Introduction
Hepatocellular carcinoma (HCC) is a common primary
liver malignancy, which represents the fifth and seventh most
common type of cancer worldwide in men and women, respectively
(1). The malignancy of HCC is
attributed to its fast progression, the rapid development of
metastasis and the absence of efficient curative therapy, all
leading to an unfavorable prognosis (2). Treatments available for HCC include
surgical resection, regional ablation and liver transplantation;
however, these options are not proposed to patients diagnosed at an
advanced stage. Treatment with near infrared-induced hyperthermia
exerts promising inhibitory effects on HCC, although the poor
penetration of the light restricts its application (3). In addition, numerous patients with HCC
present with tolerance to sorafenib, which is a common United
States Food and Drug Administration-approved chemotherapeutic drug
for patients at the advanced stage (4). Additionally, some novel tyrosine kinase
inhibitors, including lenvatinib, cabozantinib and regorafenib,
have demonstrated promising therapeutic effects in recent clinical
trials. However, the efficacy, side effects and toxicity associated
with these treatments still require further study (5–7). It is
therefore crucial to explore novel anti-HCC drugs and therapeutic
approaches.
Activation of epithelial-mesenchymal transition
(EMT) is characterized by the progression of an epithelial
phenotype to a mesenchymal phenotype. It is a crucial event for
tumor metastasis, particularly in the early stages of disease
(8). In addition, numerous
EMT-related genes are associated with metastasis and the recurrence
of cancer (9–11). In HCC, EMT is regulated by various
oncogenes and tumor suppressor genes, and stimulates venous
invasion and metastasis, leading to poor prognosis (12). Forkhead box O1 (FOXO1) belongs to the
forkhead family of transcription factors characterized by a
distinct forkhead domain. FOXO1 is considered a tumor suppressor
gene that is regulated by the phosphoinositide 3-kinase
(PI3K)/protein kinase B (AKT) and Janus kinase 1 (JAK1)/signal
transducer and activator of transcription 3 (STAT3) signaling
pathways, forming the AKT/STAT3-FOXO1 signaling pathway (13,14). Our
previous studies have revealed that FOXO1 inhibits the invasion and
metastasis of HCC by reversing zinc finger E-box-binding homeobox
2-induced EMT (15). Furthermore,
various overexpressed oncogenes in HCC, including Epidermal growth
factor receptor kinase substrate 8-like protein 3 and zinc finger
and BTB domain-containing 20, are able to promote proliferation and
inhibit apoptosis of HCC by repressing FOXO1, suggesting that FOXO1
may inhibit proliferation and stimulate apoptosis (16,17).
These findings suggested that the AKT/STAT3-FOXO1 signaling pathway
may participate in several processes involved in the progression of
HCC, including proliferation, apoptosis, and EMT-associated
migration and invasion.
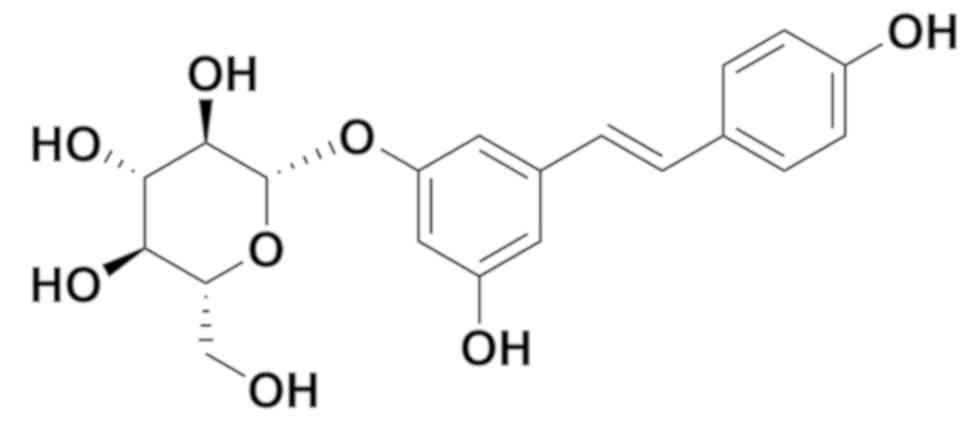
Polydatin, also named pieceid, (E)-piceid,
(E)-polydatin, trans-polydatin and
3,40,5-trihydroxystilbene-3-b-D-glucoside, is a monocrystalline
compound originally extracted from the root and rhizome of
Polygonum cuspidatum (Fig.
1). Polydatin is also detectable in grapes, peanuts, hop cones,
red wine, hop pellets, cocoa-containing products and chocolate.
Previous studies have demonstrated that polydatin has many
biological functions, including the prevention of platelet
aggregation (18), and
cardioprotective (19) and
anti-inflammatory (20) properties.
In addition, polydatin stimulates apoptosis and cell cycle arrest
in lung and colorectal cancers (21,22).
However, the effects of polydatin on HCC are currently unknown.
To the best of our knowledge, the present study
suggested for the first time that polydatin may inhibit EMT-related
migration and invasion, and proliferation (via G2/M
arrest) of HCC cells, and may promote HCC cell apoptosis by
limiting the AKT/STAT3-FOXO1 signaling pathway.
Materials and methods
Cells and reagents
The HCCLM3 human HCC cell line and LO2 normal
hepatic cell line were purchased from the American Type Culture
Collection (Manassas, VA, USA), and were cultured in Dulbecco's
modified Eagle's medium (DMEM) containing 10% fetal bovine serum,
100 IU/ml penicillin and 100 µg/ml streptomycin, which were all
obtained from Gibco, Thermo Fisher Scientific, Inc. (Waltham, MA,
USA). Cells were incubated at 37°C in a humidified incubator
containing 5% CO2. Polydatin (cat. no. P109977) was
purchased from Aladdin Industrial Corporation (Shanghai, China). A
stock solution of 350 mmol/l polydatin was prepared in dimethyl
sulfoxide (DMSO) and freshly diluted in medium prior to
experiments. MTT assay (cat. no. M2128) was purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany) and fluorescein
isothiocyanate (FITC)-conjugated Annexin V/propidium iodide (PI)
apoptosis detection kit was provided by 4A Beijing Biotech Co.,
Ltd. (Beijing, China). Primary and secondary antibodies were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
The Bio-Rad protein assay kit II was supplied by Bio-Rad
Laboratories, Inc. (Hercules, CA, USA) and the enhanced
chemiluminescent western blot detection reagents (cat. no. RPN2106)
were obtained from GE Healthcare (Chicago, IL, USA).
Cytotoxicity assay
The cytotoxicity of polydatin was measured using the
MTT assay. Cells were seeded in 96-well plates at 4×103
cells/well for 24 h. HCCLM3 and LO2 cells were treated with
increasing doses of polydatin (0–800 µmol/l) for various durations
(24–72 h). MTT solution (5 mg/ml in DMEM medium) was added (20
µl/well) and plates were further incubated for 4 h at 37°C. A
volume of 100 µl DMSO was added to each well to solubilize the
formazan product prior to measuring the absorbance with a
microplate reader at 490 nm. The assays were performed three
times.
