Introduction
Gastric carcinoma is currently the third most common
cause of cancer mortality worldwide; 50% of all gastric carcinomas
occur in Eastern Asia, and it is particularly common in China
(1,2). At present, the effective treatment
strategies for gastric carcinoma include surgery, radiotherapy,
chemotherapy and targeted therapy (3,4).
Surgical resection followed by adjuvant chemotherapy remains the
most effective therapeutic option. Unfortunately, relapse and
metastasis of gastric tumors (5),
and resistance to chemotherapy are common. Therefore, novel
therapeutic agents for gastric carcinoma therapy are urgently
required.
Increasing attention has been paid to the
application of natural products for chemopreventative cancer
therapy. Terpenoids are phytochemicals traditionally used for
medicinal purposes. Preclinical studies have demonstrated that
terpenoids present in plant foods or isolated from medicinal
plants, including germacrone from Rhizoma Curcuma or
lactucopicrin from Cichorium intybus L, exhibit significant
anti-cancer effects against various types of cancer cell in
vivo and in vitro (6–8).
Alantolactone (ALT), a sesquiterpene lactone compound isolated from
Inula helenium, has exhibited multiple biological
properties, including anti-bacterial, anti-inflammatory and
anti-cancer activities (9). Notably,
ALT was suggested to exhibit potential anti-cancer activity against
various types of cancer, including human colorectal cancer
(10), liver cancer (11,12),
leukemia (13), breast cancer
(14), lung cancer (15,16) and
cervical cancer (17,18). ALT may inhibit breast cancer growth
via anti-angiogenic activity by inhibiting vascular endothelial
growth factor receptor 2 and RAC-alpha serine/threonine-protein
kinase B (AKT) signaling (19). In
human cervical cancer cells, ALT induces apoptosis via generation
of reactive oxygen species (ROS) and inhibition of the B-cell
lymphoma 2 (Bcl-2)/Bcl-2 associated X apoptosis regulator (Bax)
signaling pathway (17). In SK-MES-1
lung squamous cancer SK-MES-1 cells, ALT may trigger apoptosis and
induce cell cycle G1/G0 phase arrest. Furthermore, ALT may enhance
the chemosensitivity of A549 cells to doxorubicin via ROS-mediated
apoptosis (20). However, the exact
mechanism underlying the anti-cancer activity of ALT in human
gastric cancer cells remains to be elucidated.
The present study aimed to elucidate the anti-cancer
effects and associated molecular mechanisms of ALT in BGC-823
cells, and to evaluate the potential of ALT for its application as
a novel naturally-derived agent for the treatment of gastric
cancer.
Materials and methods
Reagents
ALT, N-acetyl cysteine (NAC) and MTT were purchased
from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). Primary
antibodies to detect cyclin D1 (cat. no., 2978), cyclin-dependent
kinase inhibitor 1 (p21; cat. no., 2947), cyclin-dependent kinase
inhibitor 1B (p27; cat. no., 3686), Bax (cat. no., 5023), Bcl-2
(cat. no., 15071), poly (adenosine 5′diphosphate-ribose) polymerase
(PARP; cat. no., 5625), phosphorylated (p)-AKT (cat. no., 4060),
AKT (cat. no., 9272) and GAPDH (cat. no., 5174) were obtained from
Cell Signaling Technology, Inc. (Danvers, MA, USA). The Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis
detection kit and cell cycle detection kit were purchased from
Nanjing KeyGen Biotech Co., Ltd. (Nanjing, China). The ROS assay
kit was purchased from Nanjing Jiancheng Bioengineering Institute
(Nanjing, China). RPMI-1640 medium and fetal bovine serum (FBS)
were purchased from Gibco; Thermo Fisher Scientific, Inc. (Waltham,
MA, USA).
Cell culture
The BGC-823 human gastric cancer cell line was
purchased from The Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China). The cells were
cultured in RPMI-1640 medium supplemented with 10% FBS, 100 U/ml
penicillin and 100 µg/ml streptomycin at 37°C with 5%
CO2.
MTT cytotoxicity assay
Cell viability was measured using an MTT assay as
described previously (21). The
BGC-823 cells were seeded into 96-well plates at a density of
1×105 cells/well and treated following culture for 24 h.
The BGC-823 cells were treated with ALT (0, 10, 20, 40, 60, 80 or
100 µM), and incubated for an additional 24 h. In total, 0.5 mg/ml
MTT solution was then added to the medium, and the cells were
incubated for an additional 4 h at 37°C. The culture was then
removed, and dimethyl sulfoxide (150 µl/well) was added to dissolve
the solid residue. The absorbance at 490 nm (A490) was determined
using an ELISA microplate reader, and all experiments were
performed at least 3 times. Percentage of cell viability was
calculated as follows: Cell viability (%)=(A490 sample A490
blank)/(A490 control-A490 blank) ×100.
Cell cycle assay
Cell cycle arrest was detected using a cell cycle
detection kit according to manufacturer's protocol (Nanjing KeyGen
Biotech Co., Ltd.). Briefly, the BGC-823 cells were seeded at
1×106 cells/well in 6-well plates and treated with
different ALT concentrations (0, 10, 20, 40 or 60 µM) for 24 h. The
BGC-823 cells were fixed with 75% ethanol at 4°C overnight. PI was
then used to stain the DNA of the samples for 15 min at 25°C in the
dark, and flow cytometry analysis was performed using a FACSCalibur
flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA) was used
to determine the cell cycle and analyzed by Modfit LT 3.0 software
(Verity Software House, Topsham, ME, USA). All experiments were
performed at least three times.
Assessment of apoptosis
Cell apoptosis was assessed using an Annexin
V-FITC/PI kit according to manufacturer's protocol (Nanjing KeyGen
Biotech Co., Ltd.) and the apoptotic rate was analyzed by flow
cytometry on a FACSCalibur™ flow cytometer (BD Biosciences).
BGC-823 cells were seeded at 1×106 cells/well in 6-well
plates and treated different ALT concentrations (0, 10, 20, 40 or
60 µM) for 24 h. Subsequently, cells were stained with Annexin
V-FITC and PI in binding buffer for 15 min at 25°C in the dark.
According to manufacturer's protocol, 5 µl Annexin V-FITC and 5 µl
PI were added per sample. The apoptotic cells were then detected on
a FACSCalibur™ flow cytometer (BD Biosciences) and analyzed using
FlowJo 7.6 software (FlowJo LLC, Ashland, OR, USA). A total of
three independent experiments were performed.
