Introduction
Cluster of differentiation 151 (CD151) is a member
of the tetraspanins superfamily, and it is composed of
four-transmembrane-spanning proteins containing two extracellular
regions of unequal size and three short intracellular regions
(1). In various malignant tumors,
CD151 is overexpressed and characterized as a ‘facilitator’ for
tumor metastasis (2). Currently,
CD151 appears to assist metastasis of renal cell carcinoma
(3), osteosarcoma (4), hepatocellular carcinoma (5), and prostate cancer (6). CD151 is a positive regulator of
TGF-β-induced signaling in cancer metastasis (3). Therefore, CD151 is thought to be a
suitable target molecule for various malignant tumors (7).
Clear cell sarcoma (CCS) of soft tissue primarily
affects young adults (8,9). Radical surgical resection is the first
choice for treatment of CCS. However, in contrast to other
malignant soft tissue tumors, CCS often possesses a robust
lymphatic metastatic activity (8–11).
Therefore, the rate of local recurrence is as high as 84%, while
the rate of late metastases is as high as 63%, which is associated
with the 5 to 20-year survival rate of 67–10% (12). Unfortunately, CCS is relatively
insensitive to conventional soft tissue sarcoma chemotherapy
regimens. These unfavourable clinicopathological features
encouraged us for new targeted therapies development of CCS.
Unfortunately, the rare instances of CCS cases not
only pose an impediment to know the molecular mechanisms that are
responsible for metastatic activity of CCS, but also create
obstacles to develop novel therapies. To overcome this limitation,
we have recently generated an orthotropic metastatic model of CCS
through xenoplanting human clear sarcoma cells, HS-MM, to
SCID-beige mice (13). This tumor
model very well reflects clinicopathobiological features of CCS,
i.e., high lymphatic metastatic activity and distant
metastasis.
Furthermore, by serial in vivo passaging of
HS-MM from peritoneally disseminated tumor, we established a HS-MM
cell clone, designated as HS-MMhigh, which harbored the
prominent lymphatic invasion and metastatic activity. In this
study, we found that CD151, which is recently designed to be a
target molecule to regulate cancer progression (7), is related to metastatic activity of CCS
in the animal models.
Materials and methods
Antibodies
For intraperitoneal injection, anti-CD151 antibody
was obtained from ascites of nude mice (BLAB/c nu/nu, female, 8
weeks old) after intraperitoneal injection of hybridoma cells.
Hybridoma cells, which produce anti-CD151 IgG1κ antibody [clone
50-6 (14), CRL-2696] were purchased
from American Type Culture Collection (Manassas, VA, USA).
For immunoblotting, a murine antibody specific to
CD151 (clone 11G5a), and rabbit GAPDH antibody were purchased from
Abcam Inc. (Cambridge, MA, USA) and Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany), respectively. Rabbit monoclonal SMAD3
antibodies (clone C67H9) and phospho-SMAD3 (Ser423/425) (clone
C25A9) were purchased from Cell Signaling Technology (CST, Inc.,
Danvers, MA, USA).
Alexa Fluor 555-conjugated anti-rabbit and Alexa
Fluor 488-conjugated anti-mouse antibodies were purchased from
Invitrogen; Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
Control murine antibody was isolated by Protein A-affinity
chromatography from normal mouse sera (Caltag Lab, Burlingame, CA,
USA).
Xenografts
The experimental protocol was approved by the Animal
Care Committee of Gifu Graduate School of Gifu, Japan (approval no.
H30-32). Detailed procedure of orthotropic metastatic model of CCS
has been previously described (13).
Briefly, SCID-Beige
(CB17.Cg-PrkdcscidLystbg-J/CrlCrlj) mice were
purchased from Charles River Laboratories, Japan (Sizuoka, Japan).
A CCS of soft tissue cell line, HS-MM, was previously established,
characterized, and maintained as a stock in our laboratory
(15,16). HS-MM cells (1.2×106) were
subcutaneously injected into the soft tissue of the thigh of
12-week-old SCID-Beige mice. In another independent experiment,
1.0×106 HS-MM cells were similarly injected into
10-week-old SCID-Beige mice. Tumor volume was measured by calipers
using the following equation: tumor volumes (mm3) = 4/3π
× [a/2] × [b/2]2, where ‘a’ and ‘b’ correspond to the
longest and shortest diameter measured twice a week. After five
weeks later, when tumor volumes reached near 1.0 cm3,
mice were randomly divided into two groups (n=4 and n=3 for each
group), and these mice were intraperitoneally inoculated with or
without 3 mg of anti-CD151 antibody (clone 50-6). Two weeks later,
mice were sacrificed to examine the extent of metastasis. The
animals were euthanized after anesthesia, and every effort was made
to minimize suffering. The xenografts and metastatic tissues were
excised, formalin fixed, paraffin embedded, and sectioned for
histopathological analysis.
Immunofluorescence staining
Immunofluorescence staining was performed as
previously described (17). Briefly,
cells were incubated with a murine anti-CD151 antibody (clone
11G5a) for 1 h at 4°C, washed with PBS twice, and then incubated
with Alexa Fluor 488-conjugated anti-mouse antibody (1:200
dilution) for 30 min at 4°C. After re-washing with PBS, the cells
were analyzed with a Guava EasyCyte cell analyzer (Guava
Technologies, Inc., Hayward, CA, USA). Guava easyCyte™ flow
cytometry system software was used to obtain the one parameter log
histogram.
Immunoblotting
Immunoblotting was performed according to a
previously described method (18),
with the modification proposed by Towbin et al (19). Samples were analyzed by
electrophoresis on SDS-containing polyacrylamide gels under
reducing conditions. The separated proteins were then transferred
to polyvinylidene difluoride membranes (EMD Millipore Co.,
Billerica, MA, USA) and probed with various antibodies.
Immunoreactivity was assessed using the Western Blotting Detection
Kit (Promega, Madison, WI, USA). The immunoblot band was quantified
by densitometry using LI-COR C-DiGit Blot Scanner imaging software
version 3.1 (LI-COR Biosciences, Lincoln, NE, USA) and was
normalized to the GAPDH band as previously reported (20).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
cDNA synthesis from total RNA, and subsequent PCR
experiments were performed using the Reverse Transcription
Polymerase Chain Reaction Kit (Takara Bio, Inc., Otsu, Japan)
according to manufacturer instructions and as previously described
(18). Briefly, the first strand
cDNA was synthesized using a random 6-mer primer. The specific
reaction conditions were 42°C for 30 min, followed by 95°C for 5
min, and then finally 5°C for 5 min.
qPCR reactions were performed using the FastStart
Essential DNA Green Master Mix according to the manufacturer's
instructions (Roche Diagnostics, GmbH, Mannheim, Germany) using a
LightCycler (Roche Diagnostics) as previously described (21). cDNA (2 µl each) was diluted with PCR
mix containing a 0.2 pmol of primer to a final volume of 20 µl. The
following qPCR primers were used for the real-time RT-PCR:
CD151 forward 5′-CATCGCTGGTATCCTCG-3′ and reverse
5′-CTCGCTGCCCACAAAG-3′, and GAPDH forward
5′-GAAATCCCATCACCATCTTCCAGG-3′ and reverse
5′-GAGCCCCAGCCTTCTCCATG-3′. The run protocols used included a
denaturation program (95°C for 10 min), an amplification and
quantification program repeated 45 times (95°C for 10 sec, 60°C for
10 sec, 72°C for 15 sec), and a melting curve program (60-95°C with
a heating rate of 0.1°C per second and a continuous fluorescence
measurement).
