Introduction
Thyroid cancer is the most commonly diagnosed type
of head and neck cancer (1).
Differentiated thyroid cancer is the most common type of thyroid
cancer, which accounts for >95% of all cases (2). Due to increased use of diagnostic
imaging and surveillance techniques over the last 30 years, with
the exception of Africa, the incidence rates of thyroid cancer have
continued to increase worldwide, and it accounted for 3.4% of all
cancers in the USA in 2017 (3,4). The
B-Raf Proto-Oncogene, Serine/Threonine Kinase
(BRAF)V600E mutation is one of the most common mutations
observed in thyroid cancer.
Keratins are the intermediate filament (IF)-forming
proteins in epithelial cells and have been long and extensively
used as immunohistochemical markers for the pathological diagnosis
of tumors (5). The keratin 19
(KRT19) gene, located at chromosome17q21.2, encodes for the
cytokeratin (CK)19 protein, CK-19, which is an acidic protein type
I CK (6). KRT19 has been studied as
a prognostic marker and has been found to exert specific biological
roles in numerous types of cancer, including hepatocellular
carcinoma, pancreatic neuroendocrine tumors and potentially
HER2-positive breast cancer (7–10).
However, its role in thyroid cancer is currently unclear,
therefore, the aim of the present study was to examine the role of
KRT19 in thyroid cancer cell lines, and further explore the
potential mechanism of KRT19 overexpression. KRT19 expression was
observed to be higher in thyroid cancer samples when compared with
normal thyroid tissue samples and therefore, may function as a
potential oncogene that promotes thyroid cancer cell proliferation
and migration in vitro. In addition, the present study, to
the best of our knowledge, reported for the first time that the
BRAFV600E mutation may contribute to the oncogenic
function of KRT19. This suggests that KRT19 and
BRAFV600E may serve a role in epithelial-mesenchymal
transition (EMT).
Materials and methods
Data sources
The Cancer Genome Atlas (TCGA), Level 3 thyroid
cancer data were first downloaded from UCSC Xena (https://xenabrowser.net/datapages/). The TCGA
Thyroid Cancer (TCGA_THCA) dataset (ID,
TCGA.THCA.sampleMap/HiSeqV2; version 20171013) contained the
following information on 572 patients with thyroid cancer:
Tumor-Node-Metastasis (TNM) stage (11), age, grading, pathology type
(follicular subtype, classical subtype and tall cell subtype) and
other clinical parameters. The TCGA_THCA dataset was produced using
Illumina HiSeq (Illumina, Inc.), consisted of 59 normal thyroid
tissues and 505 cancer tissues, and was analyzed using the unit
log2 (count+1), and ‘count’ was considered as the
second-generation sequencing RSEM normalized count (12). For mutation analysis, the TCGA
thyroid carcinoma somatic mutation (broad automated) (ID,
TCGA.THCA.sampleMap/mutation_broad; version 20170908,) dataset,
consisting of 504 samples, was used. Analysis of gene set
enrichment was performed according to the Gene Set Enrichment
Analysis (GSEA) instructions (13,14).
Gene expression analysis results from 10 samples exhibiting the
highest levels of KRT19 expression, together with the 10 samples
with the lowest levels of KRT19 expression were submitted to GSEA
for analysis.
Cell lines, culture conditions, small
interfering (si)RNA, plasmids and transfection
The thyroid cancer cell line, 8505C was obtained
from Sigma-Aldrich (Merck KGaA; cat. no. 94090184), the cancer cell
line, BHT101, was obtained from the Leibniz Institute DSMZ-German
Collection of Microorganisms and Cell Cultures (cat. no. ACC 279),
Phoenix cells were obtained from the American Type Culture
Collection (ATCC; cat. no. CRL-3213). 8505C and BHT101 cells were
cultured in Eagle's minimum essential medium (ATCC; cat. no.
30-2003) and Phoenix cells in Dulbecco's modified Eagle's medium
(Thermo Fisher Scientific, Inc.; cat. no. 12491-015) supplemented
with 10% fetal bovine serum (FBS; cat. no. 10500064; Gibco; Thermo
Fisher Scientific, Inc.) and penicillin/streptomycin and cultured
in a humidified incubator at 37°C and 5% CO2.
Transfection was performed according to the
manufacturer's siRNA sequence transfection protocol for
Lipofectamine RNAiMAX (30 pmol siRNA per every 6 well plate well;
cat. no. 13778075; Thermo Fisher Scientific, Inc.) and the plasmid
Lipofectamine 2000 (2 µg plasmid DNA per every 6 well plate well;
cat. no. 11668027; Thermo Fisher Scientific, Inc.) protocol. The
wild type (WT) BRAFWT (pcDNA3.1, cat. no. 40775;
Addgene, Inc.) with its empty vector were transfected to the cells
followed by Lipofectamine 2000 instruction and mutant
BRAFV600E (pBabe, cat. no. 15269; Addgene, Inc.) with
its empty vector were transfected into retrovirus packaging phoenix
cells (with an VSV.G envelope plasmid; cat. no. 14888; Addgene,
Inc.), after 48 h culturing the virus supernatant was harvested and
filtered. The retrovirus was subsequently introduced into the
phoenix cells by spin infection at 1,200 × g for 90 min at 32°C. A
non-targeting RNA interference sequence (10 µM; Ambion; Thermo
Fisher Scientific, Inc.) was used as a negative control
(5′-CCTAAGGTTAAGTCGCCCTCGCTC-3′) for KRT19 siRNA experiments
(targeting sequence, 5′-AACCATGAGGAGGAAATCA-3′) (15). Transfection efficiency was evaluated
by reverse transcription-quantitative PCR (RT-qPCR) and western
blot analyses 48 h after transfection.