ROS detection
ROS generation was measured using a ROS assay kit
according to the manufacturer's protocol (Nanjing Jiancheng
Bioengineering Institute) and the ROS levels were analyzed by flow
cytometry on a FACSCalibur™ flow cytometer (BD Biosciences).
Briefly, BGC-823 cells were treated with the indicated
concentrations (0, 10, 20, 40 and 60 µM) of ALT for 6 h. In
addition, BGC-823 cells were pre-treated with 10 mM NAC for 1 h,
and with 40 µM ALT for 6 h. The cells were then incubated with 30
µM 2′,7′-dichlorodihydrofluorescein diacetate at 37°C for 30 min.
The cells were analyzed using a FACSCalibur flow cytometer (BD
Biosciences). The results were analyzed with FlowJo 7.6 software
(FlowJo LLC) and all experiments were performed at least three
times.
Western blot analysis
Cells were lysed using radioimmunoprecipitation
assay (Beyotime Institute of Biotechnology, Haimen, China) on ice
for 30 min. The proteins were quantified with Bicinchoninic Acid
Protein Assay kit (Thermo Fisher Scientific, Inc.). Proteins (40
µg) were separated by 6–12% SDS-PAGE and then transferred onto
nitrocellulose membranes (Pall Life Sciences, Port Washington, NY,
USA) and blocked with 5% non-fat milk in TBS-containing 0.05% Tween
20 for 2 h at room temperature. The membranes were incubated with
anti-cyclin D1 (1:1,000 dilution), anti-p21 (1:1,000 dilution),
anti-p27 (1:1,000 dilution), anti-Bax (1:1,000 dilution),
anti-Bcl-2 (1:1,000 dilution), anti-p-AKT (1:1,000 dilution),
anti-AKT (1:1,000 dilution), anti-cleaved PARP and anti-GAPDH
(1:3,000 dilution) antibodies at 4°C overnight. Membranes were then
incubated with horseradish peroxidase (HRP)-conjugated anti-mouse
immunoglobulin (Ig)G (cat. no. 7076; 1:25,000) and HRP-conjugated
anti-rabbit IgG (cat. no. 7074; 1:20,000; both Cell Signaling
Technology, Inc.) secondary antibodies for 2 h at 25°C.
Visualization was performed using a SuperSignal West Pico
chemiluminescent substrate (Pierce; Thermo Fisher Scientific,
Inc.).
Statistical analysis
Data are presented as the mean ± standard deviation.
Differences between groups were determined by one-way analysis of
variance followed by Dunnett's or Tukey's post-hoc tests. The
analyses were performed using SPSS v.19 statistical software (IBM
Corp., Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
ALT inhibits cell proliferation in
BGC-823 cells
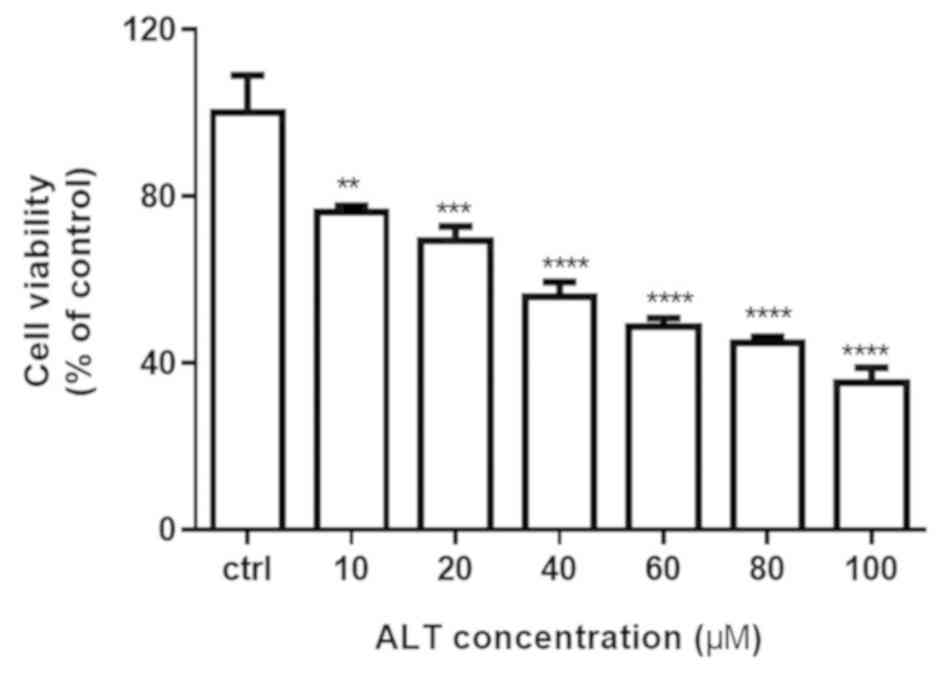
The effect of ALT on cell proliferation was
determined using an MTT assay. The cytotoxicity of ALT was
evaluated by treating BGC-823 cells with different concentrations
(0, 10, 20, 40, 60, 80 and 100 µM) for 24 h. As demonstrated in
Fig. 1, ALT significantly inhibited
the viability of BGC-823 cells in a concentration-dependent manner,
exhibiting significant differences compared with the control group.
These results indicated that ALT significantly suppressed the
growth of BGC-823 human gastric cancer cells.
ALT induces cell cycle arrest at the
G0/G1 phase in BGC-823 cells
Inhibition of cell proliferation via anti-tumor
drugs is often accompanied by changes in cell cycle progression
(22,23). In the present study, flow cytometry
was used to analyze the distribution of the cell cycle in BGC-823
cells following treatment with ALT (20–60 µM) for 24 h. G0/G1 phase
arrest was observed in BGC-823 cells exposed to ALT compared with
the control group. As demonstrated in Fig. 2A, ALT induced G0/G1 phase arrest of
BGC-823 cells in a dose-dependent manner. The percentage of cells
in the G0/G1 phase was 57.50±0.10, 66.57.50±0.05, 71.24±0.88,
72.31±0.17 and 81.97±0.53% in the 0, 10, 20, 40 and 60 µM groups,
respectively (Fig. 2A). To gain
insight into the mechanism of G0/G1 phase arrest, the expression
levels of cyclin D1, p21 and p27, which regulate G1 to S phase
progression, were detected. As presented in Fig. 2B, the level of cyclin D1 was
decreased, and levels of p21 and p27 were significantly increased
following stimulation with ALT. These results indicated that ALT
induces G0/G1 phase arrest via downregulation of cyclin D1
expression and upregulation of p21 and p27 expression in BGC-823
cells.
