Introduction
Lung cancer remains the most common form of cancer,
with the highest mortality rate globally, making it a major public
health threat (1,2). Surgery, chemotherapy, radiotherapy and
molecular targeted therapies are currently the primary treatment
options for this disease (3). Whilst
early lung cancer may be treated surgically, the treatment options
for advanced metastatic lung cancer are limited. Although
radiotherapy and chemotherapy postpone the progression of these
advanced lung cancer types, the survival rate of patients is low,
and patients are often unable to tolerate the side effects of these
therapies. Molecular targeted therapies are only effective in
patients with lung adenocarcinoma who have specific mutations in
genes including epidermal growth factor receptor (EGFR) or ALK
receptor tyrosine kinase (ALK) (4,5).
Patients who have wild-type EGFR or ALK and
non-adenovirus-associated non-small cell lung cancer lack effective
targeted therapies. Thus, there remains a clear need for the
identification of novel therapeutics suitable for treating patients
with advanced lung cancer.
The enzyme acetyl-CoA acetyltransferase 1 (ACAT-1)
is a central mediator of the cholesterol esterification pathway
(6). Previous studies have
identified that ACAT-1 is abnormally expressed in certain cancer
types, including prostate cancer, pancreatic cancer, leukemia,
glioma, breast cancer and colon cancer (7–12).
However, the functions of ACAT-1 in lung cancer are not well
understood. In the present study, the effects of the inhibition of
ACAT-1 on Lewis lung cancer (LLC) growth and metastasis were
investigated using in vitro cellular experiments and in
vivo animal models. The effect of downregulation of ACAT-1
expression on proliferation, migration and apoptosis of LLC cells
was observed at the cell level in vitro. The effects of
ACAT-1 inhibitor avasimibe on tumor growth and metastasis in LLC
mice were observed in an animal model in vivo, and the
expression of ACAT-1 in Lewis lung carcinoma tissues was detected
to add new content for lung cancer research and provide new
strategies for clinical lung cancer treatment.
Materials and methods
Ethics statement
The Research Ethics Committee of Bengbu Medical
College (Bengbu, China) ethically approved this study.
Cell culture
The LLC cell line is a malignant murine lung cancer
cell line which was obtained from the Type Culture Collection of
the Chinese Academy of Sciences (Shanghai, China). LLC cells were
grown in 25 cm3 cell culture vessels containing
Dulbecco's modified Eagle's medium (DMEM; Hyclone; GE Healthcare
Life Sciences, Logan, UT, USA), 10% foetal bovine serum (FBS;
Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.,
Hangzhou, China) and penicillin/streptomycin in standard growth
conditions (37°C). After culturing for approximately 3 days, cell
subculture was performed when the cell density reached ~80% of the
bottom of the culture bottle.
Cell viability assessment
Cell proliferation was measured using a Cell
Counting Kit-8 (CCK-8; Biosharp; Beijing Lanjieke Technology Co.,
Ltd., Hefei, China) assay, as previously described (13). Briefly, LLC cells in the logarithmic
phase of growth were harvested, resuspended at 5×104/ml,
and 100 µl cells were added to a 96 well plate (5,000 cells/well)
with five replicates per condition. Once adherence to the plates
was achieved, a concentration gradient of avasimibe (Med Chem
Express LLC, Monmouth Junction, NJ, USA) of 0.0, 2.5, 5.0, 10.0 and
20.0 µM was added to the corresponding wells. Following 24, 48, 72
or 96 h of incubation at 37°C, CCK-8 solution was added to each
well followed by a 1–4 h incubation at 37°C. The optical density at
450 nm was then determined via a microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA). Viability percentages were
determined by comparing with the blank for control and treated
samples.
Wound healing assay
This assay was conducted as described previously
(14). Briefly, LLC cells were
plated in 6-well plates (5×105 cells/well). Subsequent
to achieving 90% confluency, a vertical scratch (width, ~900 µm)
was created in the monolayer using a sterile pipette tip. Plates
were then washed using phosphate buffered saline (PBS) and
serum-free DMEM containing either 0, 5 or 10 µM avasimibe was added
to the appropriate wells. After 48 h, cell migration was observed
and photographed at 0 and 48 h after scratching. The width of the
healing wound was calculated using ImageJ software (version 1.8.0;
National Institutes of Health, Bethesda, MD, USA). The wound
healing rate was determined based on the ratio of the change in the
scratch width to the initial scratch width.
Flow cytometry
LLC cells were plated in a 6 well plate
(5×105 cells/ml) and harvested after 72 h treatment with
0, 10 or 20 µM avasimibe. Cells were then fixed at room temperature
in 4% paraformaldehyde (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) for 30 min, prior to permeabilization using 0.1% Triton
X-100 (Sigma-Aldrich; Merck KGaA) for 20 min. The cells were then
blocked in 5% goat serum (Hangzhou Sijiqing Biological Engineering
Materials Co., Ltd.) and 0.3% Triton X-100 for 30 min at 37°C.
Cells were incubated with anti-ACAT-1 antibody (pAb; 1:200; catalog
no. #44276; Cell Signaling Technology, Inc., Danvers, MA, USA) for
30 min at 37°C followed by fluorescein isothiocyanate
(FITC)-labeled goat anti-rabbit immunoglobulin G (IgG; 1:200;
catalog no. #4412; Cell Signaling Technology, Inc.) for 30 min at
37°C. The cells were washed 2–3 times with 3 ml TPBS buffer. A
Cytomics FC 500 flow cytometer (BD FACSCalibur; BD Biosciences,
Franklin Lakes, NJ, USA) was then used to detect labeled cells.
Mean fluorescence intensity was analyzed using FlowJo software
(version 7.6; FlowJo LLC, Ashland, OR, USA).
Western blotting
LLC cells were seeded (1×106 cells/ml)
into 60 mm culture dishes and harvested after 72 h treatment with 0
or 10 µM avasimibe. Subsequent to washing three times with PBS, a
cell lysis buffer (PMSF:RIPA=1:100; Beyotime Institute of
Biotechnology, Shanghai, China) was used to harvest the cells, and
protein was quantified via a BCA assay (Beyotime Institute of
Biotechnology). Samples were then separated via SDS-PAGE (10% gel)
with 30 µg per lane and then transferred to a polyvinylidene
fluoride membrane (Beyotime Institute of Biotechnology). A total of
5% skim milk was used for membrane blocking for 2 h at room
temperature, and membranes were incubated at 4°C with the primary
antibody (rabbit anti-mouse ACAT-1 polyclonal antibody; 1:1,000;
catalog no. #44276; Cell Signaling Technology, Inc.) overnight. A
horseradish peroxidase-conjugated secondary anti-rabbit IgG
antibody (goat anti-rabbit IgG; 1:8,000; catalog no. BL003A;
Biosharp; Beijing Lanjieke Technology Co., Ltd.) was then used for
detection for 2 h at 37°C. ACAT-1 was then visualized using an ECL
(EMD Millipore, Billerica, MA, USA) chromogenic reaction in a dark
room. Bands were quantified using ImageJ software (version 1.8.0;
National Institutes of Health, Bethesda, MD, USA).