RNA preparation and RT-qPCR
Total RNA was extracted from cultured 8505C and
BHT101 cells using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). For reverse transcription, 1,000 ng total RNA
was reverse transcribed to produce a final volume of 20 µl cDNA
using a Reverse Transcription kit (cat. no. RR036A; Takara
Biotechnology, Co., Ltd.), according to the manufacturer's
protocols. RT-qPCR analyses were performed using SYBR Select Master
Mix (cat. no. 4472908; Applied Biosystems; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocols. The
RT-qPCR primers for KRT19 were as follows: Forward,
5′-AACGGCGAGCTAGAGGTGA-3′, and reverse,
5′-GGATGGTCGTGTAGTAGTGGC-3′, the primers for
BRAFWT/BRAFV600E were forward,
5′-AATACACCAGCAAGCTAGATGC-3′, and reverse,
5′-AATCAGTTCCGTTCCCCAGAG-3′. β-actin was used as the reference gene
with the following primer sequences: Forward,
5′-CATGTACGTTGCTATCCAGGC-3′ and reverse,
5′-CTCCTTAATGTCACGCACGAT-3′. RT-qPCR data was collected using the
QuantStudio™ 6 Flex Real-Time PCR System (Applied Biosystems;
Thermo Fisher Scientific, Inc.) and RT-qPCR thermocycling
conditions consisted of an initial denaturation step at 95°C for 10
min, followed by 40 cycles of 92°C for 15 sec and 60°C for 1 min.
Each sample was run in triplicate and the relative expression of
the target gene was calculated and normalized relative to β-actin
using the 2−ΔΔCq method (16) in Excel 16.18 software (Microsoft
Corporation).
Protein preparation and western blot
analysis
Whole cells were lysed on ice with RIPA buffer (cat.
no. P0013; Beyotime Institute of Biotechnology), and a
bicinchoninic acid assay kit was used to quantify the protein
concentration. Equal quantities (15 µg) of protein were loaded onto
12% SDS PAGE gels. Following separation, protein samples were
transferred to nitrocellulose membranes. The membranes were blocked
in 2% bovine serum albumin (cat. no. A4737; Sigma-Aldrich; Merck
KGaA) in Tris-buffered saline and Tween 20 (TBST) for 1 h at room
temperature and incubated with antibodies at 4°C overnight. The
membranes were then washed in TBST and incubated with HRP-coupled
secondary antibodies for 2 h at room temperature. After that
membranes were treated with enhanced chemiluminescence western
blotting substrate (Pierce; cat. no., 32106) at room temperature
for 1 min, X-ray films (CL-XPosure™ Film; cat. no. 34090, Thermo
Fisher Scientific, Inc.) were used for detecting the blots. All
experiments were repeated in triplicate and the antibodies are
presented in Table I.
 | Table I.Antibody list. |
Table I.
Antibody list.
| Antibody target | Supplier and
catalogue number | Host | Dilution |
|---|
| KRT19 | Abcam; cat. no.,
ab15463 | Rabbit | 1:200 |
| β-actin | Cell Signaling
Technology, Inc.; cat. no., 8H10D10 | Mouse | 1:500 |
|
BRAFV600E | Sigma-Aldrich; Merck
KGaA; cat. no., SAB5600047 | Rabbit | 1:300 |
| E-cadherin | Cell Signaling
Technology, Inc.; cat. no., 24E10 | Rabbit | 1:500 |
| N-cadherin | Cell Signaling
Technology, Inc.; cat. no., D4R1H | Rabbit | 1:500 |
| HRP-coupled
Anti-rabbit IgG | Cell Signaling
Technology, Inc.; cat. no., 14708 |
| 1:1,000 |
| HRP-coupled
Anti-mouse IgG | Cell Signaling
Technology, Inc.; cat. no., 14709 |
| 1:1,000 |
Cell proliferation and migration
assays
The Cell Counting Kit-8 (CCK8; cat. no. CK04;
Dojindo Molecular Technologies, Inc.) was used to determine the
cell proliferation rate. Cells were seeded in 96-well plates at a
density of 2,000 cells/well (100 µl total volume), and the
absorbance was measured at a wavelength of 450 nm using an ELx800
Universal Microplate Reader (Norgen Biotek Corp.), according to the
manufacturer's protocols. Each experiment was repeated in
quadruplicate at 0, 24, 48, 72 and 96 h.
For migration assays, transfected cells (40,000
cells/well; 100 µl total volume) were seeded in the upper chamber
of Transwell assay inserts (8-mm pores; EMD Millipore) containing
200 µl serum-free media. The lower chambers were filled with media
containing 10% FBS. Following incubation for 24 h, cells on the
filter surface were fixed with 90% methanol (−20°C) for 10 min and
then stained with crystal violet for 10 min at room temperature.