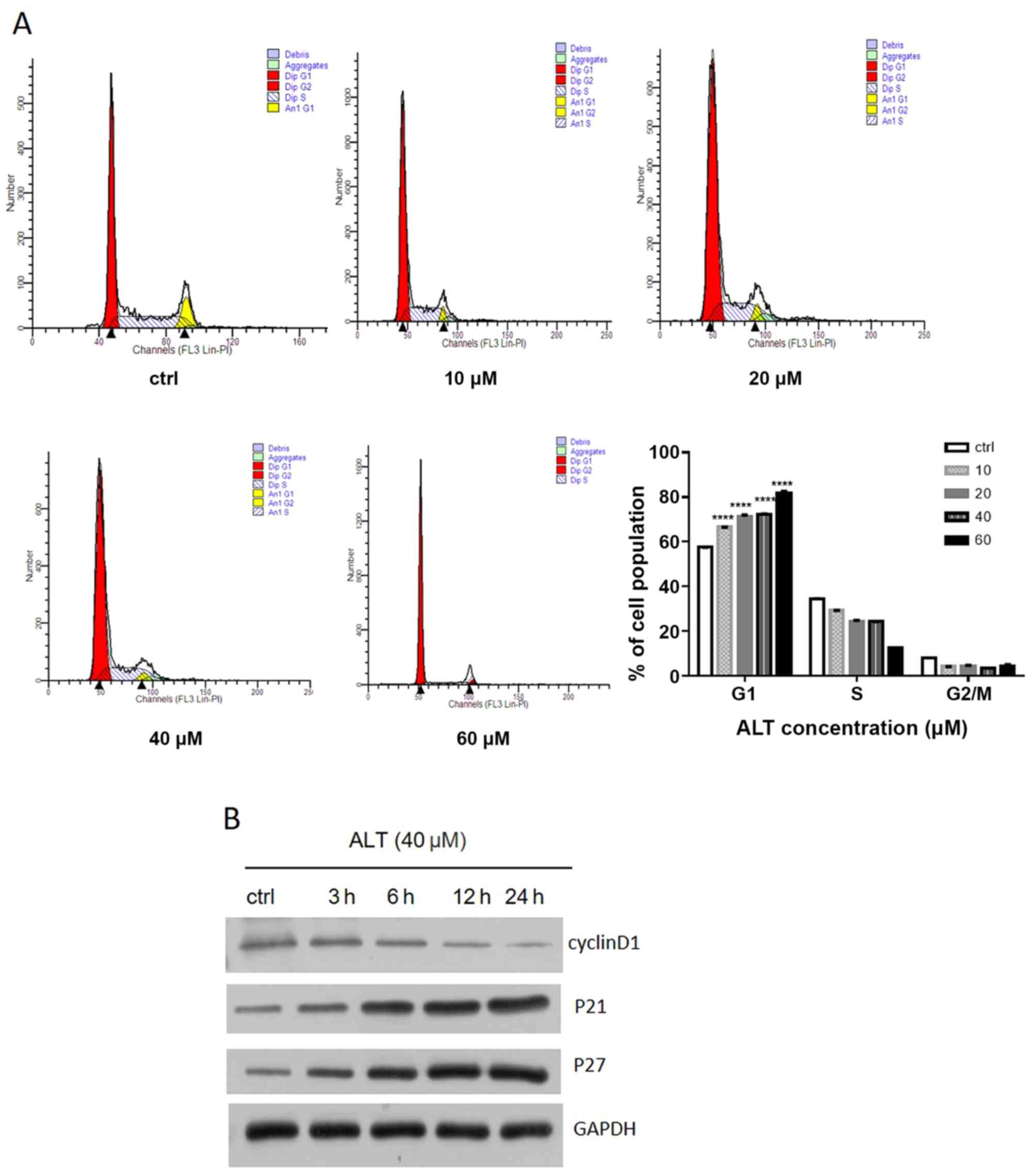 | Figure 2.Effects of ALT on cell cycle
distribution and cell cycle-associated proteins in BGC-823 cells.
(A) BGC-823 cells were incubated with different concentrations of
ALT (0–60 µM) for 24 h. The percentages of cells in G0/G1, S, and
G2/M phases were determined by flow cytometry. All data are
expressed as means ± standard deviation from 3 independent
experiments. (B) BGC-823 cells were treated with ALT (40 µM) for 3,
6, 12 and 24 h. Western blot analysis was used to detect the
expression levels of cyclin D1, p21 and p27. GAPDH was used as an
internal control. ****P<0.0001 vs. the control group. ALT, ALT,
alantolactone; ctrl, control; p21, cyclin-dependent protein kinase
1; p27, cyclin-dependent protein kinase 1B. |
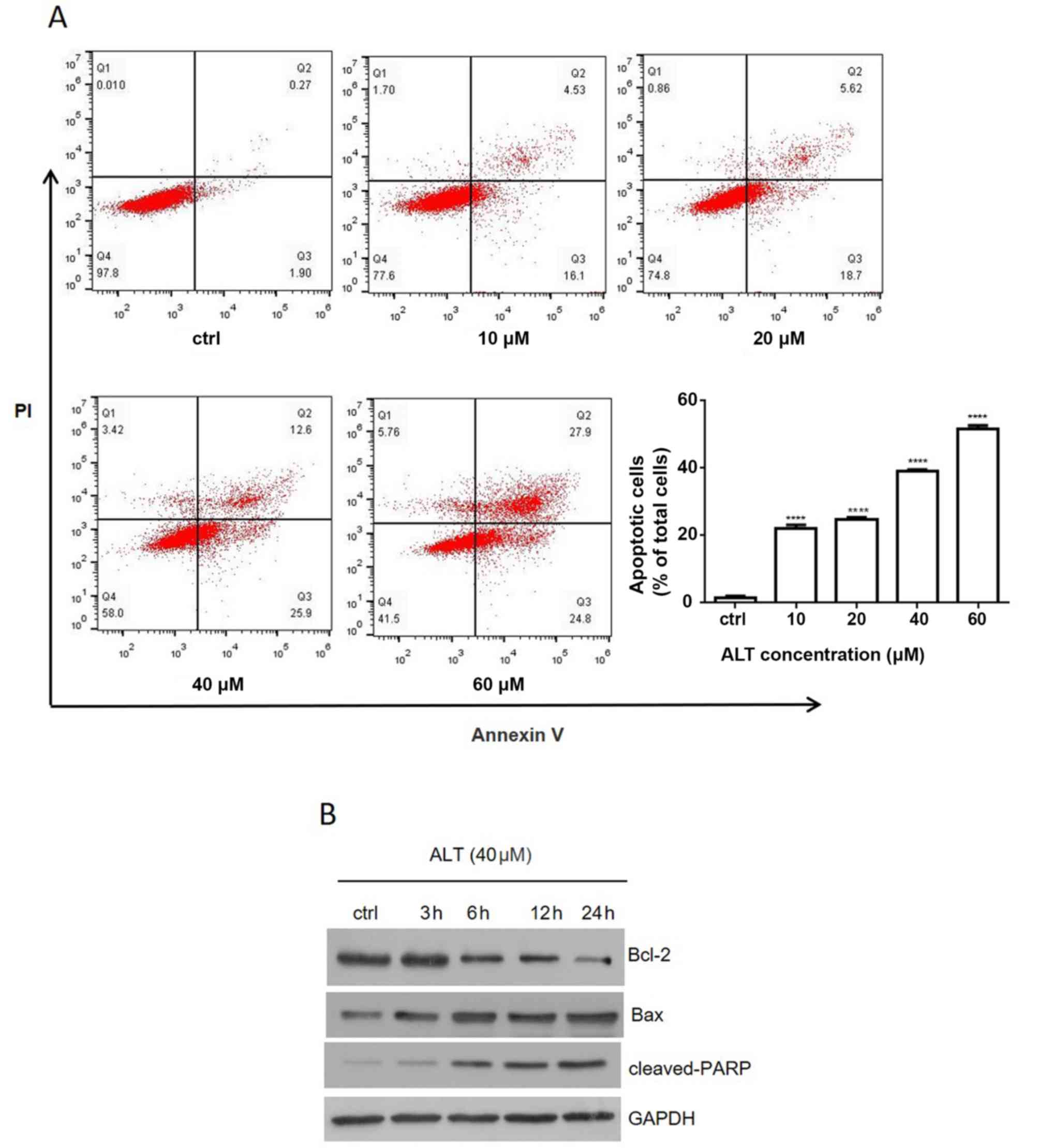
ALT induces apoptosis in BGC-823
cells
Anti-tumor drugs often execute cytotoxic effects via
the induction of apoptosis (24,25). The
effect of ALT on BGC-823 cell apoptosis was evaluated using Annexin
V-FITC/PI double staining and flow cytometry. The percentage of
apoptotic cells was 1.15±0.01, 22.69±0.13, 25.10±0.14, 39.53±0.37
and 50.93±0.29% in the 0, 10, 2, 40 and 60 µM groups, respectively,
after 24 h (Fig. 3A). The data
suggested that ALT significantly induced apoptosis in a
dose-dependent manner. To reveal the molecular mechanism involved
in ALT-induced apoptosis, the levels of anti-apoptosis protein
Bcl-2 and pro-apoptosis proteins Bax and cleaved PARP were detected
by western blot analysis. When BGC-823 cells were treated with 40