Immunofluorescence
LLC cells were seeded onto coverslips
(1×105 cells/ml). Following treatment with 0 and 10 µM
avasimibe for 72 h, the coverslips were fixed using 4%
paraformaldehyde and permeabilized as aforementioned, and were then
blocked in 5% goat serum and 0.3% Triton X-100 for 30 min at 37°C.
The coverslips were then incubated with anti-ACAT-1 antibody (pAb;
1:200; catalog no. #44276; Cell Signaling Technology, Inc.,
Danvers, MA, USA) for 30 min at 37°C and then incubated with
FITC-labeled goat anti-rabbit IgG (1:50; catalog no. BL003A;
Biosharp; Beijing Lanjieke Technology Co., Ltd.) for 30 min at
37°C. DAPI (Beyotime Institute of Biotechnology) nuclear staining
was then performed while samples were in the dark for 5 min at room
temperature. Subsequent to rinsing with PBS, the coverslips were
fixed on glass slides using 1:1 glycerol:water for 5 min at room
temperature. Confocal microscopy (magnification, ×600; FV-1200MPE
SHARE; Olympus Corporation, Tokyo, Japan) was used for imaging, and
ImageJ software (version 1.8.0; National Institutes of Health,
Bethesda, MD, USA) was used for quantitative analyses.
Apoptosis detection
Apoptosis was detected using an Apoptosis Detection
kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China) according to
the manufacturer's protocol. Subsequent to LLC cell treatment with
avasimibe (0 and 10 µM) for 72 h, the cells were digested with
EDTA-free trypsin and resuspended in 500 µl binding buffer. Samples
were then stained with 1:100 annexin V and propidium iodide for 10
min at 4°C in the dark, and cells were detected using flow
cytometry as described above. The rate of apoptosis was analyzed
using FlowJo software (version 7.6; FlowJo LCC
In vivo mouse model experiments
A total of 24 male C57BL/6 mice (age range, 8–10
weeks; weight, 18–22 g) were obtained from the Experimental Animal
Center of Bengbu Medical College (Bengbu, China). The mice were
housed in a pathogen-free central animal facility (temperature,
20~26°C; relative humidity, 40~70%; light-dark alternate time,
12/12 h; food and water were disinfected and sterilized, the mice
had continuous access to the food and water) at the Bengbu Medical
College. The animal experiments were performed based on the
recommendations provided in the National Institutes of Health
Laboratory Animal Care and Use Guidelines (15). LLC cells were washed twice with PBS
and filtered through a 40 µm filter membrane prior to being
resuspended at a density of 2×107 cells/ml and
subcutaneously injected into the left forelimbs of these mice.
Following 10 days, a total of 34 mice meeting the experimental
requirements, that the tumor sizes were similar, were
identified.
In the avasimibe treatment trial, 24 mice were
randomized into 4 groups: A control group, a cyclophosphamide (CTX;
20 mg/kg, once every other day) group, an avasimibe (15 mg/kg, once
every 2 days) group and a CTX + avasimibe group. From days 10–35
following tumor inoculation, CTX or avasimibe were injected
intraperitoneally and mouse body weight was monitored once weekly.
On day 35, all mice were euthanized and tumors, livers, lungs and
spleens were harvested. Spleens were then weighed, tumor volume was
determined, and liver and lung samples were inspected for evidence
of metastases.
For the remaining 10 tumor model mice, the lungs
were collected on day 35. In parallel, lung samples from 10 healthy
tumor-free mice were also collected. Lung tissue protein was then
extracted and ACAT-1 expression was detected by western
blotting.
Statistical analysis
All in vitro experiments were conducted at
least three times. SPSS v16.0 (SPSS, Inc., Chicago, IL, USA) was
used for all statistical analyses. Data are provided as the mean ±
standard deviation, and were assessed using one-way analyses of
variance and Student's t-tests. The least significant difference-t
test was used as a post-hoc test for comparison between multiple
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Decreased expression levels of ACAT-1
inhibit LLC cell proliferation
LLC cells were treated with different avasimibe
concentrations to inhibit ACAT-1, revealing a significant dose- and
time-dependent suppression of proliferation as revealed using a
CCK-8 assay (P<0.05; Fig. 1A and
B). The cell viability rates in the blank group and the
avasimibe (2.5, 5, 10 and 20 µM) groups were 100.00±0.00,
63.57±4.88, 45.47±5.35, 37.66±3.72 and 30.59±1.24%, respectively
(Fig. 1A). In addition, the
viability of the control group and the groups at 1, 2, 3 and 4 days
were 100.00±0.00, 72.21±4.50, 58.60±5.25, 46.11±3.9 and
39.02±3.04%, respectively (Fig. 1B).
These results therefore demonstrated that avasimbe inhibits LLC
cell proliferation compared with the controls.