Colony formation efficiency assay
HCCLM3 cells were seeded in 6-well plates at a
density of 1×103 cells/well for 24 h. Culture medium was
replaced with DMEM containing different doses of polydatin (0–800
µmol/l) and cells were incubated for 24 h at 37°C. The supernatant
was replaced with 2 ml regular DMEM containing 10% FBS, and cells
were cultured for 2 weeks at 37°C until visible cell clones were
formed. Once colonies were formed, cells were fixed with 4%
paraformaldehyde (PFA) for 25 min and washed three times with PBS
at room temperature. Cells were stained with crystal violet for 25
min and rinsed three times with PBS at room temperature. Colonies
were counted in a double blind manner using a light microscope
(Shanghai CSOIF Co., Ltd., Shanghai, China). Results were presented
as the percentage of colony numbers (average colony numbers of each
group compared with control), and the assays were replicated three
times.
Wound healing assay
HCCLM3 cells were seeded into 6-well plates at
2.5×105 cells/well and cultured at 37°C until a
monolayer was formed. Cells were scratched with a sterile
micropipette tip and treated with 20 µg/ml mitomycin (Aladdin
Industrial Corporation) for 20 min. Cells were washed with PBS to
remove debris, and further cultivated with serum-free medium
containing different doses of polydatin (0–150 µmol/l) for 24 and
48 h. The migration distance was measured and analyzed by Image J
v1.8.0 (National Institutes of Health, Bethesda, MD, USA), and the
assays were repeated three times.
Migration and invasion assays
Transwell chambers (pore size, 8 µm) were used to
detect the migration and invasion of HCCLM3 cells pretreated with
polydatin (0–150 µmol/l) for 48 h at 37°C. For the migration assay,
5×104 cells were seeded in serum-free DMEM (200 µl) into
the upper chambers and 500 µl DMEM containing 10% FBS was added to
the lower chamber. For the invasion assay, the upper polycarbonate
membranes of the chambers were coated with 5 mg/ml Matrigel.
Following a 48 h incubation, cells were fixed with 4% PFA for 20
min and stained with crystal violet for 20 min. Cells in three
randomly chosen fields were assessed under a light microscope
(Shanghai CSOIF Co., Ltd). Each assay was performed three
times.
Analysis of apoptosis
Annexin V-FITC/PI double staining was conducted to
examine the apoptosis of HCCLM3 cells following treatment with
polydatin. Briefly, cells were seeded in 6-well plates at a density
of 2.5×105 cells/well and were cultured overnight to
allow adhesion. Following treatment with increasing doses of
polydatin (0–800 µmol/l) for 48 h at 37°C, cells were stained with
Annexin V-FITC/PI for 15 min in the dark, according to the
manufacturer's protocol. Apoptotic cells were detected by flow
cytometry using the FACSCalibur flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA). The assays were carried out three
times.
Cell cycle analysis
HCCLM3 cells were seeded in 6-well plates
(2.5×105/well). At 24 h, cells were treated with 200
µmol/l polydatin for an additional 48 h at 37°C. Following
trypsinization, cells were washed with PBS and fixed with 75%
ethanol for 4 h at 4°C. Cells were washed twice with PBS and
covered with 0.5 ml PBS containing 20 µl RNaseA and PI for 30 min
at 4°C. The cell cycle progression of HCC cells was detected using
the FACSCalibur flow cytometer with an excitation wavelength of 488
nm, and emission wavelength of at 670 nm. Cell cycle progression
was analyzed using ModFit LT 4.1 (Verity Software House, Topsham,
ME, USA) by gating live cells.
Western blotting
HCCLM3 cells were seeded into 6-well plates at
2.5×105 cells/well and incubated at 37°C overnight. The
supernatant was replaced by fresh medium containing increasing
concentrations of polydatin (0–800 µmol/l) for 48 h at 37°C.
Following treatment, cells were lysed with RIPA buffer (Aladdin
Industrial Corporation) and subjected to protein extraction.
Protein concentrations were determined using the Pierce Micro BCA
protein assay system (Pierce; Thermo Fisher Scientific, Inc.).
Equal amounts of protein (25 µg) from each sample were separated
via SDS-PAGE on a 10% gel, and transferred to nitrocellulose
membranes that were further blocked with 5% non-fat milk (diluted
in TBST containing 0.1% Tween-20) for 1 h at room temperature.
Target proteins were immunoblotted with the appropriate primary and
horseradish peroxidase (HRP)-conjugated secondary antibodies, which
were all diluted at 1:1,000. The following antibodies were
purchased from Wuhan Sanyang Biotechnology (Wuhan, China): Cyclin
B1 (cat. no. 55004-1-AP) Cyclin D1 (cat. no. 60186-1-Ig) Bax (cat.
no. 60267-1-Ig) Bcl-2 (cat. no. 60178-1-Ig) β-actin (cat. no.
20536-1-Ig) Vimentin (cat. no. 60,330-1-Ig) N-Cadherin (cat. no.
66219-1-Ig) E-Cadherin (cat. no. 60335-1-Ig), p-AKT (cat. no.
66444-1-Ig), AKT (cat. no. 60203-2-Ig) and FOXO1 (cat. no.
66457-1-Ig). The following antibodies were obtained from Abcam
(Shanghai, China): p-JAK1 (cat. no. ab138005), JAK1 (cat. no.
ab47435), p-STAT3 (cat. no. ab76315), STAT3 (cat. no. ab119352),
p-p38 (cat. no. ab4822) and p38 (cat. no. ab31828). Primary
antibodies were incubated at room temperature for 2 h. Following
washing the membranes with PBST containing 0.1% Tween-20 three
times, the bound antibodies were then detected using the secondary
goat anti-rabbit or goat anti-mouse antibodies for 1 h at room
temperature and protein signals were visualized by enhanced
chemiluminescence using ECL Western blotting detection reagents
(Beyotime Institute of Biotechnology, Haimen, China) for 1 min and
exposed to Kodak Biomax XAR film. The experiments were repeated
three times.
Statistical analysis
All data were analyzed using SPSS 22.0 software (IBM
Corp., Armonk, NY, USA). Results were expressed as the mean ±
standard deviation. One-way analysis of variance was used to
compare two groups of data. Multiple comparisons between the groups
were performed using the Student-Newman-Keuls method. P<0.05 was
considered to indicate a statistically significant difference.