To ensure that SYBR Green was not incorporated,
resulting in primer dimers or non-specific amplification in the
qPCR runs, the CD151 and GAPDH PCR products were
analyzed by polyacrylamide gel electrophoresis in preliminary
experiments. Single intense bands at their respective expected
sizes were observed. The samples were cultured in triplicates, and
the expression of each target gene was analyzed with a LightCycler
system using the 2−ΔΔCT method described by Livak and
Schmittgen (22). For each
triplicate set, the ΔCT values were normalized to GAPDH
expression in both control and target cells. The values of the
target group were then calculated as the fold change relative to
the mean values of the control group, i.e., original HS-MM cells or
HS-MMhigh cells treated by a green fluorescent protein
(GFP)-siRNA duplex (control; set to 1.0). Following this, the
standard deviations were calculated for the triplicate sets, and
the fold changes for the target genes were plotted/calculated.
siRNA-mediated gene silencing
The detailed procedure followed for gene silencing
has been described previously (23).
In this study, we used the FlexiTube GeneSolution GS977, which is
composed of 4 siRNAs: SI02777257, SI02777250, SI04434570, and
SI03070018 (Qiagen, Inc., Valencia, CA, USA), to silence the
CD151 gene. A green fluorescent protein (GFP)-siRNA duplex
with the target sequence 5′-CGGCAAGCUGACCCUGAAGUUCAU-3′ was used as
a non-silencing control. The siRNAs were transfected into cells
with Lipofectamine RNAiMAX (Invitrogen; Thermo Fisher Scientific,
Inc.). At 72 h post transfection, the cells were used for
subsequent experiments.
Cell proliferation and Matrigel
invasion assays
Cell proliferation was evaluated by counting the
number of viable cells as previously described (24). Briefly, 1×104 cells were
cultured in triplicates in standard 35-mm tissue culture dishes (BD
Falcon; BD Biosciences, San Jose, CA, USA). After 24, 48, and 72 h,
the live cells were counted. The assay was performed in triplicates
and repeated twice.
The invasiveness of the cultured cells was
determined using 24-well Corning BioCoat Matrigel Invasion Chamber
Plates (Discovery Labware Inc., Bedford, MA, USA) according to the
manufacturer's protocol and as described previously (23). Briefly, 5×104 cells were
placed in the upper compartment of the invasion chamber. After 48 h
of incubation with DMEM containing 10% (lower chamber) or 2% (upper
chamber) FBS, the non-invading cells were gently removed from the
filter by swiping with a cotton-tipped swab. The cells on the lower
surface of the filter were counted under a microscope.
Statistical analysis
We repeated experiments twice to evaluate CD151
expression in HS-MM and HS-MMhigh cells. Xenotransplant
assay of anti-CD151 inoculation was repeated twice. To assess the
involvement of CD151 in invasion activity, we performed two
independent experiments. The zymography assay was repeated three
times and quantification of enzymatic activity assay was repeated
twice. Statistical analysis was performed by Student's t-test for
unpaired observations or ANOVA using Tukey's test. Findings with
P<0.05 were considered significant.
Zymography analysis
Functional activity of matrix metalloproteinase-9
(MMP-9) was evaluated by gelatin zymography, as previously
described by Heussen and Dowdle (25). Briefly, cells were incubated with
serum-free DMEM for 24 h. Supernatant of culture medium was
collected and mixed with non-reducing SDS sample buffer. After
SDS-PAGE on 10% polyacrylamide gels containing 1 mg/ml gelatin, SDS
was removed from the gels by incubating in 2.5% Triton X-100 for 1
h at room temperature. Subsequently, the gels were incubated in a
buffer containing 50 mM Tris-HCl, 5 mM CaCl2, pH 7.6,
for 24 h at 37°C, and stained with Coomassie blue R 250 (0.25%).
Proteolytic activities of MMP-9 were detected as clear bands
against the blue background of stained gelatin.
MMP-9 activity quantification
assay
The MMP-9 total activity (already active plus latent
MMP-9) was assayed using QuickZyme Human MMP-9 Activity Assay kit
(QuickZyme BioSciences, Leiden, The Netherlands) according to the
manufacturer's instructions. Briefly, culture supernatant MMP-9 was
captured by specific antibody to MMP-9. After treatment with
p-aminophenyl mercuric acetate, active MMP-9 altered pro-detection
enzyme to active detection enzyme. Subsequently, this active
detection enzyme recognized peptide substrate to form colored
product, which was measured at 405 nm. Assay was performed in
triplicate.
Cytostaining
Cells were fixed with 4% (m/v) paraformaldehyde,
permeabilized with 0.1% Triton X-100, and blocked with 10% goat
serum. Subsequently, cells were incubated with 1 µg/ml rabbit
anti-phospho-SMAD3 (Ser423/425) and murine anti-CD151 (clone 11G5a)
antibodies at room temperature for 1 h. After washing with PBS,
cells were incubated with 1:200 diluted Alexa Fluor 555-conjugated
anti-rabbit antibody and Alexa Fluor 488-conjugated anti-mouse
antibody (Invitrogen; Thermo Fisher Scientific, Inc.). After
staining, images were acquired with the help of a confocal laser
scanning microscope (Leica TCS SP8; Leica Microsystems GmbH,
Wetzlar, Germany).
Results
Increased CD151 expression in HS-MM
clear cell sarcoma cells with high metastatic activity
As previously reported, metastasis of HS-MM cells to
lung, liver, and lymph nodes was observed in xenoplanted SCID-Beige
mice (13). Notably, peritoneal
dissemination was also found in xenoplanted mice. Serial in
vivo passaging of peritoneally disseminated tumor cells
accelerated metastasis following implantation. After four passages,
we obtained HS-MM cells with high-metastatic activity, and these
cells were designated HS-MMhigh.
As recent advances indicate that CD151 enhances
metastasis of various malignant tumors, we examined the expression
status of original HS-MM and HS-MMhigh cells.