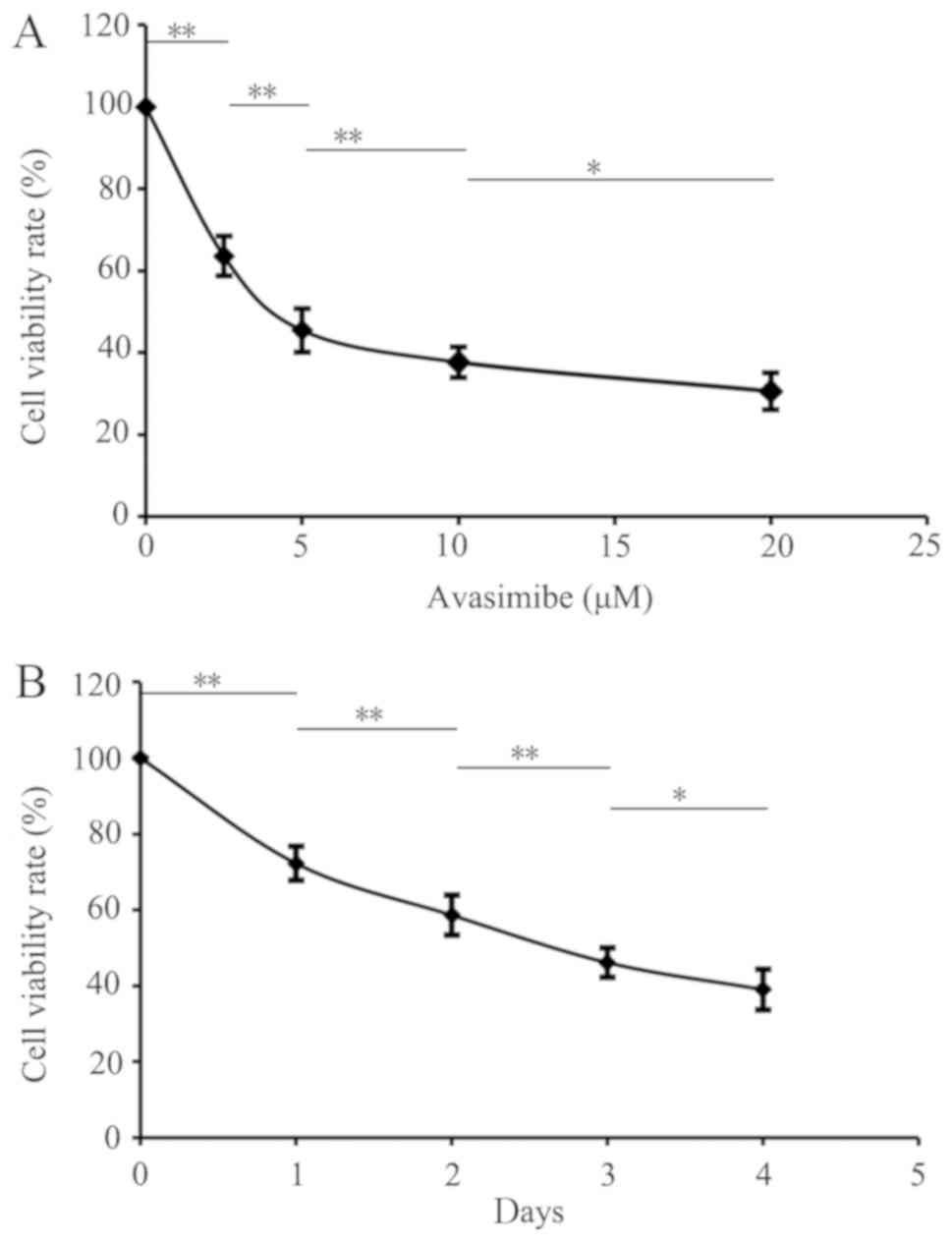 | Figure 1.Downregulating ACAT-1 expression
using avasimibe may inhibit the viability of LLC cells. (A) Effect
of different concentrations of avasimibe (0, 2.5, 5, 10 and 20 µM)
on LLC cell viability at 72 h, with 0 µM avasimibe used as a
control. (B) Effects of avasimibe (10 µM) on the viability of LLC
cells at 0, 24, 48, 72 and 96 h, with 0 h used as a control.
*P<0.05 and **P<0.01 with comparisons shown by lines. ACAT-1,
acetyl-CoA acetyltransferase 1; LLC, Lewis lung cancer. |
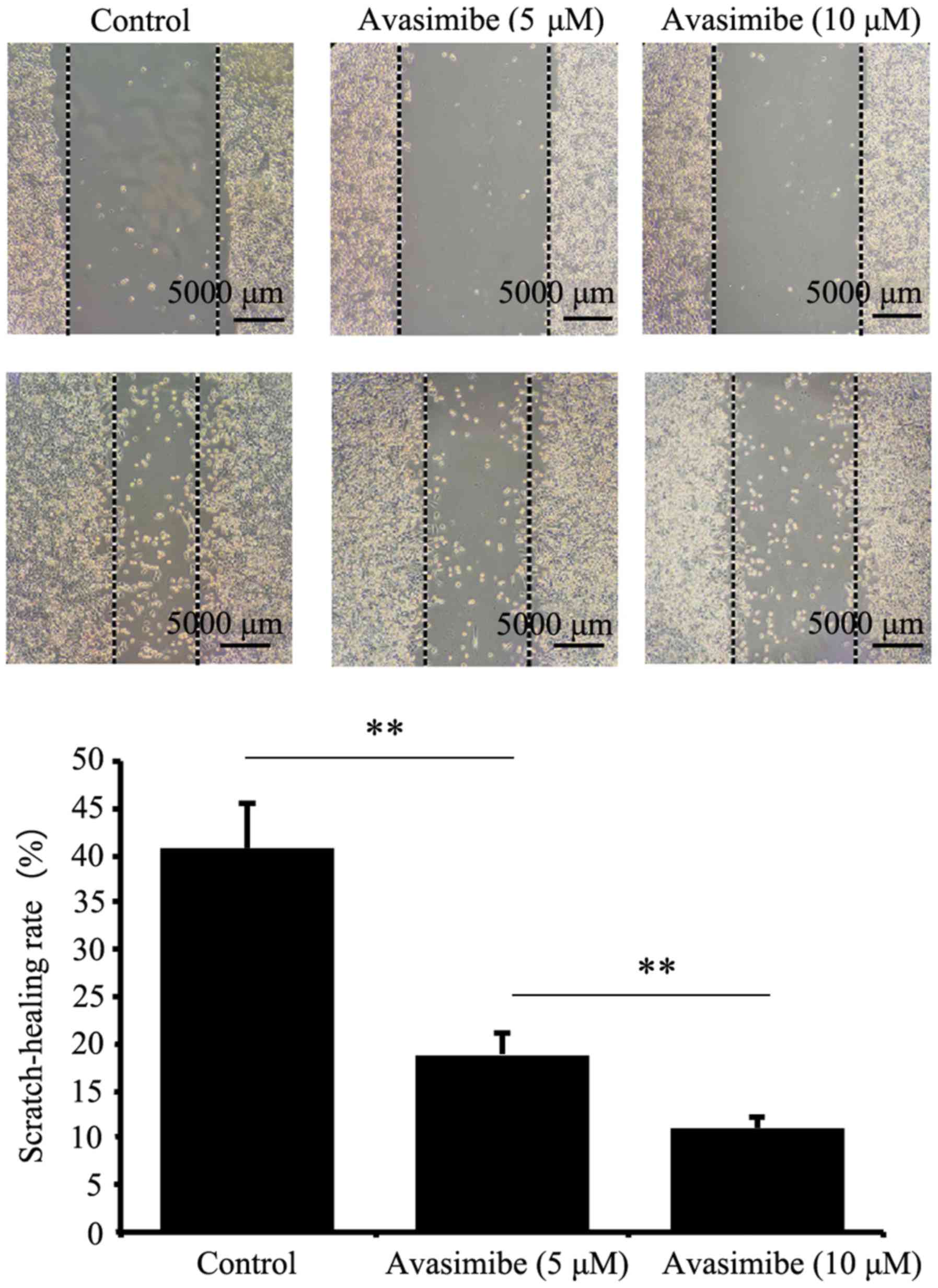
Decreased ACAT-1 expression inhibits
LLC cell migration
As presented in Fig.
