Introduction
Metastasis is the primary cause of mortality in
patients with cancer; however, it is poorly understood due to its
complexity and multiple step nature (1). Adhesive interactions between the
circulating tumor cells and the target organ are an important early
step in the formation of metastasis at secondary sites (1,2). Since
adhesive interactions are the first step of cancer metastasis, it
is imperative to understand the mechanisms contributing to this
process, as inhibiting this early metastatic step will prevent
subsequent metastatic processes, including migration,
intravasation, invasion and new metastatic growths (3–5).
Disseminated cancer cells exhibit varying degrees of
adhesive potential, which is dependent on environmental factors,
including serum growth factors, extracellular matrix composition,
physical forces, temperature and humidity (6,7). In
particular, mechanical forces generated in the circulation during
laparoscopic insufflation, surgical manipulation, postoperative
abdominal third spacing and passage through the lymphatics and
circulation may increase the adhesive potential of a wide variety
of cancer cells from colonic cancer (8), breast cancer (9), head and neck squamous cell cancer
(10) and sarcoma types (11). The stimulation of cancer cell
adhesion by extracellular pressure therefore represents an
interesting mechanism that regulates cancer cell adhesiveness and a
probe to delineate the relevant intracellular signal pathway, in
the hope that the identified novel targets can be blocked to
inhibit metastasis. Our previous investigation reported that this
effect is statistically significant at 10 mmHg, and was maximal at
15 mmHg (12), yet increased
pressures do not yield an increased effect, at least in
vitro (12). Since the effect is
maximal at 15 mmHg and as 15 mmHg is the pressure most commonly
used to inflate the abdomen in laparoscopic surgery (13), 15 mmHg was selected to investigate
increased pressures above ambient in the present study.
Mechanical forces, such as increased extracellular
pressure and shear stress, activate cytoskeletally-dependent and
cytoskeletally-independent pathways that converge on the activation
of focal adhesion kinase (FAK) (14). Although FAK-Tyr-397 phosphorylation
is generally conceived as the initial step in FAK activation
(15), it was observed that FAK
activation in cancer cells in response to physical forces requires
the upstream activation of Akt1 at the catalytic region Ser-473,
yet not at the regulatory site Thr-308, which then binds to FAK and
induces its phosphorylation at serine 517, 601 and 695 and
threonine 600 (16,17). As FAK activation does not generally
require Akt, this novel interaction may be a promising target for
pharmacologic blockade, which may not interfere with other aspects
of FAK signaling, and therefore provides a more specific targeting
mechanism than the global FAK inhibitors currently in trials
(18–21). Serial truncation of the FAK FERM
domain demonstrated that FAK-Akt1 binding occurs in sub domain
1–126 amino acids (22). Further
serial truncation of this domain narrowed down to 33 amino acids
NT1-2-2 FERM (amino acid sequence 94–126 of FERM) subdomain of FAK
is required for Akt1 binding (23),
and further studies condensed this further to the 7 amino acid
sequence 113LAHPPEEQ119, which is a short helical structure
(24). Adenoviral expression of this
peptide in colon cancer cells prevents activation of FAK in
response to extracellular pressure, attenuates FAK-Akt1
interaction, and inhibits the pressure-induced adhesion of colon
cancer cells to a collagen I substrate (24). Furthermore, cells transiently
overexpressing this peptide are less adherent in murine surgical
wound tumor recurrence models, decreasing tumor incidence and
improving subsequent survival (24).
However, small molecules may be more advantageous as
therapeutic agents than peptides with respect to half-life, cell
permeability and stability (25,26). For
instance, small molecules are more stable than peptides in various
physiological environments, including those at varying pH, or
containing degradation enzymes, or with different cation or anion
concentrations (25,26). In a previous preliminary study, a
three dimensional ligand-based virtual screen was performed to
identify small molecules that may mimic the structure of this
peptide within FAK, and reported that two such molecules,
ZINC04085549 and ZINC4085554, could inhibit the pressure-induced
adhesion of SW620 colon cancer cells to collagen I (27). The aim of the present study was to
examine the effects of ZINC4085554 on adhesion to collagen I in
vitro and to wound extracellular matrix (of which type I
collagen is the dominant collagen) in vivo. The promising
molecule, ZINC4085554 was selected, to delineate the mechanism of
this anti-adhesive effect, and examine the hypothesis that
ZINC4085554 reduces cancer cell adhesion by inhibiting the
interaction of Akt1 with FAK, as well as the additional hypothesis
that this molecule would not interfere with more conventional FAK
signaling. The results of the present study confirmed that
ZINC4085554 can inhibit cancer cell adhesion not only to purified
proteins in cell culture, but also to intact tissues in living
mice.
Materials and methods
Cell culture and Reagents
The SW620 human colon cancer cell line was purchased
from the American Tissue Culture Collection (Manassas, VA, USA),
and were routinely cultured in Leibovitz's L-15 (L15) media (cat.
no. 11415-064; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing penicillin and streptomycin (cat. no. 15140-122; Thermo
Fisher Scientific, Inc.) at 100 U/mL and 100 µg/ml, respectively,
and 10% fetal bovine serum (FBS). Cells placed at 37°C in a
humidified incubator containing 5% CO2.
Extracellular pressure treatment
Cells (50,000 cells/well) were seeded into 24 well
plates that were then subjected to 15 mmHg increased extracellular
pressure for 30 min at 37°C in a mixture of 5% CO2 with
95% room air in a pre-warmed air tight box with inlet and outlet
valves and a pressure gauge, as described previously (17,24,27). It
has been previously reported that this technique, if the box is
appropriately pre-warmed, allows for the control temperature,
pressure and oxygen partial pressure within the cell culture
medium. A preliminary study demonstrated the ability to maintain
constant temperature and pressure conditions of 2°C and 1.5 mmHg,
respectively, using this method (12).
Adhesion assay
Our previous studies performed cell counting via
optical microscope (23,28) and Tag-it dye (Tag-it Violet
Proliferation Cell Tracking Dye; Biolegend, Inc., San Diego, CA,
USA) staining method (24,27) for cell counting and have revealed
that dye staining methods are more objective as they do not require
the observer to select which microscopic fields to count and are
more accurate as they compensate for any unevenness of distribution
of cells across the well. Therefore, the adherent cells were not
imaged, and only their optical densities were measured. Our studies
(24,27) and other studies (29,30)
routinely used such techniques to measure adhesion.
The adhesion assay was performed in petri dishes
pre-coated with type I collagen (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany), as previously described (12). Cells treated at 37°C for 1 h with
vehicle [0.1% dimethyl sulfoxide (DMSO)] or ZINC4085554 (10, 25 and
50 µM) were seeded into collagen I pre-coated 24 well plate at
50,000 cells/well. One plate was maintained at ambient pressure
while the other was subjected to 15 mmHg pressure, each in 5%
CO2 at 37°C. At 30 min, the non-adherent cells were
washed away PBS. The remaining adherent cells were stained with MTS
(CellTiter 96 Aqueous One Solution Cell Proliferation Assay;
Promega Corporation, Madison, WI, USA) at 37°C for 1 h and
absorbance was measured at 490 nm.