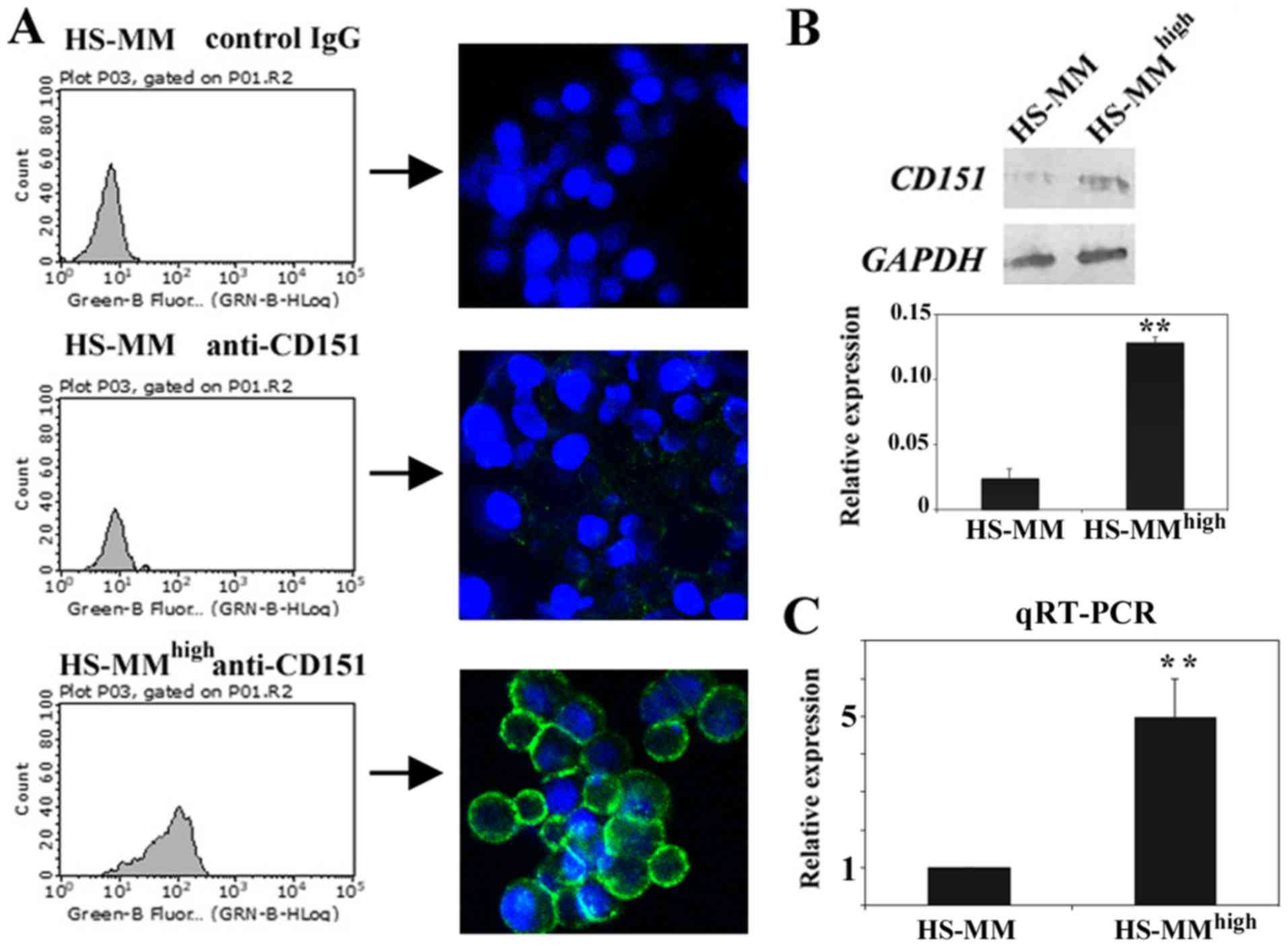
Immunofluorescent staining followed by cell analyzer
analysis revealed that HS-MMhigh cells expressed
significantly higher levels of cell surface CD151 compared with
those of original HS-MM cells (Fig.
1A). Subsequent immunoblotting also demonstrated that CD151
protein levels were more abundant in the cell lysates of
HS-MMhigh cells than they were from those derived from
HS-MM cells (Fig. 1B). RT-qPCR
showed that transcription of the CD151 gene is significantly
upregulated in HS-MMhigh cells compared to HS-MM cells
(Fig. 1C). We concluded that CD151
was quite abundantly expressed in HS-MMhigh cells
relative to expression in the original HS-MM cells, and this was
increase in expression was detectable at the cell membrane surface,
in total proteins, and at the transcript level. This experiment was
performed in duplicate, and both times we obtained similar
results.
Anti-CD151 antibody treatment
suppressed metastasis of HS-MM cells in xenoplanted SCID-Beige
mice
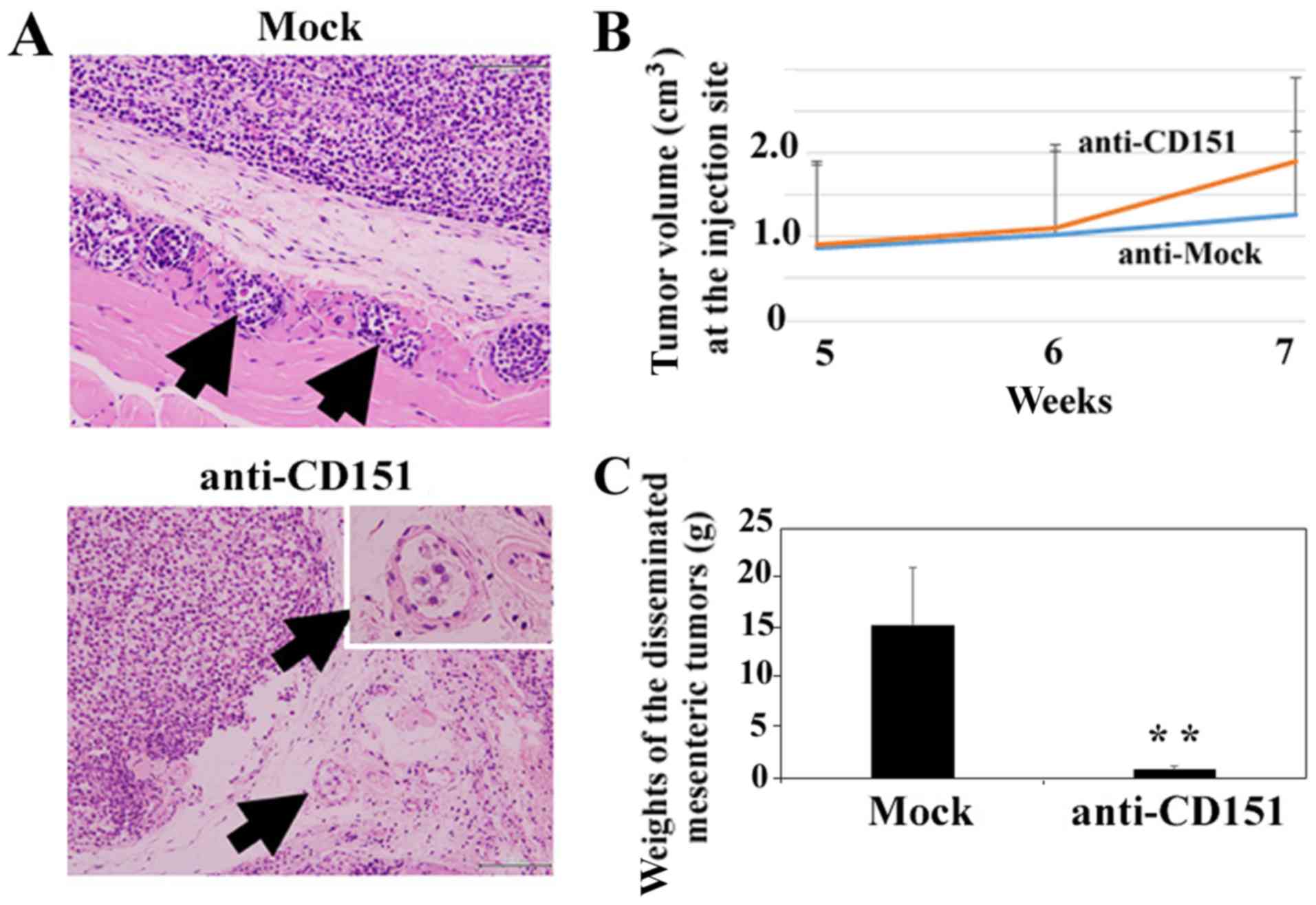
As demonstrated in Fig.
