Introduction
Despite advances in surgical techniques, radio- and
chemotherapy, the outcome of patients suffering from glioblastoma
multiforme (GBM) remains poor, with a median survival of <15
months (1). In recent years,
immunotherapeutic approaches to malignant glioma have advanced
rapidly. This is based on findings revisiting the traditional
concept of the central nervous system (CNS) as an immunoprivileged
locus. The discovery of the meningeal lymphatic system (2) and an improved understanding of brain T
cell trafficking into the brain via the leptomeninges (3) and the blood-brain-barrier (4) represent important communication
channels between the CNS and the peripheral immune system.
Whereas research has focused on T-cells as the
critical component of the specific antigen-mediated antitumor
response in malignant glioma, there is increasing evidence that
non-specific local immunotherapies may aid in glioma defense.
Clinical observations that postoperative infections within or close
to tumor sites promote a prolonged survival or even complete
remission in patients with GBM (5–9) were
supported by results of two retrospective single-center studies
(10,11) that reported an effect of non-specific
systemic immune responses on glioma growth and surveillance. This
has been confirmed experimentally with a novel approach using the
local administration of heat-inactivated staphylococci as potent
immunomodulators in an experimental gliosarcoma model, which led to
oncolysis and prolonged survival associated with a distinct peri-
and intratumoral infiltration of macrophages (3).
In contrast to the effects of a topical application,
it is still debated if systemic immunostimulation exerts a
significant antitumor effect. This is the basis and potential key
for immunotherapies. Non-specific local immunotherapies have proven
to be ineffective, whereas specific local immunotherapies have
suggested a response (12).
Non-specific systemic immunotherapies have not been validated in
studies, but numerous suggestions were made regarding effects in
allergies and brain abscesses (13).
Host factors, including the tumor microenvironment, considerably
influence glioma growth and targeting angiogenetic and inflammatory
properties recently evolved as an effective treatment strategy
(14).
In 2004, spontaneous regression was observed in
animal studies with experimental gliomas (15). Spheroids of C6 cells were implanted
into the brain and tumor growth was measured by magnetic resonance
imaging (MRI)-based volumetry. The aim of the current study was to
evaluate effects of a stimulation of the innate immune response,
particularly of macrophages, on the proliferation of glioblastoma.
For that purpose, recombinant human granulocyte macrophage colony
stimulating factor (rhGM-CSF), which has been established in the
treatment of neutropenia with a known safety profile, was used in
this study.
Materials and methods
Tumor cell culture and generation of
3D spheroids
Multicellular spheroids from C6 tumor cells (CLS
Cell Lines Service GmbH, Eppelheim, Germany) were generated by
seeding cells in 75 cm2 culture flasks filled with
Dulbecco's modified Eagle's medium (DMEM). Flasks were base-coated
with 1% noble agar (Difco; BD Biosciences, Franklin Lakes, NJ, USA)
dissolved in medium. Incubation was performed at 37°C in the
presence of 5% CO2 with 100% humidity. Spheroids were screened for
signs of necrosis using inverted light microscopy. Following 6
days, spheroids with a diameter of 200–300 µm without a necrotic
core were selected for implantation.
Animal preparation, operative
procedure and stimulation of systemic macrophages
The animal care committee of the district
authorities (Regierung von Unterfranken, Veterinärwesen, Bavaria,
Germany; AZ 55.2–2531.01-65/10) approved the experimental
procedures. The implantation procedure has previously been
described in detail (15,16). For the current study, 29 male
Sprague-Dawley rats (weight, 250–300 g) were anesthetized by an
intraperitoneal (i.p.) injection of ketamine hydrochloride
(Ketavet™; Pfizer, Inc., New York, NY, USA) and xylazine
hydrochloride (Rompun™; Bayer AG, Leverkusen, Germany). The
animals' heads were fixed in a stereotactic frame using
non-perforating bars, a midline incision of the scalp was performed
and a 2 mm burr hole was placed 2 mm left of the bregma. Following
excision of the dura, the cortex was incised in a semicircular
fashion using a microscalpel. Under microscopic view, a single
spheroid was then placed 2–3 mm subcortically. Following surgery,
animals were housed using a 12-h dark/light cycle with free access
to food and water and were monitored for signs of discomfort or
neurological abnormalities daily. Of the 29 animals implanted with
C6 glioma spheroids, 20 rats received a subcutaneous injections of
10 µg/kg rhGSM-CSF every other day. Nine animals served as a
control group.
MRI and tumor volumetry
MRI exams were performed on postoperative days (POD)
7, 14, 21, 28, 32 and 42 with a 3 T clinical scanner (Magnetom
Trio®; Siemens Healthineers, Erlangen, Germany) using a
round surface coil. The following sequences were performed: T1 TSE
cor (0.9 mm), cor T2 TSE rs (0.7 mm) and cor T2 CISS 3D (0.3 mm).
Animals were anesthetized using Ketavet® as described
and were then administered 0.1 ml contrast agent i.p. 10 min prior
to MRI examination. Tumor volumes were calculated using the T2 3D
CISS sequences by using MRI Convert® and
MIPAV® software.
Tissue preparation
Following each MRI exam, two randomly selected
animals were sacrificed for histological examination. Brains and
spleens were removed and immediately fixed in paraformaldehyde
solution for 24 h and stored in cold PBS (pH 7.4; 4°C) for one week
prior to paraffin embedding. For histological studies, spleen
sections and coronal brain sections cut into 4-µm slices.
Histological and immunohistochemical
analyses
Hematoxylin and eosin (H&E) staining was
performed for an estimation of the gross morphology of tumors and
spleen sections.
For immunohistochemistry, sections were stained with
Ki67 and CD8 antibodies targeting CD8+ lymphocytes (AK CD8α;
eBioscience; Thermo Fisher Scientific, Inc., Waltham, MA, USA; cat.
no. 14-0084; AK CD68, Zymed; Thermo Fisher Scientific, Inc.; cat.
no. 603–2210; AK Ki67; Zymed; Thermo Fisher Scientific, Inc.; cat.
no. RMPD 004). Omission of primary antibodies in the control
experiments resulted in the expected absence of any cellular
labeling. The extent of infiltration of different immune-cell
subsets was quantified by cell counting of five representative
high-power fields (HPF) in each section, including the tumor
margins.
In order to estimate the putative effects of
rhGM-CSF stimulation, macrophages and CD8+ lymphocytes in tumor
tissues were counted and the corresponding spleen tissue functioned
as positive control. Sections were counted at a magnification of
×100 using a microscope. In each case, five contiguous fields of
view were counted and the mean was determined. For brain sections,
five visual fields were counted starting from the tumor margin. As
ED1 stains macrophages and microglia, only cells with distinct
phagocyte morphology were considered. In CD8+ cells, only cells
with clear lymphocyte morphology were included.