Images were captured using a light inverted microscope
(magnification, ×40; Axio Observer; Carl Zeiss AG) and cell
migration was assessed by counting the number of stained cells in
five random fields of view/filter for each group. Experiments were
repeated in triplicate.
Flow cytometry analysis
Apoptosis was analyzed by flow cytometry using an
Annexin V-fluorescein isothiocyanate (FITC) Apoptosis Detection kit
(BD Biosciences) with propidium iodide (PI), according to the
manufacturer's instructions. Briefly, cells were first washed and
resuspended at a concentration of 1×106 cells/ml. The
cells were then incubated with Annexin V-FITC (1:20 dilution) at
room temperature in the dark for 20 min and after that, PI (1:20
dilution) was added. The samples were immediately analyzed using a
FACScan flow cytometer (BD Biosciences). FlowJo software (version
10; FlowJo LLC) was used for data analysis. Each assay was
performed in triplicate.
Statistical analysis
SPSS Statistics 20.0 software (IBM Corp.) and
GraphPad Prism 7.04 (GraphPad Software, Inc.) software were used
for statistical analyses and the production of graphs. Data are
presented as the mean ± standard deviation, and were analyzed using
a Student's t-test, and one-way analysis of variance with Tukey's
multiple comparisons post-hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
KRT19 expression is increased in
thyroid cancer tissues and is associated with more aggressive
clinical characteristics
The TCGA thyroid cancer dataset was downloaded and
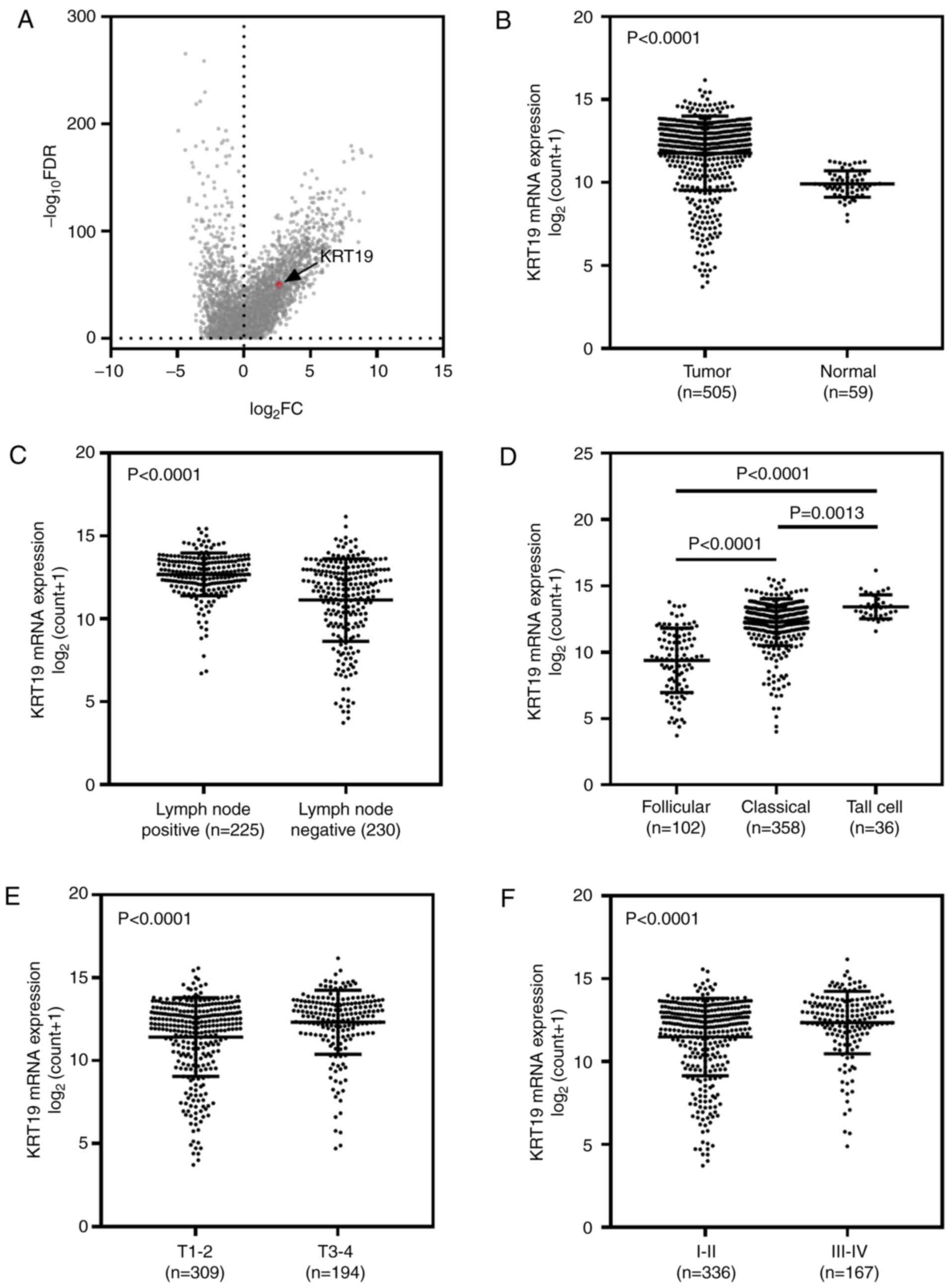
used for the purposes of this study. As demonstrated in Fig. 1A, the expression profile of genes in
the thyroid cancer dataset are presented as a volcano plot (cancer
vs. normal), and KRT19 was observed to be at an upregulated
position. As demonstrated in Fig.