Results
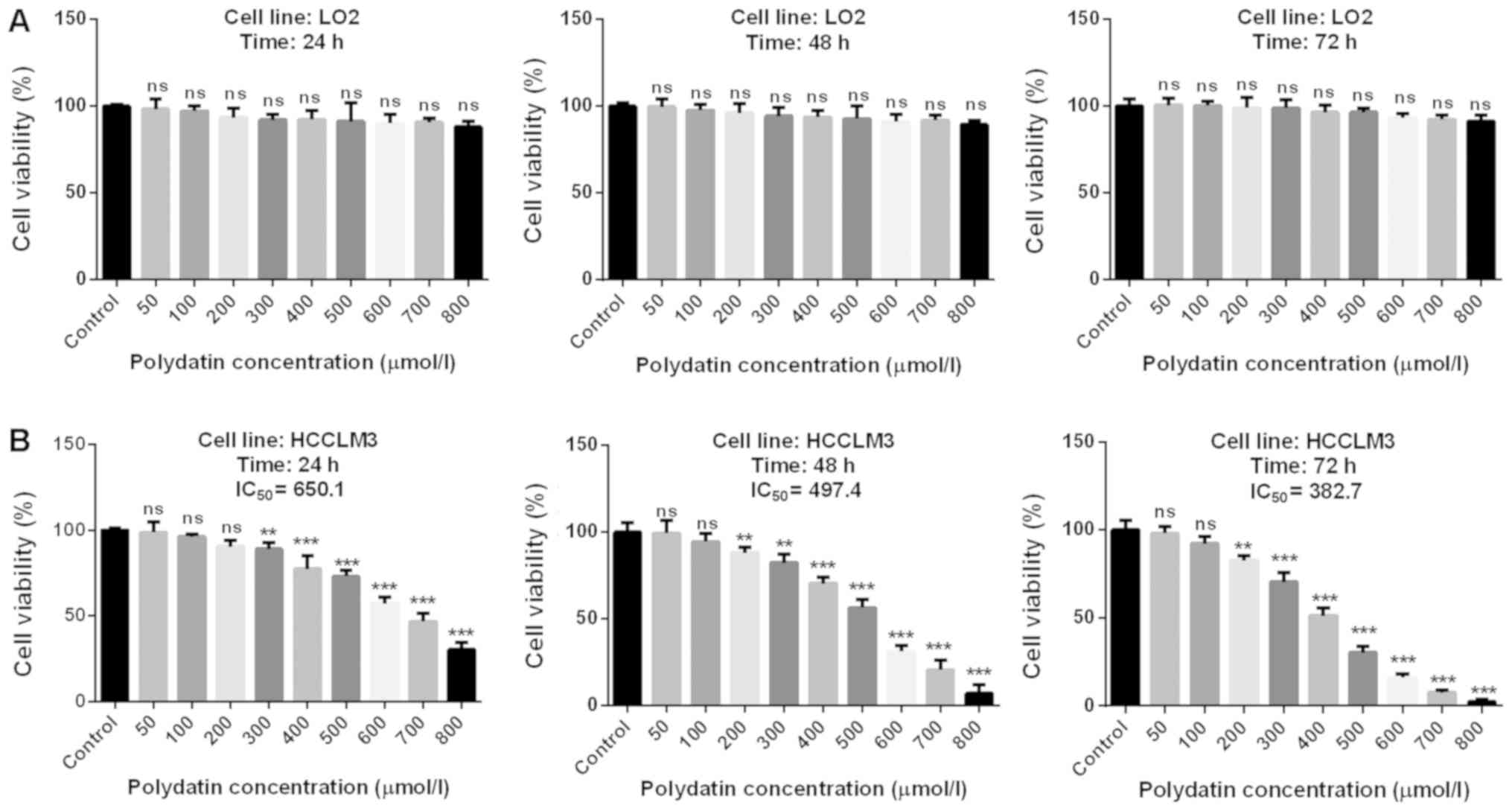
Cytotoxic effects of polydatin on
HCCLM3 cells
The associations between drug cytotoxicity and
dose/time-dependent drug efficacy were the first parameters
considered. In addition, an absence of toxicity of chemotherapeutic
drugs on human normal cells is essential when developing a novel
treatment, particularly for liver cancer, since the liver is the
major organ involved in drug metabolism.
Results revealed that polydatin was not toxic for
LO2 cells (Fig. 2A). Furthermore,
polydatin was significantly toxic to HCCML3 cells, at
concentrations of 300 and 200 µmol/l after 24 and 48 h,
respectively. However, concentrations <200 µmol/l did not
exhibit any cytotoxicity for all treatment durations. Furthermore,
for concentrations >200 µmol/l, polydatin displayed toxicity in
a dose-dependent manner. In combination with the drug half maximal
inhibiting concentration illustrated in Fig. 2B, these results suggested that
polydatin may effectively destroy HCC cells with low toxicity in
healthy human hepatic cells.
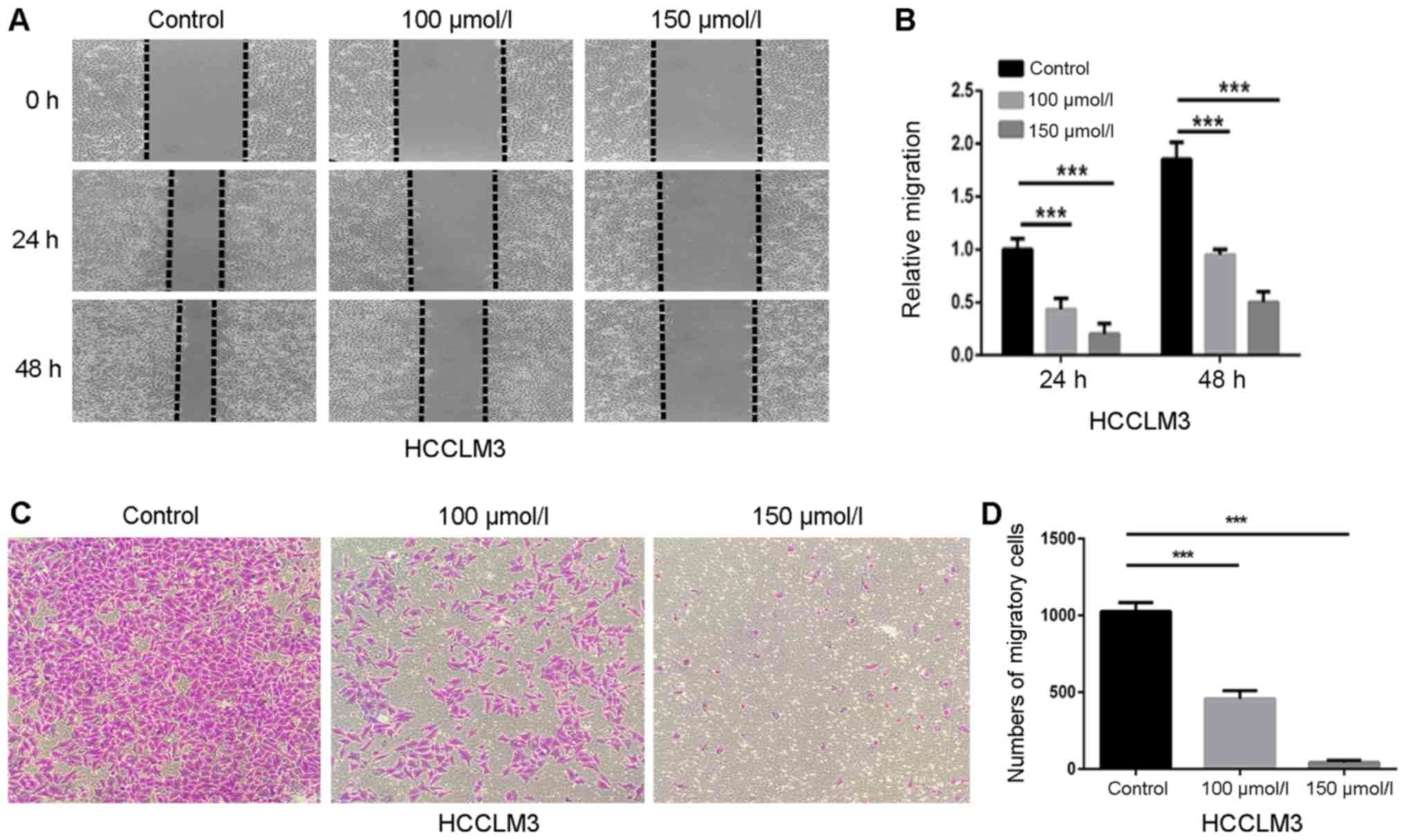
Effects of polydatin on HCCLM3 cell
migration
Wound healing and transwell chamber assays were
performed to investigate the effects of polydatin on the migration
of HCC cells. HCCLM3 cells were cultured with nontoxic doses of
polydatin (0–150 µmol/l) to avoid interference with cell toxicity.