µM ALT, Bax and cleaved PARP protein levels were increased in a
time-dependent manner, whereas Bcl-2 levels were decreased in a
time-dependent manner (Fig. 3B).
These results suggested that ALT may trigger apoptosis in BGC-823
cells partially via a mitochondrial-dependent pathway.
ROS accumulation is associated with
ALT-induced apoptosis
Several studies have revealed that anti-tumor
drug-induced cell death is associated with intracellular ROS
accumulation in multiple cancer types (26–28).
Therefore, whether ROS were involved in ALT-induced apoptosis was
also investigated. Intracellular ROS generation was detected
following treatment with different ALT concentrations (0, 10, 20,
40 or 60 µM) for 6 h. ALT significantly increased intracellular ROS
generation in a concentration-dependent manner in BGC-823 cells
(data not shown). Subsequently, to determine whether the increased
ROS generation served an important role in ALT-induced cell
apoptosis, the cells were pretreated with NAC 1 h prior to
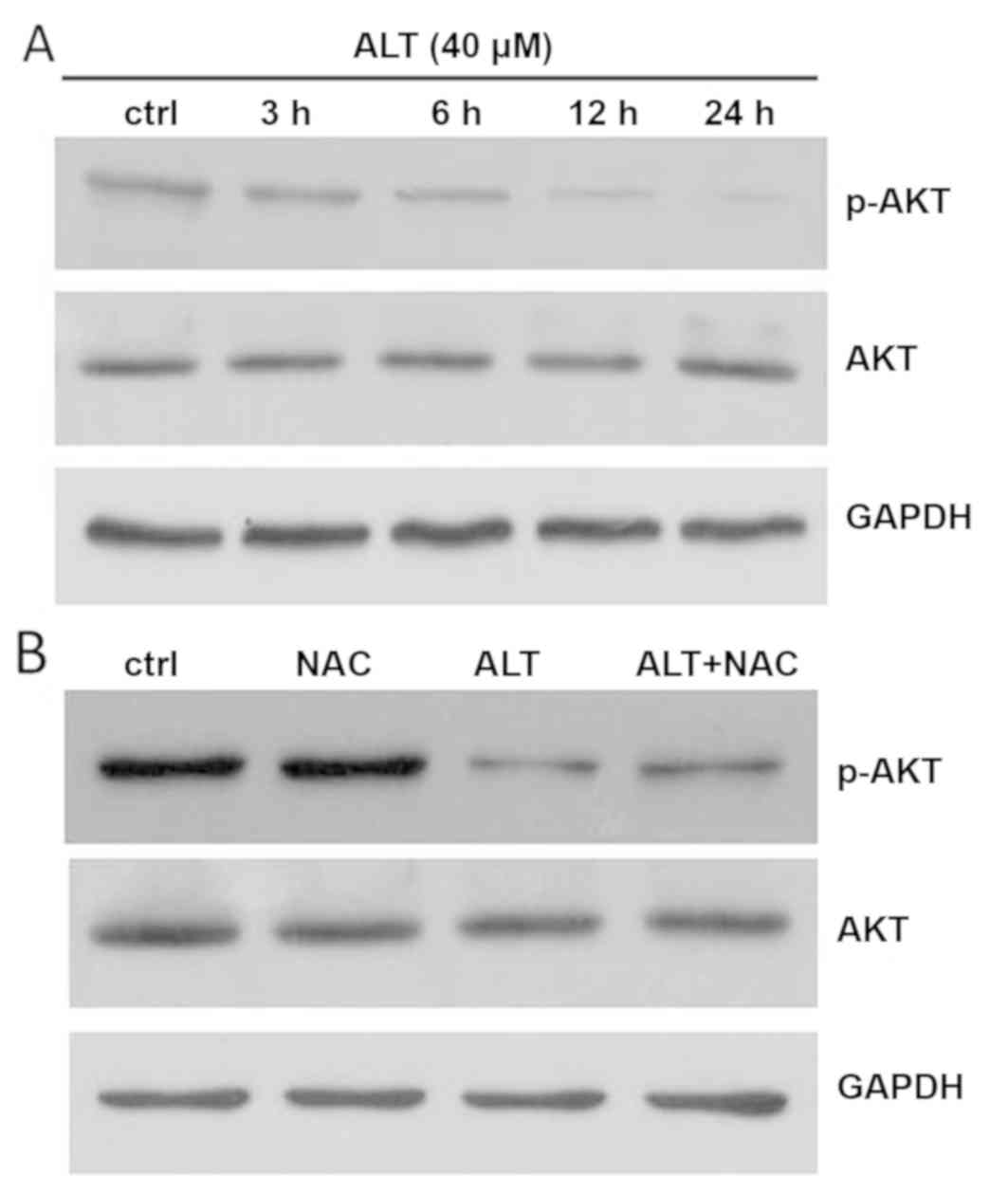
treatment with ALT for 24 h. Pretreatment with NAC effectively
prevented ALT-induced ROS accumulation (Fig. 4A). Additionally, NAC also abolished
ALT-inhibited cell proliferation and ALT-induced cell apoptosis
(Fig. 4B and C). Furthermore,
western blot analysis revealed that NAC decreased the expression of
Bax and the cleavage of PARP, and increased Bcl-2 expression
(Fig. 4D). Collectively, the results
indicated that ALT-induced apoptosis was associated with ROS
accumulation.