Data analysis
Survival time is presented using box plots. Tumor
volumes and cell counts are presented as the median ± standard
deviation. Due of the small sample size, all analyses were of
explorative nature. The results (survival times, tumor growth, cell
counts) were evaluated graphically. Animals surviving 42 days
without MRI-based evidence of tumor growth were excluded from the
analysis. Overall survival was assessed by Kaplan-Meier analysis
and differences between survival curves were calculated by using
Graph Pad Prism® (GraphPad Software, Inc, La Jolla, CA,
USA). P<0.05 was considered to indicate a statistically
significant difference. Student's t-tests were used in pairwise
comparisons.
Results
MRI studies
rhGM-CSF group
On POD 7, 2/20 (10%) animals treated with rhGM-CSF
developed a visible tumor. Two animals were sacrificed on POD 7 for
analysis and further two rats were excluded from the trial due to
anesthesia-associated complications, leaving 16 animals for
evaluation on POD 14. At that time, solid tumors were observed in
11/16 (65%) animals and on POD 21, tumors were visible in 14/15
animals. A further animal exhibited a visible tumor on POD 28.
Solid tumors developed in 15/20 (75%) animals. Tumor regression was
observed in six (30%) animals. As a result of severe symptoms
associated with tumor growth, seven (35%) animals of the rhGM-CSF
group were sacrificed during the trial.
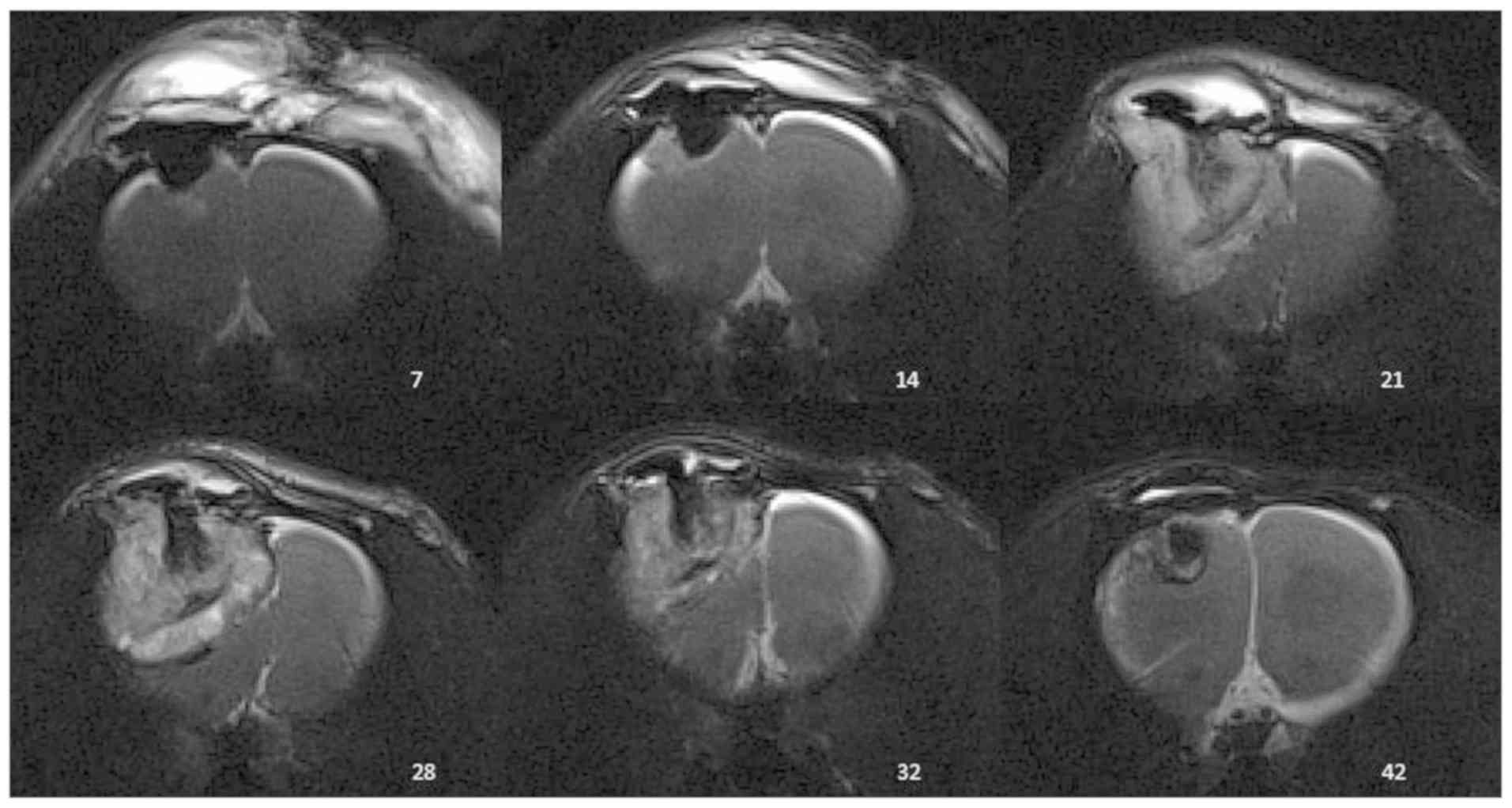
An exemplary course of tumor growth is presented in
Fig. 1. On POD 7, the first
extracranial fluid accumulation caused by the surgical trauma was
detected. On POD 14, minimal tumor growth with contrast uptake at
the edges of the surgical cavity was observed. An increase in size
caused peritumoral edema. On POD 28, a midline shift to the
opposite side developed due to a mass effect. In addition, the
central sparing of contrast enhancement representing necrotic
changes was accompanied by a decrease of tumor volume. The
peritumoral edema disappeared and midline shift was no longer
visible. A small defect with a fading contrast enhancement remained
at POD 32 and 42.