2A, HS-MMhigh cells showed high levels of invasion
in the absence of anti-CD151 antibody inoculation (Fig. 2A, Mock). High numbers of
HS-MMhigh cells within lymphatic vessels are indicated
by arrows, and a single inoculation of anti-CD151 antibody markedly
impaired the infiltration of HS-MMhigh cells into
lymphatic vessels (Fig. 2A,
anti-CD151). We did observe a small number of HS-MMhigh
cells in lymphatic vessels following treatment as indicated by
arrows; however, anti-CD151 antibody did not alter the tumor volume
of HS-MMhigh cells at the injection site as demonstrated
in Fig. 2B (Student's t-test.
P=0.37). In contrast, administration of anti-CD151 antibody
significantly decreased total weights of collected, disseminated
mesenteric tumors (Student's t-test, P=0.0036).
Representative data using an individual experiment
is shown. We also obtained similar results from another independent
experiment, in which 1.0×106 HS-MMhigh cells
were injected into 10-week-old SCID-Beige mice (n=3 for each
group). Given this, it is clear that anti-CD151 antibody treatment
suppressed metastasis of HS-MMhigh cells in xenoplanted
SCID-Beige mice.
siRNA-mediated gene silencing of CD151
decreased the Matrigel-invasion activity of HS-MMhigh
cells
Further, we investigated if CD151 plays a role in
the invasion of HS-MMhigh cells by using a Matrigel
invasion assay. Successful downregulation of CD151 mRNA
levels (>90% as determined by RT-qPCR) was achieved using any of
4 siRNAs (SI02777257, SI02777250, SI04434570, and SI03070018;
Qiagen). Both SI02777257 (indicated by siCD151#1) and SI02777250
(indicated by siCD151#2) siRNA treatments decreased CD151 protein
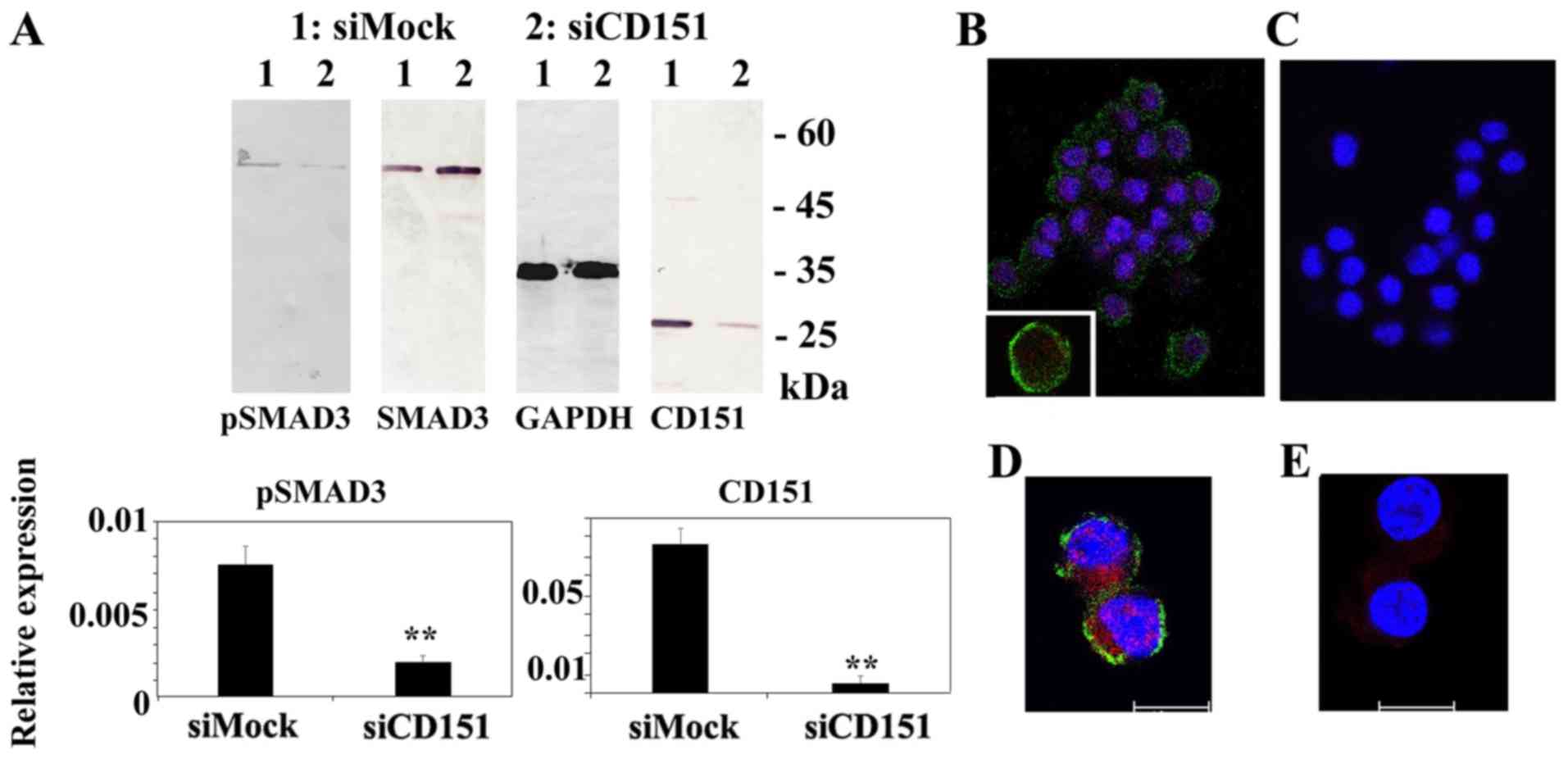
expression. Representative data are shown in Fig. 3A. Downregulation of CD151
using SI02777257 did not affect the cell growth of
HS-MMhigh cells in vitro (Student's t-test,
P=0.54, Fig. 3B). We obtained
similar results using SI02777250 siRNA. In contrast, downregulation
of CD151 using SI02777257 (indicated by siCD151#1) or
SI02777250 (indicated by siCD151#2) significantly reduced Matrigel
invasion activity (ANOVA, Tukey's test. P<0.05, Fig. 3C).