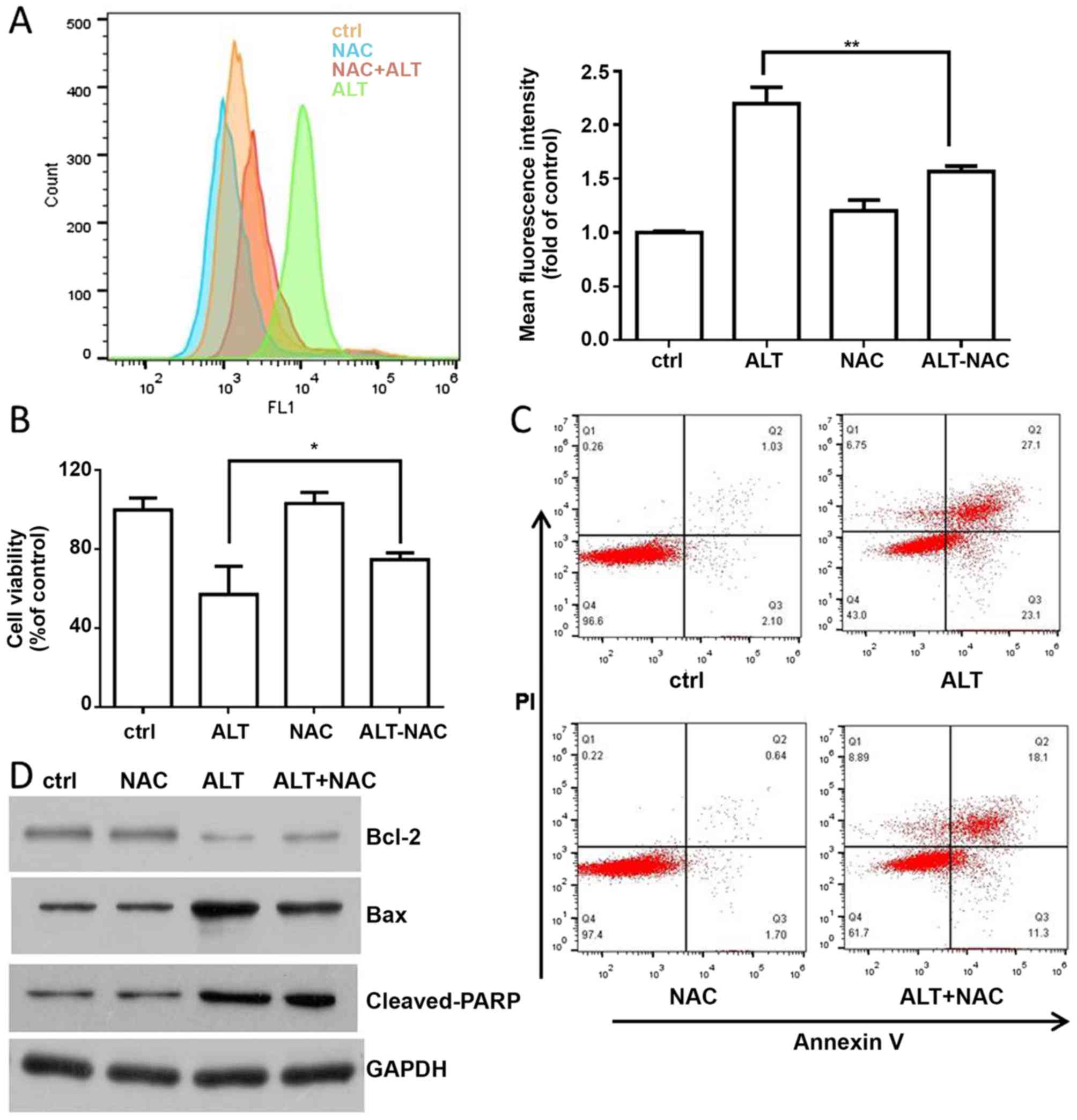 | Figure 4.ROS are involved in ALT-induced
apoptosis. (A) BGC-823 cells were pre-treated with NAC (10 mM) for
1 h, and then ALT (40 µM) was added to the cells for 6 h. Flow
cytometry was used to detect ROS levels. (B) BGC-823 cells were
pre-treated with NAC (10 mM) for 1 h, and then ALT (40 µM) was
added to the cells for 24 h. MTT was used to measure cell
viability. (C) BGC-823 cells were incubated with ALT and NAC as
aforementioned. Flow cytometry was used to detect apoptosis in
BGC-823 cells. (D) BGC-823 cells were pre-treated with NAC (10 mM)
for 1 h, and then ALT (40 µM) was added to the cells for 12 h.