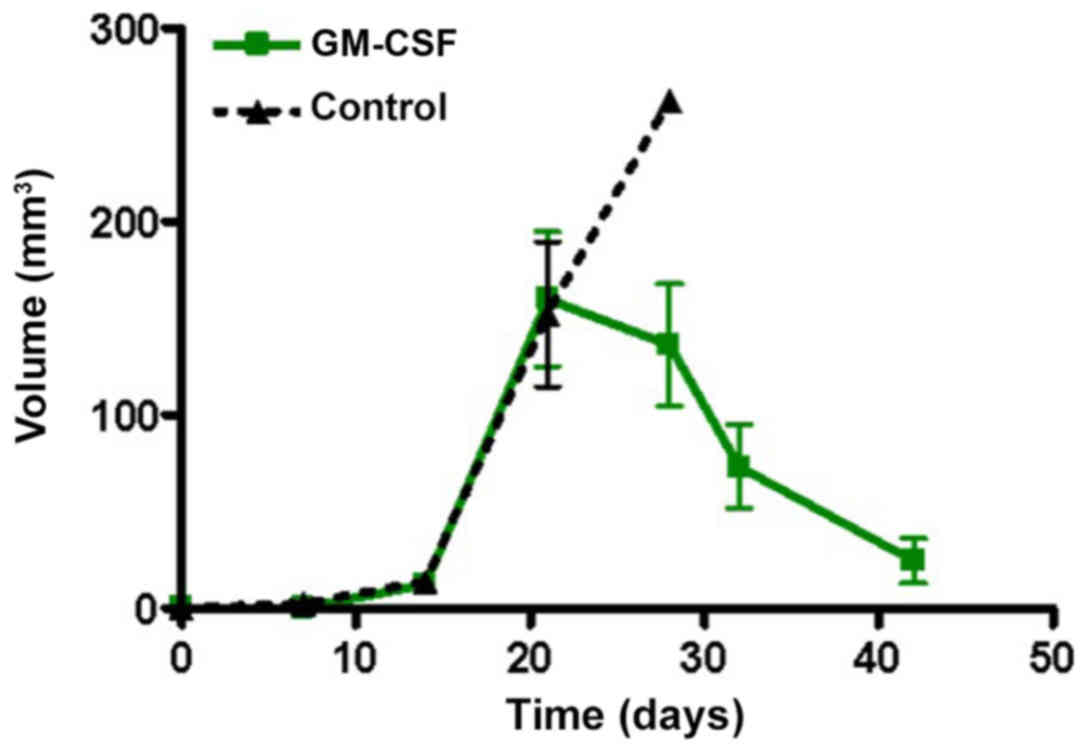
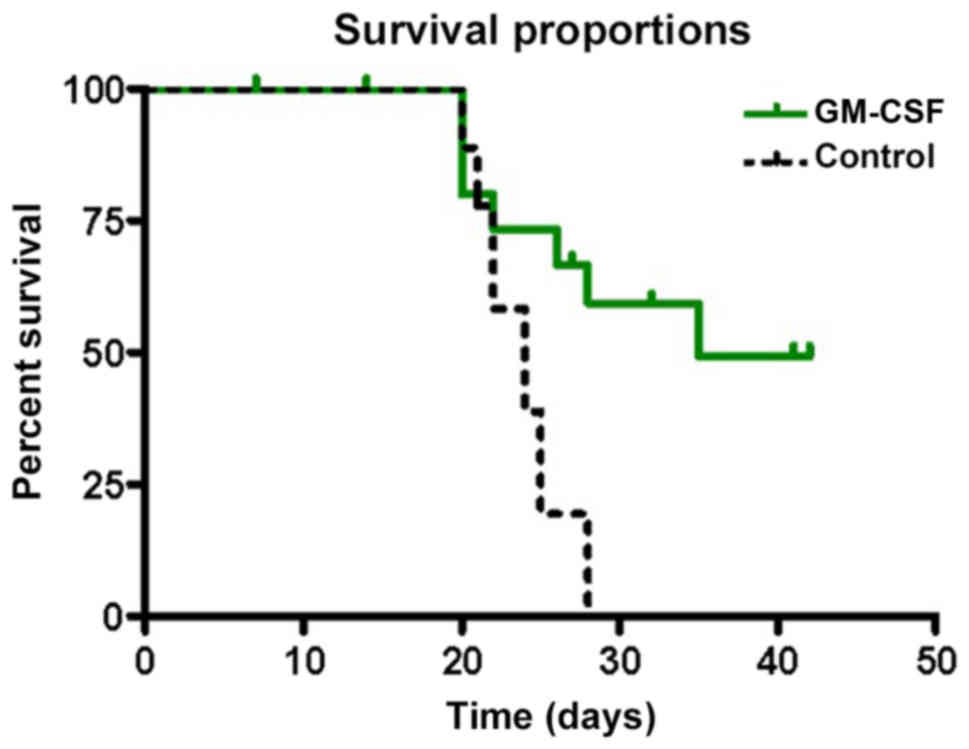
Effects of rhGM-CSF on tumor growth and survival are
documented in Figs. 2 and 3. Until POD 20, no significant differences
were determined. The mean survival time was 35 days in the
rhGM-CSF.
Control group
On POD 7, small tumors were visible in 3/9 (30%)
animals. On POD 14, tumor growth was observed in all control
animals. A total of 6/9 (66%) animals were sacrificed due to severe
symptoms of tumor growth. A total of 3 animals were used for
histological examination. On POD 28, one animal remained, which was
then sacrificed due to tumor mass and occurring symptoms. In
contrast to the rhGM-CSF-group, none of the control animals
exhibited spontaneous regression (Fig.
2). On POD 28, only one measurement was obtained for the
control group and no statistical comparison was performed.
Comparison of treatment and control
groups
Treatment with rhGM-CSF significantly prolonged
survival in the rhGM-CSF group compared with the control group (35
vs. 24 days; P=0.0343). Tumor volume measurements suggested a
delayed growth onset in the rhGM-CSF group compared with the
control and a reduced median volume (134 vs. 262
mm3).
Histology
Spheroids were orthotopically implanted and in 15/20
animals tumor growth was observed. In addition, some animals
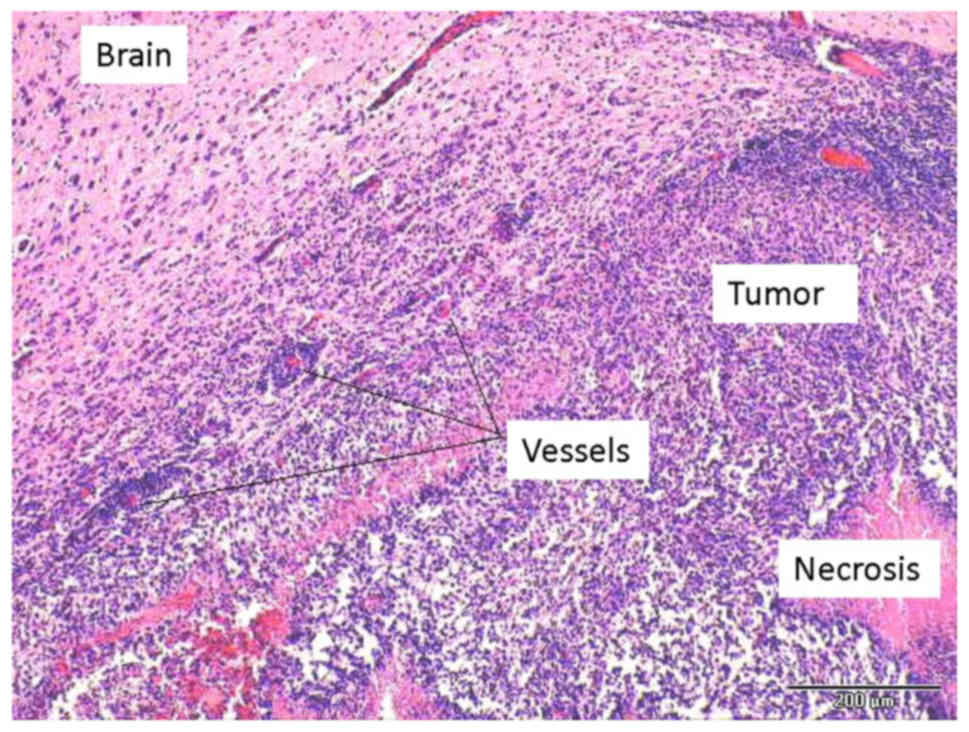
exhibited extracranial tumor components. H&E staining revealed
typical growth characteristics of GBM-type neovascularity, necrosis
and palisade-type arrangement of the tumor cells (Fig. 4).