1B, KRT19 expression was significantly higher in tumor samples
when compared with adjacent normal samples (P<0.001). In
addition, KRT19 expression was associated with lymph node
metastasis and significantly higher expression levels were observed
in the lymph node positive group (P<0.001; Fig. 1C). When patients were categorized by
pathological subtype, the follicular subtype (best prognosis) group
exhibited the lowest KRT19 expression levels, while the tall cell
subtype (worst prognosis) group exhibited the highest KRT19
expression levels (follicular subtype vs. classical subtype,
P<0.0001; follicular subtype vs. tall cell subtype, P<0.0001;
and classical subtype vs. tall cell subtype, P=0.0013; Fig. 1D). Patients in the TNM stage III–IV
group exhibited significantly higher KRT19 expression levels when
compared with those in the TNM stage I–II group (P<0.001;
Fig. 1F). Similar results were
observed for tumor stage analysis (P<0.001; Fig. 1E; Table
II).
| Table II.Clinical analysis in TCGA_THCA. |
Table II.
Clinical analysis in TCGA_THCA.
| Characteristics | Number of
patients | Percentage (%) | P-value |
|---|
| Age (years) |
|
| 0.010 |
|
<60 | 385 | 76.2 |
|
| ≥60 | 120 | 23.8 |
|
| Sex |
|
| 0.871 |
| Male | 136 | 26.9 |
|
|
Female | 369 | 73.1 |
|
| Primary tumor |
|
| <0.001 |
|
T1-2 | 310 | 61.4 |
|
|
T3-4 | 195 | 38.6 |
|
| Lymph node
metastasis |
|
| <0.001 |
|
Positive | 225 | 49.5 |
|
|
Negative | 230 | 50.5 |
|
| TNM stage |
|
| <0.001 |
|
I–II | 336 | 66.8 |
|
|
III–IV | 167 | 33.2 |
|
| Metastasis |
|
| 0.780 |
|
Positive | 282 | 96.9 |
|
|
Negative | 9 |
3.1 |
|
KRT19-knockdown inhibits thyroid
cancer cell proliferation and migration, however, not apoptosis in
vitro
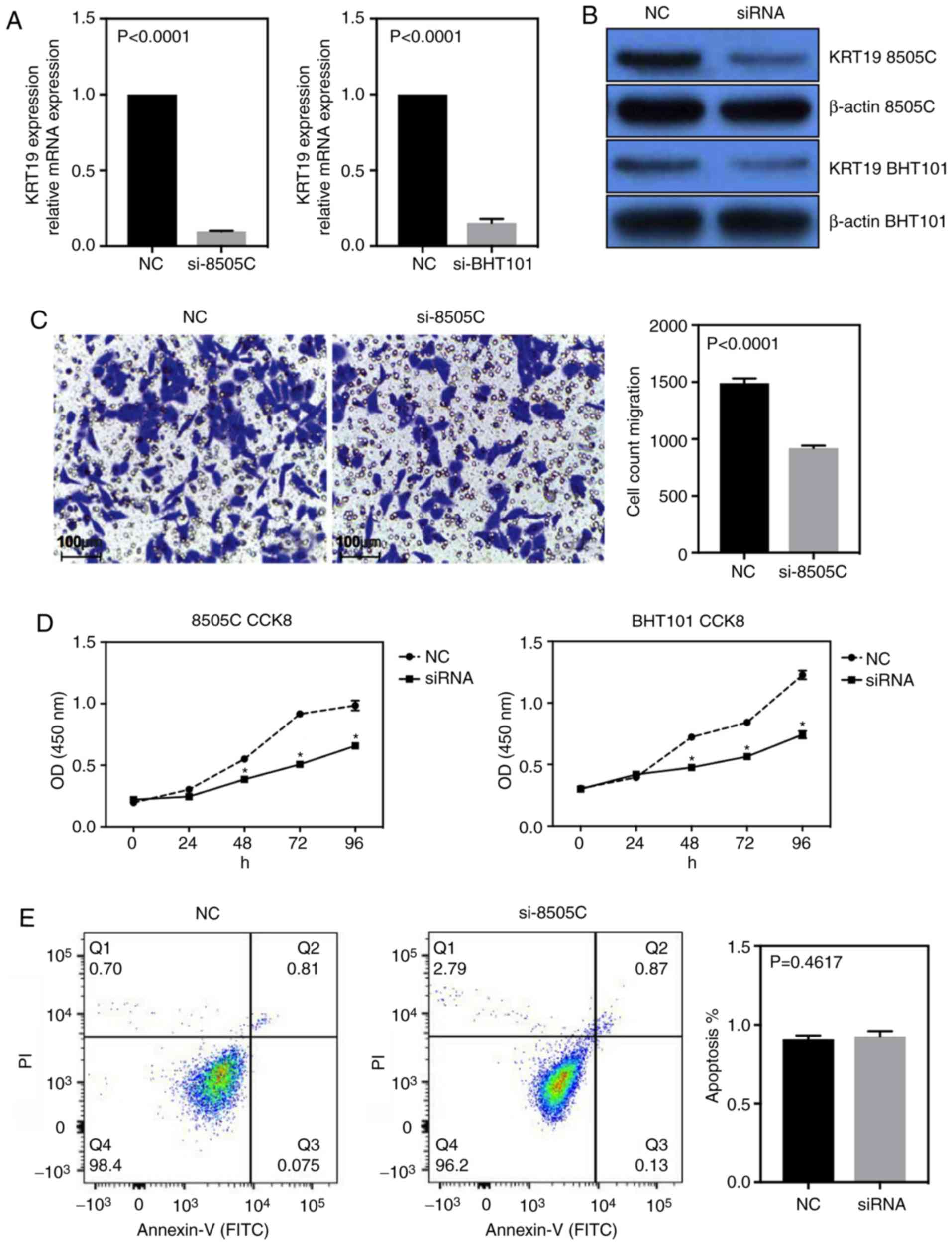
To investigate the role of KRT19 in thyroid cancer
cell lines the siRNA knockdown system was used. Using two different
thyroid cancer cell lines, 8505C and BHT101, the efficiency of
siRNA-knockdown was measured by RT-qPCR and western blot analyses
(Fig. 2A and B). Using Transwell
assays, the migration ability of 8505C cells was also measured. As
demonstrated in Fig. 2C, the
migration ability of 8505C cells was significantly reduced
following knockdown of KRT19 (P<0.001). The proliferation of