2, cell migration was significantly reduced in the avasimibe
group compared with the control group (P<0.01). The
wound-healing rates of the avasimibe-treated (5 and 10 µM) groups
and the blank group were 18.90±2.37, 11.07±1.27 and 40.63±4.98%,
respectively. This indicated that avasimibe inhibited the migration
of LLC cells compared with the controls.
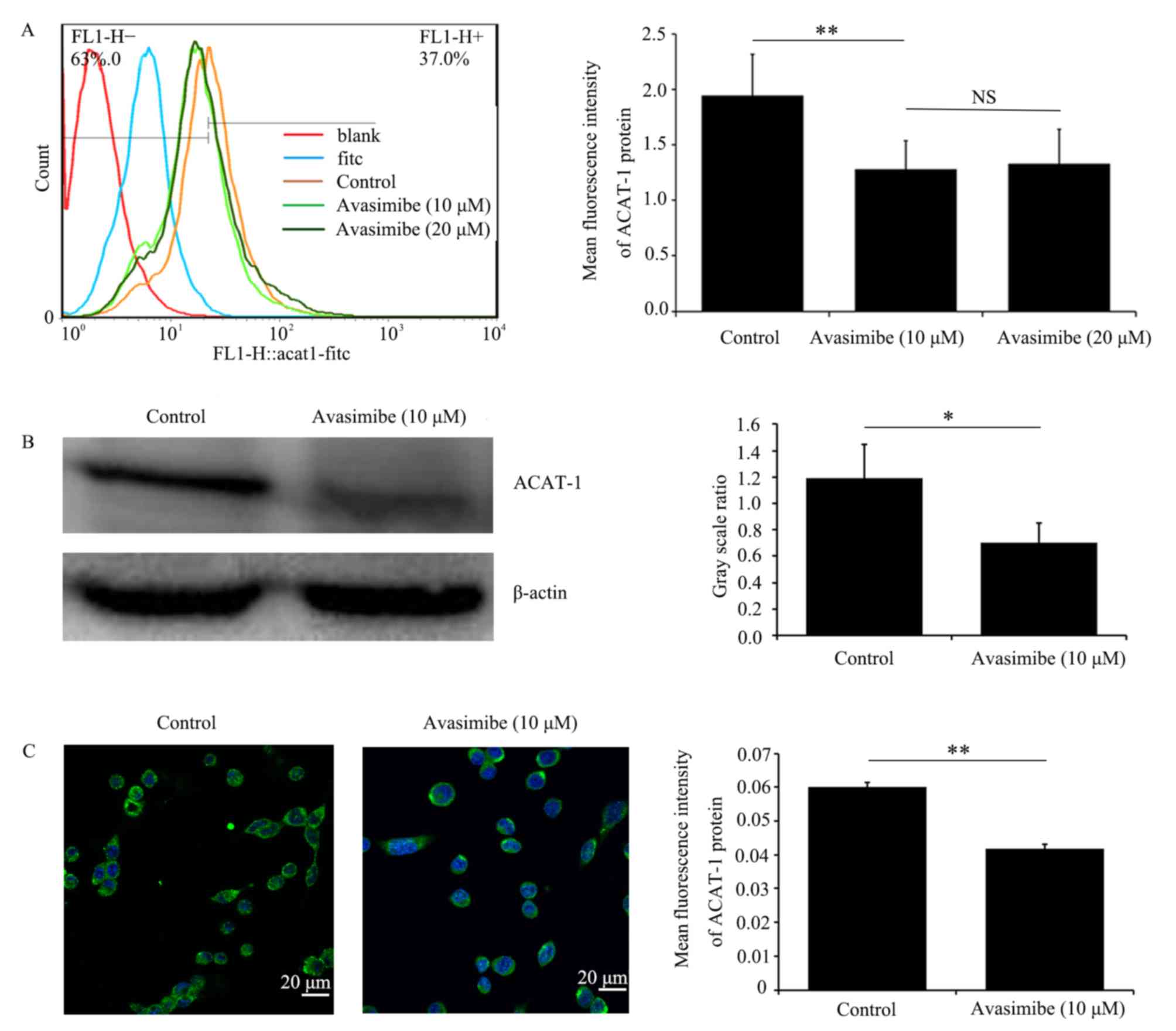
Avasimibe downregulates ACAT-1
expression in LLC cells
LLC cells were treated with different avasimibe
doses for 72 h, following which the ability of avasimibe to inhibit
ACAT-1 expression was validated using flow cytometry, western
blotting and immunofluorescence microscopy. As presented in
Fig. 3, it was revealed that
avasimibe treatment resulted in the significant downregulation of
ACAT-1 protein expression levels in LLC cells compared with the
control groups (P<0.05).