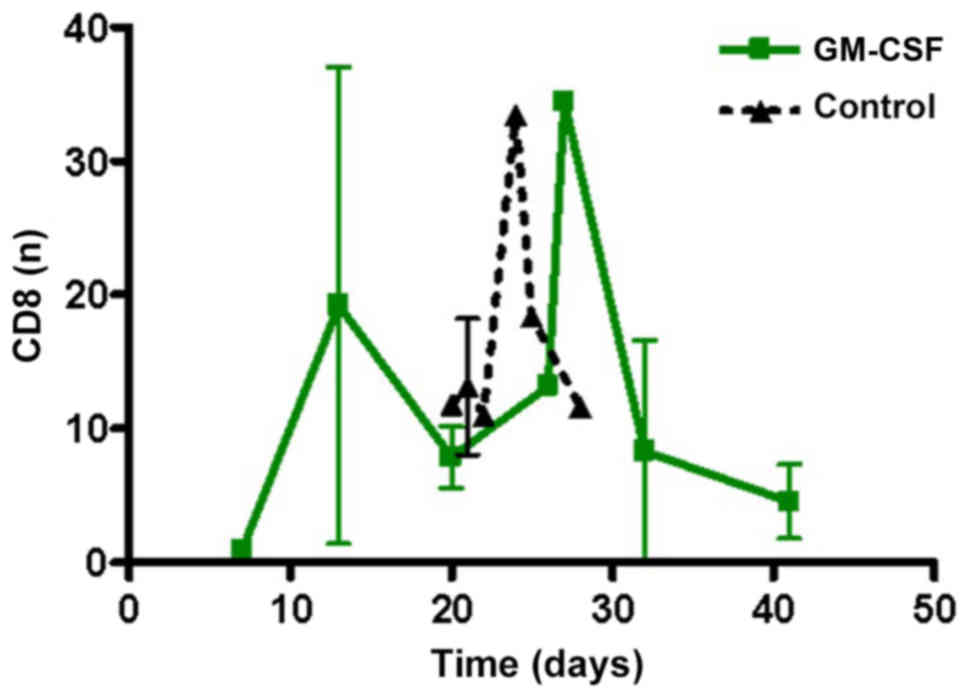
CD8+ cells, including cytotoxic T-cells, were
monitored to reveal potential effects on tumor growth. There was no
significant difference in CD8+ positive cells in the two groups
(Fig. 5).
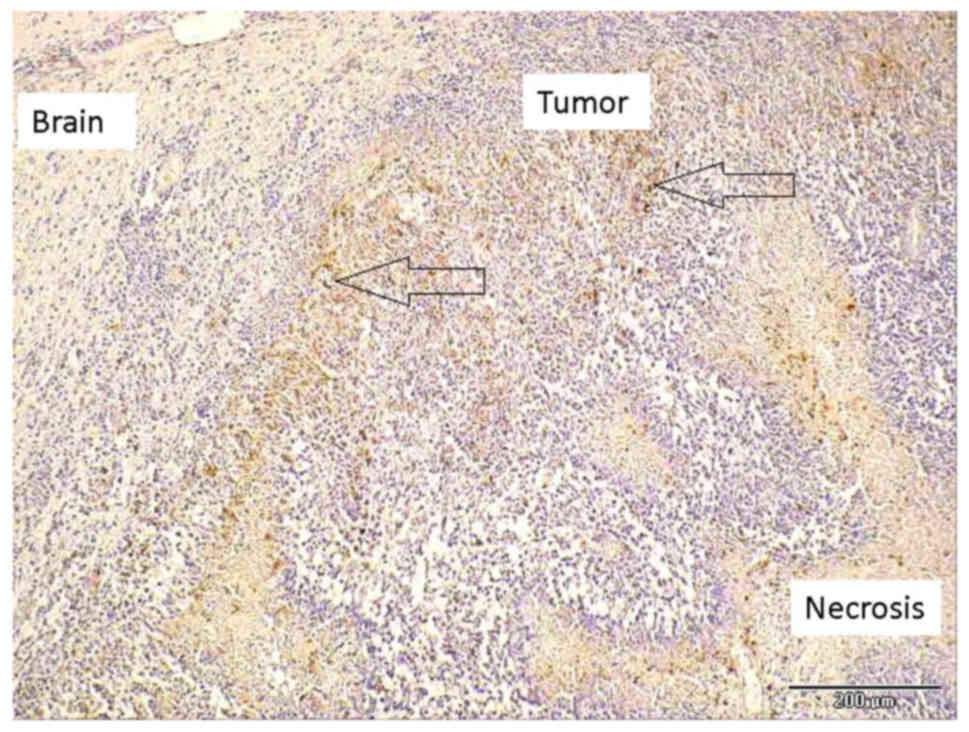
CD68 staining revealed brown-colored macrophages
that were located in necrotic areas and in solid tumor tissues
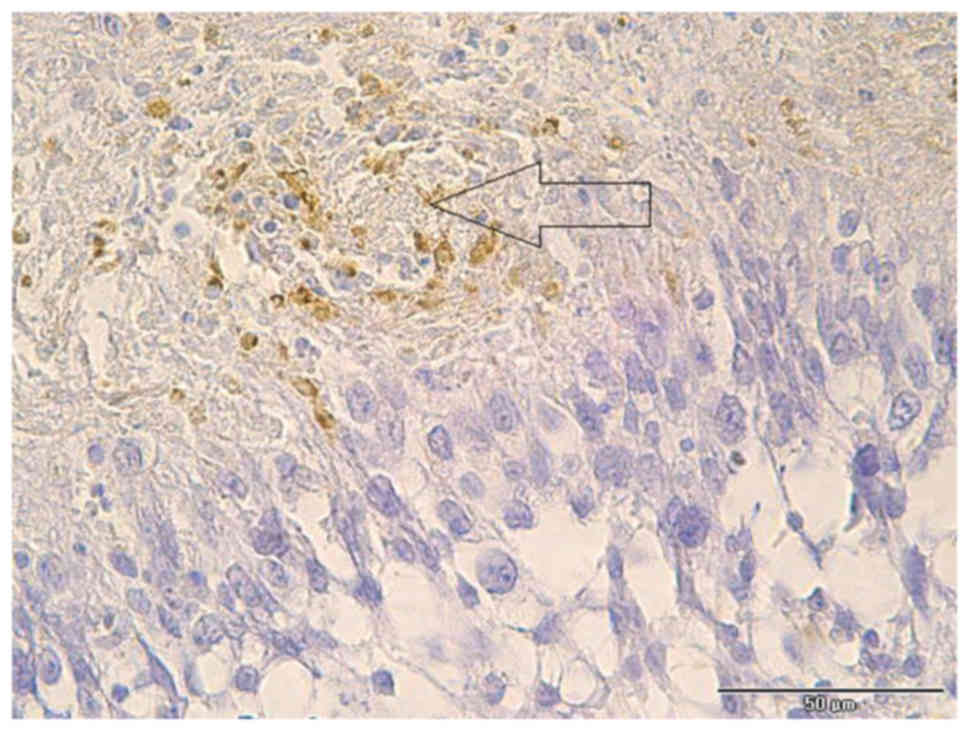
(Fig. 6). In addition, numerous
macrophages were grouped around tumor vessels (Fig. 7).