Western blotting and
co-immunoprecipitation (co-IP)
As the signal cascade examined herein occurs in
suspended cells prior to adhesion, the majority of signaling
studies were performed in dishes pre-coated with 1% heat
inactivated bovine serum albumin (BSA) (Sigma-Aldrich; Merck KGaA)
to prevent adhesion. SW620 cells were treated with vehicle (0.1%
DMSO) or ZINC4085554 (50 µM) and subjected to ambient and 15 mmHg
pressure in 1% heat inactivated BSA pre-coated dishes, for 30 min
at 37°C in 5% CO2. The cells were then collected and
lysed in lysis buffer (50 mM Tris, 150 mM NaCl, 1 mM EDTA, 1 mM
EGTA, 1% Triton-X-100, 1% deoxycholic acid, 0.1% SDS, 10% glycerol,
and protease and phosphatase inhibitors) and concentration of
protein was estimated by bicinchoninic acid. Proteins (50 µg) were
separated by 10% SDS-PAGE and transferred onto nitrocellulose
membranes. Membranes were blocked with blocking buffer (Odyssey
Blocking Buffer; LI-COR Biosciences, Lincoln, NE, USA) at room
temperature for 1 h and immunoblotted at 4°C overnight with primary
antibodies as follows: FAK (1:1,000; cat. no. 05-537, clone 4.47;
mouse monoclonal; Merck KGaA, Darmstadt, Germany), FAK-Try-397
(1:1,000; cat. no. ab81298; rabbit monoclonal; Abcam, Cambridge,
UK), Akt1 (1:1,000; cat. no. 2967, clone 2H10; mouse monoclonal;
Cell Signaling Technology, Inc., Danvers, MA, USA), Akt1-Ser-473
(1:1,000; cat. no. 9276; mouse monoclonal; Cell Signaling
Technology, Inc.) and Akt-Thr-308 (1:1,000; cat. no. 9275; Cell
Signaling Technology, Inc.). Infrared fluorescence dye (IRDye)
detection method was used to visualize the immunoblots. IRDye
conjugated secondary antibodies were the rabbit IRDye 680 (cat. no.
P/N 925-68073), rabbit IRDye 800 (cat. no. P/N 925-32213), mouse
IRDye 680 (cat. no. P/N 925-68072) and mouse IRDye 800 (cat. no.
P/N 925-32212). All secondary antibodies were purchased from LI-COR
Biosciences, used at 1:15,000 and incubated at room temperature for
1 h. The IRDye 680 and IRDye 800 conjugated secondary antibodies
were used to detect the signal at 700 nm and 800 nm channel,
respectively, using an Odyssey Fc Imaging system (LI-COR
Biosciences). Images were captured and analyzed using LI-COR
software Image Studio Lite v.5.x (LI-COR Biosciences).
For comparative studies of the effects of
ZINC4085554 on collagen I stimulated FAK-Tyr-397 phosphorylation in
SW620 cells, the dishes were coated with 1% heat inactivated BSA or
collagen I.
For the co-IP studies, cells were transfected using
Lipofectamine™ 2000 transfection reagent (Thermo Fisher Scientific,
Inc.) with 2 µg control hemagglutinin (HA), Tag-plasmid DNA
(pcDNA3; Addgene, Inc., Cambridge, MA, USA) and HA-FAK plasmid DNA
construct (HA-FAKpcDNA3; kindly donated by Dr. Guan; Department of
Cancer Biology, University of Cincinnati College of Medicine,
Cincinnati, OH, USA) to overexpress FAK (PTK2) tagged with HA.
After 72 h transfection, these cells were treated with vehicle
(0.1% DMSO) and 50 µM ZINC4085554 in suspension and subjected to
ambient and 15 mmHg pressure for 30 min in 1% heat inactivated BSA
coated dishes at 37°C in 5% CO2. These cells were then
lysed in non-denaturing lysis buffer (as aforementioned without
SDS). A total of 1–2 mg total protein was used for the
immunoprecipitation. The pre-cleared total protein was incubated
with anti-HA antibody (1:200; cat. no. 05-904; mouse monoclonal;
Merck KGaA) at 4°C overnight. Normal mouse IgG (1:200; cat. no.
SC-2025; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) was used
as a control antibody for the co-IP. Protein G plus/protein A
agarose suspension beads (Merck KGaA) were added to the lysate
containing the anti-HA antibody (1:200; cat. no. 05-904, mouse
monoclonal; Merck KGaA) and incubated for additional 2 h at 4°C.
The beads were washed three times with the same lysis buffer and
the protein was eluted in 6X Laemmli buffer (Morganville
Scientific, Morganville, NJ, USA). The eluted protein samples from
the original 1–2 mg of protein were resolved by 10% SDS-PAGE,
transferred onto nitrocellulose membranes, and immunoblotted with
anti-HA antibody (1:1,000; cat. no. 3724 (C29F4); rabbit; Cell
Signaling Technology, Inc.) and/or anti-Akt1 antibody (1:1,000;
cat. no. 2967 (2H10); mouse; Cell Signaling Technology, Inc.). The
total lysates loaded on gel and western blotted for HA-FAK and Akt1
served as input control for the co-IP. Densitometric quantitation
was performed with Kodak Scientific Imaging Systems 1D v.3.5.4 or
LI-COR imaging software (LI-COR Biosciences).
Surgical wound adhesion assay
This assay was designed to study cancer cell
adhesion, which is the first step in cancer recurrence at surgical
wounds (2). We previously
extensively validated this assay (4,24,31). A
total of 24 male mice (8–10 week old) were included in this study.
Mice were housed in temperature-controlled rooms with 12 h light
and 12 h dark cycle at 22–24°C and 50–60% humidity. All animals had
free access food and water. As the animals were sacrificed
post-adhesion and the non-adherent cells were washed away, wound
healing was not studied in these animals.
The present study was approved by the University of
North Dakota institutional animal use committee, and performed
using a technique previously described (4,24). SW620
cells were treated with DMSO or 50 µM ZINC4085554 at 37°C for 1 h
in an incubator. Following incubation, cells (~2 million/well of a
24-well plate) were trypsinized with 1 ml of 0.05% trypsin,
neutralized with 1 ml of culture medium containing 10% FBS, and
labeled with 50 µM Tag-it dye in PBS at 37°C for 20 min in
suspension (Tag-it Violet Proliferation Cell Tracking Dye;
Biolegend, Inc.). PBS containing 5% FBS was added to quench the
excess dye and then cells were pelleted and suspended in complete
L15 media. The suspended cells were treated with DMSO or 50 µM
ZINC4085554 and maintained at ambient or 15 mmHg increased
extracellular pressure as aforementioned. Following pressure
treatment, the cells were collected and counted. C57BL/6 mice were
anesthetized intraperitoneally using ketamine (100 mg/kg), xylazine
(10 mg/kg) and aceproject maleate (3 mg/kg) cocktail. A small
incision was made at the thigh and armpit region. The 400,000 cells
in 30 µl PBS were seeded for each group in these surgical wounds
and permitted to adhere for 30 min. The non-adherent cells were
washed away with 50 µl PBS five times. The mice were then
euthanized by cervical dislocation and the wounds were completely
excised along with a margin of unwounded tissue. The excised tissue
was digested using type I collagenase (Thermo Fisher Scientific,
Inc.) and a single cell suspension was prepared. These cells were
fixed using a FoxP3/Transcription factor fixation/permeabilization
kit from eBioscience (Thermo Fisher Scientific, Inc.) at 4°C for 30
min and acquired using a LSRII flow cytometer (BD Biosciences, San
Jose, CA, USA), gating so as to count only the Tag-it-labeled cells
that had been adherent to the surgical wounds at time of sacrifice.
No fluorophore kit or antibody was required since cells could be
detected by the Tag-it label. FlowJo software v.10 (FlowJo LLC,
Ashland, OR, USA) was used to analyze the data. It should be noted
that eBioscience Foxp3/Transcription factor
fixation/permeabilization concentrate and diluent is the name of a
fixation/permeabilization buffer (eBioscience; Thermo Fisher
Scientific, Inc.). Although originally used to investigate Foxp3,
it has a general purpose as a fixation permeabilization buffer and
it was determined to be beneficial to permeabilize digested
surgical wounds so that flow cytometry can be conducted to
quantitate pre-labeled adherent cancer cells within the wound. The
use of this buffer for flow cytometry has been discussed previously
(32,33).
Statistical analysis
All assays were performed within the linear range.
Data was normalized against the vehicle (0.1% DMSO)-treated ambient
pressure condition, and represented as the mean ± standard error of
the mean. SigmaPlot v.13 software (Systat Software, Inc., San Jose,
CA, USA) was used for statistical analysis. Unpaired student's
t-test was used to analyze the data. P<0.05 was considered to
indicate a statistically significant difference.