thyroid cancer cell lines was then analyzed using a CCK8 assay and
fluorescence-activated cell sorting (FACS) in both cell lines. As
presented in Fig. 2D, the negative
control (NC) groups demonstrated higher cell counts, and the
results of FACS analysis indicated no significant alterations in
the apoptosis rate between the NC and the si-KRT19 group (Fig. 2E). These results suggest that
knockdown of KRT19 decreased the proliferation ability of thyroid
cancer cells.
Increased KRT19 expression may be
induced by the BRAFV600E mutation
In the TCGA thyroid cancer dataset, it was observed
that compared with normal group, the expression of KRT19 was higher
only in the BRAFV600E group, however, not the WT group
(Fig. 3A). Therefore,
BRAFV600E and BRAFWT sequences were then
overexpressed in 8505C cells. qRT-PCR results indicated these
plasmids could significantly upregulate the expression of
BRAFWT or BRAFV600E (Fig. 3B and C). As demonstrated in Fig. 3D and E, the mutation group exhibited
higher expression levels of KRT19 when compared with WT group. The
results demonstrated that high expression levels of KRT19 were only
observed in patients harboring the BRAFV600E mutation,
which suggests that BRAFV600E may induce KRT19
expression.
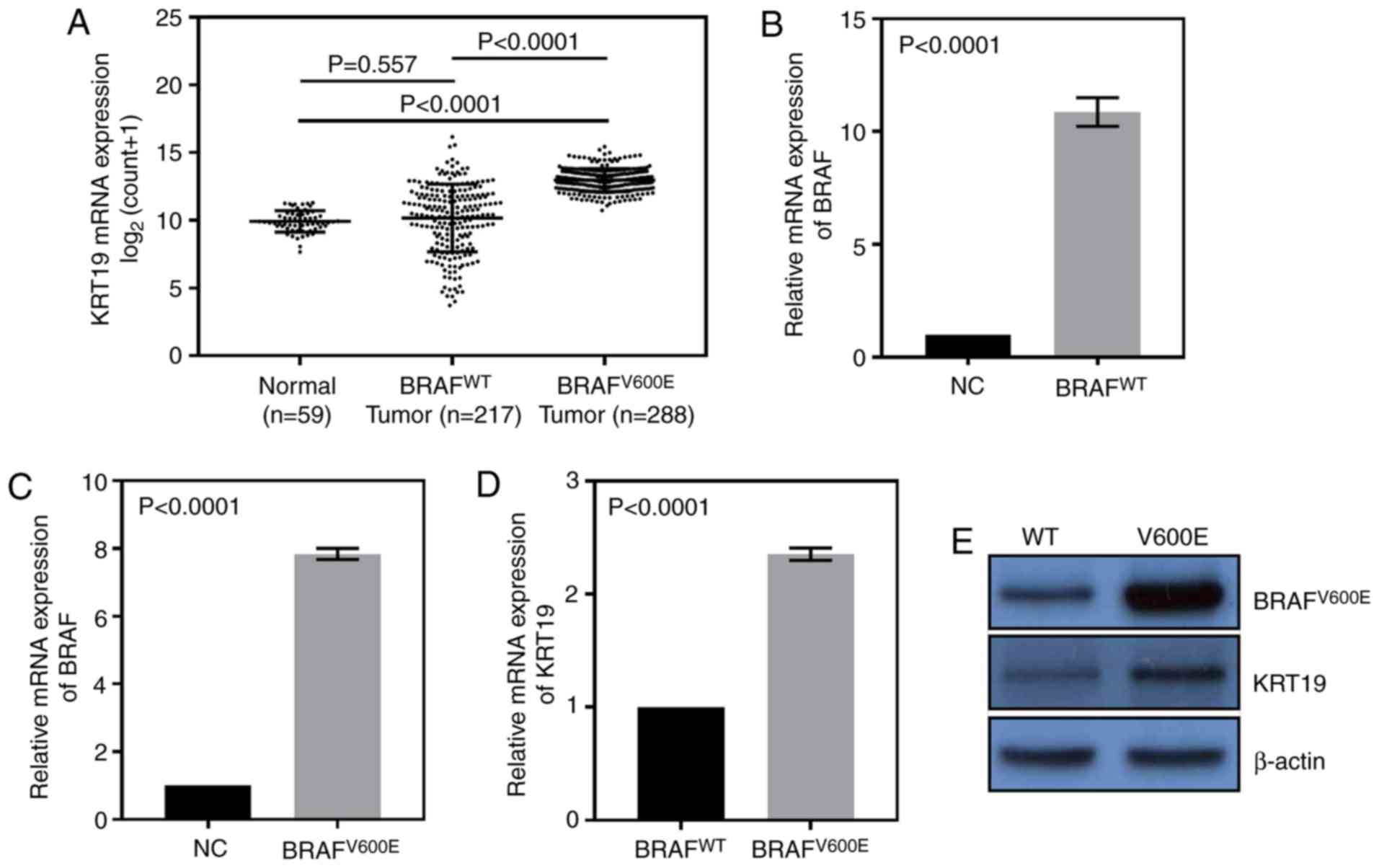 | Figure 3.The TCGA_THCA dataset was used for the
following analysis. (A) Compared with the BRAF wild type group, the
expression of KRT19 was increased in BRAFV600E group,
however, not in BRAF wild type group. The (B) wild type BRAF or (C)
V600E mutated BRAF overexpression plasmids were transfected into
8505C cells, and may be overexpressed significantly compared with
their empty vectors, BRAFWT/BRAFV600E. (D)
mRNA and (E) protein expression levels of the BRAFV600E
mutation group exhibited higher expression levels of KRT19 in 8505C
cells when compared with BRAF wild type group. BRAF, B-Raf
Proto-Oncogene, Serine/Threonine Kinase; TCGA, The Cancer Genome
Atlas Network; THCA, thyroid cancer; KRT19, keratin 19; WT, wild
type. |
KRT19 exerts its oncogenic activity
via EMT
To further investigate the role of KRT19 and the
BRAFV600E mutation in thyroid cancer, GSEA was used.
Patients from the TCGA thyroid cancer dataset were grouped
according to BRAF mutation status (BRAFV600E and WT BRAF