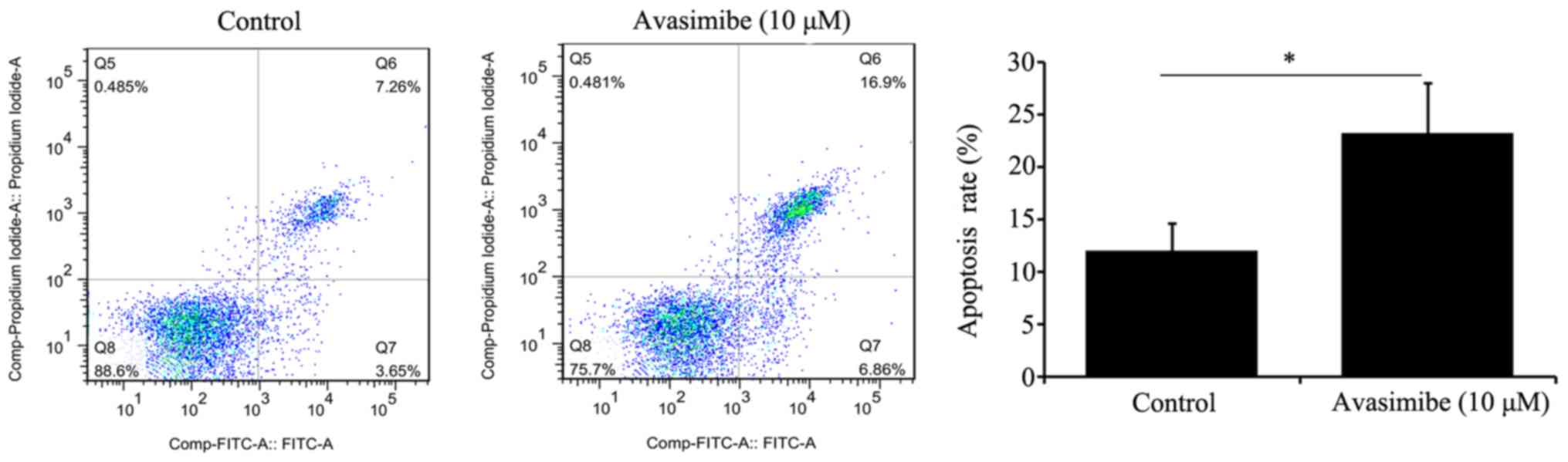
Avasimibe promotes the apoptosis of
LLC cells
Based on a flow cytometry-based analysis, avasimibe
(10 µM) treatment of LLC cells for 72 h significantly increased the
apoptotic rate of these cells compared with the control group
(P<0.05; Fig. 4).
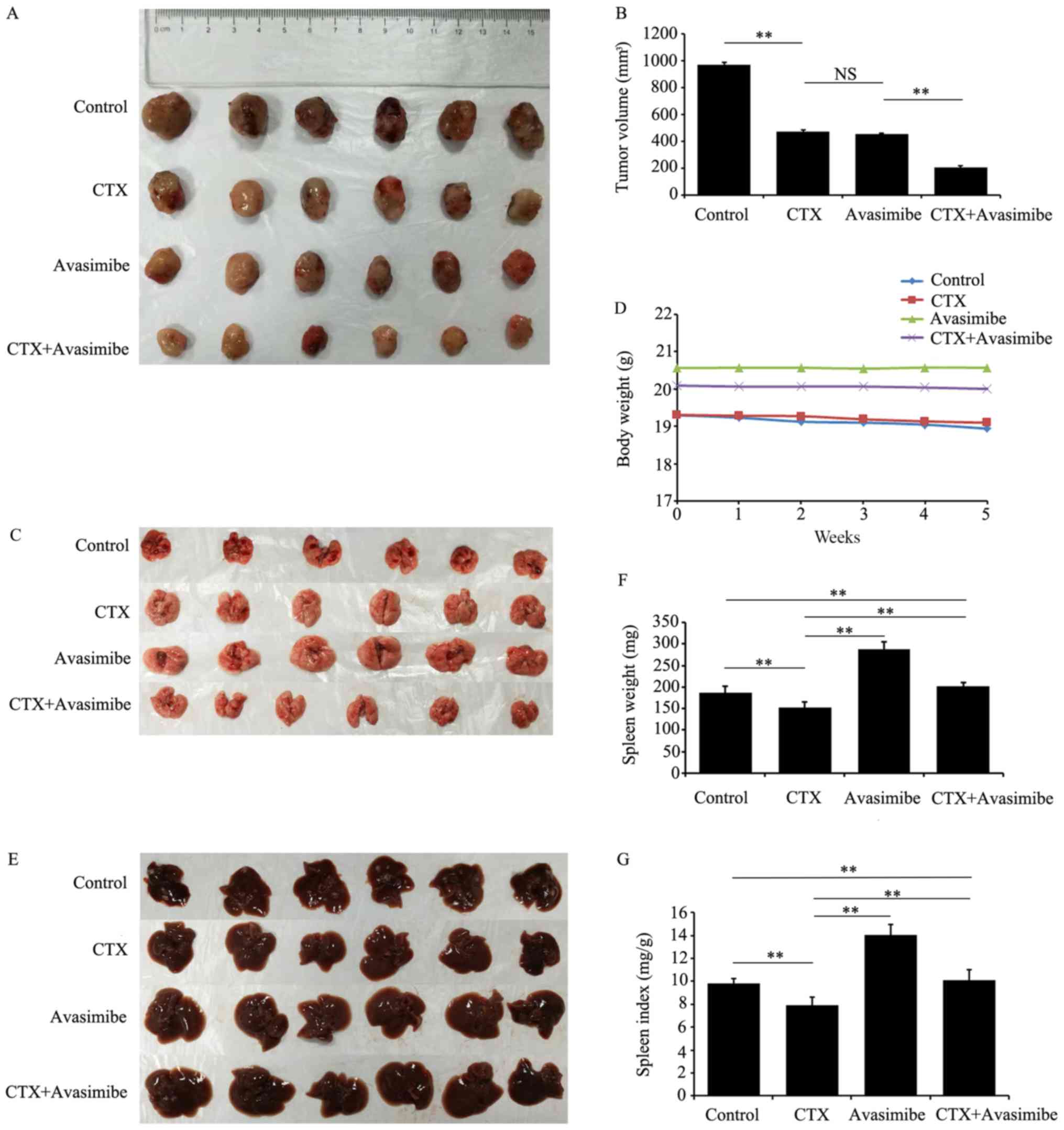
Avasimibe inhibits LLC growth and
metastasis in mice
In the present study, a subcutaneous mouse LLC model
was established in order to validate the anti-tumor effects of
ACAT-1 inhibition in vivo. The results revealed that
avasimibe may significantly reduce tumor size (P<0.01; Fig. 5A and B), and resulted in a
significant increase in spleen weights and spleen indexes
(P<0.01; Fig. 5F and G) compared
with the control groups. Compared with the CTX and avasimibe
mono-treatment groups, the combination of CTX+avasimibe was able to
significantly inhibit tumor growth more effectively than either
single treatment (P<0.01; Fig.
5B). Avasimibe treatment did not result in notable changes in
mouse body weight over time (Fig.