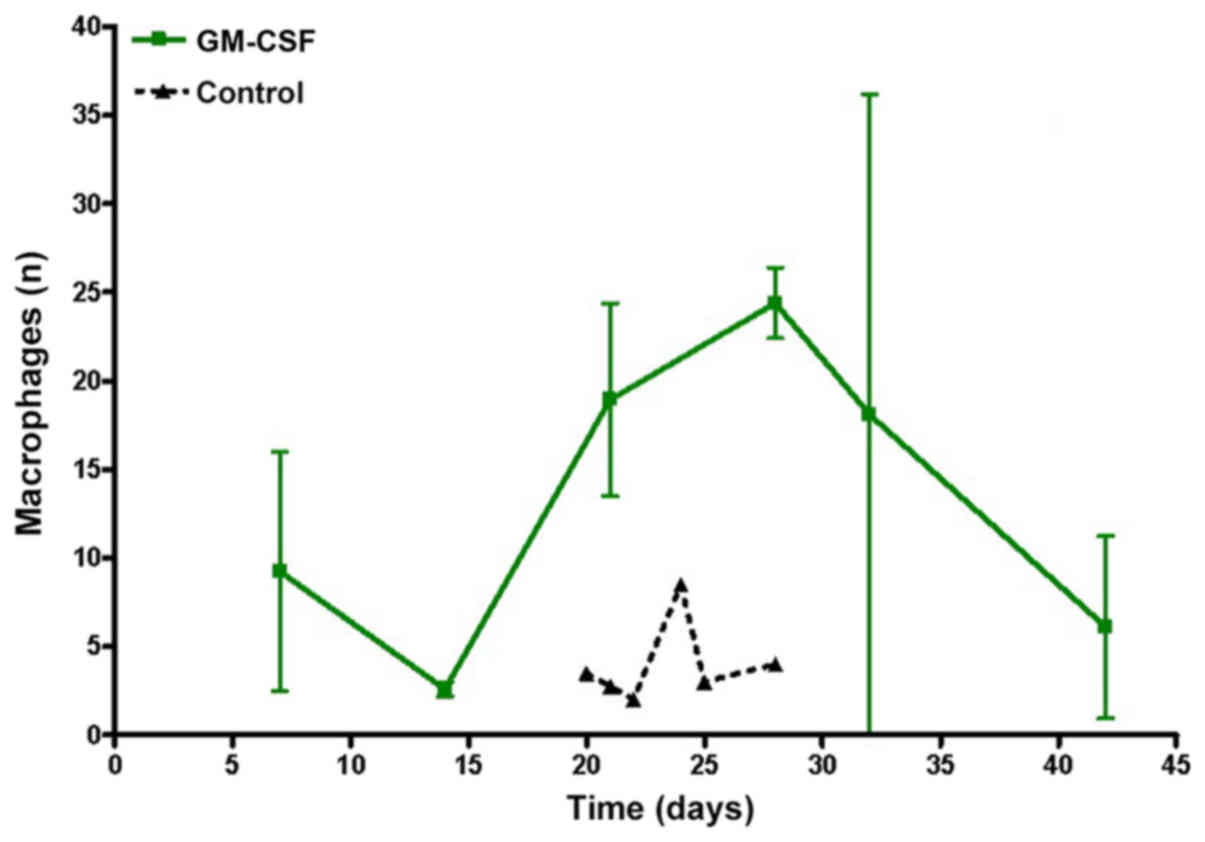
Analyzing the time course of macrophage invasion,
the maximum number was observed when regression started (Fig. 8). Decreasing tumor size led to
decreasing numbers of macrophages. Compared with the control group,
rhGM-CSF animals presented a significantly higher numbers of
macrophages in brain slices (P=0.0275).
Discussion
In the current study, tumor tissues of rhGM-CSF
animals exhibited significantly higher numbers of macrophages in
five-fields-of-view compared with the controls. In addition,
animals of the experimental group survived significantly longer
compared with animals of the control group. Based on the knowledge
that macrophage invasion is correlated with tumor growth in
glioblastoma and experimental models, the current data describing
reduced tumor growth associated with increased macrophage invasion
suggested that the pharmacological induction of macrophages may
attenuate or inhibit tumor growth.
A rat brain tumor implantation model was established
to investigate macrophage infiltration and tumor growth in
rhGM-CSF-treated animals. Compared with mouse models, the larger
size of the rat brain facilitates a more precise intracerebral
implantation of tumor cells and better in vivo imaging and
volumetry of the growing mass (15).