groups), and the expression profiles for each group were submitted
for GSEA analysis. The results indicated that the BRAF mutation
status was significantly associated with EMT (Fig. 4A and D). As demonstrated in Fig. 4B and C, GSEA was used to analyze the
gene expression profiles of the top 10 tumor samples exhibiting the
highest levels of KRT19 expression and the top 10 tumor samples
exhibiting the lowest expression of KRT19, which revealed EMT.
These results suggest that KRT19 may influence EMT. Therefore, the
expression of EMT markers was analyzed in 8505C cells, and the
results demonstrated that the level of E-cadherin was significantly
higher in cells transfected with si-KRT19, whereas N-cadherin
expression levels were significantly lower when compared with the
controls.
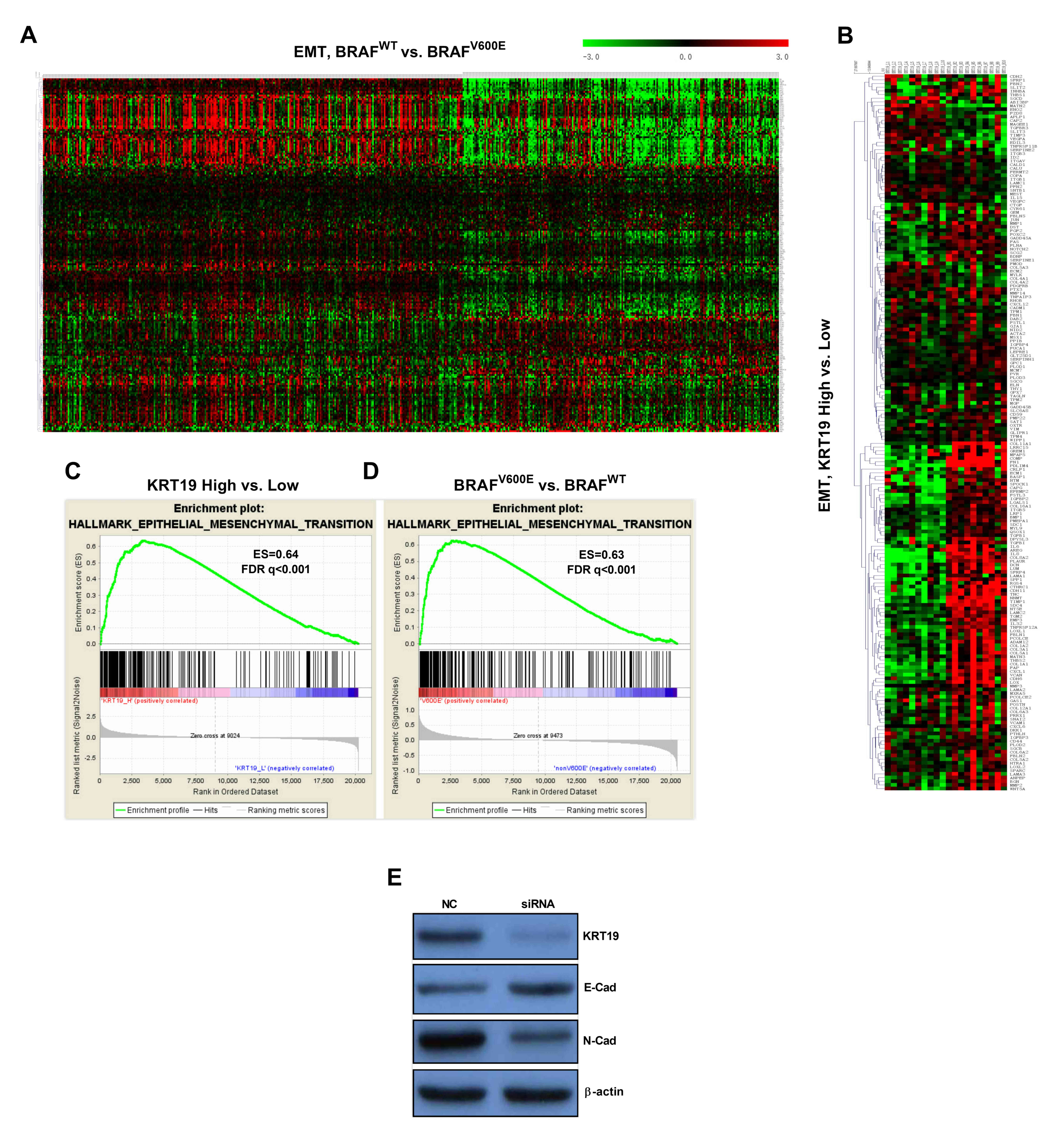 | Figure 4.The TCGA_THCA dataset was used for the
following analysis. (A and D) Heatmap and GSEA analysis in EMT and
BRAFV600E tumor vs. wild type BRAF tumor samples from
the TCGA_THCA dataset. (B and C) Heatmap of EMT genes in the top 10
samples exhibiting high KRT19 expression vs. 10 samples with the
lowest KRT19 expression from the TCGA dataset. (E) Western blot
analysis of EMT markers in the 8505C cell line transfected with
controls or si-KRT19. Knockdown of KRT19 was associated with
increased E-cadherin and reduced N-cadherin levels when compared
with the control group. BRAF, B-Raf Proto-Oncogene,
Serine/Threonine Kinase; TCGA, The Cancer Genome Atlas Network;
THCA, thyroid cancer; GSEA, Gene Set Enrichment Analysis; EMT,
epithelial-mesenchymal-transition; KRT19, keratin-19; siRNA, small
interfering RNA; FDR, false discovery rate; ES, enrichment
score. |
Discussion
KRT19 is a member of the keratin family of IF
proteins, which can be further subdivided into CKs and hair
keratins (17). KRT19, as a type I
CK, has been reported to be abundantly expressed in epithelial
tumor cells and serves as a marker for metastatic tumors (18–20). A
previous study suggested that KRT19 expression may be activated by
certain signaling pathways. For example, Ju et al (10) reported that the erb-b2 receptor
tyrosine kinase-2 (ERBB2/HER2)-downstream MEK/ERK signaling pathway
stimulated KRT19 expression, and membrane-localized KRT19 can bind