5C). Metastatic lesions in distant organs (liver and lung) were
also assessed at the study endpoint, with at least one metastatic
lesion being observed in the liver and lung of each control mouse,
compared with the lack of evidence of metastatic lesions in other
groups (Fig. 5D and E). These
results further confirmed that avasimibe inhibited tumor growth and
enhanced immune responses in mice with LLC tumor types.
Furthermore, avasimibe enhanced the anti-tumor effects of CTX.
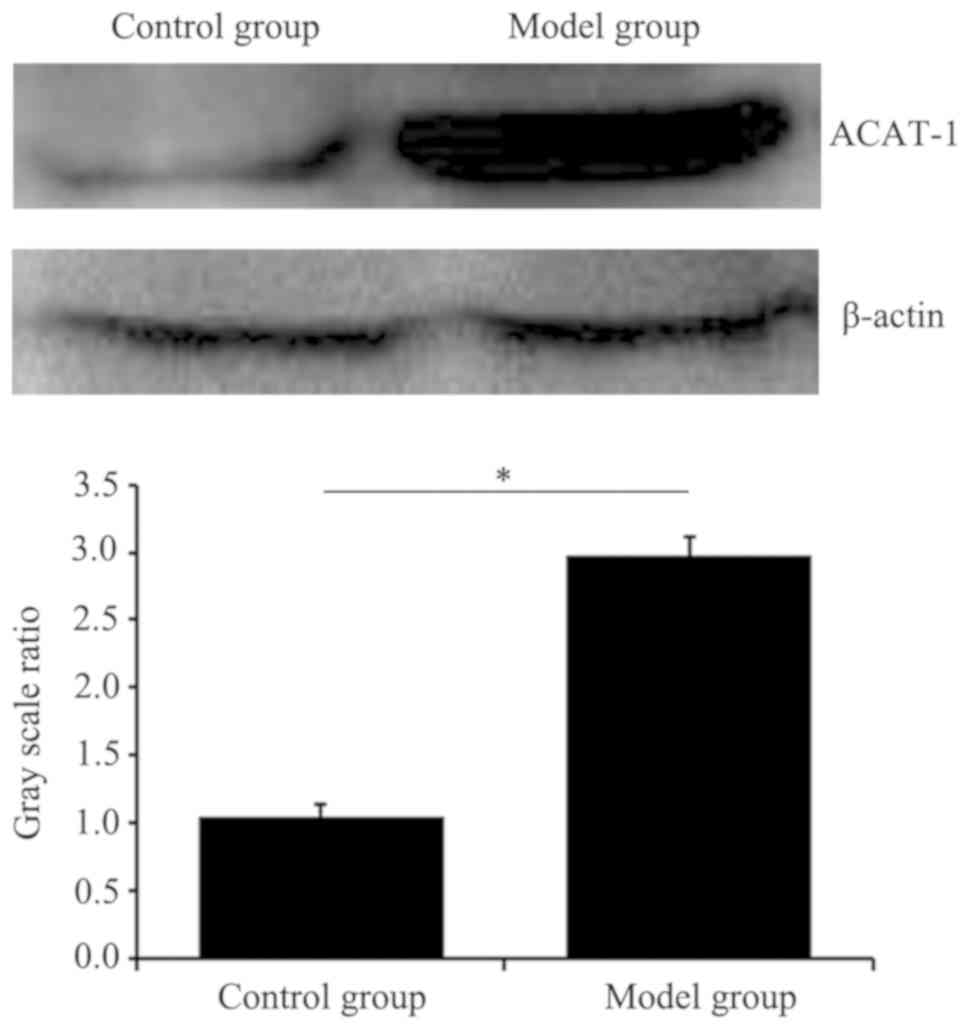
Expression of ACAT-1 in the tumor
model mice is higher compared with in the control mice
As presented in Fig.
6, the expression levels of ACAT-1 in LLC mice were
significantly higher compared with that in the control mice
(P<0.05).
Discussion
In the present study, an association between ACAT-1
and lung cancer growth and metastasis was identified. ACAT-1 has
primarily been studied in the context of cardiovascular diseases,
with numerous ACAT inhibitors having been developed to treat
diseases including cardiovascular diseases and Alzheimer's disease
(16,17). Recently, researchers have determined
that ACAT-1 expression and activity are upregulated in numerous
tumor cells, and ACAT-1 inhibitors exhibit anti-tumor activity in
certain experimental models in vivo and in vitro
(9,18–22),
including in renal cell carcinoma, colon cancer, breast cancer,
glioma, pancreatic ductal adenocarcinoma, chronic myelogenous
leukemia and lung cancer. A previous paper studied the function of
ACAT-1 in lung cancer cells, uncovering results which differed from
those of the present study (19).
This may be due to a number of reasons; firstly, this may be that
this previous study did not perform migration experiments to
investigate the effect of avasimibe on lung cancer cell migration.
Secondly, this previous study used A549 cells, which are distinct
from LLC cells. Thirdly, this previous study did not conduct animal
model experiments assessing lung cancer. Finally, this previous
study investigated the anti-tumor effects of avasimbe by detecting
cholesterol in cells, whereas the present study focused
specifically on the expression of ACAT-1. Here, it was demonstrated
that the inhibition of ACAT-1 may be effective in treating lung
cancer.
The present study provides novel insights into the
importance of ACAT-1 in LLC cells, suggesting that this protein
serves an important function in the growth and metastasis of LLC
cells and the development of LLC in mice. Targeting ACAT-1 is thus
a potential novel therapeutic strategy for treating lung cancer. By
treating LLC cells with avasimibe, an ACAT-1 inhibitor, it was
revealed that avasimibe reduced the expression of ACAT-1 and
significantly inhibited LLC cell proliferation and migration
compared with the control cells (P<0.05), further promoting the
apoptosis of these LLC cells. The present study additionally
established a mouse model of LLC to verify the anti-tumor effects
of ACAT-1 inhibitors in vivo. Mice were treated with saline,
CTX (20 mg/kg), avasimibe (15 mg/kg) or CTX+avasimibe. It was
revealed that avasimibe did not cause weight loss in mice. No
evidence of avasimibe toxicity in mice was observed, and avasimibe
alone or in combination with the existing chemotherapy CTX was
sufficient to inhibit tumor growth and metastasis, with avasimibe
further increasing tumor sensitivity to CTX. Studies have reported
that avasimibe may enhance the efficacy of gemcitabine as a means
of combatting pancreatic ductal adenocarcinoma proliferation
(22), and may additionally enhance
the efficacy of imatinib as a means of disrupting the growth of
chronic myelogenous leukemia (9).