Results
ZINC4085554 inhibits
pressure-stimulated adhesion of SW620 cells
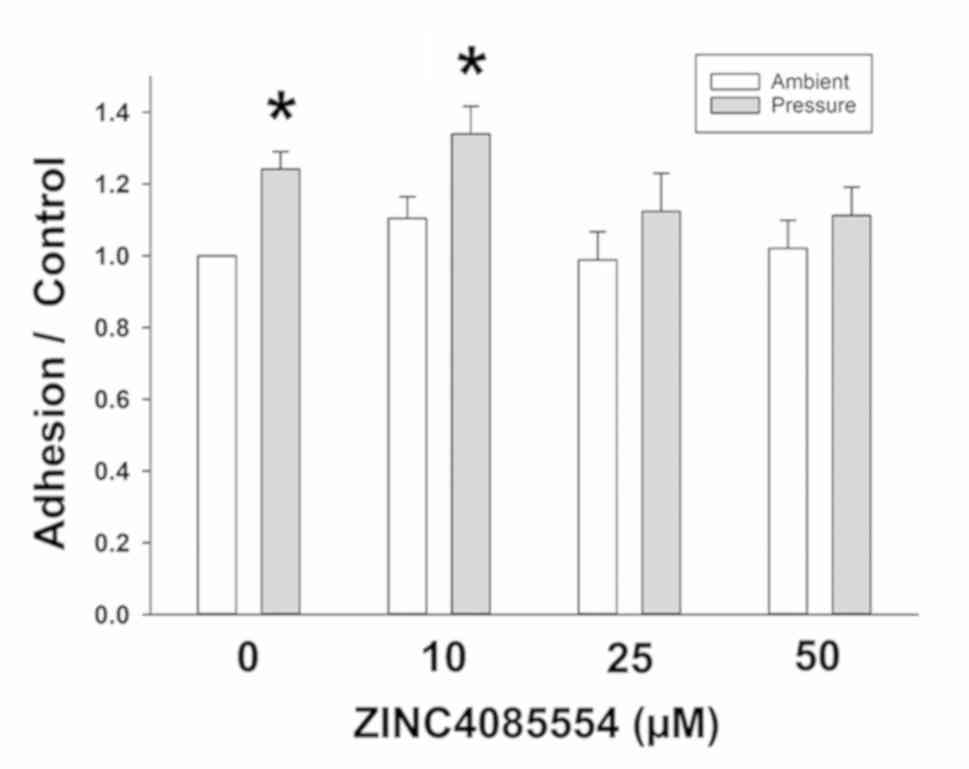
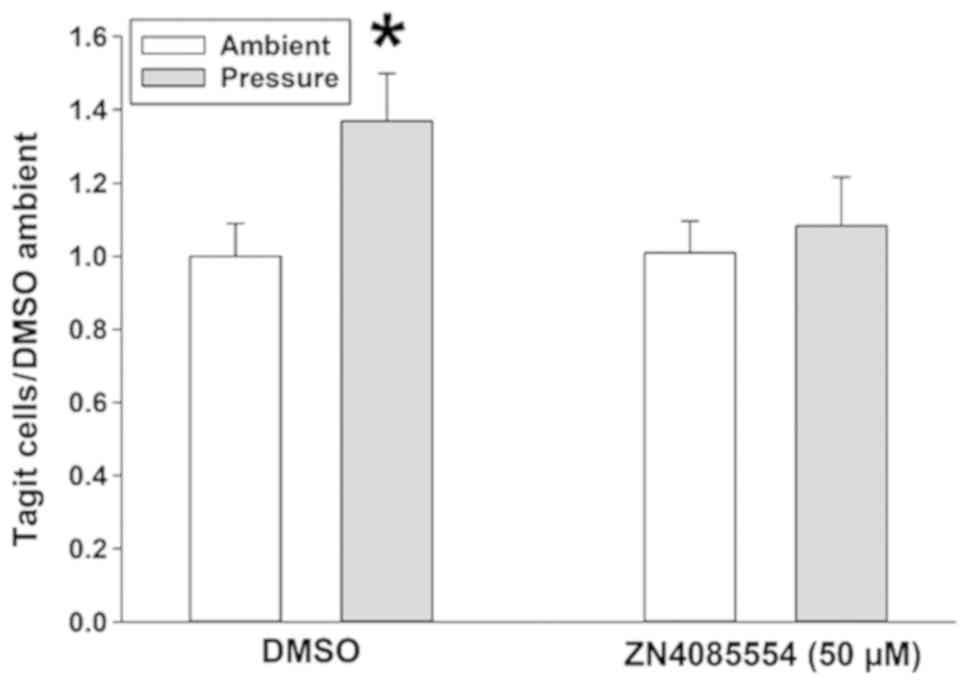
The present study first confirmed our previous cell
adhesion results with ZINC4085554. The data revealed that 15 mmHg
of pressure increases 24.1±4.9% of the adhesion vs. ambient
pressure in DMSO-treated SW620 cells. ZINC4085554 at 10 µM did not
affect the adhesion of SW620 cells at ambient pressure or prevent
the stimulation of adhesion by 15 mmHg extracellular pressure
(33.9±8.6% increase in ambient vs. pressure). In contrast, at 25
µM, an attenuation of the pressure effect was observed
(12.3±10.6%), and at 50 µM the pressure effect on adhesion was
blocked further (11.2±7.9%), reducing adhesion compared with that
observed at ambient pressure (Fig.
1). Therefore, although an effect was observed at 25 µM, 50 µM
was selected for the remainder of this investigation due to its
increased potency.
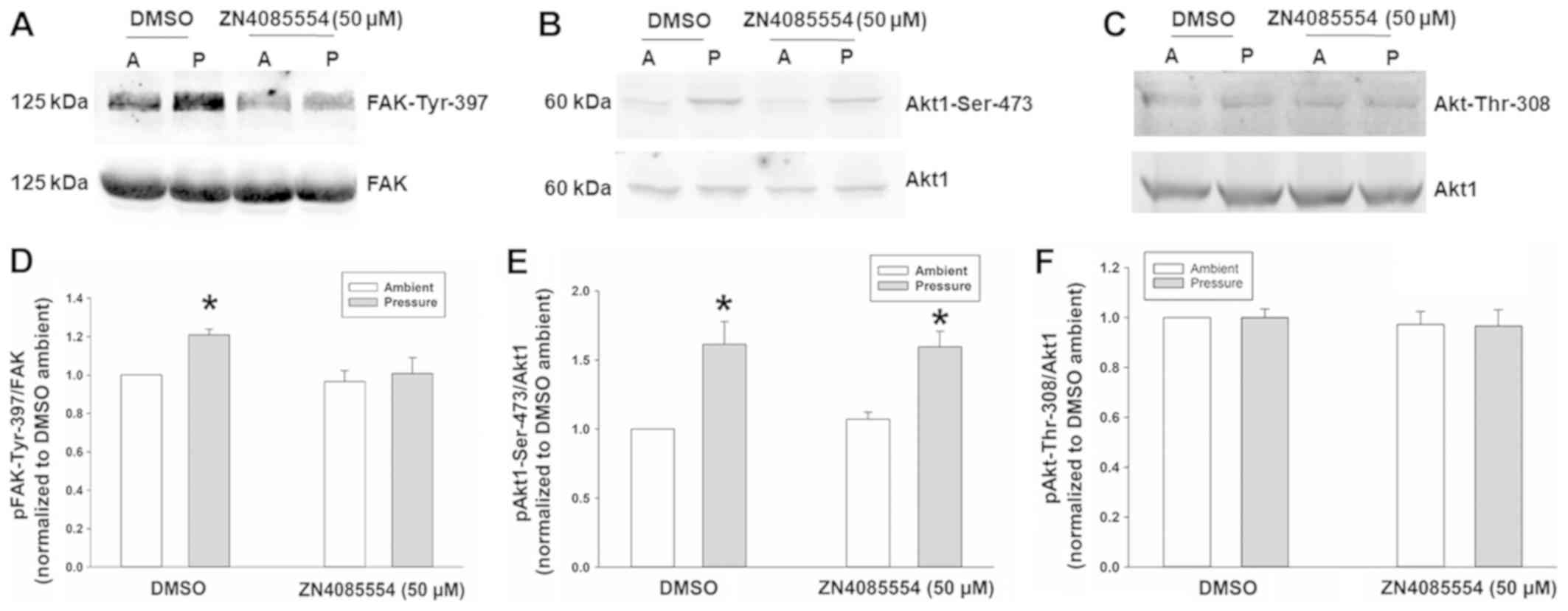
Pressure stimulates FAK-Tyr-397
phosphorylation yet not Akt1-Ser-473, and is inhibited by
ZINC4085554, while Akt-Thr-308 remains unaffected by pressure
and/or ZINC4085554
It has previously been reported that Akt1
phosphorylation at Ser-473 (but not at Thr-308) precedes and is
required for FAK-Tyr-397 phosphorylation in the stimulation of
cancer cell adhesion by pressure (16,28),
although FAK-Tyr-397 phosphorylation usually does not require Akt1
in other settings (34). The present
study therefore examined the effect of 50 µM ZINC4085554 on FAK and
Akt1 phosphorylation. There was a FAK-Tyr-397 increase of 18.7±3.2%
in pressure-stimulated SW620 cells (Fig.