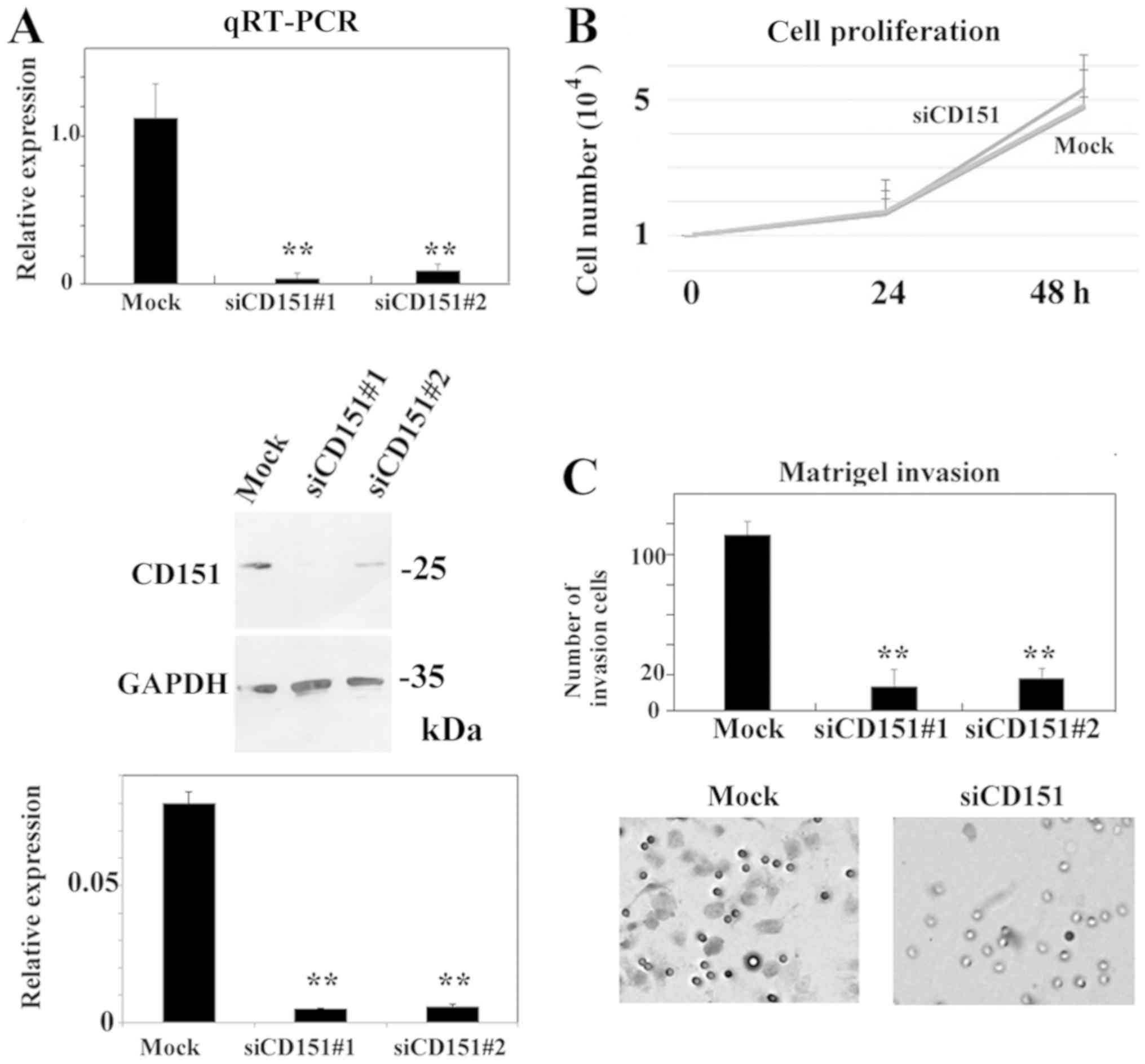 | Figure 3.siRNA-mediated gene-silencing of
CD151 does not affect cell proliferation, but significantly
decreases Matrigel invasion activity of HS-MMhigh in
vitro. (A) Both SI02777257 (indicated by siCD151#1) and
SI02777250 (indicated by siCD151#2) siRNA treatments reduced CD151
expression at both mRNA and protein level. Reverse
transcription-quantitative-polymerase chain reaction indicated that
both SI02777257 and SI02777250 significantly reduced CD151
expression. Immunoblotting also demonstrated little CD151 protein
in SI02777257 and SI02777250 siRNA treated HS-MMhigh
cells. The intensity ratio of each band to GAPDH is shown. The data
represent the mean ± SD of triplicate assays. ANOVA, Tukey's
t-test. was performed to confirm the significant differences,
**P<0.01 vs. Mock group. (B) Representative cell proliferation
assay using siRNA targeting CD151. At 24 h, the cell number
was 1.67±0.47 (Mock), 1.67±0.67 (SI02777257), 1.73±0.35
(SI02777250). Respective numbers at 48 h were 5.33±0.75 (Mock),
4.77±0.30 (SI02777257), and 4.83±1.07 (SI02777250). The data
represent mean ± SD in triplicate assays. ANOVA, Tukey's t-test.
P>0.5). Data using Mock and SI02777257 siRNAs are shown in this
cell proliferation line graph. (C) SI02777257 and SI02777250 siRNAs
specific to CD151 significantly reduced Matrigel invasion
activity of HS-MMhigh cells. The number of invading
cells was 113.7±6.42 (Mock), 13.7±8.33 (SI02777257, indicated by
#1), and 19.3±11.0 (SI02777250, indicated by #2). Data from
triplicate assay are expressed as the mean ± SD (n=3; ANOVA,
Tukey's t-test. **P<0.01 vs. Mock group). A representative image
of Matrigel invasion assay using Mock and SI02777257 siRNAs are
shown. Cells that migrated to the undersurface of the membrane are
shown. Original magnification, ×100. CD151, cluster of
differentiation 151; SD, standard deviation; siRNA, small
interfering RNA. |
Our results indicate that silencing of CD151
decreased the Matrigel invasion activity of metastatic
HS-MMhigh cells without affecting cell proliferation.