The present study further observed that the inhibition of ACAT-1
enhanced immune responses in mice, as evidenced by measuring the
spleen weight and spleen indexes of treated mice. Additionally, it
was revealed that the expression of ACAT-1 in the lung tissue of
the LLC mice was higher compared with in healthy mice. These
results will provide the basis for further biological research to
fully understand the function of ACAT-1 in the occurrence and
development of cancer.
The mechanisms linking ACAT-1 to lung cancer growth
and metastasis require further study. One potential link is the
fact that ACAT-1 may reduce the levels of the pyruvate
dehydrogenase complex (PDC) (23).
As early as the 1950s, German scientists proposed the Warburg
effect (24); unlike the oxidative
metabolism of normal cells, tumor cells undergo extensive aerobic
glycolysis to provide the energy necessary for growth in an
oxygen-rich environment. PDC is extremely important mediator of the
interface between glycolysis and oxidative phosphorylation
(25). Studies have demonstrated
that decreased PDC levels are able to enhance glycolysis in tumor
cells, thus providing a metabolic advantage for tumor growth
(26–28). ACAT-1 is an upstream regulator of
pyruvate dehydrogenase and pyruvate dehydrogenase phosphatase
(PDP), which acetylates pyruvate dehydrogenase E1-a subunit and
PDP-1 to downregulate PDC levels (23). In the present study, the expression
of ACAT-1 was successfully inhibited using avasimibe, which in
theory may increase PDC levels and thereby inhibit tumor
growth.
In addition to promoting glycolysis, ACAT-1 serves a
key role in cholesterol metabolism, catalyzing free cholesterol
conversion into cholesterol esters (29). ACAT-1 may increase cholesterol ester
levels and elevate blood sugar in the body, resulting in insulin
secretion which may in turn increase the activity of insulin-like
growth factor (IGF). Increased IGF function may promote cell
mitosis, thereby accelerating cell proliferation and promoting
tumor growth (30–32). Studies have demonstrated that IGF is
closely associated with lung cancer progression (33,34).
Thus, the inhibition of ACAT-1 expression reduces cholesterol ester
levels, and may thereby result in decreased IGF activity, resulting
in the inactivation of phosphoinositide 3 kinase/protein kinase B
or RAS type GTPase family/mitogen-activated protein kinase
signaling pathways, thereby inhibiting tumor growth and metastasis
(35).
Recent studies (36–38) have
also identified an association between ACAT-1 and the immune
system, as inhibiting ACAT-1 expression may enhance the anti-tumor
activity of cluster of differentiation (CD)8+ T cells.
During T cell activation, lipid metabolism is required for membrane
biosynthesis and cell growth. Cholesterol regulates membrane
protein function, participates in membrane trafficking and
regulates transmembrane signaling (39,40).
Previous studies have also reported that ACAT-1 inhibitors
potentiate the anti-tumor effects of human chimeric antigen
receptor-modified T cells, and that CD8+ T cell
responses to melanoma may be enhanced by inhibiting cholesterol
esterification in mice via ACAT-1 inhibition (41,42). The
present study similarly observed that avasimibe may improve immune
responses in tumor-bearing mice, which may additionally be
associated with ACAT-1 affecting immune system function. The
avasimibe and CTX+avasimibe group exhibited increased spleen
weights and spleen index values compared with the control group,
but these values were higher in the avasimibe treatment group
compared with the CTX+avasimibe group, which may be due to the
effects of CTX on the immune system in these mice. CTX is commonly
used as an immunosuppressant to inhibit tumor growth, which may
inhibit immune system function (43–45).
Furthermore, the inhibition of ACAT-1 by avasimibe may improve the
immune function in mice (38,41,42).
Therefore, the regulation of cholesterol metabolism may have a
profound impact on anti-tumor responses through these other
signaling pathways. Further research will therefore be required to
fully elucidate the underlying molecular mechanisms.
The safety and toxicity of avasimibe and CTX have
been evaluated in clinical trials. In prior clinical trials for the
treatment of atherosclerosis, avasimibe proved to have good safety
in humans (46,47). CTX is a commonly used
chemotherapeutic drug, and its clinical safety has also been
confirmed (48–50). At present, to the best of our
knowledge there has been no clinical trial assessing the
combination of avasimibe and CTX. It is therefore necessary to
conduct clinical trials to confirm the effectiveness and safety of
this combination.
In summary, a preliminary conclusion may be drawn
from the present study; that inhibiting the expression of ACAT-1
may weaken the proliferation and metastasis of LLC cells. Avasimibe
may not only inhibit the tumor growth and development of distant
metastases in mice, but may additionally enhance the anti-tumor
efficacy of chemotherapeutic drug treatment and enhance immune
responses in vivo. Therefore, targeted blocking of the
ACAT-1 metabolism pathway has potential for use as a novel
treatment strategy for lung cancer that may be combined with
chemotherapy to provide novel treatment strategies for this
disease. This treatment has the potential to delay the progression
and metastasis of cancer, thereby prolonging patient survival time
and improving quality of life. However, the further study of ACAT-1
is still required in order to fully understand the function of
ACAT-1 in the development and progression of lung cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Medical
Research Project of Anhui Provincial Department of Health (grant
no. 09C171), the key project of Anhui Provincial University Natural
Science Research Project (grant no. KJ2016A483) and grants from the
National Natural Science Foundation of China (grant no. 21707002)
and the Graduate School of Innovation (grant no. Byycxz1722).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
MB designed the study, contributed to the conception
of the study and the analysis and intepretation of the experimental
data, revised the manuscript, and gave final approval of the
version to be published. XuxQ, WL, YL and SZ performed the
experiments. HaoZ, ZG, HW and JH performed the data analysis. HaiZ,
MS, YW, JY, XujQ and ZZ participated in developing the animal
model. All authors read and approved the manuscript and agreed to
be accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All experiments were performed in accordance with a
protocol ethically approved by the Institutional Animal Care and
Use Committee of Bengbu Medical College (Bengbu, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Zhang S, Zeng H, Zuo T,
Xia C, Yang Z and He J: Cancer incidence and mortality in China in
2013: An analysis based on urbanization level. Chin J Cancer Res.
29:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang C, Leighl NB, Wu YL and Zhong WZ:
Emerging therapies for non-small cell lung cancer. J Hematol Oncol.