Fig. 3A demonstrated that polydatin
decreased the wound healing of HCCLM3 cells in a dose-dependent
manner. In comparison with control cells, the relative migration
level of HCCLM3 cells treated with 150 µmol/l polydatin was
~0.27-fold at 48 h (Fig. 3B). In
addition, the migration assay demonstrated that migrated cells on
the lower side of the membrane were reduced in a dose-dependent
manner (Fig. 3C and D), which
confirmed the results of the wound healing assay. These data
suggested that polydatin may prevent HCCML3 migration in a
dose-dependent manner.
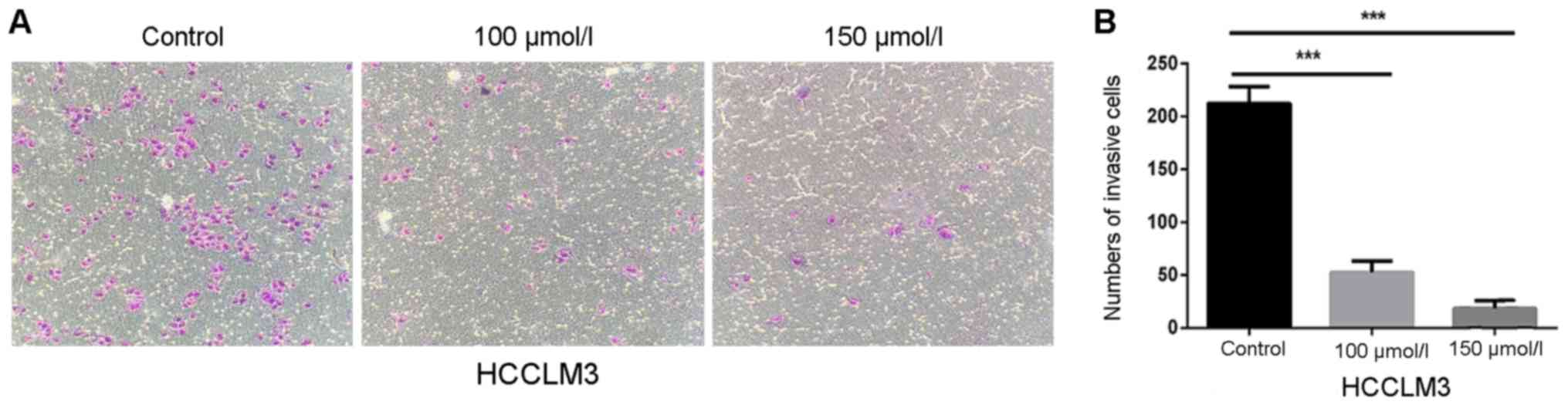
Effects of polydatin on HCCLM3 cell
invasion
Compared with cell migration, which only involves
mobility, cell invasion also involves the ability of cells to
degrade the extracellular matrix. Therefore, Transwell chambers
covered with Matrigel were used to detect the effects of polydatin
on HCC cell invasion. Fig. 4A and B
revealed that the number of invasive cells was significantly
decreased following polydatin treatment in a dose-dependent manner.
In comparison with control cells, the relative invasion of HCCLM3
cells treated with 150 µmol/l polydatin was ~0.09-fold. These
findings suggested that polydatin may inhibit the invasive ability
of HCCLM3 cells.
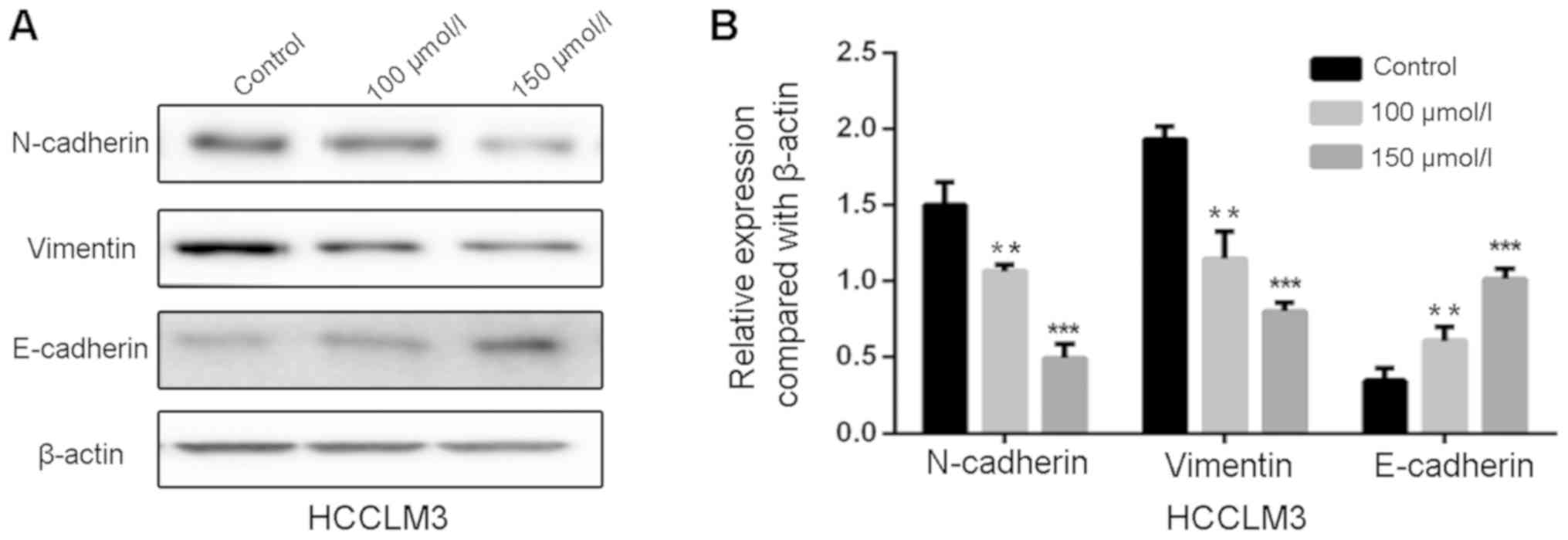
Polydatin inhibits EMT in HCCLM3
cells
EMT is a crucial process for migration and invasion
of cancer cells. In the present study, polydatin was suspected to
suppress migration and invasion of HCCLM3 cells by inhibiting EMT.
To further investigate the effects of polydatin on EMT, western
blotting was used to detect the EMT-associated markers in HCCLM3
cells following treatment with polydatin for 48 h. The results
suggested that the expression levels of the mesenchymal markers
N-cadherin and vimentin were decreased by polydatin in a
dose-dependent manner, whereas the expression levels of the
epithelial marker E-cadherin were increased (Fig. 5A and B). These findings suggested
that polydatin may prevent migration and invasion of HCCLM3 cells
by reversing the EMT process.
