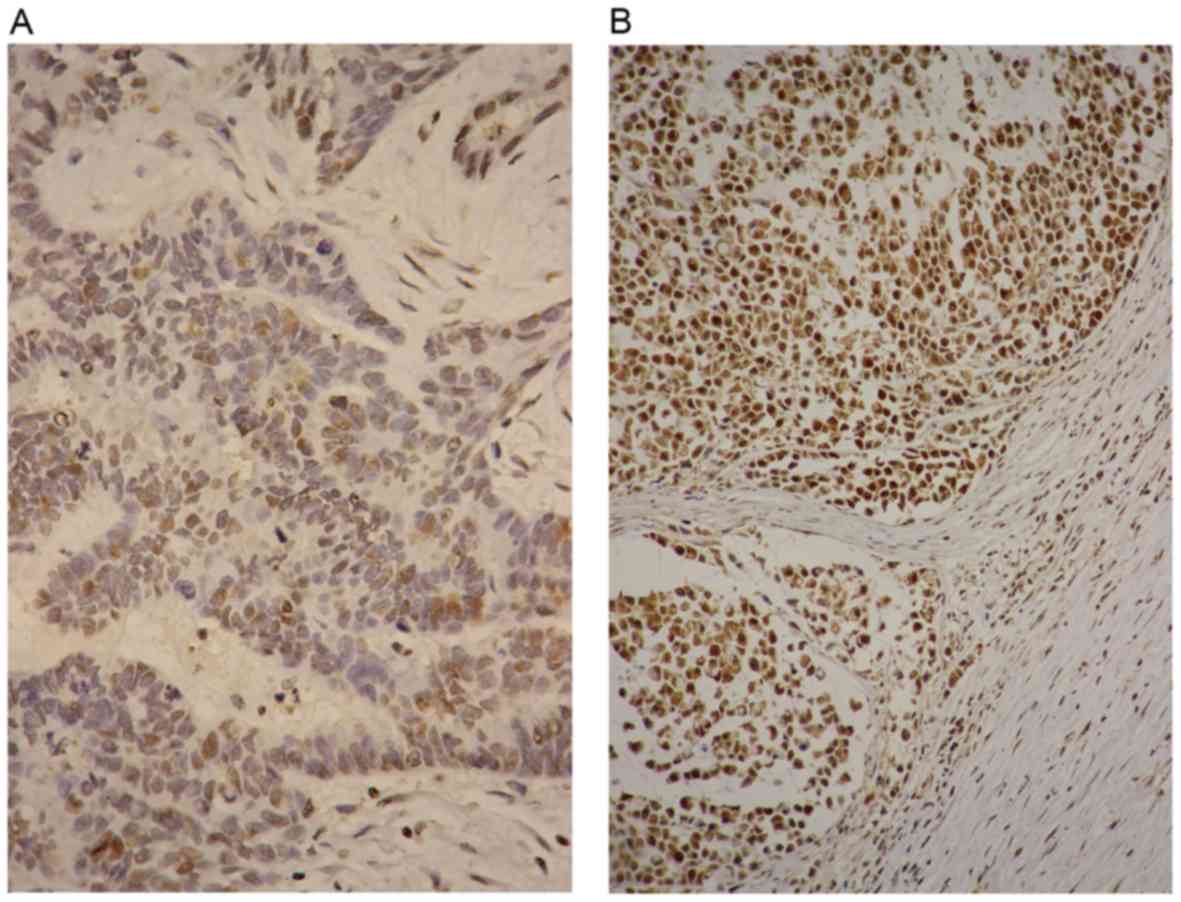
The association between the miR-502-binding site SNP
rs16917496 and SET8 expression was investigated in CRC tissues
(Fig. 2). The HScore of SET8 was
calculated and the SNP rs16917496-based SET8 expression in patients
with CRC is presented in Table
III. The SET8 CC genotype was associated with lower
levels of SET8 expression compared with that of the TT genotype by
χ2 test (P<0.001). Survival analysis referring to
SET8 expression was performed, the patients with low SET8
expression had a longer lifespan compared with that of patients
with high SET8 expression (5-year survival rate, 75.7 vs. 47.2%;
P=0.005).
| Table III.Association of SET8 expression with
SET8 genotype. |
Table III.
Association of SET8 expression with
SET8 genotype.
| Genotype | Low expression | High
expression | P-value |
|---|
| CC | 17 | 8 | <0.001 |
| CT+TT | 20 | 64 |
|
Discussion
The methyltransferase SET8 has been implicated in a
number of cancer processes (23–28). The
results of the present study indicated that the SNP rs16917496 in
the miR-502-binding site of the SET8 3′ UTR was associated
with the survival time of patients with CRC. In addition, this SNP
may eliminate the binding affinity between miR-502 and SET8
so as to modulate SET8 expression, with altered SET8 expression
contributing to the progression of CRC.
The association of the miRNA-binding site SNP with
cancer risk and outcome has been well-studied (25,29–32).