Western blot analysis was used to detect the expression levels of
Bax, Bcl-2 and cleaved PARP. *P<0.05 and **P<0.01 vs. the
control group. ROS, reactive oxygen species; ALT, alantolactone;
NAC, N-acetyl cysteine; Bcl-2, B cell lymphoma 2; Bax,
Bcl-2-associated protein; PARP, poly (adenosine
5′diphosphate-ribose) polymerase; PI, propidium iodide; ctrl,
control. |
ALT induces BGC-823 cell apoptosis via
inhibition of AKT signaling
Accumulating evidence has revealed that the AKT
pathway is one of the major signaling pathways closely associated
with cancer progression (29,30). To
determine whether the AKT signaling pathway is involved in
ALT-induced apoptosis, western blot analysis was used to assay the
effects of ALT on p-AKT and AKT protein expression. As presented in
Fig. 5A, BGC-823 cells were treated
with 40 µM ALT for 3, 6, 12 and 24 h. The phosphorylation of AKT
was significantly decreased in a time-dependent manner. ROS have
been demonstrated to be associated with the apoptosis induced by
anti-cancer drugs via regulation of AKT pathways (31). To additionally investigate the
association between ROS and the AKT pathway, cells were treated
with or without NAC for 1 h, and then western blot analysis was
used to examine p-AKT and AKT protein expression levels in BGC-823
cells. As presented in Fig. 5B, NAC
significantly increased AKT phosphorylation. Together, these
results suggest that ALT induced apoptosis through ROS-mediated
inactivation of the AKT signaling pathway in BGC-823 cells.
Discussion
The emergence of drug resistance remains a major
barrier for successful cancer treatments. Therefore, exploring and
developing novel and effective anti-cancer drugs is essential. ALT,
a sesquiterpene lactone, is a potential anti-cancer therapeutic
agent. In a previous study, the anti-cancer effects of ALT were
demonstrated in various tumor cells, including breast (14), lung (20,32),
cervical (17,18), liver (12), colorectal (10) and colon cancer (33). However, to the best of our knowledge,
there is a lack of thorough experimental data describing the
anti-tumor effects of ALT in BGC-823 human gastric cancer cells.
The present study demonstrated that ALT exhibited significant
anti-cancer effects in BGC-823 cells: ALT inhibited cell
proliferation, and induced G0/G1 cell cycle arrest and apoptosis in
BGC-823 cells. Furthermore, ALT promoted BGC-823 cell apoptosis via
ROS-induced AKT signaling.
Cell cycle arrest is an important mechanism
associated with anti-cancer drug-induced proliferation inhibition
(34,35). Previous studies have indicated that
ALT may inhibit cell cycle progression in lung and colorectal
cancer cells (10,32). Similarly, flow cytometric analysis
demonstrated that ALT induced G0/G1 cell cycle arrest of BGC-823
cells in a concentration-dependent manner. Furthermore, previous
studies have demonstrated that anti-cancer drugs promote G0-G1/S
transition and inhibit cell proliferation by regulating the
expression of cyclin D1 (36,37).
Additionally, p27 and p21 serve suppressive roles in G0-G1/S
transition by inhibiting the activity of cyclin/cyclin-dependent
kinase complexes (37,38). To additionally investigate the
molecular basis by which ALT inhibited G0/G1 transition in BGC-823
cells, the expression of cell cycle-associated proteins was
determined by western blot analysis. ALT markedly decreased the
expression levels of cyclin D1, whereas the expression of p21 and
p27 were increased. These data provide evidence that ALT inhibited
the proliferation of BGC-823 cells via ALT-induced G0/G1 phase
arrest.
Apoptosis is a critical homeostatic mechanism
involved in anti-cancer drug-induced proliferation inhibition
(35). The mitochondrial apoptotic
pathway has a critical role in drug-mediated apoptosis and is often
regarded as a potential anti-cancer target (39–41).
Bcl-2 family protein members, including Bax and Bcl-2, serve
important roles in the mitochondrial apoptotic pathway. Previous
data has indicated that ALT induces apoptosis in a variety of human
cancer cells: Cui et al (14)
suggested that ALT induced the mitochondrial-mediated apoptotic
pathway by increasing the Bax/Bcl-2 ratio and PARP cleavage in
MDA-MB-231 cells; Jiang et al (17) also demonstrated that the Bcl-2/Bax
signaling pathway was associated with ALT-induced HeLa cell
apoptosis. In the present study, treatment with ATL significantly
induced apoptosis of BGC-823 cells by increasing the Bax/Bcl-2
ratio and PARP cleavage in BGC-823 cells. These data suggest that
the mitochondrial apoptotic pathway serves a key role in
ALT-mediated BGC-823 cell apoptosis.
The AKT signaling pathway is an important
anti-apoptosis pathway that promotes cell survival and resistance
to cell apoptosis induced by chemotherapeutic agents in various
cancer types (42). Inactivation of
the AKT signaling pathway may inhibit cell growth and induce cell
apoptosis in various cancer cells. Therefore, whether the AKT
pathway was associated with ALT-induced apoptosis of BGC-823 cells
was examined in the present study. The data demonstrated that ALT
decreased the phosphorylation of AKT in a time-dependent manner.
This indicated that AKT signaling may be involved in ALT-mediated
apoptosis of BGC-823 cells.
ROS have been suggested to be involved in the
initiation and the promotion of tumor development at different
stages of carcinogenesis (43,44). In
fact, numerous studies have demonstrated that various anti-cancer
drugs exert their effects via ROS-dependent pathways (45–47). ALT
was demonstrated to induce apoptosis of MDA-MB-231 cells via
ROS-mediated mitochondrial dysfunction (14). Jiang et al (17) also revealed that ROS may mediate
apoptosis in human cervical cancer cells by increasing the
Bax/Bcl-2 ratio. The results of the present study demonstrated that
ALT treatment increased ROS generation in a concentration-dependent
manner in BGC-823 cells (data not shown). Furthermore, pretreatment
with NAC for 1 h reversed the ALT-induced production of ROS and
cell apoptosis, and NAC significantly decreased the Bax/Bcl-2 ratio
and PARP cleavage. The results suggested that ALT induced
ROS-dependent apoptosis in BGC-823 cells. In addition, ROS
generation was demonstrated to be involved in chemotherapeutic
agent-mediated apoptosis and may be an upstream regulator of
AKT-mediated signaling pathways (48,49). In
the present study, pretreatment with NAC for 1 h reversed the AKT
inhibition induced by ALT. These results suggested that the
apoptosis of BGC-823 cells was induced by ALT via ROS generation,
which was then modulated the AKT signaling.
In conclusion, the results demonstrated that ALT
induced apoptosis and G0/G1 phase arrest in BGC-823 cells in a
concentration-dependent manner. In addition, ALT induced the
apoptosis of BGC-823 cells via ROS-mediated inactivation of the AKT
signaling pathway; therefore, ALT may be a promising candidate drug
for the treatment of gastric cancer. However, additional studies
are required to validate the anti-cancer activity of ALT in
xenograft mouse models in vivo.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Xinjiang
Uygur Autonomous Region Natural Science Foundation Project (grant
no. 2015211C244).