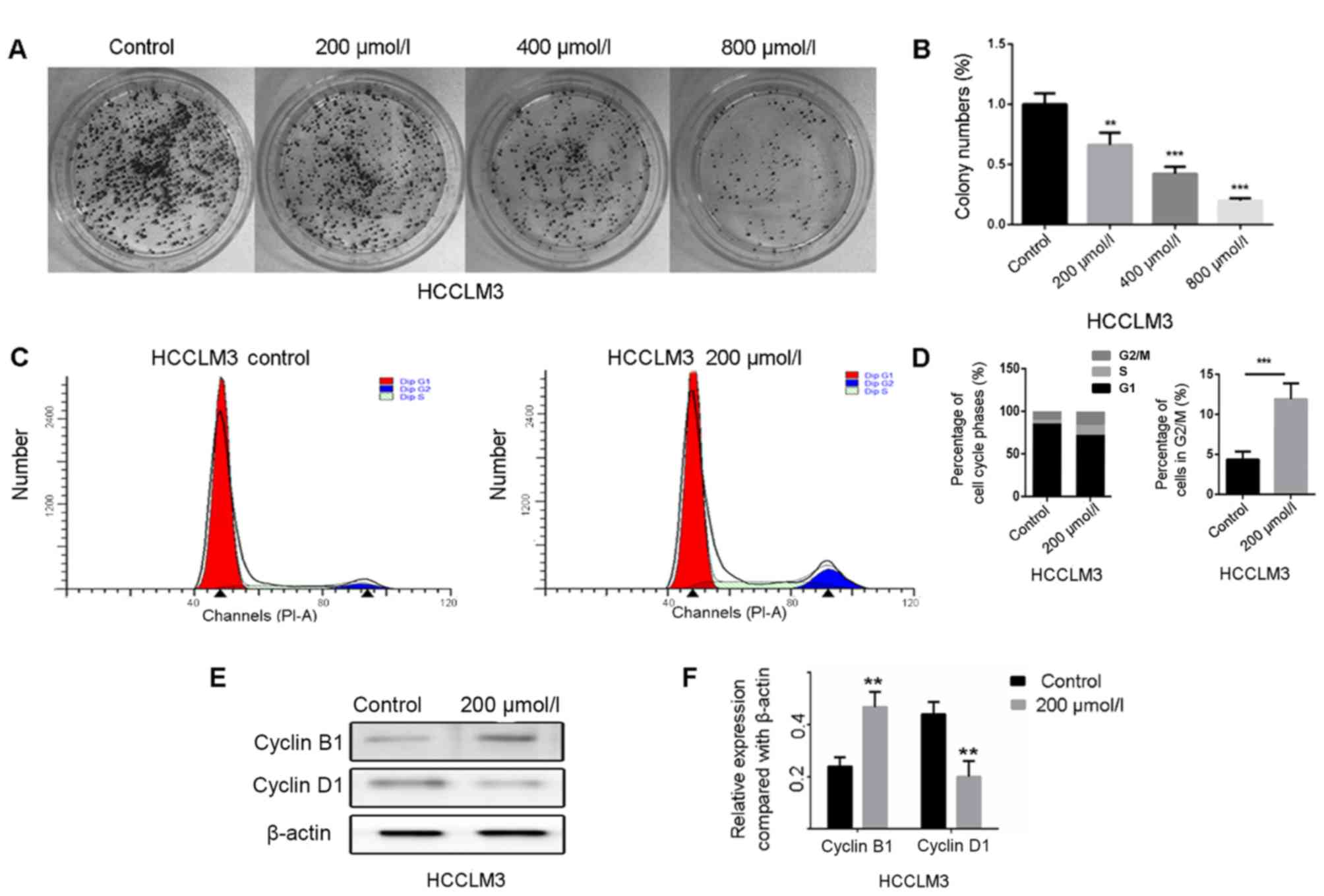
Anti-proliferative effects of
polydatin on HCCLM3 cells
Proliferation is an essential process for
tumorigenesis and development. In the present study, the colony
formation assay was performed to evaluate the proliferative ability
of HCCLM3 cells following treatment with polydatin. The results
demonstrated that polydatin significantly inhibited HCCLM3 cell
proliferation in a dose-dependent manner (Fig. 6A and B). It is generally accepted
that cell proliferation is controlled by the cell cycle. The effect
of polydatin on cell cycle progression was therefore assessed. As
presented in Fig. 6C and D,
polydatin arrested the HCCLM3 cell cycle at the G2/M
phase. To further confirm this result, the expression levels of the
proteins cyclin D1 and B1, which regulate the
G0/G1 and G2/M phases,
respectively, were analyzed by western blotting. The results
revealed that the expression levels of cyclin B1 were markedly
increased following polydatin treatment, whereas a decreased
expression of cyclin D1 was observed (Fig. 6E and F). These results suggested that
polydatin may arrest the cell cycle at the G2/M phase,
combined with a significantly reduced proportion of HCCML3 cells in
the G0/G1 phase. Taken together, these data
suggested that polydatin may inhibit HCCLM3 cell proliferation by
inducing cell cycle arrest at the G2/M phase.
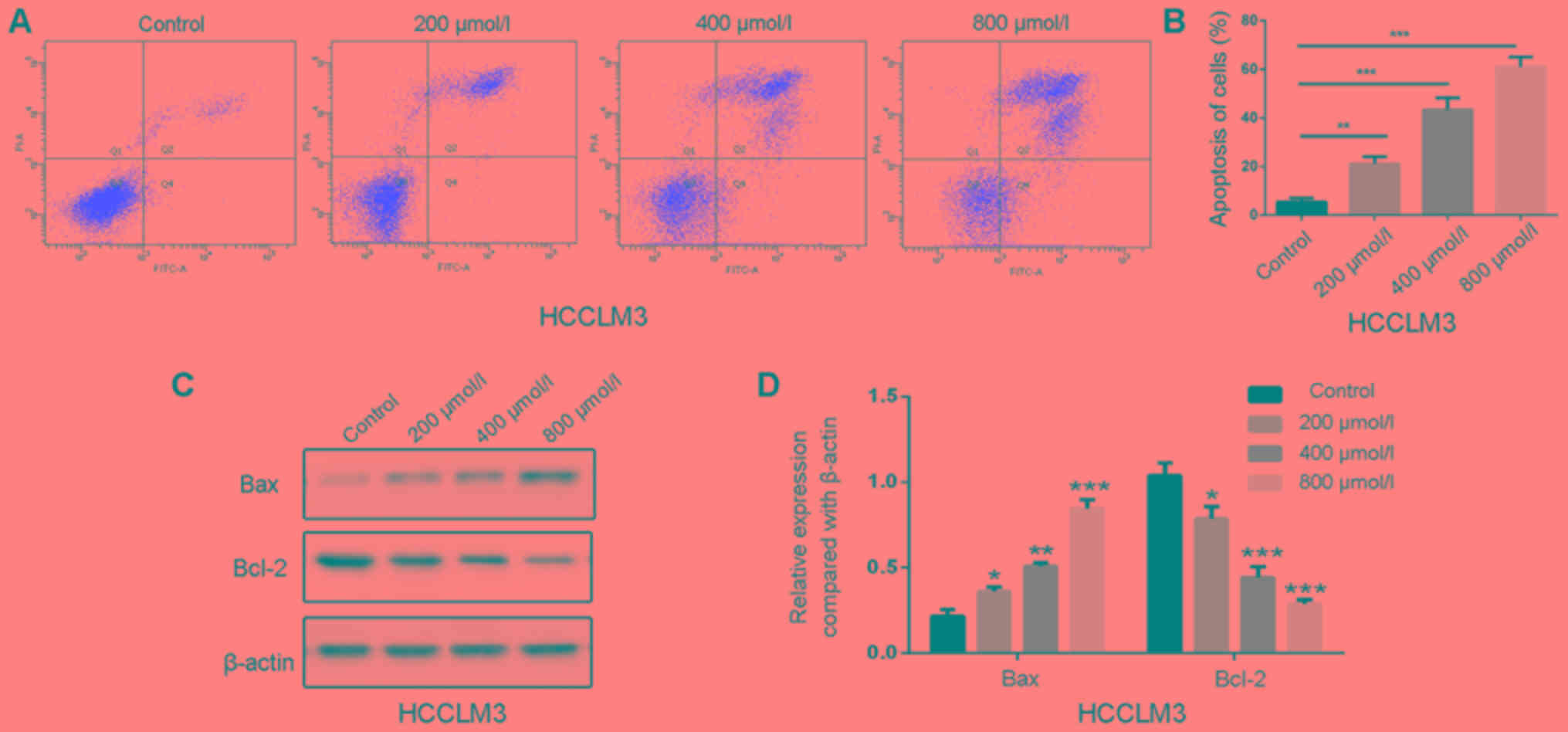
Polydatin induces apoptosis in HCCLM3
cells
Apoptosis is a primary type of cell death induced by
chemotherapeutic drugs, and the effects of polydatin on apoptosis
have been reported in human lung cancer cells (21). The effects of 48 h treatment of
polydatin on HCCML3 cell apoptosis were therefore evaluated. As
shown in Fig. 7A, polydatin induced
HCCML3 cell apoptosis in a dose-dependent manner. At a
concentration of 800 µmol/l, the percentage of apoptotic cells
reached ~60% compared with untreated cells (Fig. 7B).