Among the various rat brain tumor models, the C6 glioma model has
been extensively used and is the best characterized experimental
model to investigate a wide array of biological properties of glial
tumors, including, their mechanisms of invasion and angiogenesis,
intratumoral signaling pathways and efficacy of novel therapeutic
modalities (15,17). The C6 glioma model was originally
developed by Benda et al (18) by
repetitive administration of methylnitrosourea to Wistar rats and
was further successfully established in Long-Evans and
Sprague-Dawley rats (19,20). C6 cells share certain morphologic
features of human glioblastoma-type pleomorphic cells, including
nuclear polymorphism, a high mitotic index, areas of necrosis and
invasion into the surrounding brain tissue (21). A limitation of the model is its
immunogenicity. Several rat strains challenged with C6 cells
develop a vigorous immune response. Hence, studies on
immunotherapies using the C6 model require careful interpretation
(22).
In brain tumors, high numbers of macrophages are
detected, increasing with the degree of malignancy (23,24). A
total of 5–30% are typified as microglia/macrophages and account
for the majority of infiltrating immune cells in gliomas (25). The majority of studies does not
distinguish between microglia and systemic macrophages (CD68 and
CD11b/c positive) (26–28). Badie et al (23) have reported that a differentiation of
the CD11b/c positive cells is possible through quantification due
to low expression of CD45 in microglia and increased expression in
macrophages (1,23,29).
Initially, it was assumed that high numbers of microglial cells
indicate a strong antitumor response in gliomas; however, certain
studies postulated a positive effect on tumor growth and a support
of the immunosuppressive peritumoral region (23,30).
There are various synergistic mechanisms between glioma cells and
microglia/macrophages that ensure an immunosuppressant milieu. On
the one hand, microglia express low levels of major
histocompatibility complex class II (MHC II) in the vicinity of the
glioma. The production of transforming growth factor (TGF) β by
glioma cells causes downregulation of MHC class II expression
(31). These molecules are essential
in the interaction of antigen-presenting cells and T-lymphocytes
(32–34). On the other hand, glioma cells
produce substances with immunosuppressive effects. These substances
include interleukin (IL) 4, IL 6, TGF β [21] and prostaglandin (PG)
E2 (32).
Further studies reported a positive effect of high
numbers of tumor-macrophages (35,36).
Galarneau et al (37) analyzed the
influence of macrophages on the growth of glioma cells using a
transgenic mouse model. Depletion of the macrophages leads to an
increase of tumor volume by ≤33%. It is a well-known fact that
macrophages are able to recruit T cells by release of tumor
necrosis factor (TNF) (38).
However, it remains to be elucidated whether effects on tumor
growth are driven by macrophages alone or through TNF-mediated
activation of T cells. In addition, a lower vessel density of ~12%
in macrophage-depleted animals was observed. Thus, it is unlikely
that increased tumor growth is associated with increased vascular
supply (27). Furthermore,
Villeneuve et al (38) struggled to
determine whether an altered vascular supply in the tumor is caused
by the depletion of macrophages.
To determine effects of rhGM-CSF treatment, the M1
and M2 status of macrophages has to be considered. Macrophages
present in two different forms, the M1 and M2 status (32,33,39). The
M1 status describes classically-activated macrophages associated
with inflammation (22,29). In the presence of various cytokines,
including rhGM-CSF and interferon γ, monocytes develop to
macrophages with M1 status (33).
Expression of signal transducer and activator of transcription
(STAT) 1, M1 macrophages exhibit antimicrobial, immunostimulatory
and antitumor functions (40). TNF
is another factor that contributes to the activation and
recruitment of microglia/macrophages. Production of this
proinflammatory cytokine contributes to recruitment (33). The M2 status arises from alternative
activation and describes macrophages under normal conditions. Among
other factors, this status is responsible for preventing excessive
immune reactions and explains the more immunosuppressive character
of M2 (38). The conversion from M1
to M2 status is induced by IL 4, IL 6, IL 10 and M-CSF. Expression
of STAT3 causes M2 macrophage activation, including tissue repair
and support of angiogenesis, and favors a tumor progression. This
is mediated by the release of various compounds, including IL 10
and TGF (33,41).
During the early stages of glioma development,
macrophages are arrested in the M1 tumor suppressive status. The
percentage of microglia/macrophages arresting in M2 correlates with
the histological grade of malignancy in glioma (35,37,39).
Certain mediators produced by the tumor induce the conversion from
M1 to M2; these include IL 4, IL 10, TGF β and M-CSF (35,39,40,42).
M-CSF produced by tumor cells increases M2 microglia and
macrophages and promotes tumor growth and proliferation (40). Reduced tumor growth is detected when
inhibiting the M2 status (42,43).
Macrophage activation by rhGM-CSF may be a novel therapeutic
approach in preserving the M1 status for prolonged periods,
preventing a transition into the M2 status (43). Microglial cells treated with rhGM-CSF
exhibit upregulated growth and form a heterogeneous cell population
in vitro, similar to macrophages. Their function is strongly
associated with local environmental factors, such as rhGM-CSF and
M-CSF (30,44).
Expression of the Fas-ligand enable
macrophage/microglia to support the immunosuppressive environment
of gliomas (41,45). There is evidence that cancer stem
cells represent the major cause for reprogramming
microglia/macrophages to adapt the immunosuppressive M2 status.
Tumor stem cells produce soluble (s) CSF, macrophage cytokine 1
(MIC1) and TGF β1. These substances polarize microglia/macrophages
in the M2 status, block phagocytosis and the production of
immunosuppressive cytokines is induced, including IL 10 and TGF β1.
In addition, inhibition of T-cell proliferation is observed
(41,45,46).
Macrophages exhibit tumor suppressive functions at later stages in
the development of gliomas. Changes may be influenced
pharmacologically during this period (41,47). It
has been postulated that the therapeutic approach using
tumor-associated macrophages is more successful with a staggered
induction (43). Considering that
different means of activation trigger various responses explains
the comparatively late tumor growth observed in a number of rats of
the GM-CSF group in the current study. A novel therapeutic approach
to treating GBM via targeting macrophages at different states of
activity may be considered in the future (32,39,40).
Additionally, it has to be clarified, which type of macrophages is
promoted by administration of rhGM-CSF that further stimulates
tumor progression (35,48). Depletion of rhGM-CSF led to a reduced
number of tumor-promoting macrophages and interfered with the
development of proinvasive macrophages. In animal studies,
rhGM-CSF-depleted animals exhibit increased overall survival
(48). In contrast, Grabstein et al
(49) described enhanced tumor
suppressing properties of macrophages by stimulating with rhGM-CSF
in vitro and noted that macrophages exhibit improved antitumor
activities.
Effects of rhGM-CSF on an immune system with tumor
control are associated with the dose (50). High systemic concentrations lead to
recruitment of regulatory T cells, which contribute to the
immunosuppressive environment and interfere with the response
against the tumor exerted by the immune system itself. Low doses in
the context of vaccinations with tumor antigens lead to an immune
stimulation and enhance antitumor activity (50,51). To
reduce inter-individual differences, investigations were performed
using genetically identical mice. It is postulated that rhGM-CSF at
high doses reduces the expression of M-CSF receptor, in contrast to
a promotion of receptor expression that is observed at low doses
(52,53). In humans, treatment with defined
amounts of GM-CSF lead to varying serum levels of M-CSF receptor
due to individual variance (51).