2A and B) and a 71.2±16.9% increase in Akt1-Ser-473
phosphorylation (Fig. 2C and D) in
comparison to ambient pressure SW620 cells treated with the DMSO
vehicle. Consistent with our previous findings, the Akt-Thr-308
remains unaltered by pressure (16)
or ZINC4085554 (Fig. 2E and F).
ZINC4085554 did not affect basal FAK-Tyr-397 or Akt1
phosphorylation at ambient pressure. However, ZINC4085554 did
inhibit the stimulation of FAK-Tyr-397 phosphorylation at 15 mmHg,
yet did not affect pressure-stimulated Akt1 phosphorylation at
Ser-473 (Fig. 2), indicating that
ZINC4085554 inhibits FAK-Tyr-397 phosphorylation downstream of
Akt1.
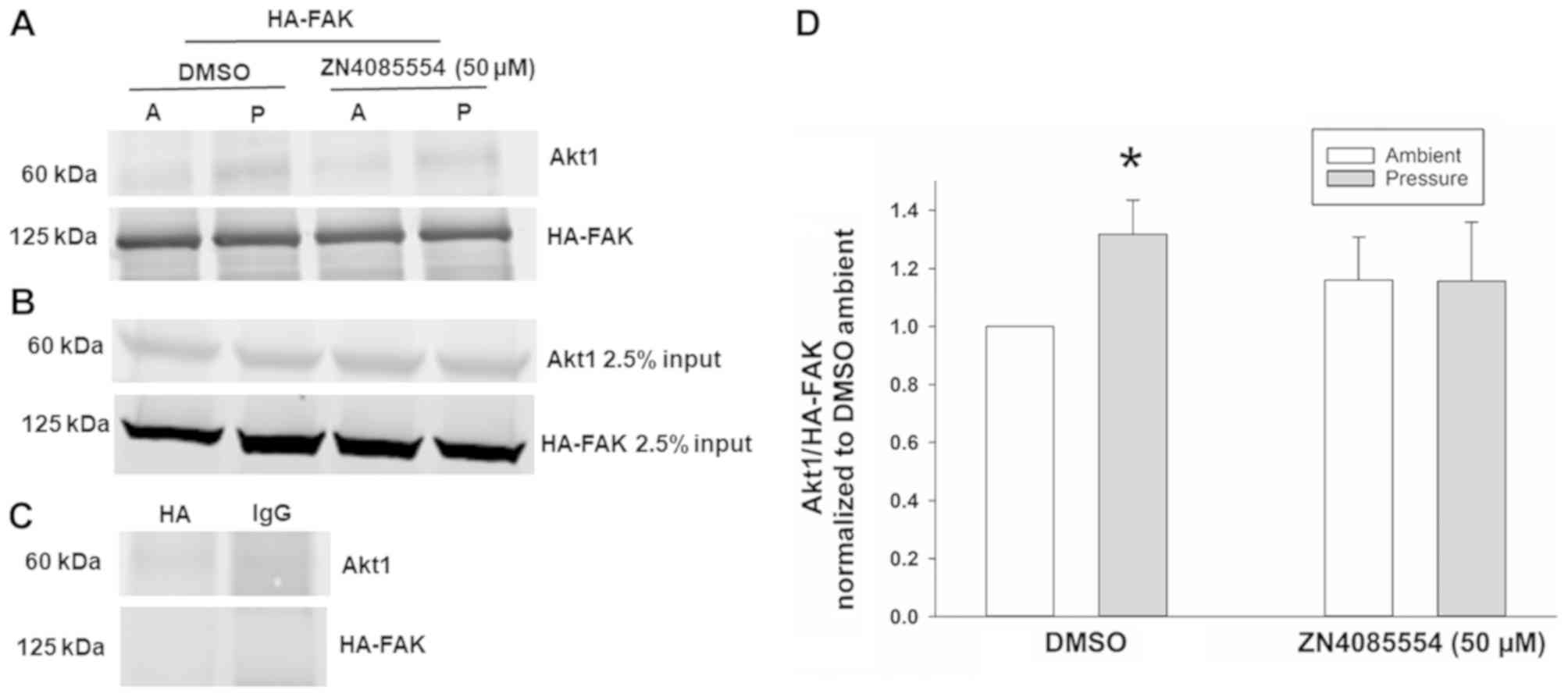
ZINC4085554 inhibits the
pressure-stimulated interaction of FAK and Akt1
As ZINC4085554 may block pressure-induced
FAK-Tyr-397 phosphorylation without affecting upstream Akt1
activation at Ser-473, the present study then examined the effects
of ZINC4085554 on interactions of FAK-Akt1 following exposure to
increased extracellular pressure. Co-IP studies demonstrated a
29.5±13.2% increase in the pull-down fractions of Akt1 using HA-FAK
vs. ambient DMSO in cells treated with DMSO at 15 mmHg increased
pressure. In contrast, in SW620 cells treated with 50 µM
ZINC4085554 and exposed to 15 mmHg increased pressure did not
stimulate the increased pull-down of Akt1 using HA-FAK; however,
yielded Akt1 pull-down similar to that observed at ambient pressure
(Fig. 3). This indicates that
ZINC4085554 interferes with the pressure-stimulated interaction of
FAK-Akt1.