Apoptosis is mediated by death receptors,
mitochondria and the endoplasmic reticulum (23). In the present study, the expression
levels of the pro-apoptotic factor B-cell lymphoma 2
(Bcl-2)-associated X protein (Bax) increased following polydatin
treatment, whereas the expression levels of the anti-apoptotic
factor Bcl-2 were decreased (Fig. 7C and
D). These results suggested that polydatin may induce HCCML3
cell apoptosis by regulating the expression of Bax and Bcl-2 in a
dose-dependent manner.
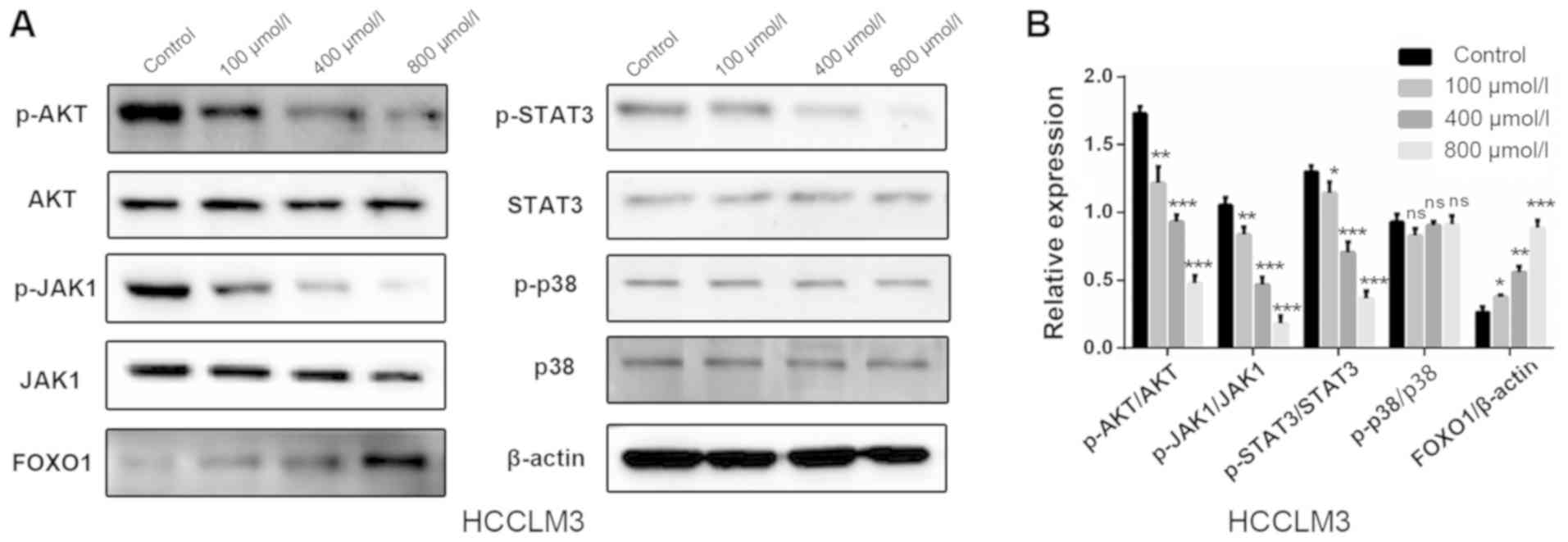
Polydatin inhibits HCC via the
AKT/STAT3-FOXO1 signaling pathway
Results from the present study demonstrated that
polydatin affected proliferation, apoptosis and EMT-associated
migration and invasion in HCCLM3 cells. The possible underlying
mechanism involved in these processes were therefore further
investigated. Numerous signaling pathways, including PI3K/AKT,
JAK1/STAT3 and p38, are known to serve important roles in the
progression of HCC, including the EMT process (24–27). The
effect of polydatin on these signaling pathways was thus evaluated.
Following treatment with increasing doses of polydatin for 48 h,
the expression levels of the central proteins involved in the
aforementioned pathways were evaluated in HCCLM3 cells by western
blotting. The following were detected: Activated AKT
[phosphorylated (p)-AKT, Ser 473], AKT, activated JAK1 (p-JAK1, Tyr
1034/1035), JAK1, activated STAT3 (p-STAT3, Tyr 705), STAT3,
activated p38 (p-p38, Thr 180/Tyr 182) and p38. The results
demonstrated that polydatin induced a downregulation of p-AKT,
p-JAK1 and p-STAT3 as compared with the control group; however, no
significant changes were observed in p-p38 levels (Fig. 8A and B). These results suggested that
polydatin may suppress HCC via the regulation of the PI3K/AKT and
JAK1/STAT3 signaling pathways. As a downstream gene of these two
signaling pathways, FOXO1 has garnered interest for its involvement
in EMT, in addition to its well-known involvement in cell
proliferation and apoptosis. The expression of FOXO1 was therefore
evaluated. The results revealed that following treatment of HHCML3
cells with polydatin, the expression levels of FOXO1 were
upregulated in a dose-dependent manner (Fig. 8A and B). These results suggested that
polydatin may exert anti-HCC effects via the AKT/STAT3-FOXO1
signaling pathway.
Discussion
Natural herbal medicines have gained increasing
attention in the development of chemotherapeutic regimens.
Polydatin is a glycoside of resveratrol, in which the glycoside
group is bound in the C-3 position, leading to alterations in its
biological properties. Compared with resveratrol, polydatin is more
efficiently absorbed by the digestive system via an active
mechanism using glucose carriers (28–30).
Polydatin is also more resistant to enzymatic oxidation. In
addition, polydatin preserves the active groups of resveratrol,
including the two benzene rings with a hydroxyl group and double
bonds between the two rings, whose anticancerous properties have
previously been demonstrated (31,32).
Polydatin may therefore have some anticancer potential. To the best
of our knowledge, studies investigating the anticancer potential of
polydatin are lacking (21,22), with none suggesting an anti-EMT
effect on HCC cells.
Tumor metastasis is a characteristic feature of
malignant tumors, which is primarily responsible for
cancer-associated mortality. Migration and invasion are essential
conditions for tumor metastasis. In the present study, polydatin
was demonstrated to inhibit the migration and invasion of HCCLM3
cells in a dose-dependent manner. In order to evaluate the anti-HCC
effects of polydatin, numerous proliferation-associated experiments
were performed, including colony formation efficiency, cell cycle
analysis and apoptosis analysis. Results revealed that polydatin
significantly inhibited proliferation and promoted apoptosis of
HCCML3 cells in a dose-dependent manner.