Representative data are shown in Fig.
3. We also obtained similar results from another independent
experiment.
siRNA-mediated gene silencing of CD151
decreased MMP-9 activity and phosphorylation status of in SMAD3 of
HS-MMhigh cells
CD151 correlates with MMP-9 activity in melanoma,
osteosarcoma, hepatocellular carcinoma, and a number of other
malignant tumors (26–29). Additionally, recent evidence
indicates that CD151 appears to increase phosphorylation status of
SMAD3, an event that can lead to epithelial-mesenchymal transition
in renal cell carcinoma (3). Given
this, we investigated if CD151 participates in MMP-9 activation and
active phosphorylation status of SMAD3.
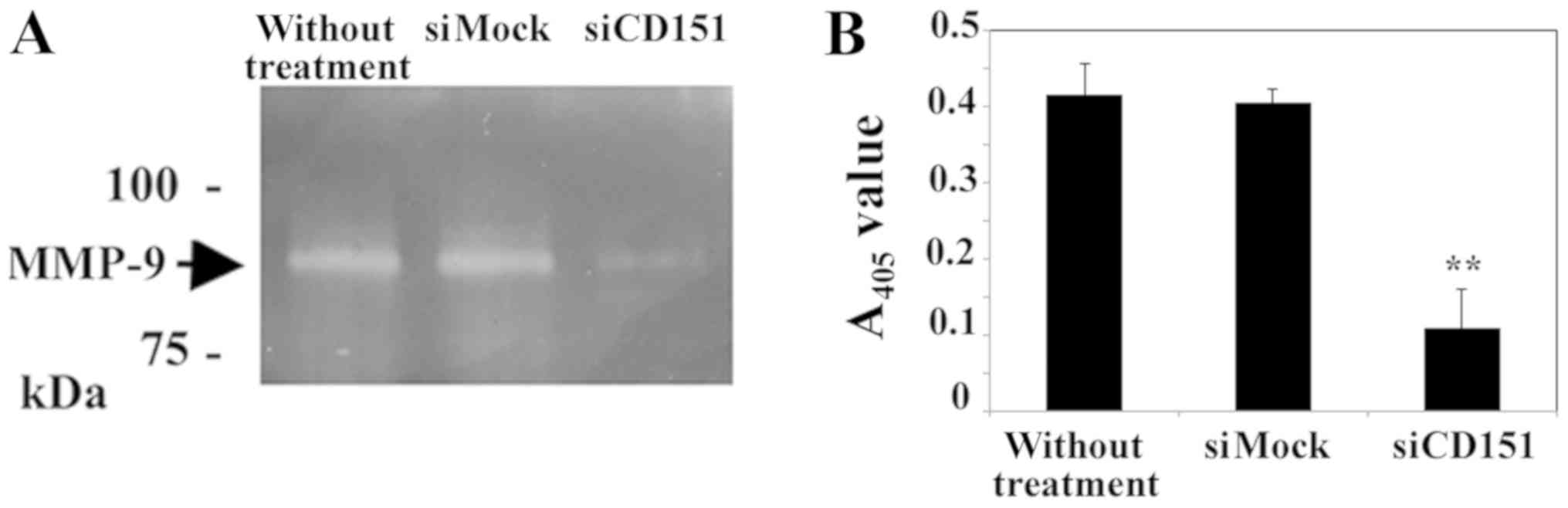
Results of a gelatin zymography and quantification
assay indicated that siRNA-mediated downregulation of CD151
decreased MMP-9 activation in HS-MMhigh cells. The MMP-9
active band in CD151 downregulated HS-MMhigh cells is
weak when compared to that of control cells (Fig. 4A). Furthermore, the present
quantification assay demonstrated that the amount of already active
plus latent MMP-9 was decreased by downregulation of CD151
(Fig. 4B).
Additionally, downregulation of CD151 impaired the
phosphorylation status and nuclear transport of SMAD3 (Fig. 5). Immunoblot analysis demonstrated
little or no phospho-SMAD family member 3 (pSMAD3), band in CD151
knockout HS-MMhigh cells. Immunocytostaining indicated
that HS-MMhigh cells exhibited CD151 immunoreactivity at
the cell membrane surface (green staining) and pSMAD3 reactivity in
both cytoplasm and nucleus (red staining). In contrast,
downregulation of CD151 reduced the immunoreactivity of both CD151
and pSMAD3.
Representative data using SI02777257 siRNA is
presented in Figs. 4 and 5. Similar results were obtained using
another siRNA, SI02777250, to silence the CD151 gene.
Discussion
CCS is a rare, high-grade malignant soft tissue
tumor (8,12). This tumor was previously known as
‘melanoma of soft part’ (9), but it
is now thought to be derived from the neural crest (30,31).
Most CCS tumors are characterized by a t(12;22)(q13;q12)
translocation harboring an EWS-ATF1 fusion gene (32). The resulting fusion protein directly
activates expression of the melanocyte master transcription factor
that drives the same down-stream pathways in both CCS and melanoma
(33). As a result, this chimeric
molecular alteration leads to significant clinicopathological
parallels between CCS and melanoma.
Surgical resection, if necessary, in combination
with adjuvant radiotherapy is the clinical mainstay for treatment
of patients diagnosed with CCS. Robust lymphatic metastatic
activity, however, often impedes effective treatment, and currently
the overall survival rate for patients diagnosed with this
malignancy is approximately 60% for 5 years. In this context, it is
important to understand the molecular mechanisms, underlying the
elevated lymphatic metastatic activity of CCS.
The present study strongly suggests an important
role for CD151 in lymphatic invasion, and we identify potent
suppressor activity of anti-CD151 antibody in the context of
lymphatic metastases using an orthotropic metastatic model of CCS.
Specifically, we demonstrate that CD151 is highly expressed in
HS-MMhigh cells, and that these cells possessed an
elevated potential for lymphatic invasion and metastasis compared
to that of the original HS-MM cells. Lymphatic infiltration of
HS-MMhigh cells was also found at transplant sites in
the control mice (Fig. 2A, Mock),
but was absent at these sites in anti-CD151 treated mice (Fig. 2A, anti-CD151). Additionally,
treatment with anti-CD151 significantly abrogated intraperitoneal
lymph node metastasis (Fig. 2C). We
expect that the combination of local surgical resection with
anti-CD151 treatment may provide significant improvement in
clinical outcomes for patients diagnosed with CCS.