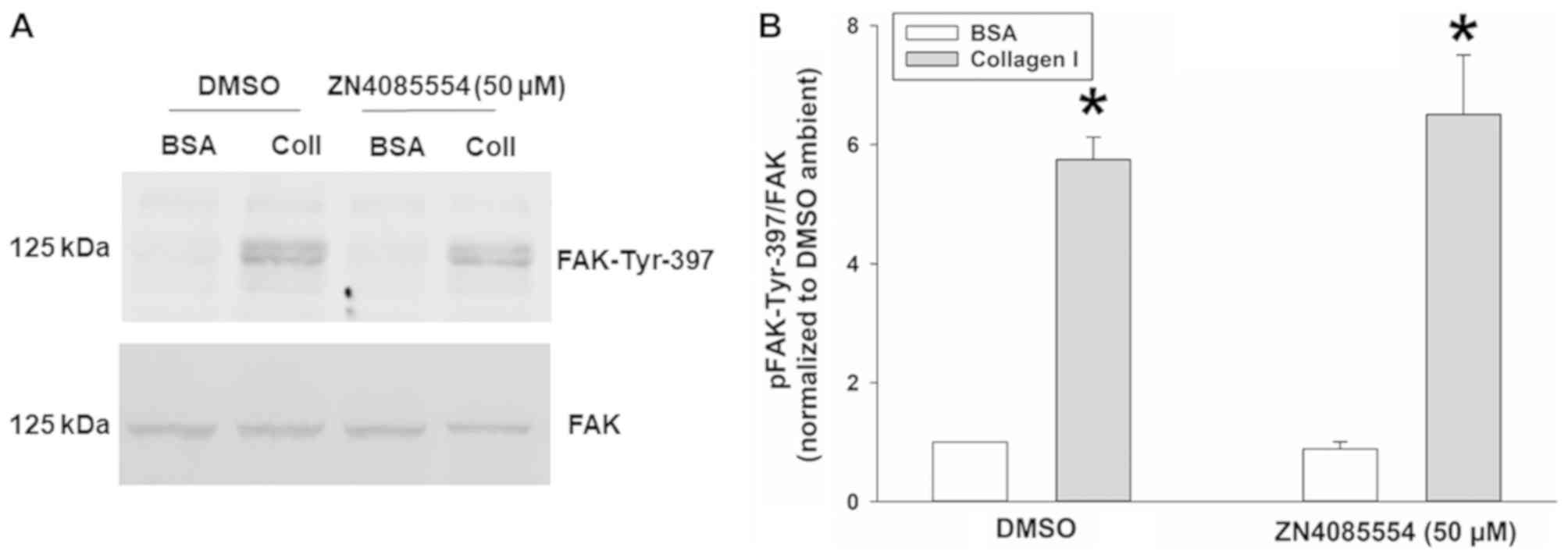
Collagen I stimulated FAK-Tyr-397
phosphorylation is unaffected by treatment with ZINC4085554
Although the present study focused on the
stimulation of FAK-Tyr-397 phosphorylation by increased
extracellular pressure in suspended cancer cells prior to adhesion
to a matrix substrate, FAK phosphorylation is much more commonly
studied in epithelial cells in response to adhesion (15,35–39).
This canonical pathway involves integrin heterodimer engagement
with extracellular matrix proteins, and shifts of integrin
conformation across the cell membrane that result in FAK
phosphorylation within the focal adhesion complex (36). This pathway does not require Akt for
FAK phosphorylation (34), and the
reason the present study focused on inhibiting Akt-FAK interaction
is that an agent that specifically blocks pressure-induced Akt-FAK
interaction should prevent pressure-induced downstream FAK
activation in cancer cells without interfering with the more common
FAK activation in other cells and tissues that is engendered by
cell-matrix interactions. Therefore, the present study sought to
address the specificity of the effect of ZINC4085554 on
pressure-stimulated FAK phosphorylation.
The effects of ZINC4085554 on pressure-stimulated
FAK-Tyr-397 phosphorylation were compared with the induction of
FAK-Tyr-397 phosphorylation by adhesion to type I collagen by
examining the effect of ZINC4085554 on collagen I stimulated
FAK-Tyr-397 phosphorylation. DMSO-treated SW620 cells exhibited a
475.0±37.7% increase in FAK-Tyr-397 phosphorylation on collagen I
vs. BSA coated plates. The same effect was observed (550.8±99.7%
increase in FAK-Tyr-397 phosphorylation on collagen I vs. BSA
coated plates) even in the presence of 50 µM ZINC4085554 (Fig. 4). Consequently, collagen I induced
FAK-Tyr-397 phosphorylation was unaffected by ZINC4085554, in
contrast to the induction of FAK-Tyr-397 phosphorylation by
increased extracellular pressure.
ZINC4085554 inhibits the
pressure-stimulated surgical wound adhesion in mice models
The present study then sought to confirm the effect
of ZINC4085554 on the adhesion of SW620 cells to more biologically
complex intact tissues in the surgical wounds of mice. SW620 cells
treated with DMSO and/or ZINC4085554 at ambient or 15 mmHg
increased pressure were allowed to adhere to surgical wounds in
mice, prior to non-adherent cells were washed away, in order to
mimic the cancer cell adhesion that occurs in surgical fields
during cancer resection, in which shed cancer cells initially may
adhere locally prior to subsequent irrigation of the surgical
field. An 84.4±42.1% increase was observed in the wound adhesion of
the DMSO-treated SW620 cells exposed to 15 mmHg pressure vs. cells
maintained at ambient pressure. In contrast, the cells treated with
50 µM ZINC4085554 did not exhibit significantly increased adhesion
following stimulation by 15 mmHg increased pressure yet rather
displayed adhesion to surgical wounds similar to that observed by
cells exposed to ambient pressure (Fig.
5).
Discussion
Mechanical forces stimulate the cancer cell
adhesiveness required for metastasis (8,40).
FAK-Akt1 interaction serves a crucial role in 15 mmHg pressure
above ambient stimulated adhesion of colon cancer cells (24). The interaction between Akt1 and FAK
represents an opportune target for intervention, which unlike
conventional FAK or Akt inhibitors, may specifically ablate the
mechanosensitive signaling pathways that promotes cancer cell
adhesion without interfering with other FAK or Akt signaling in
host tissues. Building upon a preliminary in silico
screening and confirmation that at least two of the candidate
molecules may inhibit cancer cell adhesion (27), the present study demonstrated that
ZINC4085554 inhibits pressure-stimulated FAK activation; however,
it does not prevent the upstream activation of Akt1 by increased
extracellular pressure or the activation of FAK by more
conventional outside-in signaling induced by adhesion to type I
collagen, a dominant interstitial matrix component (41). Instead, ZINC4085554 attenuated
FAK-Akt1 interaction. Lastly, this molecule inhibited the 15 mmHg
pressure-stimulated adhesion of colon cancer cells in the surgical
wounds of mice.
FAK activation is one of the central steps in the
adhesive processes of cancer cells, and also critical to numerous
other signaling pathways, including cell migration, response to
cyclic strain and response to growth factors (15,18,42). A
wide variety of scaffolding proteins, adapter molecules, signaling
messengers and transmembrane proteins regulate FAK while FAK also
regulates its own activation and deactivation via dimerization and
auto-phosphorylation (35,43,44).
While FAK activation in response to extracellular forces requires
Akt (17,37,45), FAK
activation in other contexts occurs independently of Akt (34), and Akt may or may not be activated
downstream of FAK, depending on the context (34,38,46). The
phosphorylation of Akt1 at Thr-308 and Ser-473 has previously been
reported to potentiate its full activation. PDK1 phosphorylates
Akt1 at Thr-308 in the catalytic region or activation loop and this
phosphorylation does increase Akt1 activity (47). However, mTORC2 phosphorylation of
Akt1 at Ser-473, in the C-terminal hydrophobic motif, is
hypothesized to be required for Akt1 maximal activation (47). The literature indicates that certain
signaling pathways/proteins exclusively phosphorylate each site for
Akt activation. For instance, tumor necrosis factor-α exclusively
phosphorylates Akt at Akt-Ser-473, but not at Thr-308 (48). Similarly, glutamate-exposed neurons
exhibit Akt phosphorylation at Akt-Ser-473, yet not at Thr-308
(49). We have previously reported
that exposure to increased extracellular pressure also results in
the phosphorylation of Akt-Ser-473, but not of Akt-Thr-308
(16). The results of the present
study are consistent with the previous observation of the absence
of Akt-Thr-308 phosphorylation in response to extracellular
pressure.
The observation that ZINC4085554 does not prevent
FAK activation in response to adhesion is not unexpected. FAK is
activated in response to a diverse range of stimuli, including
cell-matrix interactions (36,39) and
transactivation by the epidermal growth factor receptor (42,50).