to and stabilize HER2 by inhibiting its ubiquitination. In thyroid
cancer, increased KRT19 expression was observed in papillary
carcinoma when compared with follicular adenoma (21). However, the role of KRT19 in thyroid
cancer is currently unclear, so in order to clarify the phonotype
of KRT19 in thyroid cancer cell lines and examine its downstream
mechanism the present study designed and performed the
aforementioned experiments.
The current study presents evidence to suggest that
KRT19 expression is elevated in thyroid cancer, which is associated
with lymph node metastasis, increased tumor size and advanced TNM
stage. Using CCK8 assays and apoptosis analysis, KRT19 knockdown
was demonstrated to reduce the proliferation of thyroid cancer
cells; however, this was not via the induction of apoptosis. In
agreement with the observation that KRT19 was associated with lymph
node metastasis, the results of the Transwell assay indicated that
KRT19-knockdown decreased the migration of thyroid cancer
cells.
BRAFV600E is the most frequently mutated
gene in thyroid cancer and, according to the cBioPortal, this
mutation is present in ~60% of all cases (22). Therefore, the association between
BRAFV600E and KRT19 expression was investigated in the
current study. Notably, the expression of KRT19 was observed to be
significantly higher in BRAFV600E tumors compared with
WT tumors. By transfecting thyroid tumor cells with a plasmid
containing BRAFV600E, this mutation was observed to
induce the expression of KRT19. Therefore, to the best of our
knowledge, this is the first study to report that increased KRT19
expression could be caused by BRAFV600E mutations.
Consistent with this hypothesis, GSEA analysis of
BRAFV600E vs. WT BRAF thyroid cancer samples indicated
EMT. In addition, KRT19 expression was associated with lymph node
metastasis in thyroid cancer patients, and GSEA analysis yielded
similar results. KRT19 is also known to be involved in EMT
(8). Therefore, the level of
specific EMT makers were measured in thyroid cancer cell lines in
the present study. E-cadherin levels were increased while
N-cadherin levels were decreased in thyroid cancer cells
transfected with si-KRT19, which indicated that knockdown of KRT19
may influence EMT.