Consistent with previous studies that identified the prognostic
value of the rs16917496 on the outcome of patients with small cell
lung cancer, hepatocellular carcinoma and non-Hodgkin's lymphomas
with the CC genotype being associated with longer survival
(23–26), the results of the present study
indicate that the CC genotype tends to lead to a long lifespan in
patients with CRC. The convergence of the SET8 CC genotype
and low SET8 expression which was identified in patients with
breast cancer and hepatocellular carcinoma was also confirmed in
CRC tissue (25,33).
SET8 methylates Lys382 of p53 to modulate
p53 activity, which is implicated in cell death and cell cycle
arrest following DNA damage; furthermore, SET8 depletion
could abrogate the accumulation of 53BP1 in DNA double-strand
breaks to render cells sensitive to apoptosis (21,27).
SET8 could methylate the promoters of the TWIST target genes
of breast cancer cell to promote epithelial-mesenchymal transition
and enhance the invasive potential (22). Although SET8 would be a novel
therapy target for CRC treatment, the underlying molecular
mechanism of miR-502-mediated SET8 expression and whether SET8
modifies the development of CRC via its methyltransferase requires
further investigation.
Acknowledgements
The authors would like to thank Professor Xiaoling
Wang (Department of Pathology, The Fourth Hospital of Hebei Medical
University) for their assistance with immuohistochemistry.
Funding
The present study was supported by Key Basic
Research Program of Hebei (grant no. 14967713D).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SL collected PCR and sequence data, and wrote the
manuscript. HD collected the immunostaining data. JW contributed to
data collection and writing of the manuscript. CW contributed to
project design, data collection and manuscript writing. All authors
read and approved the final manuscript. The authors agreed to be
accountable for all aspects of the study, ensuring that questions
related to the accuracy or integrity of any part of the study are
appropriately investigated and resolved.
Ethics approval and consent to
participate
All procedures were supervised and approved by the
Human Tissue Research Committee of The Fourth Hospital of Hebei
University (Shijiazhuang, China). Written informed consent was
obtained from all patients enrolled in the present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Ervik M,
Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D and
Bray F: Globocan 2012: Estimated cancer incidence, mortality and
prevalence worldwide in 2012 v1.0. IARC CancerBase. (11): IARC,
Lyon. 2014, http://globocan.iarcfr
|
|
2
|
Center MM, Jemal A and Ward E:
International trends in colorectal cancer incidence rates.
CancerEpidemiol Biomarkers Prev. 18:1688–1694. 2009. View Article : Google Scholar
|
|
3
|
World Cancer Research Fund, American
Institute for Cancer Research Imperial College, London, . WCRF/AICR
Systematic Literature Review Continuous Update Project Report: The
Associations between Food, Nutrition and Physical Activity and the
Risk of Colorectal Cancer. https://www.wcrf.org/sites/default/files/SLR_colorectal_cancer_2010.pdfOctober.
2010
|
|
4
|
Bishehsari F, Mahdavinia M, Vacca M,
Malekzadeh R and Mariani-Costantini R: Epidemiological transition
of colorectal cancer in developing countries: Environmental
factors, molecular pathways, and opportunities for prevention.
World J Gastroenterol. 20:6055–6072. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Migliore L, Migheli F, Spisni R and
Coppedè F: Genetics, cytogenetics, and epigenetics of colorectal
cancer. J Biomed Biotechnol. 2011:7923622011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Peters U, Jiao S, Schumacher FR, Hutter
CM, Aragaki AK, Baron JA, Berndt SI, Bézieau S, Brenner H,
Butterbach K, et al: Colon cancer family registry and the genetics
and epidemiology of colorectal cancer consortium: Identification of
genetic susceptibility loci for colorectal tumors in a genome-wide
meta-analysis. Gastroenterology. 144:799–807. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Markowitz SD and Bertagnolli MM: Molecular
origins of cancer: Molecular basis of colorectal cancer. N Engl J
Med. 361:2449–2460. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zeng Y, Yi R and Cullen BR: MicroRNAs and
small interfering RNAs can inhibit mRNA expression by similar
mechanisms. Proc Natl Acad Sci USA. 100:9779–9784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeng Y, Wagner EJ and Cullen BR: Both
natural and designed micro RNAs can inhibit the expression of
cognate mRNAs when expressed in human cells. Mol Cell. 9:1327–1333.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chin LJ, Ratner E, Leng S, Zhai R, Nallur
S, Babar I, Muller RU, Straka E, Su L, Burki EA, et al: A SNP in a
let-7 microRNA complementary site in the KRAS 3′ untranslated
region increases non-small cell lung cancer risk. Cancer Res.
68:8535–8540. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brendle A, Lei H, Brandt A, Johansson R,
Enquist K, Henriksson R, Hemminki K, Lenner P and Försti A:
Polymorphisms in predicted microRNA-binding sites in integrin genes
and breast cancer: ITGB4 as prognostic marker. Carcinogenesis.
29:1394–1399. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fang J, Feng Q, Ketel CS, Wang H, Cao R,
Xia L, Erdjument-Bromage H, Tempst P, Simon JA and Zhang Y:
Purification and functional characterization of SET8, a nucleosomal
histone H4-lysine 20-specific methyltransferase. Curr Biol.