These canonical pathways do not require Akt to interact with FAK
(21,34,51,52). For
instance, adhesion itself engages integrin heterodimers with
moieties on the extracellular matrix, inducing a conformational
shift in the β-1 integrin subunit that transmits across the cell
membrane and is eventually associated with FAK-Tyr-397
phosphorylation without involving Akt, and this pathway has been
extensively reviewed (21,34,36,51). In
contrast, the activation of FAK in cancer cells in response to
extracellular pressure requires a preliminary FAK-Akt interaction,
which makes FAK-Akt interaction an attractive target to
specifically block pressure-induced FAK activation in cancer cells
without interfering with other FAK signaling that may be notable in
other tissues and organs within the patient (17,22–24,27). The
results of the present study are consistent with this model, since
ZINC4085554 prevents FAK-Tyr-397 phosphorylation in suspended cells
in response to pressure but does not prevent the FAK-Tyr-397
phosphorylation that occurs post- and in response to adhesion to a
matrix.
Indeed, these results further indicate that
ZINC4085554 is neither a conventional FAK inhibitor as it permits
FAK activation in response to adhesion, nor a conventional Akt
inhibitor, as it allows Akt activation at Ser-473 in response to
increased extracellular pressure. ZINC4085554 may interfere with
the binding of Akt1 to FAK after Akt1 has been activated in the
setting of increased extracellular pressure, and thereby more
specifically prevents FAK activation by increased extracellular
pressure while allowing FAK activation by a different pathway. This
indicates the possibility that molecules based on ZINC4085554 may
exhibit less off-target specificity than conventional FAK or Akt
inhibitors by allowing other FAK and Akt signaling to continue
unabated. It is notable that physical force effects on other cell
types may invoke different signaling pathways. For instance,
increased extracellular pressure consistent with the edema in
inflamed tissues stimulates macrophage phagocytosis by a pathway
that involves the activation of Akt2 rather than Akt1, and the
inhibition of FAK rather than its stimulation (53).
The concept that ZINC4085554 inhibits adhesion by
interfering with FAK-Akt1 interaction would be consistent with our
original hypothesis that ZINC4085554 resembles the subdomain of FAK
to which Akt1 binds. ZINC4085554 was identified by in silico
screening as a small molecule resembling a two site mutant peptide
derived from a helical subdomain in FAK that was revealed to
compete with FAK for binding to Akt1 (27). As FAK itself dimerizes, it will be
notable to determine in future studies whether ZINC4085554 binds to
FAK itself, thereby occupying the Akt1 binding site, or whether
ZINC4085554 binds to Akt1, occupying the FAK binding site. This
awaits further study.
The present study further confirmed that ZINC4085554
not only inhibits cancer cell adhesion to a type I collagen
substrate in vitro but also to the biological tissues in a
murine surgical wound. This was notable as metastatic adhesion must
occur to a complex tissue in vivo offering multiple binding
sites beyond those that are present in type I collagen (1,4). It has
previously been reported that increased extracellular pressure also
stimulates cancer cell adhesion to endothelial cells (54). Although the increase in adherent cell
numbers in vitro and to the mouse wounds with extracellular
pressure and its blockade by ZINC4085554 may seem small in
magnitude, we previously reported that such small changes in cancer
cell adhesion translate to substantial changes in subsequent tumor
development and survival in mouse models (4,24,31,55).
This may be because tumor development is a stochastic process that
requires tumor cells to not only adhere, but to subsequently
proliferate and elude host immune surveillance (1,3). Others
examined changes of similar magnitude in FAK activation and
adhesion in other contexts (56–59). For
instance, Hic-5−/−;PyMT tumor cells present reduced
FAK-Tyr-397 phosphorylation compared to Hic-5+/+;PyMT
cells (56).
ZINC4085554 may not be a therapeutic agent in
itself, as the concentration required for efficacy is relatively
high and its dinitrobenzene group may raise chemical safety issues
(27). However, these studies
represent proof of principle that a small molecule can be developed
to inhibit cancer cell adhesiveness by interfering with FAK-Akt1
interaction. It may be easier to modify this molecule or similar
molecules for therapeutic testing than to develop an approach based
upon the previous demonstration that a seven amino acid FAK-derived
peptide can reduce adhesiveness (24) since the latter would necessitate
solving problems of delivery and stability.
Postoperative tumor recurrence can reflect the
emergence of previously unrecognized and therefore unresected
tumors that had previously metastasized, yet not grown sufficiently
for detection or the dissemination of new tumor cells at the time
of surgery that implants and develop into recurrent disease. Such
tumor implantation can occur in surgical wounds, or other aspects
of the surgical site such as the peritoneum, or distant organs via
lymphatic or circulatory dissemination (4,8,60–67).
Although there may be differences in the pathophysiology and
mechanics underlying local tumor dissemination to wounds to the
peritoneum and dissemination through the circulation, it has
previously been demonstrated that increased extracellular pressure
promotes wound implantation and peritoneal metastasis (4). Shear stress occurs during circulation,
and also activates the same pathway (68). Wound implantation is easier to
investigate from a methodological standpoint, and was consequently
selected for the first attempt to validate the potential effects of
ZINC4085554 in vivo. However, further study is warranted to
extend these results to the effects of ZINC4085554 on tumor
dissemination via the lymphatics or circulation.
It is not always possible to distinguish metastasis
detected postoperatively, which was simply not detected
preoperatively from postoperative metastasis that reflects surgical
dissemination. However, the increased shedding of tumor cells into
the portal and peripheral circulation following colon cancer
resection is well documented (67,69), and
the presence of such tumor cells in the circulation is an adverse
prognostic factor (61,62) even if it is not possible determine
which recurrences reflect perioperative shedding and which reflect
preexisting disease. Additionally, postoperative recurrence of the
tumor within the surgically-created wound may represent a
consequence of surgery itself. The pivotal New England Journal of
Medicine trial comparing laparoscopic and open colectomy for cancer
(60) documented a 0.2% incidence of
wound recurrence in the open surgical group and a 0.5% incidence of
wound recurrence following laparoscopic surgery involving increased
intraperitoneal pressure, although the study did not have
sufficient sample size to demonstrate a statistical difference in
wound recurrence between the two groups. Postoperative wound
recurrence is a consequence of surgery in the majority of cases, as
the wound was not present preoperatively, and a previous study
described an increased incidence of wound metastasis in increased
intraperitoneal pressure surgical procedures modeling laparoscopic
surgery in animals (64). This
effect disappears when so-called ‘gasless laparoscopy’ models using
traction (negative pressure) to expand the abdomen rather than
pressure to inflate it are used (63). While much of this work has been
performed on colorectal cancer, it has now become clear that
laparoscopic resections for cervical cancer are more prone to
recurrence and no longer represent the standard of care (65,66).
Therefore, while it may not be possible to distinguish preoperative
undetected disease from perioperative tumor spread in any specific
patient, it may not to conclude that at least some postoperative
metastasis does unfortunately reflect the effects of surgery.
Although much work must still be done before human trials could be
contemplated for this approach, these studies are aimed at
understanding and blocking the pathways by which physical forces
engendered either during surgery or during passage through the
lymphatics or circulation in the non-surgical patient can activate
the adhesion of metastasizing tumor cells, in the hope that this
approach may ultimately inhibit tumor metastasis.
In conclusion, the present study demonstrated that a
small molecule that mimics a subdomain of FAK can inhibit cancer
cell adhesiveness in vitro and in live murine tissue by
interfering with Akt1 binding to FAK. As other FAK and Akt1
signaling were not interfered with, such a molecule may well be
used therapeutically at higher concentrations than conventional FAK
or Akt inhibitors as FAK or Akt signaling would not be expected to
be interfered with. Molecules such as ZINC4085554 therefore seem
promising targets for further investigation, while these results
also further validate our initial in silico structure-based
screening strategy by suggesting that ZINC4085554 works by a
mechanism consistent with that which was targeted by that
strategy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SKM and MDB designed and planned the study. SKM
performed all the experiments. EEVD acquired and analyzed the flow
cytometry data. SKM and MDB wrote the manuscript and EEVD reviewed
it before submission.
Ethics approval and consent to
participate
All animal studies were approved by the University
of North Dakota Institutional Animal Use and Care Committee.
Patient consent for publication
Not applicable.