Availability of data and materials
The datasets used and/or analyzed in the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XZ conducted the experiments, analyzed the data,
contributed to the design of the study and prepared the manuscript.
XZ and HMZ performed the western blotting and analyzed the data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Theuer CP, Kurosaki T, Ziogas A, Butler J
and Anton-Culver H: Asian patients with gastric carcinoma in the
United States exhibit unique clinical features and superior overall
and cancer specific survival rates. Cancer. 89:1883–1892. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lemjabbar-Alaoui H, Hassan OU, Yang YW and
Buchanan P: Lung cancer: Biology and treatment options. Biochim
Biophys Acta. 1856:189–210. 2015.PubMed/NCBI
|
|
4
|
Dikken JL, van de Velde CJ, Coit DG, Shah
MA, Verheij M and Cats A: Treatment of resectable gastric cancer.
Therap Adv Gastroenterol. 5:49–69. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ueno T, Iida M, Yoshino S, Takeda S,
Kubota H, Higashida M, Oka Y, Tsuruta A, Matsumoto H and Nagano H:
East versus west: Differences in surgical management in asia
compared with Europe and North America. Surg Clin North Am.
97:453–466. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kubatka P, Kapinova A, Kruzliak P, Kello
M, Výbohová D, Kajo K, Novák M, Chripková M, Adamkov M, Péč M, et
al: Antineoplastic effects of Chlorella pyrenoidosa in the breast
cancer model. Nutrition. 31:560–569. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pan J, Miao D and Chen L: Germacrone
reverses adriamycin resistance in human chronic myelogenous
leukemia K562/ADM cells by suppressing MDR1 gene/P-glycoprotein
expression. Chem Biol Interact. 288:32–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang X, Lan D, Ning S and Ruan L:
Anticancer action of lactucopicrin in SKMEL-5 human skin cancer
cells is mediated via apoptosis induction, G2/M cell cycle arrest
and downregulation of m=TOR/PI3K/AKT signalling pathway. J BUON.
23:224–228. 2018.PubMed/NCBI
|
|
9
|
Khan M, Yi F, Rasul A, Li T, Wang N, Gao
H, Gao R and Ma T: Alantolactone induces apoptosis in glioblastoma
cells via GSH depletion, ROS generation, and mitochondrial
dysfunction. IUBMB Life. 64:783–794. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ding Y, Wang H, Niu J, Luo M, Gou Y, Miao
L, Zou Z and Cheng Y: Induction of ROS overload by alantolactone
prompts oxidative DNA damage and apoptosis in colorectal cancer
cells. Int J Mol Sci. 17:5582016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lei JC, Yu JQ, Yin Y, Liu YW and Zou GL:
Alantolactone induces activation of apoptosis in human hepatoma
cells. Food Chem Toxicol. 50:3313–3319. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Khan M, Li T, Ahmad Khan MK, Rasul A,
Nawaz F, Sun M, Zheng Y and Ma T: Alantolactone induces apoptosis
in HepG2 cells through GSH depletion, inhibition of STAT3
activation, and mitochondrial dysfunction. Biomed Res Int.
2013:7198582013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang C, Yang J, Sun M, Yan J, Meng X and
Ma T: Alantolactone inhibits growth of K562/adriamycin cells by
downregulating Bcr/Abl and P-glycoprotein expression. IUBMB Life.
65:435–444. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cui L, Bu W, Song J, Feng L, Xu T, Liu D,
Ding W, Wang J, Li C, Ma B, et al: Apoptosis induction by
alantolactone in breast cancer MDA-MB-231 cells through reactive
oxygen species-mediated mitochondrion-dependent pathway. Arch Pharm
Res. 41:299–313. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee SS, Jeong HE, Liu KH, Ryu JY, Moon T,
Yoon CN, Oh SJ, Yun CH and Shin JG: Identification and functional
characterization of novel CYP2J2 variants: G312R variant causes
loss of enzyme catalytic activity. Pharmacogenet Genomics.
15:105–113. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gaedigk A, Baker DW, Totah RA, Gaedigk R,
Pearce RE, Vyhlidal CA, Zeldin DC and Leeder JS: Variability of
CYP2J2 expression in human fetal tissues. J Pharmacol Exp Ther.
319:523–532. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang Y, Xu H and Wang J: Alantolactone
induces apoptosis of human cervical cancer cells via reactive
oxygen species generation, glutathione depletion and inhibition of
the Bcl-2/Bax signaling pathway. Oncol Lett. 11:4203–4207. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang J, Li Y, Duan D, Yao J, Gao K and
Fang J: Inhibition of thioredoxin reductase by alantolactone
prompts oxidative stress-mediated apoptosis of HeLa cells. Biochem
Pharmacol. 102:34–44. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu YR, Cai QY, Gao YG, Luan X, Guan YY,
Lu Q, Sun P, Zhao M and Fang C: Alantolactone, a sesquiterpene
lactone, inhibits breast cancer growth by antiangiogenic activity
via blocking VEGFR2 signaling. Phytother Res. 32:643–650. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Maryam A, Mehmood T, Zhang H, Li Y, Khan M
and Ma T: Alantolactone induces apoptosis, promotes STAT3
glutathionylation and enhances chemosensitivity of A549 lung
adenocarcinoma cells to doxorubicin via oxidative stress. Sci Rep.
7:62422017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu ZY, Wang Z, Lee KY, Yuan P and Ding J:
Effect of silencing colon cancer-associated transcript 2 on the
proliferation, apoptosis and autophagy of gastric cancer BGC-823
cells. Oncol Lett. 15:3127–3132. 2018.PubMed/NCBI
|
|
22
|
Wen W, Lowe G, Roberts CM, Finlay J, Han
ES, Glackin CA and Dellinger TH: Pterostilbene Suppresses ovarian
cancer growth via induction of apoptosis and blockade of cell cycle
progression involving inhibition of the STAT3 pathway. Int J Mol
Sci. 19(pii): E19832018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ling Z, Guan H, You Z, Wang C, Hu L, Zhang
L, Wang Y, Chen S, Xu B and Chen M: Aloperine executes antitumor
effects through the induction of apoptosis and cell cycle arrest in
prostate cancer in vitro and in vivo. Onco Targets Ther.