Data presented in the current study supported the
hypothesis of an antitumor effect of rhGM-CSF. The number of
macrophages counted in tumor tissues of the rhGM-CSF group was
increased compared with the control group. An analysis of
macrophage counts and tumor volume over time produced supporting
results; initially an increase was observed that towards the end of
the trial almost reached baseline levels. It was observed that
rhGM-CSF increased macrophage accumulation and exhibited a positive
effect on tumor suppression.
In conclusion, a systemic stimulation of macrophages
by rhGM-CSF led to significantly reduced and delayed tumor growth
in a rodent C6 glioma model. Model being aware that studies on
immunotherapies using the C6 rat model have to be interpreted
carefully because of its immunogenicity.
The host control of experimental gliomas by
macrophages may be combined with other promising immune-based
approaches, including chimeric antigen receptor T-cell technology
or PD-1/PD-L1 checkpoint inhibitors. The role macrophages serve in
host tumor control may have been underestimated in the past.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TL and AJ performed the experiments to collect the
data. TL drafted the manuscript. CMM performed the histological
examinations. AFK, TW made substantial contributions to conception
and design and analyzing the data. ML made substantial
contributions to conception and design, was involved in drafting
the manuscript, revised it critically for important intellectual
content and gave the final approval of the version to be published.
GHV and RIE made substantial contributions to conception and
design, interpretation of the data, were involved in drafting the
manuscript and revised it critically for important intellectual
content. GH performed the neuroradiological examinations. ML was
involved in drafting the manuscript and gave final approval for
publication. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The experimental procedures were approved by the
Animal Care Committee of the district authorities (Regierung von
Unterfranken, Veterinärwesen, Bavaria, Germany; AZ
55.2–2531.01-65/10).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: Effects of radiotherapy with concomitant and
adjuvant temozolomide versus radiotherapy alone on survival in
glioblastoma in a randomised phase III study: 5-year analysis of
the EORTC-NCIC trial. Lancet Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Louveau A, Smirnov I, Keyes TJ, Eccles JD,
Rouhani SJ, Peske JD, Derecki NC, Castle D, Mandell JW, Lee KS, et
al: Structural and functional features of central nervous system
lymphatic vessels. Nature. 523:337–341. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schläger C, Körner H, Krueger M, Vidoli S,
Haberl M, Mielke D, Brylla E, Issekutz T, Cabañas C, Nelson PJ, et
al: Effector T-cell trafficking between the leptomeninges and the
cerebrospinal fluid. Nature. 530:349–353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ratnam NM, Gilbert MR and Giles AJ:
Immunotherapy in CNS cancers: The role of immune cell trafficking.
Neuro Oncol. 21:37–46. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bowles AP Jr and Perkins E: Long-term
remission of malignant brain tumors after intracranial infection: A
report of four cases. Neurosurgery. 44:636–643. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kapp JP: Microorganisms as antineoplastic
agents in CNS tumors. Arch Neurol. 40:637–642. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Margolis J and West D: Spontaneous
regression of malignant disease: Report of three cases. J Am
Geriatr Soc. 15:251–253. 1967. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Naganuma H, Sasaki A, Satoh E, Nagasaka M,
Isoe S, Nakano S and Nukui H: Long-term survival in a young patient
with anaplastic glioma. Brain Tumor Pathol. 14:71–74. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Walker DG and Pamphlett R: Prolonged
survival and pulmonary metastases after local cure of glioblastoma
multiforme. J Clin Neurosci. 6:67–68. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
De Bonis P, Albanese A, Lofrese G, de
Waure C, Mangiola A, Pettorini BL, Pompucci A, Balducci M,
Fiorentino A, Lauriola L, et al: Postoperative infection may
influence survival in patients with glioblastoma: Simply a myth?
Neurosurgery. 69:864–869. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bohman LE, Gallardo J, Hankinson TC,
Waziri AE, Mandigo CE, McKhann GM II, Sisti MB, Canoll P and Bruce
JN: The survival impact of postoperative infection in patients with
glioblastoma multiforme. Neurosurgery. 64:828–835. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Platten M, Bunse L, Wick W and Bunse T:
Concepts in glioma immunotherapy. Cancer Immunol Immunother.
65:1269–1275. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Löhr M, Molcanyi M, Poggenborg J,
Spuentrup E, Runge M, Röhn G, Härtig W, Hescheler J and Hampl JA:
Intracerebral administration of heat-inactivated Staphylococcus
epidermidis enhances oncolysis and prolongs survival in a 9L
orthotopic gliosarcoma model. Cell Physiol Biochem. 31:614–624.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thomas AA, Brennan CW, DeAngelis LM and
Omuro AM: Emerging therapies for glioblastoma. JAMA Neurol.
71:1437–1444. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vince GH, Bendszus M, Schweitzer T,
Goldbrunner RH, Hildebrandt S, Tilgner J, Klein R, Solymosi L,
Christian Tonn J and Roosen K: Spontaneous regression of
experimental gliomas-an immunohistochemical and MRI study of the C6
glioma spheroid implantation model. Exp Neurol. 190:478–485. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goldbrunner RH, Bernstein JJ, Plate KH,
Vince GH, Roosen K and Tonn JC: Vascularization of human glioma
spheroids implanted into rat cortex is conferred by two distinct
mechanisms. J Neurosci Res. 55:486–495. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Grobben B, De Deyn PP and Slegers H: Rat
C6 glioma as experimental model system for the study of
glioblastoma growth and invasion. Cell Tissue Res. 310:257–270.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Benda P, Lightbody J, Sato G, Levine L and
Sweet W: Differentiated rat glial cell strain in tissue culture.
Science. 161:370–371. 1968. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nagano N, Sasaki H, Aoyagi M and Hirakawa
K: Invasion of experimental rat brain tumor: Early morphological
changes following microinjection of C6 glioma cells. Acta
Neuropathol. 86:117–125. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Whittle IR, MacArthur DC, Malcom GP, Li M,
Washington K and Ironside JW: Can experimental models of rodent
implantation glioma be improved? A study of pure and mixed glioma
cell line tumours. J Neurooncol. 36:231–242. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chicoine MR and Silbergeld DL: Invading C6
glioma cells maintaining tumorigenicity. J Neurosurg. 83:665–671.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Parsa AT, Chakrabarti I, Hurley PT, Chi
JH, Hall JS, Kaiser MG and Bruce JN: Limitations of the C6/Wistar
rat intracerebral glioma model: Implications for evaluating
immunotherapy. Neurosurgery. 47:993–1000. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Badie B and Schartner J: Role of microglia
in glioma biology. Microsc Res Tech. 54:106–113. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Roggendorf W, Strupp S and Paulus W:
Distribution and characterization of microglia/macrophages in human
brain tumors. Acta Neuropathol. 92:288–293. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Badie B and Schartner JM: Flow cytometric
characterization of tumor-associated macrophages in experimental
gliomas. Neurosurgery. 46:957–962. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
da Fonseca AC and Badie B: Microglia and
macrophages in malignant gliomas: Recent discoveries and
implications for promising therapies. Clin Dev Immunol.