Competing interests
The University of North Dakota and Michigan State
University have jointly applied for a patent on “inhibiting FAK-AKT
interaction to inhibit metastasis” on which MDB is named as a
co-inventor. The authors declare that they have no competing
interests.
References
|
1
|
Lambert AW, Pattabiraman DR and Weinberg
RA: Emerging Biological Principles of Metastasis. Cell.
168:670–691. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Craig DH and Basson MD: Biological impact
of mechanical stimuli on tumor metastasis. Cell Cycle. 8:828–831.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brooks SA, Lomax-Browne HJ, Carter TM,
Kinch CE and Hall DM: Molecular interactions in cancer cell
metastasis. Acta Histochem. 112:3–25. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Craig DH, Owen CR, Conway WC, Walsh MF,
Downey C and Basson MD: Colchicine inhibits pressure-induced tumor
cell implantation within surgical wounds and enhances tumor-free
survival in mice. J Clin Invest. 118:3170–3180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lafrenie RM, Buchanan MR and Orr FW:
Adhesion molecules and their role in cancer metastasis. Cell
Biophys. 23:3–89. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kavic SM and Basson MD: Environmental
factors of temperature, humidity, serum accumulation, and cell
seeding increase colon cancer cell adhesion in vitro, with partial
characterization of the serum component responsible for
pressure-stimulated adhesion. J Surg Res. 98:89–96. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wirtz D, Konstantopoulos K and Searson PC:
The physics of cancer: The role of physical interactions and
mechanical forces in metastasis. Nat Rev Cancer. 11:512–522. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Basson MD: An intracellular signal pathway
that regulates cancer cell adhesion in response to extracellular
forces. Cancer Res. 68:2–4. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Downey C, Alwan K, Thamilselvan V, Zhang
L, Jiang Y, Rishi AK and Basson MD: Pressure stimulates breast
cancer cell adhesion independently of cell cycle and apoptosis
regulatory protein (CARP)-1 regulation of focal adhesion kinase. Am
J Surg. 192:631–635. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Conway WC, Van der Voort van Zyp J,
Thamilselvan V, Walsh MF, Crowe DL and Basson MD: Paxillin
modulates squamous cancer cell adhesion and is important in
pressure-augmented adhesion. J Cell Biochem. 98:1507–1516. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Perry BC, Wang S and Basson MD:
Extracellular pressure stimulates adhesion of sarcoma cells via
activation of focal adhesion kinase and Akt. Am J Surg.
200:610–614. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Basson MD, Yu CF, Herden-Kirchoff O,
Ellermeier M, Sanders MA, Merrell RC and Sumpio BE: Effects of
increased ambient pressure on colon cancer cell adhesion. J Cell
Biochem. 78:47–61. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Daskalakis M, Scheffel O and Weiner RA:
High flow insufflation for the maintenance of the pneumoperitoneum
during bariatric surgery. Obes Facts. 2 (Suppl 1):37–40. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thamilselvan V and Basson MD: The role of
the cytoskeleton in differentially regulating pressure-mediated
effects on malignant colonocyte focal adhesion signaling and cell
adhesion. Carcinogenesis. 26:1687–1697. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Parsons JT: Focal adhesion kinase: The
first ten years. J Cell Sci. 116:1409–1416. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang S and Basson MD: Integrin-linked
kinase: Α multi-functional regulator modulating extracellular
pressure-stimulated cancer cell adhesion through focal adhesion
kinase and AKT. Cell Oncol. 31:273–289. 2009.PubMed/NCBI
|
|
17
|
Wang S and Basson MD: Akt directly
regulates focal adhesion kinase through association and serine
phosphorylation: Implication for pressure-induced colon cancer
metastasis. Am J Physiol Cell Physiol. 300:C657–C670. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Golubovskaya VM: Targeting FAK in human
cancer: From finding to first clinical trials. Front Biosci.
19:687–706. 2014. View
Article : Google Scholar
|
|
19
|
Lv PC, Jiang AQ, Zhang WM and Zhu HL: FAK
inhibitors in Cancer, a patent review. Expert Opin Ther Pat.
28:139–145. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schultze A and Fiedler W: Therapeutic
potential and limitations of new FAK inhibitors in the treatment of
cancer. Expert Opin Investig Drugs. 19:777–788. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sulzmaier FJ, Jean C and Schlaepfer DD:
FAK in cancer: Mechanistic findings and clinical applications. Nat
Rev Cancer. 14:598–610. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Basson MD, Zeng B and Wang S: Akt1 binds
focal adhesion kinase via the Akt1 kinase domain independently of
the pleckstrin homology domain. J Physiol Pharmacol. 66:701–709.
2015.PubMed/NCBI
|
|
23
|
Basson MD, Zeng B and Wang S: The
C-terminal region of the focal adhesion kinase F1 domain binds Akt1
and inhibits pressure-induced cell adhesion. J Physiol Pharmacol.
68:375–383. 2017.PubMed/NCBI
|
|
24
|
Zeng B, Devadoss D, Wang S, Vomhof-DeKrey
EE, Kuhn LA and Basson MD: Inhibition of pressure-activated cancer
cell adhesion by FAK-derived peptides. Oncotarget. 8:98051–98067.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marqus S, Pirogova E and Piva TJ:
Evaluation of the use of therapeutic peptides for cancer treatment.
J Biomed Sci. 24:212017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Otvos L Jr and Wade JD: Current challenges
in peptide-based drug discovery. Front Chem. 2:622014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Raschka S, More SK, Devadoss D, Zeng B,
Kuhn LA and Basson MD: Identification of potential small-molecule
protein-protein inhibitors of cancer metastasis by 3D epitope-based
computational screening. J Physiol Pharmacol. 69:doi:
10.26402/jpp.2018.2.11. PubMed/NCBI
|
|
28
|
Thamilselvan V, Craig DH and Basson MD:
FAK association with multiple signal proteins mediates
pressure-induced colon cancer cell adhesion via a Src-dependent
PI3K/Akt pathway. FASEB J. 21:1730–1741. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Allain F, Vanpouille C, Carpentier M,
Slomianny MC, Durieux S and Spik G: Interaction with
glycosaminoglycans is required for cyclophilin B to trigger
integrin-mediated adhesion of peripheral blood T lymphocytes to
extracellular matrix. Proc Natl Acad Sci USA. 99:2714–2719. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Canavan HE, Cheng X, Graham DJ, Ratner BD
and Castner DG: Cell sheet detachment affects the extracellular
matrix: A surface science study comparing thermal liftoff,
enzymatic, and mechanical methods. J Biomed Mater Res A. 75:1–13.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Craig DH, Downey C and Basson MD:
SiRNA-mediated reduction of alpha-actinin-1 inhibits
pressure-induced murine tumor cell wound implantation and enhances
tumor-free survival. Neoplasia. 10:217–222. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Claycombe KJ, Vomhof-DeKrey EE, Garcia R,
Johnson WT, Uthus E and Roemmich JN: Decreased beige adipocyte
number and mitochondrial respiration coincide with increased
histone methyl transferase (G9a) and reduced FGF21 gene expression
in Sprague-Dawley rats fed prenatal low protein and postnatal
high-fat diets. J Nutr Biochem. 31:113–121. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vomhof-DeKrey E, Darland D, Ghribi O,
Bundy A, Roemmich J and Claycombe K: Maternal low protein diet
leads to placental angiogenic compensation via dysregulated M1/M2
macrophages and TNFα expression in Sprague-Dawley rats. J Reprod
Immunol. 118:9–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Velling T, Nilsson S, Stefansson A and
Johansson S: beta1-Integrins induce phosphorylation of Akt on
serine 473 independently of focal adhesion kinase and Src family
kinases. EMBO Rep. 5:901–905. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huveneers S and Danen EH: Adhesion
signaling - crosstalk between integrins, Src and Rho. J Cell Sci.