We also studied the molecular pathway by which CD151
influenced lymphatic invasion and lymph node metastasis in our
Xenoplant model. Metastasis is a complex process composed of
consecutive steps, including initiation of local invasion,
lymphovascular invasion, migration to a distant site,
extravasation, formation of a micrometastasis, and, finally, the
development of overt metastatic tumors (19). Certain steps, especially initial
steps or those that occur during local invasion and at the end of
the cascade, appear to be more challenging for tumor cells
(20). The tetraspanin molecule
CD151 is believed to be a ‘facilitator’ of tumor metastasis. The
role of this molecule in the promotion of early stage metastasis,
particularly invasion and intravasation at the primary tumor site,
has been demonstrated using in vivo models (21). Consistent with these experimental
findings, clinicopathological studies also demonstrated that
overexpression of CD151 is associated with poor prognosis, and
there is significant correlation between overexpression of CD151
and lymph node metastasis of oesophageal squamous cell carcinoma
(22), breast cancer (23), and pancreatic ductal adenocarcinoma
(24).
In this study, silencing of CD151 impaired the
Matrigel invasion activity of HS-MMhigh cells through
downregulation of activated MMP-9. MMP-9 is a well-recognized key
enzyme that functions in the proteolytic degradation of the
extracellular matrix during tumor invasion (25). Homophilic interactions of CD151
upregulate MMP-9 expression in human melanoma cells. Anti-CD151
antibody may impair the homophilic interactions of CD151, resulting
in decreased expression of MMP-9 and abrogation of the initiation
of lymphatic vessel invasion.
Additionally, the present study demonstrated that
downregulation of CD151 affected the phosphorylation status of
SMAD3. Phosphorylation, followed by the nuclear import of SMAD3, is
a key signal for the induction of EMT (26,27).
This process not only facilitates tumor invasion, but also
contributes to tumor progression via pleiotropic pathobiological
mechanisms such as the acquisition of cancer stem cell features
(28).
Our results suggest that anti-CD151 antibody
treatment may impair activation of MMP-9, thus decreasing the
lymphatic invasion activity of HS-MMhigh cells. It is
also likely that reduced levels of phosphorylated SMAD3 may
contribute to reduced lymphatic invasion by abrogating EMT
activity.
As CD151 possesses a tetraspanin with a long
extracellular protein domain, CD151 may be an ideal candidate for
use in antibody-based target therapies to regulate CCS
metastasis.
Numerous monoclonal antibodies have been
successfully generated to regulate various malignant tumors;
however, applications for monoclonal antibodies in the context of
soft tissue sarcoma regulation remain limited. This is surprising,
given that breast cancer, lymphoma, and gastric cancer outcomes can
all be significantly improved by the use of newly developed
antibody-based therapies. Recently, a monoclonal antibody called
Olaratumab that targets platelet-derived growth factor receptor
(PDGFR)-α (34), when used in
combination with doxorubicin received accelerated FDA approval
based on improvement of overall survival compared with doxorubicin
monotherapy in a phase 1b/2 trial.
The monoclonal antibody 50-6, which is used in the
present study, was initially generated by subtractive immunization,
and this antibody inhibited the metastasis of a human epidermoid
carcinoma cell, HEp-3, in a chicken embryo assay (14). Another monoclonal antibody, SFA1.2B4,
was also found to suppress the in vivo pulmonary metastasis
of the CD151-overexpressing human colon cancer cell line RPMI4788
induced by direct intravenous challenge to BALB/c nu/nu mice
(35). These findings may indicate
that anti-CD151 antibodies may be of clinical benefit to patients
not only suffering from CSS, a rare soft part tumor, but also those
diagnosed with other malignant tumors.
We believe that anti-CD151 antibody treatment may
impair activation of MMP-9 to decrease the lymphatic invasion
activity of HS-MMhigh cells. Additionally, reduced
amounts of phosphorylated SMAD3 may also inhibit lymphatic invasion
by abrogating EMT activity. Currently, experiments to delineate the
mechanisms by which anti-CD151 antibody treatment can abrogate
lymphatic invasion and metastasis are underway.
In conclusion, the present findings indicate that
CD151 may provide a therapeutic target to regulate CCS metastasis.
Antibody-based therapy combined with surgical resection could
provide an effective treatment approach for patients diagnosed with
CCS.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the Ministry
of Education of Japan (grant nos. KAKEN 15K08361 and 15K19051).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
CS and TT participated in the design of the study,
the data interpretation and manuscript drafting. KK, YK, YH and YE
performed the experiments. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The experimental protocol was approved by the Animal
Care Committee of Gifu Graduate School of Gifu, Japan (approval no.
H30-32).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Romanska HM and Berditchevski F:
Tetraspanins in human epithelial malignancies. J Pathol. 223:4–14.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zöller M: Tetraspanins: Push and pull in
suppressing and promoting metastasis. Nat Rev Cancer. 9:40–55.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yu Y, Liang C, Wang S, Zhu J, Miao C, Hua
Y, Bao M, Cao Q, Qin C, Shao P and Wang Z: CD151 promotes cell
metastasis via activating TGF-β1/Smad signaling in renal cell
carcinoma. Oncotarget. 9:13313–13323. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang Z, Wang C, Zhou Z, Sun M, Zhou C,
Chen J, Yin F, Wang H, Lin B, Zuo D, et al: CD151-mediated adhesion
is crucial to osteosarcoma pulmonary metastasis. Oncotarget.
7:60623–60638. 2016.PubMed/NCBI
|
|
5
|
Ke AW, Zhang PF, Shen YH, Gao PT, Dong ZR,
Zhang C, Cai JB, Huang XY, Wu C, Zhang L, et al: Generation and
characterization of a tetraspanin CD151/integrin α6β1-binding
domain competitively binding monoclonal antibody for inhibition of
tumor progression in HCC. Oncotarget. 7:6314–6322. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Palmer TD, Martínez CH, Vasquez C, Hebron
KE, Jones-Paris C, Arnold SA, Chan SM, Chalasani V, Gomez-Lemus JA,
Williams AK, et al: Integrin-free tetraspanin CD151 can inhibit
tumor cell motility upon clustering and is a clinical indicator of
prostate cancer progression. Cancer Res. 74:173–187. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zeng P, Wang YH, Si M, Gu JH, Li P, Lu PH
and Chen MB: Tetraspanin CD151 as an emerging potential poor
prognostic factor across solid tumors: A systematic review and
meta-analysis. Oncotarget. 8:5592–5602. 2017.PubMed/NCBI
|
|
8
|
Enzinger FM: Clear-cell sarcoma of tendons
and aponeuroses. An analysis of 21 cases. Cancer. 18:1163–1174.
1965. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chung EB and Enzinger FM: Malignant
melanoma of soft parts. A reassessment of clear cell sarcoma. Am J
Surg Pathol. 7:405–413. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Andreou D and Tunn PU: Sentinel node
biopsy in soft tissue sarcoma. Recent Results Cancer Res.