11:2735–2743. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cho HD, Lee JH, Moon KD, Park KH, Lee MK
and Seo KI: Auriculasin-induced ROS causes prostate cancer cell
death via induction of apoptosis. Food Chem Toxicol. 111:660–669.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lai ZQ, Ip SP, Liao HJ, Lu Z, Xie JH, Su
ZR, Chen YL, Xian YF, Leung PS, Lin ZX and Brucein D: a naturally
occurring tetracyclic triterpene quassinoid, induces apoptosis in
pancreatic cancer through ROS-associated PI3K/Akt signaling
pathway. Front Pharmacol. 8:9362017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xia S, Miao Y and Liu S: Withaferin A
induces apoptosis by ROS-dependent mitochondrial dysfunction in
human colorectal cancer cells. Biochem Biophys Res Commun.
503:2363–2369. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu Y, Tong Y, Ying J, Lei Z, Wan L, Zhu X,
Ye F, Mao P, Wu X, Pan R, et al: Chrysin induces cell growth
arrest, apoptosis, and ER stress and inhibits the activation of
STAT3 through the generation of ROS in bladder cancer cells. Oncol
Lett. 15:9117–9125. 2018.PubMed/NCBI
|
|
28
|
Sun Q, Lu NN and Feng L: Apigetrin
inhibits gastric cancer progression through inducing apoptosis and
regulating ROS-modulated STAT3/JAK2 pathway. Biochem Biophys Res
Commun. 498:164–170. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sathe A and Nawroth R: Targeting the
PI3K/AKT/mTOR Pathway in Bladder Cancer. Methods Mol Biol.
1655:335–350. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Costa RLB, Han HS and Gradishar WJ:
Targeting the PI3K/AKT/mTOR pathway in triple-negative breast
cancer: A review. Breast Cancer Res Treat. 169:397–406. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang B, Zhou TY, Nie CH, Wan DL and Zheng
SS: Bigelovin, a sesquiterpene lactone, suppresses tumor growth
through inducing apoptosis and autophagy via the inhibition of mTOR
pathway regulated by ROS generation in liver cancer. Biochem
Biophys Res Commun. 499:156–163. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao P, Pan Z, Luo Y, Zhang L, Li X, Zhang
G, Zhang Y, Cui R, Sun M and Zhang X: Alantolactone induces
apoptosis and cell cycle arrest on lung squamous cancer SK-MES-1
cells. J Biochem Mol Toxicol. 29:199–206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y, Bao YL, Wu Y, Yu CL, Huang YX,
Sun Y, Zheng LH and Li YX: Alantolactone induces apoptosis in RKO
cells through the generation of reactive oxygen species and the
mitochondrial pathway. Mol Med Rep. 8:967–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
King KL and Cidlowski JA: Cell cycle
regulation and apoptosis. Annu Rev Physiol. 60:601–617. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sithara T, Dhanya BP, Arun KB, Sini S, Dan
M, Kokkuvayil Vasu R and Nisha P: Zerumbone, a cyclic sesquiterpene
from Zingiber zerumbet induces apoptosis, cell cycle arrest, and
antimigratory effects in SW480 colorectal cancer cells. J Agric
Food Chem. 66:602–612. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang J, Li XM, Bai Z, Chi BX, Wei Y and
Chen X: Curcumol induces cell cycle arrest in colon cancer cells
via reactive oxygen species and Akt/GSK3β/cyclin D1 pathway. J
Ethnopharmacol. 210:1–9. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sikander M, Hafeez BB, Malik S, Alsayari
A, Halaweish FT, Yallapu MM, Chauhan SC and Jaggi M: Cucurbitacin D
exhibits potent anti-cancer activity in cervical cancer. Sci Rep.
6:365942016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cicenas J, Kalyan K, Sorokinas A, Jatulyte
A, Valiunas D, Kaupinis A and Valius M: Highlights of the latest
advances in research on CDK inhibitors. Cancers (Basel).
6:2224–2242. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sithara T, Arun KB, Syama HP, Reshmitha TR
and Nisha P: Morin inhibits proliferation of SW480 colorectal
cancer cells by inducing apoptosis mediated by reactive oxygen
species formation and uncoupling of warburg effect. Front
Pharmacol. 8:6402017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Choi AY, Choi JH, Yoon H, Hwang KY, Noh
MH, Choe W, Yoon KS, Ha J, Yeo EJ and Kang I: Luteolin induces
apoptosis through endoplasmic reticulum stress and mitochondrial
dysfunction in Neuro-2a mouse neuroblastoma cells. Eur J Pharmacol.
668:115–126. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shahali A, Ghanadian M, Jafari SM and
Aghaei M: Mitochondrial and caspase pathways are involved in the
induction of apoptosis by nardosinen in MCF-7 breast cancer cell
line. Res Pharm Sci. 13:12–21. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Park J, Ko YS, Yoon J, Kim MA, Park JW,
Kim WH, Choi Y, Kim JH, Cheon Y and Lee BL: The forkhead
transcription factor FOXO1 mediates cisplatin resistance in gastric
cancer cells by activating phosphoinositide 3-kinase/Akt pathway.
Gastric Cancer. 17:423–430. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zheng QS and Zheng RL: Effects of ascorbic
acid and sodium selenite on growth and redifferentiation in human
hepatoma cells and its mechanisms. Pharmazie. 57:265–269.
2002.PubMed/NCBI
|
|
44
|
Balaban RS, Nemoto S and Finkel T:
Mitochondria, oxidants, and aging. Cell. 120:483–495. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chetram MA, Bethea DA, Odero-Marah VA,
Don-Salu-Hewage AS, Jones KJ and Hinton CV: ROS-mediated activation
of AKT induces apoptosis via pVHL in prostate cancer cells. Mol
Cell Biochem. 376:63–71. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pavithra PS, Mehta A and Verma RS:
Aromadendrene oxide 2, induces apoptosis in skin epidermoid cancer
cells through ROS mediated mitochondrial pathway. Life Sci.
197:19–29. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cui YQ, Liu YJ and Zhang F: The
suppressive effects of Britannin (Bri) on human liver cancer
through inducing apoptosis and autophagy via AMPK activation
regulated by ROS. Biochem Biophys Res Commun. 497:916–923. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhong WF, Wang XH, Pan B, Li F, Kuang L
and Su ZX: Eupatilin induces human renal cancer cell apoptosis via
ROS-mediated MAPK and PI3K/AKT signaling pathways. Oncol Lett.
12:2894–2899. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xu H, Li X, Ding W, Zeng X, Kong H, Wang H
and Xie W: Deguelin induces the apoptosis of lung cancer cells
through regulating a ROS driven Akt pathway. Cancer Cell Int.
15:252015. View Article : Google Scholar : PubMed/NCBI
|