122:1059–1069. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hynes RO: Integrins: Bidirectional,
allosteric signaling machines. Cell. 110:673–687. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Turecková J, Vojtechová M, Krausová M,
Sloncová E and Korínek V: Focal adhesion kinase functions as an akt
downstream target in migration of colorectal cancer cells. Transl
Oncol. 2:281–290. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xia H, Nho RS, Kahm J, Kleidon J and Henke
CA: Focal adhesion kinase is upstream of phosphatidylinositol
3-kinase/Akt in regulating fibroblast survival in response to
contraction of type I collagen matrices via a beta 1 integrin
viability signaling pathway. J Biol Chem. 279:33024–33034. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao J and Guan JL: Signal transduction by
focal adhesion kinase in cancer. Cancer Metastasis Rev. 28:35–49.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gayer CP and Basson MD: The effects of
mechanical forces on intestinal physiology and pathology. Cell
Signal. 21:1237–1244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vleming LJ, Baelde JJ, Westendorp RG, Daha
MR, van Es LA and Bruijn JA: Progression of chronic renal disease
in humans is associated with the deposition of basement membrane
components and decorin in the interstitial extracellular matrix.
Clin Nephrol. 44:211–219. 1995.PubMed/NCBI
|
|
42
|
Tomar A and Schlaepfer DD: A PAK-activated
linker for EGFR and FAK. Dev Cell. 18:170–172. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nagano M, Hoshino D, Koshikawa N, Akizawa
T and Seiki M: Turnover of focal adhesions and cancer cell
migration. Int J Cell Biol. 2012:3106162012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Schlaepfer DD, Hauck CR and Sieg DJ:
Signaling through focal adhesion kinase. Prog Biophys Mol Biol.
71:435–478. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Higuchi M, Kihara R, Okazaki T, Aoki I,
Suetsugu S and Gotoh Y: Akt1 promotes focal adhesion disassembly
and cell motility through phosphorylation of FAK in growth
factor-stimulated cells. J Cell Sci. 126:745–755. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao X and Guan JL: Focal adhesion kinase
and its signaling pathways in cell migration and angiogenesis. Adv
Drug Deliv Rev. 63:610–615. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Manning BD and Toker A: AKT/PKB Signaling:
Navigating the Network. Cell. 169:381–405. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
O'toole A, Moule SK, Lockyer PJ and
Halestrap AP: Tumour necrosis factor-alpha activation of protein
kinase B in WEHI-164 cells is accompanied by increased
phosphorylation of Ser473, but not Thr308. Biochem J. 359:119–127.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kitagawa K, Takasawa K, Kuwabara K, Sasaki
T, Tanaka S, Mabuchi T, Sugiura S, Omura-Matsuoka E, Matsumoto M
and Hori M: Differential Akt phosphorylation at Ser473 and Thr308
in cultured neurons after exposure to glutamate in rats. Neurosci
Lett. 333:187–190. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Salazar EP, Hunger-Glaser I and Rozengurt
E: Dissociation of focal adhesion kinase and paxillin tyrosine
phosphorylation induced by bombesin and lysophosphatidic acid from
epidermal growth factor receptor transactivation in Swiss 3T3
cells. J Cell Physiol. 194:314–324. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Mitra SK, Hanson DA and Schlaepfer DD:
Focal adhesion kinase: In command and control of cell motility. Nat
Rev Mol Cell Biol. 6:56–68. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mitra SK and Schlaepfer DD:
Integrin-regulated FAK-Src signaling in normal and cancer cells.
Curr Opin Cell Biol. 18:516–523. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shiratsuchi H and Basson MD: Akt2, but not
Akt1 or Akt3 mediates pressure-stimulated serum-opsonized latex
bead phagocytosis through activating mTOR and p70 S6 kinase. J Cell
Biochem. 102:353–367. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Thamilselvan V and Basson MD: Pressure
activates colon cancer cell adhesion by inside-out focal adhesion
complex and actin cytoskeletal signaling. Gastroenterology.
126:8–18. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
van Zyp J, Conway WC, Craig DH, van Zyp N,
Thamilselvan V and Basson MD: Extracellular pressure stimulates
tumor cell adhesion in vitro by paxillin activation. Cancer Biol
Ther. 5:1169–1178. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Goreczny GJ, Ouderkirk-Pecone JL, Olson
EC, Krendel M and Turner CE: Hic-5 remodeling of the stromal matrix
promotes breast tumor progression. Oncogene. 36:2693–2703. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lee BY, Hochgräfe F, Lin HM, Castillo L,
Wu J, Raftery MJ, Martin Shreeve S, Horvath LG and Daly RJ:
Phosphoproteomic profiling identifies focal adhesion kinase as a
mediator of docetaxel resistance in castrate-resistant prostate
cancer. Mol Cancer Ther. 13:190–201. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Srinivas V, Datta SA, Ramakrishna T and
Rao CM: Studies on the alpha-crystallin target protein binding
sites: Sequential binding with two target proteins. Mol Vis.
7:114–119. 2001.PubMed/NCBI
|
|
59
|
Wang H, Zhu Y, Zhao M, Wu C, Zhang P, Tang
L, Zhang H, Chen X, Yang Y and Liu G: miRNA-29c suppresses lung
cancer cell adhesion to extracellular matrix and metastasis by
targeting integrin β1 and matrix metalloproteinase2 (MMP2). PLoS
One. 8:e701922013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Nelson H, Sargent DJ, Wieand HS, Fleshman
J, Anvari M, Stryker SJ, Beart RW Jr, Hellinger M, Flanagan R Jr,
Peters W, et al Clinical Outcomes of Surgical Therapy Study Group,
: A comparison of laparoscopically assisted and open colectomy for
colon cancer. N Engl J Med. 350:2050–2059. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Fujita S, Kudo N, Akasu T and Moriya Y:
Detection of cytokeratin 19 and 20 mRNA in peripheral and
mesenteric blood from colorectal cancer patients and their
prognosis. Int J Colorectal Dis. 16:141–146. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Guller U, Zajac P, Schnider A, Bösch B,
Vorburger S, Zuber M, Spagnoli GC, Oertli D, Maurer R, Metzger U,
et al: Disseminated single tumor cells as detected by real-time
quantitative polymerase chain reaction represent a prognostic
factor in patients undergoing surgery for colorectal cancer. Ann
Surg. 236:768–775; discussion 775–776. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ishida H, Hashimoto D, Takeuchi I,
Yokoyama M, Okita T and Hoshino T: Liver metastases are less
established after gasless laparoscopy than after carbon dioxide
pneumoperitoneum and laparotomy in a mouse model. Surg Endosc.
16:193–196. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lee SW, Whelan RL, Southall JC and Bessler
M: Abdominal wound tumor recurrence after open and
laparoscopic-assisted splenectomy in a murine model. Dis Colon
Rectum. 41:824–831. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Melamed A, Margul DJ, Chen L, Keating NL,
Del Carmen MG, Yang J, Seagle BL, Alexander A, Barber EL, Rice LW,
et al: Survival after Minimally Invasive Radical Hysterectomy for
Early-Stage Cervical Cancer. N Engl J Med. 379:1905–1914. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Ramirez PT, Frumovitz M, Pareja R, Lopez
A, Vieira M, Ribeiro R, Buda A, Yan X, Shuzhong Y, Chetty N, et al:
Minimally Invasive versus Abdominal Radical Hysterectomy for
Cervical Cancer. N Engl J Med. 379:1895–1904. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Umpleby HC, Fermor B, Symes MO and
Williamson RC: Viability of exfoliated colorectal carcinoma cells.
Br J Surg. 71:659–663. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Thamilselvan V, Patel A, van der Voort van
Zyp J and Basson MD: Colon cancer cell adhesion in response to Src
kinase activation and actin-cytoskeleton by non-laminar shear
stress. J Cell Biochem. 92:361–371. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Sugarbaker PH: Successful management of
microscopic residual disease in large bowel cancer. Cancer
Chemother Pharmacol. 43 (Suppl):S15–S25. 1999. View Article : Google Scholar : PubMed/NCBI
|