179:25–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Andreou D, Boldt H, Werner M, Hamann C,
Pink D and Tunn PU: Sentinel node biopsy in soft tissue sarcoma
subtypes with a high propensity for regional lymphatic
spread-results of a large prospective trial. Ann Oncol.
24:1400–1405. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mavrogenis A, Bianchi G, Stavropoulos N,
Papagelopoulos P and Ruggieri P: Clinicopathological features,
diagnosis and treatment of clear cell sarcoma/melanoma of soft
parts. Hippokratia. 17:298–302. 2013.PubMed/NCBI
|
|
13
|
Egawa Y, Saigo C, Kito Y, Moriki T and
Takeuchi T: Therapeutic potential of CPI-613 for targeting tumorous
mitochondrial energy metabolism and inhibiting autophagy in clear
cell sarcoma. PLoS One. 13:e01989402018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Testa JE, Brooks PC, Lin JM and Quigley
JP: Eukaryotic expression cloning with an antimetastatic monoclonal
antibody identifies a tetraspanin (PETA-3/CD151) as an effector of
human tumor cell migration and metastasis. Cancer Res.
59:3812–3820. 1999.PubMed/NCBI
|
|
15
|
Sonobe H, Furihata M, Iwata J, Ohtsuki Y,
Mizobuchi H, Yamamoto H and Kumano O: Establishment and
characterization of a new human clear-cell sarcoma cell-line,
HS-MM. J Pathol. 169:317–322. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sonobe H, Takeuchi T, Taguchi T, Shimizu
K, Iwata J, Furihata M and Ohtsuki Y: Further characterization of
the human clear cell sarcoma cell line HS-MM demonstrating a
specific t(12;22)(q13;q12) translocation and hybrid EWSR1/ATF-1
transcript. J Pahol. 187:594–597. 1999.
|
|
17
|
Takeuchi T, Kuro-o M, Miyazawa H, Ohtsuki
Y and Yamamoto H: Transgenic expression of a novel thymic
epithelial cell antigen stimulates abberant development of
thymocytes. J Immunol. 159:726–733. 1997.PubMed/NCBI
|
|
18
|
Takeuchi T, Adachi Y, Sonobe H, Furihata M
and Ohtsuki Y: A ubiquitin ligase, skeletrophin, is a negative
regulator of melanoma invasion. Oncogene. 25:7059–7069. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Towbin H, Staehelin T and Gordon J:
Electrophoretic transfer of proteins from polyacrylamide gels to
nitrocellulose sheets: Procedure and some applications. Proc Natl
Acad Sci USA. 76:4350–4354. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kito Y, Saigo C and Takeuchi T: Novel
transgenic mouse model of polycystic kidney disease. Am J Pathol.
187:1916–1922. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Morikawa A, Takeuchi T, Kito Y, Saigo C,
Sakuratani T, Futamura M and Yoshida K: Expression of beclin-1 in
the microenvironment of invasive ductal carcinoma of the breast:
Correlation with prognosis and the cancer-stromal interaction. PLoS
One. 10:e01257622015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takeuchi T, Adachi Y and Nagayama T: A
WWOX-binding molecule, transmembrane protein 207, is related to the
invasiveness of gastric signet-ring cell carcinoma. Carcinogenesis.
33:548–554. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takeuchi T, Misaki A, Liang SB, Tachibana
A, Hayashi N and Sonobe H: Expression of T-cadherin (CDH13,
H-cadherin) in human brain and its characteristics as a negative
growth regulator of epidermal growth factor in neuroblastoma cells.
J Neurochem. 74:1489–1497. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Heussen C and Dowdle EB: Electrophoretic
analysis of plasminogen activators in polyacrylamide gels
containing sodium dodecyl sulfate and copolymerized substrates.
Anal Biochem. 102:196–202. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hong IK, Jin YJ, Byun HJ, Jeoung DI, Kim
YM and Lee H: Homophilic interactions of Tetraspanin CD151
up-regulate motility and matrix metalloproteinase-9 expression of
human melanoma cells through adhesion-dependent c-Jun activation
signaling pathways. J Biol Chem. 281:24279–24292. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Z, Wang F, Li Q, Zhang H, Cui Y, Ma
C, Zhu J, Gu X and Sun Z: CD151 knockdown inhibits osteosarcoma
metastasis through the GSK-3β/β-catenin/MMP9 pathway. Oncol Rep.
35:1764–1770. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi GM, Ke AW, Zhou J, Wang XY, Xu Y, Ding
ZB, Devbhandari RP, Huang XY, Qiu SJ, Shi YH, et al: CD151
modulates expression of matrix metalloproteinase 9 and promotes
neoangiogenesis and progression of hepatocellular carcinoma.
Hepatology. 52:183–196. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li P, Zeng H, Qin J, Zou Y, Peng D, Zuo H
and Liu Z: Effects of tetraspanin CD151 inhibition on A549 human
lung adenocarcinoma cells. Mol Med Rep. 11:1258–1265. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mii Y, Miyauchi Y, Hohnoki K, Maruyama H,
Tsutsumi M, Dohmae K, Tamai S, Konishi Y and Yamanouchi T: Neural
crest origin of clear cell sarcoma of tendons and aponeuroses.
Ultrastructural and enzyme cytochemical study of human and nude
mouse-transplanted tumours. Virchows Arch A Pathol Anat
Histopathol. 415:51–60. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yamada K, Ohno T, Aoki H, Semi K, Watanabe
A, Moritake H, Shiozawa S, Kunisada T, Kobayashi Y, Toguchida J, et
al: EWS/ATF1 expression induces sarcomas from neural crest-derived
cells in mice. J Clin Invest. 123:600–610. 2013.PubMed/NCBI
|
|
32
|
Zucman J, Delattre O, Desmaze C, Epstein
AL, Stenman G, Speleman F, Fletchers CD, Aurias A and Thomas G: EWS
and ATF-1 gene fusion induced by t(12;22) translocation in
malignant melanoma of soft parts. Nat Genet. 4:341–345. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Davis IJ, Kim JJ, Ozsolak F, Widlund HR,
Rozenblatt-Rosen O, Granter SR, Du J, Fletcher JA, Denny CT,
Lessnick SL, et al: Oncogenic MITF dysregulation in clear cell
sarcoma: Defining the MiT family of human cancers. Cancer Cell.
9:473–484. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Antoniou G, Lee ATJ, Huang PH and Jones
RL: Olaratumab in soft tissue sarcoma-Current status and future
perspectives. Eur J Cancer. 92:33–39. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kohno M, Hasegawa H, Miyake M, Yamamoto T
and Fujita S: CD151 enhances cell motility and metastasis of cancer
cells in the presence of focal adhesion kinase. Int J Cancer.
97:336–343. 2002. View Article : Google Scholar : PubMed/NCBI
|