Previous studies have revealed that EMT leads to
increased cell migration and invasion in various types of cancer
(33,34). Subsequently, the present study
further explored the influences of polydatin on EMT. The results
revealed that treatment with polydatin increased the expression
levels of E-cadherin, and decreased expression levels of N-cadherin
and vimentin. These results suggested that polydatin may promote
EMT in HCCML3 cells in order to block their migration and
invasion.
Three common signaling pathways have proven critical
in the progression of HCC, including PI3K/AKT, JAK1/STAT3 and p38,
and are closely associated with EMT in cancer metastasis (35). These pathways were therefore
evaluated in the present study in HCCML3 cells following treatment
with polydatin. The activation of AKT leads to the loss of tumor
cell-cell junctions, disruption of tumor cell polarity and
morphological alterations of tumor cells, thus stimulating tumor
cell motility (36,37). Furthermore, activated AKT inhibits
the transcription of E-cadherin (24). Similarly, activation of STAT3
increases the mesenchymal level in various types of cancer cells
(25,26), whereas p38 participates in EMT in
colorectal cancer progression (27).
The results of the present study indicated that the levels of
phosphorylated AKT, JAK1 and STAT3 were markedly decreased
following treatment of HCCML3 cells with polydatin; however, no
change was observed in p38 and p-p38 expression. The protein p38 is
involved in oxidative stress regulation, suggesting that polydatin
may not increase intracellular reactive oxygen species, which is
consistent with the findings of previous studies (38,39).
FOXO1 belongs to the forkhead family and serves numerous roles,
including cellular differentiation, proliferation, cell cycle
progression, apoptosis and glucose metabolism (40). Previous studies have demonstrated
that the expression levels of FOXO1 are closely associated with the
EMT process in HCC (15,41). By searching the Kyoto Encyclopedia of
Genes and Genomes Pathway Database, FOXO1 was revealed to be a
downstream gene of PI3K/AKT and JAK1/STAT3 signaling pathways
(42,43). This suggested that polydatin may
control cell proliferation (G2/M arrest) and
EMT-associated migration and invasion, and promote apoptosis of HCC
cells via FOXO1 regulation. The expression of FOXO1 was therefore
assessed and was increased in HCCLM3 cells following polydatin
treatment. These results suggested that polydatin may serve
therapeutic roles via increasing FOXO1 expression, which was
regulated by PI3K/AKT and JAK1/STAT3 signaling pathways.
In conclusion, the present study demonstrated that
polydatin may inhibit proliferation via G2/M arrest,
suppress migration and invasion of HCC cells and promote HCC cell
apoptosis in a dose dependent-manner. Overall, polydatin exhibited
some therapeutic potential for HCC, and induced no toxicity in
normal human cells. The study suggested that polydatin may exert
therapeutic effects by inhibiting the AKT/STAT3-FOXO1 signaling
pathway, and reducing proliferation, apoptosis and EMT-related
migration and invasion. These findings may provide a novel
theoretical foundation for the development of polydatin-based
chemotherapeutic treatment for HCC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grants no. 81771894) and the
Fund of Scientific Research Innovation of The First Affiliated
Hospital of Harbin Medical University (grant no. 2018B009).
Availability of data and materials
The datasets used and/or data analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
JJ, YC, PL and XY conceived this study. JJ and YC
designed the experiments. JJ, YC, TD, MY and YZ performed the
experiments. TA and JZ conducted the statistical analysis. JJ and
YC wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Heindryckx F, Colle I and Van Vlierberghe
H: Experimental mouse models for hepatocellular carcinoma research.
Int J Exp Pathol. 90:367–386. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wei KR, Yu X, Zheng RS, Peng XB, Zhang SW,
Ji MF, Liang ZH, Ou ZX and Chen WQ: Incidence and mortality of
liver cancer in China, 2010. Chin J Cancer. 33:388–394.
2014.PubMed/NCBI
|
|
3
|
Yang CY, Chen YD, Guo W, Gao Y, Song CQ,
Zhang Q, Zheng NN, Han XJ and Guo CS: Bismuth ferrite-based
nanoplatform design: An ablation mechanism study of solid tumor and
NIR-triggered photothermal/photodynamic combination cancer therapy.
Adv Funct Mater. 17068272018. View Article : Google Scholar
|
|
4
|
Raza A and Sood GK: Hepatocellular
carcinoma review: Current treatment, and evidencebased medicine.
World J Gastroenterol. 20:4115–4127. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Abou-Alfa GK, Meyer T, Cheng AL,
El-Khoueiry AB, Rimassa L, Ryoo BY, Cicin I, Merle P, Park JW,
Blanc JF, et al: Cabozantinib (C) versus placebo (P) in patients
(pts) with advanced hepatocellular carcinoma (HCC) who have
received prior sorafenib: Results from the randomized phase III
CELESTIAL trial. J Clin Oncol. 36:2072018. View Article : Google Scholar
|
|
6
|
Bruix J, Qin S, Merle P, Granito A, Huang
YH, Bodoky G, Pracht M, Yokosuka O, Rosmorduc O, Breder V, et al:
Regorafenib for patients with hepatocellular carcinoma who
progressed on sorafenib treatment (RESORCE): A randomised,
double-blind, placebo-controlled, phase 3 trial. Lancet. 389:56–66.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kudo M, Finn RS, Qin S, Han KH, Ikeda K,
Piscaglia F, Baron A, Park JW, Han G, Jassem J, et al: Lenvatinib
versus sorafenib in frst-line treatment of patients with
unresectable hepatocellular carcinoma: A randomised phase 3
non-inferiority trial. Lancet. 391:1163–1173. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Puisieux A, Brabletz T and Caramel J:
Oncogenic roles of EMT-inducing transcription factors. Nat Cell
Biol. 16:488–494. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yopp AC and Singal AG: Epithelial to
mesenchymal transition expression profiles as predictive biomarkers
of recurrence following resection of HCC: Implications for current
clinical use and future stratification for systemic therapy. Ann
Surg Oncol. 21:3723–3724. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fransvea E, Angelotti U, Antonaci S and
Giannelli G: Blocking transforming growth factor-beta up-regulates
E-cadherin and reduces migration and invasion of hepatocellular
carcinoma cells. Hepatology. 47:1557–1566. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yamada S, Okumura N, Wei L, Fuchs BC,
Fujii T, Sugimoto H, Nomoto S, Takeda S, Tanabe KK and Kodera Y:
Epithelial to mesenchymal transition is associated with shorter
disease-free survival in hepatocellular carcinoma. Ann Surg Oncol.
21:3882–3890. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang CH, Guo ZY, Chen ZT, Zhi XT, Li DK,
Dong ZR, Chen ZQ, Hu SY and Li T: TMPRSS4 facilitates
epithelial-mesenchymal transition of hepatocellular carcinoma and
is a predictive marker for poor prognosis of patients after
curative resection. Sci Rep. 5:123662015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Boreddy SR, Pramanik KC and Srivastava SK:
Pancreatic tumor suppression by benzyl isothiocyanate is associated
with inhibition of PI3K/AKT/FOXO pathway. Clin Cancer Res.