12:1086–1099. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nishioka K, Rice JC, Sarma K,
Erdjument-Bromage H, Werner J, Wang Y, Chuikov S, Valenzuela P,
Tempst P, Steward R, et al: PR-Set7 is a nucleosome-specific
methyltransferase that modifies lysine 20 of histone H4 and is
associated with silent chromatin. Mol Cell. 9:1201–1213. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu S, Wang W, Kong X, Congdon LM, Yokomori
K, Kirschner MW and Rice JC: Dynamic regulation of the PR-Set7
histone methyltransferase is required for normal cell cycle
progression. Genes Dev. 24:2531–2542. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tardat M, Murr R, Herceg Z, Sardet C and
Julien E: PR-Set7-dependent lysine methylation ensures genome
replication and stability through S phase. J Cell Biol.
179:1413–1426. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jorgensen S, Elvers I, Trelle MB, Menzel
T, Eskildsen M, Jensen ON, Helleday T, Helin K and Sorensen CS: The
histone methyltransferase SET8 is required for S-phase progression.
J Cell Biol. 179:1337–1345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Abbas T, Shibata E, Park J, Jha S, Karnani
N and Dutta A: CRL4(Cdt2) regulates cell proliferation and histone
gene expression by targeting PR-Set7/Set8 for degradation. Mol
Cell. 40:9–21. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Huen MS, Sy SM, van Deursen JM and Chen J:
Direct interaction between SET8 and proliferating cell nuclear
antigen couples H4-K20 methylation with DNA replication. J Biol
Chem. 283:11073–11077. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dulev S, Tkach J, Lin S and Batada NN:
SET8 methyltransferase activity during the DNA double-strand break
response is required for recruitment of 53BP1. EMBO Rep.
15:1163–1174. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang F, Sun L, Li Q, Han X, Lei L, Zhang H
and Shang Y: SET8 promotes epithelial-mesenchymal transition and
confers TWIST dual transcriptional activities. EMBO J. 31:110–123.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang C, Guo Z, Wu C, Li Y and Kang S: A
polymorphism at the miR-502 binding site in the 3′ untranslated
region of the SET8 gene is associated with the risk of epithelial
ovarian cancer. Cancer Genet. 205:373–376. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ding C, Li R, Peng J, Li S and Guo Z: A
polymorphism at the miR-502 binding site in the 3′ untranslated
region of the SET8 gene is associated with the outcome of
small-cell lung cancer. Exp Ther Med. 3:689–692. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo Z, Wu C, Wang X, Wang C, Zhang R and
Shan B: A polymorphism at the miR-502 binding site in the
3′-untranslated region of the histone methyltransferase SET8 is
associated with hepatocellular carcinoma outcome. Int J Cancer.
131:1318–1322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Diao L, Su H, Wei G, Li T, Gao Y, Zhao G
and Guo Z: Prognostic value of microRNA 502 binding site SNP in the
3′-untranslated region of the SET8 gene in patients with
non-Hodgkin's lymphoma. Tumori. 100:553–558. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shi X, Kachirskaia I, Yamaguchi H, West
LE, Wen H, Wang EW, Dutta S, Appella E and Gozani O: Modulation of
p53 function by SET8-mediated methylation at lysine 382. Mol Cell.
27:636–646. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dhami GK, Liu H, Galka M, Voss C, Wei R,
Muranko K, Kaneko T, Cregan SP, Li L and Li SS: Dynamic methylation
of Numb by Set8 regulates its binding to p53 and apoptosis. Mol
Cell. 50:565–576. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Landi D, Gemignani F, Naccarati A, Pardini
B, Vodicka P, Vodickova L, Novotny J, Forsti A, Hemminki K, Canzian
F and Landi S: Polymorphisms within micro-RNA-binding sites and
risk of sporadic colorectal cancer. Carcinogenesis. 29:579–584.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao Y, He Y, Ding J, Wu K, Hu B, Liu Y, Wu
Y, Guo B, Shen Y, Landi D, et al: An insertion/deletion
polymorphism at miRNA-122-binding site in the interleukin-1alpha 3′
untranslated region confers risk for hepatocellular carcinoma.
Carcinogenesis. 30:2064–2069. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Horikawa Y, Wood CG, Yang H, Zhao H, Ye Y,
Gu J, Lin J, Habuchi T and Wu X: Single nucleotide polymorphisms of
microRNA machinery genes modify the risk of renal cell carcinoma.
Clin Cancer Res. 14:7956–7962. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hu Z, Chen J, Tian T, Zhou X, Gu H, Xu L,
Zeng Y, Miao R, Jin G, Ma H, et al: Genetic variants of miRNA
sequences and non-small cell lung cancer survival. J Clin Invest.
118:2600–2608. 2008.PubMed/NCBI
|
|
33
|
Song F, Zheng H, Liu B, Wei S, Dai H,
Zhang L, Calin GA, Hao X, Wei Q, Zhang W and Chen K: An
miR-502-binding site single-nucleotide polymorphism in the
3′-untranslated region of the SET8 gene is associated with early
age of breast cancer onset. Clin Cancer Res. 19:6292–6300. 2009.
View Article : Google Scholar
|