In conclusion, the results of the current study
suggest that BRAFV600E mutations may increase the
expression of KRT19 in thyroid cancer, which may subsequently lead
to lymph node metastasis via induction of EMT.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Health and
Family Planning Commission of Shanxi Province (grant no.
2017070).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
XW and HZ designed the study. XW, CP, TG and XX
performed the experiments. YQ and JJ performed the statistical
analysis and XW and YQ wrote the manuscript. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Smith-Bindman R, Lebda P, Feldstein VA,
Sellami D, Goldstein RB, Brasic N, Jin C and Kornak J: Risk of
thyroid cancer based on thyroid ultrasound imaging characteristics:
Results of a population-based study. JAMA Intern Med.
173:1788–1796. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cabanillas ME, McFadden DG and Durante C:
Thyroid cancer. Lancet. 388:2783–2795. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pellegriti G, Frasca F, Regalbuto C,
Squatrito S and Vigneri R: Worldwide increasing incidence of
thyroid cancer: Update on epidemiology and risk factors. J Cancer
Epidemiol. 2013:9652122013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Randle RW, Bushman NM, Orne J, Balentine
CJ, Wendt E, Saucke M, Pitt SC, Macdonald CL, Connor NP and Sippel
RS: Papillary thyroid cancer: The good and bad of the ‘good
cancer’. Thyroid. 27:902–907. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karantza V: Keratins in health and cancer:
More than mere epithelial cell markers. Oncogene. 30:127–138. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paiva F, Duarte-Pereira S, Costa VL,
Ramalho-Carvalho J, Patrício P, Ribeiro FR, Lobo F, Oliveira J,
Jerónimo C and Henrique R: Functional and epigenetic
characterization of the KRT19 gene in renal cell neoplasms. DNA
Cell Biol. 30:85–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cen D, Chen J, Li Z, Zhao J and Cai X:
Prognostic significance of cytokeratin 19 expression in pancreatic
neuroendocrine tumor: A meta-analysis. PLoS One. 12:e01875882017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takano M, Shimada K, Fujii T, Morita K,
Takeda M, Nakajima Y, Nonomura A, Konishi N and Obayashi C: Keratin
19 as a key molecule in progression of human hepatocellular
carcinomas through invasion and angiogenesis. BMC Cancer.
16:9032016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Masai K, Nakagawa K, Yoshida A, Sakurai H,
Watanabe S, Asamura H and Tsuta K: Cytokeratin 19 expression in
primary thoracic tumors and lymph node metastases. Lung Cancer.
86:318–323. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ju JH, Oh S, Lee KM, Yang W, Nam KS, Moon
HG, Noh DY, Kim CG, Park G, Park JB, et al: Cytokeratin19 induced
by HER2/ERK binds and stabilizes HER2 on cell membranes. Cell Death
Differ. 22:665–676. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: the 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li B and Dewey CN: RSEM: Accurate
transcript quantification from RNA-Seq data with or without a
reference genome. BMC Bioinformatics. 12:3232011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mootha VK, Lindgren CM, Eriksson KF,
Subramanian A, Sihag S, Lehar J, Puigserver P, Carlsson E,
Ridderstråle M, Laurila E, et al: PGC-1alpha-responsive genes
involved in oxidative phosphorylation are coordinately
downregulated in human diabetes. Nat Genet. 34:267–273. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ohtsuka T, Sakaguchi M, Yamamoto H, Tomida
S, Takata K, Shien K, Hashida S, Miyata-Takata T, Watanabe M,
Suzawa K, et al: Interaction of cytokeratin 19 head domain and HER2
in the cytoplasm leads to activation of HER2-Erk pathway. Sci Rep.
6:395572016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Saha SK, Choi HY, Kim BW, Dayem AA, Yang
GM, Kim KS, Yin YF and Cho SG: KRT19 directly interacts with
β-catenin/RAC1 complex to regulate NUMB-dependent NOTCH signaling
pathway and breast cancer properties. Oncogene. 36:332–349. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stathopoulos EN, Sanidas E, Kafousi M,
Mavroudis D, Askoxylakis J, Bozionelou V, Perraki M, Tsiftsis D and
Georgoulias V: Detection of CK-19 mRNA-positive cells in the
peripheral blood of breast cancer patients with histologically and
immunohistochemically negative axillary lymph nodes. Ann Oncol.
16:240–246. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang XR, Xu Y, Shi GM, Fan J, Zhou J, Ji
Y, Sun HC, Qiu SJ, Yu B, Gao Q, et al: Cytokeratin 10 and
cytokeratin 19: Predictive markers for poor prognosis in
hepatocellular carcinoma patients after curative resection. Clin
Cancer Res. 14:3850–3859. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen TF, Jiang GL, Fu XL, Wang LJ, Qian H,
Wu KL and Zhao S: CK19 mRNA expression measured by
reverse-transcription polymerase chain reaction (RT-PCR) in the
peripheral blood of patients with non-small cell lung cancer
treated by chemo-radiation: An independent prognostic factor. Lung
Cancer. 56:105–114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Arcolia V, Journe F, Renaud F, Leteurtre
E, Gabius HJ, Remmelink M and Saussez S: Combination of galectin-3,
CK19 and HBME-1 immunostaining improves the diagnosis of thyroid
cancer. Oncol Lett. 14:4183–4189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|