2013:2641242013.PubMed/NCBI
|
|
27
|
Coniglio SJ and Segall JE: Review:
Molecular mechanism of microglia stimulated glioblastoma invasion.
Matrix Biol. 32:372–380. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wagner S, Czub S, Greif M, Vince GH, Süss
N, Kerkau S, Rieckmann P, Roggendorf W, Roosen K and Tonn JC:
Microglial/macrophage expression of interleukin 10 in human
glioblastomas. Int J Cancer. 82:12–16. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sedgwick JD, Schwender S, Imrich H,
Dörries R, Butcher GW and ter Meulen V: Isolation and direct
characterization of resident microglial cells from the normal and
inflamed central nervous system. Proc Natl Acad Sci USA.
88:7438–7442. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang I, Han SJ, Kaur G, Crane C and Parsa
AT: The role of microglia in central nervous system immunity and
glioma immunology. J Clin Neurosci. 17:6–10. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Czarniecki CW, Chiu HH, Wong GH, McCabe SM
and Palladino MA: Transforming growth factor-beta 1 modulates the
expression of class II histocompatibility antigens on human cells.
J Immunol. 40:4217–4223. 1988.
|
|
32
|
Charles NA, Holland EC, Gilbertson R,
Glass R and Kettenmann H: The brain tumor microenvironment. Glia.
59:1169–1180. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Watters JJ, Schartner JM and Badie B:
Microglia function in brain tumors. J Neurosci Res. 81:447–455.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Badie B, Bartley B and Schartner J:
Differential expression of MHC class II and B7 costimulatory
molecules by microglia in rodent gliomas. J Neuroimmunol.
133:39–45. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hambardzumyan D, Gutmann DH and Kettenmann
H: The role of microglia and macrophages in glioma maintenance and
progression. Nat Neurosci. 19:20–27. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zeiner PS, Preusse C, Blank AE, Zachskorn
C, Baumgarten P, Caspary L, Braczynski AK, Weissenberger J, Bratzke
H, Reiß S, et al: MIF receptor CD74 is restricted to
microglia/macrophages, associated with a M1-polarized immune milieu
and prolonged patient survival in gliomas. Brain Pathol.
25:491–504. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Galarneau H, Villeneuve J, Gowing G,
Julien JP and Vallières L: Increased glioma growth in mice depleted
of macrophages. Cancer Res. 67:8874–8881. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Villeneuve J, Tremblay P and Vallières L:
Tumor necrosis factor reduces brain tumor growth by enhancing
macrophage recruitment and microcyst formation. Cancer Res.
65:3928–3936. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Komohara Y, Ohnishi K, Kuratsu J and
Takeya M: Possible involvement of the M2 anti-inflammatory
macrophage phenotype in growth of human gliomas. J Pathol.
216:15–24. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wei J, Gabrusiewicz K and Heimberger A:
The controversial role of microglia in malignant gliomas. Clin Dev
Immunol. 2013:2852462013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mantovani A, Marchesi F, Malesci A, Laghi
L and Allavena P: Tumour-associated macrophages as treatment
targets in oncology. Nat Rev Clin Oncol. 14:399–416. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Alterman RL and Stanley ER: Colony
stimulating factor-1 expression in human glioma. Mol Chem
Neuropathol. 21:177–188. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gabrusiewicz K, Ellert-Miklaszewska A,
Lipko M, Sielska M, Frankowska M and Kaminska B: Characteristics of
the alternative phenotype of microglia/macrophages and its
modulation in experimental gliomas. PLoS One. 6:e239022011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fischer HG and Reichmann G: Brain
dendritic cells and macrophages/microglia in central nervous system
inflammation. J Immunol. 166:2717–2726. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Badie B, Schartner J, Prabakaran S, Paul J
and Vorpahl J: Expression of Fas ligand by microglia: Possible role
in glioma immune evasion. J Neuroimmunol. 120:19–24. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wu A, Wei J, Kong LY, Wang Y, Priebe W,
Qiao W, Sawaya R and Heimberger AB: Glioma cancer stem cells induce
immunosuppressive macrophages/microglia. Neuro Oncol. 12:1113–1125.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kennedy BC, Maier LM, D'Amico R, Mandigo
CE, Fontana EJ, Waziri A, Assanah MC, Canoll P, Anderson RC,
Anderson DE and Bruce JN: Dynamics of central and peripheral
immunomodulation in a murine glioma model. BMC Immunol. 10:112009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sielska M, Przanowski P, Wylot B,
Gabrusiewicz K, Maleszewska M, Kijewska M, Zawadzka M, Kucharska J,
Vinnakota K, Kettenmann H, et al: Distinct roles of CSF family
cytokines in macrophage infiltration and activation in glioma
progression and injury response. J Pathol. 230:310–321. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Grabstein KH, Urdal DL, Tushinski RJ,
Mochizuki DY, Price VL, Cantrell MA, Gillis S and Conlon PJ:
Induction of macrophage tumoricidal activity by
granulocyte-macrophage colony-stimulating factor. Science.
232:506–508. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Serafini P, Carbley R, Noonan KA, Tan G,
Bronte V and Borrello I: High-dose granulocyte-macrophage
colony-stimulating factor-producing vaccines impair the immune
response through the recruitment of myeloid suppressor cells.
Cancer Res. 64:6337–6343. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Rapoport AP, Abboud CN and DiPersio JF:
Granulocyte-macrophage colony-stimulating factor (GM-CSF) and
granulocyte colony- stimulating factor (G-CSF): Receptor biology,
signal transduction, and neutrophil activation. Blood Rev. 6:43–57.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gliniak BC and Rohrschneider LR:
Expression of the M-CSF receptor is controlled
posttranscriptionally by the dominant actions of GM-CSF or
multi-CSF. Cell. 63:1073–1083. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
McLay RN, Kimura M, Banks WA and Kastin
AJ: Granulocyte-macrophage colony-stimulating factor crosses the
blood-brain and blood-spinal cord barriers. Brain. 120:2083–2091.
1997. View Article : Google Scholar : PubMed/NCBI
|