17:1784–1795. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Aghazadeh S and Yazdanparast R:
Mycophenolic acid potentiates HER2-overexpressing SKBR3 breast
cancer cell line to induce apoptosis: Involvement of AKT/FOXO1 and
JAK2/STAT3 pathways. Apoptosis. 21:1302–1314. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dong T, Zhang Y, Chen Y, Liu P, An T,
Zhang J, Yang H, Zhu W and Yang X: FOXO1 inhibits the invasion and
metastasis of hepatocellular carcinoma by reversing ZEB2-induced
epithelial-mesenchymal transition. Oncotarget. 8:1703–1713.
2017.PubMed/NCBI
|
|
16
|
Zeng CX, Tang LY, Xie CY, Li FX, Zhao JY,
Jiang N, Tong Z, Fu SB, Wen FJ and Feng WS: Overexpression of
EPS8L3 promotes cell proliferation by inhibiting the transactivity
of FOXO1 in HCC. Neoplasma. 65:701–707. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kan H, Huang Y, Li X, Liu D, Chen J and
Shu M: Zinc finger protein ZBTB20 is an independent prognostic
marker and promotes tumor growth of human hepatocellular carcinoma
by repressing FoxO1. Oncotarget. 7:14336–14349. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu LT, Guo G, Wu M and Zhang WG: The
progress of the research on cardio-vascular effects and acting
mechanism of polydatin. Chin J Integr Med. 18:714–719. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao JP, Chen CX, Gu WL, Wu Q, Wang Y and
Lü J: Effects of polydatin on attenuating ventricular remodeling in
isoproterenol-induced mouse and pressure-overload rat models.
Fitoterapia. 81:953–960. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi YW, Wang CP, Liu L, Liu YL, Wang X,
Hong Y, Li Z and Kong LD: Antihyperuricemic and nephroprotective
effects of resveratrol and its analogues in hyperuricemic mice. Mol
Nutr Food Res. 56:1433–1444. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Y, Zhuang Z, Meng Q, Jiao Y, Xu J
and Fan S: Polydatin inhibits growth of lung cancer cells by
inducing apoptosis and causing cell cycle arrest. Oncol Lett.
7:295–301. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
De Maria S, Scognamiglio I, Lombardi A,
Amodio N, Caraglia M, Cartenì M, Ravagnan G and Stiuso P:
Polydatin, a natural precursor of resveratrol, induces cell cycle
arrest and differentiation of human colorectal Caco-2 cell. J
Transl Med. 11:2642013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Redza-Dutordoir M and Averill-Bates DA:
Activation of apoptosis signalling pathways by reactive oxygen
species. Biochim Biophys Acta. 1863:2977–2992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Larue L and Bellacosa A:
Epithelial-mesenchymal transition in development and cancer: Role
of phosphatidylinositol 3′ kinase/AKT pathways. Oncogene.
24:7443–7454. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen YD, Zhang Y, Dong TX, Xu YT, Zhang W,
An TT, Liu PF and Yang XH: Hyperthermia with different temperatures
inhibits proliferation and promotes apoptosis through the
EGFR/STAT3 pathway in C6 rat glioma cells. Mol Med Rep.
16:9401–9408. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Luwor RB, Baradaran B, Taylor LE, Iaria J,
Nheu TV, Amiry N, Hovens CM, Wang B, Kaye AH and Zhu HJ: Targeting
Stat3 and Smad7 to restore TGF-β cytostatic regulation of tumor
cells in vitro and in vivo. Oncogene. 32:2433–2441. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang B, Zhang L and Zhao L, Zhou R, Ding
Y, Li G and Zhao L: LASP2 suppresses colorectal cancer progression
through JNK/p38 MAPK pathway meditated epithelial-mesenchymal
transition. Cell Commun Signal. 15:212017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hollman PC, de Vries JH, van Leeuwen SD,
Mengelers MJ and Katan MB: Absorption of dietary quercetin
glycosides and quercetin in healthy ileostomy volunteers. Am J Clin
Nutr. 62:1276–1282. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Paganga G and Rice-Evans CA: The
identifcation of flavonoids as glycosides in human plasma. FEBS
Lett. 401:78–82. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Krasnow MN and Murphy TM: Polyphenol
glucosylating activity in cell suspensions of grape (Vitis
vinifera). J Agric Food Chem. 52:3467–3472. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Delmas D, Aires V, Limagne E, Dutartre P,
Mazué F, Ghiringhelli F and Latruffe N: Transport, stability, and
biological activity of resveratrol. Ann N Y Acad Sci. 1215:48–59.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chalal M, Klinguer A, Echairi A, Meunier
P, Vervandier-Fasseur D and Adrian M: Antimicrobial activity of
resveratrol analogues. Molecules. 19:7679–7688. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tsai JH and Yang J: Epithelial-mesenchymal
plasticity in carcinoma metastasis. Genes Dev. 27:2192–2206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mitra A, Mishra L and Li S: EMT, CTCs and
CSCs in tumor relapse and drug-resistance. Oncotarget.
6:10697–10711. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hsu CY, Lin CH, Jan YH, Su CY, Yao YC,
Cheng HC, Hsu TI, Wang PS, Su WP, Yang CJ, et al:
Huntingtin-interacting protein-1 is an early-stage prognostic
biomarker of lung adenocarcinoma and suppresses metastasis via
AKT-mediated epithelial-mesenchymal transition. Am J Respir Crit
Care Med. 193:869–880. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Attoub S, Arafat K, Hammadi NK, Mester J
and Gaben AM: AKT2 knock-down reveals its contribution to human
lung cancer cell proliferation, growth, motility, invasion and
endothelial cell tube formation. Sci Rep. 5:127592015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Liu S, Wang L, Wu Y, Hao J, Wang
Z, Lu W, Wang XA, Zhang F, Cao Y, et al: A novel PI3K/AKT signaling
axis mediates Nectin-4-induced gallbladder cancer cell
proliferation, metastasis and tumor growth. Cancer Lett.
375:179–189. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu J, Chang F, Li F, Fu H, Wang J, Zhang
S, Zhao J and Yin D: Palmitate promotes autophagy and apoptosis
through ROS-dependent JNK and p38 MAPK. Biochem Biophys Res Commun.
463:262–267. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ince S, Avdatek F, Demirel HH,
Arslan-Acaroz D, Goksel E and Kucukkurt I: Ameliorative effect of
polydatin on oxidative stress-mediated testicular damage by chronic
arsenic exposure in rats. Andrologia. 48:518–524. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ponugoti B, Dong G and Graves DT: Role of
forkhead transcription factors in diabetes-induced oxidative
stress, Exp. Diabetes Res. 2012:9397512012.
|
|
41
|
Du M, Wang Q, Li W, Ma X, Wu L, Guo F,
Zhao S, Huang F, Wang H and Qin G: Overexpression of FOXO1
ameliorates the podocyte epithelial-mesenchymal transition induced
by high glucose in vitro and in vivo. Biochem Biophys Res Commun.
471:416–422. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Duan S, Huang W, Liu X, Liu X, Chen N, Xu
Q, Hu Y, Song W and Zhou J: IMPDH2 promotes colorectal cancer
progression through activation of the PI3K/AKT/mTOR and
PI3K/AKT/FOXO1 signaling pathways. J Exp Clin Cancer Res.
37:3042017. View Article : Google Scholar
|
|
43
|
Jiang G and Huang C, Li J, Huang H, Jin H,
Zhu J, Wu XR and Huang C: Role of STAT3 and FOXO1 in the divergent
therapeutic responses of non-metastatic and metastatic bladder
cancer cells to miR-145. Mol Cancer Ther. 16:924–935. 2017.
View Article : Google Scholar : PubMed/NCBI
|