12:452019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu SG and Shih JY: Management of acquired
resistance to EGFR TKI-targeted therapy in advanced non-small cell
lung cancer. Mol Cancer. 17:382018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Solomon BJ, Besse B, Bauer TM, Felip E,
Soo RA, Camidge DR, Chiari R, Bearz A, Lin CC, Gadgeel SM, et al:
Lorlatinib in patients with ALK-positive non-small-cell lung
cancer: Results from a global phase 2 study. Lancet Oncol.
19:1654–1667. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu Y, Chen CY, Li J, Cheng JX, Jang M and
Kim KH: In vitro exploration of ACAT contributions to lipid droplet
formation during adipogenesis. J Lipid Res. 59:820–829. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Allott EH, Masko EM, Freedland AR, Macias
E, Pelton K, Solomon KR, Mostaghel EA, Thomas GV, Pizzo SV, Freeman
MR, et al: Serum cholesterol levels and tumor growth in a PTEN-null
transgenic mouse model of prostate cancer. Prostate Cancer
Prostatic Dis. 21:196–203. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li J, Gu D, Lee SS, Song B, Bandyopadhyay
S, Chen S, Konieczny SF, Ratliff TL, Liu X, Xie J, et al:
Abrogating cholesterol esterification suppresses growth and
metastasis of pancreatic cancer. Oncogene. 35:6378–6388. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bandyopadhyay S, Li J, Traer E, Tyner JW,
Zhou A, Oh ST and Cheng JX: Cholesterol esterification inhibition
and imatinib treatment synergistically inhibit growth of BCR-ABL
mutation-independent resistant chronic myelogenous leukemia. PLoS
One. 12:e01795582017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ohmoto T, Nishitsuji K, Yoshitani N,
Mizuguchi M, Yanagisawa Y, Saito H and Sakashita N: K604, a
specific acyl-CoA:cholesterol acyltransferase 1 inhibitor,
suppresses proliferation of U251-MG glioblastoma cells. Mol Med
Rep. 12:6037–6042. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
de Gonzalo-Calvo D, López-Vilaró L,
Nasarre L, Perez-Olabarria M, Vázquez T, Escuin D, Badimon L,
Barnadas A, Lerma E and Llorente-Cortés V: Intratumor cholesteryl
ester accumulation is associated with human breast cancer
proliferation and aggressive potential: A molecular and
clinicopathological study. BMC Cancer. 15:4602015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang L, Kim SB, Luitel K and Shay JW:
Cholesterol depletion by TASIN-1 induces apoptotic cell death
through the ER stress/ROS/JNK signaling in colon cancer cells. Mol
Cancer Ther. 17:943–951. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao H, Wu M, Zhu L, Tian Y, Wu M, Li Y,
Deng L, Jiang W, Shen W, Wang Z, et al: Cell-penetrating
peptide-modified targeted drug-loaded phase-transformation lipid
nanoparticles combined with low-intensity focused ultrasound for
precision theranostics against hepatocellular carcinoma.
Theranostics. 8:1892–1910. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu Y, Rao SS, Wang ZX, Cao J, Tan YJ, Luo
J, Li HM, Zhang WS, Chen CY and Xie H: Exosomes from human
umbilical cord blood accelerate cutaneous wound healing through
miR-21-3p-mediated promotion of angiogenesis and fibroblast
function. Theranostics. 8:169–184. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stephenson W: Deficiencies in the National
Institute of Health's guidelines for the care and protection of
laboratory animals. J Med Philos. 18:375–388. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zanoni P, Khetarpal SA, Larach DB,
Hancock-Cerutti WF, Millar JS, Cuchel M, DerOhannessian S, Kontush
A, Surendran P, Saleheen D, et al: Rare variant in scavenger
receptor BI raises HDL cholesterol and increases risk of coronary
heart disease. Science. 351:1166–1171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang TY, Yamauchi Y, Hasan MT and Chang
C: Cellular cholesterol homeostasis in Alzheimer's disease. J Lipid
Res. 58:2239–2254. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao Z, Wu F, Ding S, Sun L, Liu Z, Ding K
and Lu J: Label-free quantitative proteomic analysis reveals
potential biomarkers and pathways in renal cell carcinoma. Tumour
Biol. 36:939–951. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee SS, Li J, Tai JN, Ratliff TL, Park K
and Cheng JX: Avasimibe encapsulated in human serum albumin blocks
cholesterol esterification for selective cancer treatment. ACS
Nano. 9:2420–2432. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Antalis CJ, Uchida A, Buhman KK and
Siddiqui RA: Migration of MDA-MB-231 breast cancer cells depends on
the availability of exogenous lipids and cholesterol
esterification. Clin Exp Metastasis. 28:733–741. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bemlih S, Poirier MD and Andaloussi AE:
Acyl-coenzyme A: cholesterol acyltransferase inhibitor Avasimibe
affect survival and proliferation of glioma tumor cell lines.
Cancer Biol Ther. 9:1025–1032. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li J, Qu X, Tian J, Zhang JT and Cheng JX:
Cholesterol esterification inhibition and gemcitabine
synergistically suppress pancreatic ductal adenocarcinoma
proliferation. PLoS One. 13:e01933182018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fan J, Shan C, Kang HB, Elf S, Xie J,
Tucker M, Gu TL, Aguiar M, Lonning S, Chen H, et al: Tyr
phosphorylation of PDP1 toggles recruitment between ACAT1 and SIRT3
to regulate the pyruvate dehydrogenase complex. Mol Cell.
53:534–548. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harris RA, Bowker-Kinley MM, Huang B and
Wu P: Regulation of the activity of the pyruvate dehydrogenase
complex. Advan Enzyme Regul. 42:249–259. 2002. View Article : Google Scholar
|
|
26
|
Zhao H, Jiang H, Li Z, Zhuang Y, Liu Y,
Zhou S, Xiao Y, Xie C, Zhou F and Zhou Y: 2-Methoxyestradiol
enhances radiosensitivity in radioresistant melanoma MDA-MB-435R
cells by regulating glycolysis via HIF-1α/PDK1 axis. Int J Oncol.
50:1531–1540. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hitosugi T, Fan J, Chung TW, Lythgoe K,
Wang X, Xie J, Ge Q, Gu T, Polakiewicz RD, Roesel JL, et al:
Tyrosine phosphorylation of mitochondrial pyruvate dehydrogenase
kinase 1 is important for cancer metabolism. Mol Cell. 44:864–877.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin R, Elf S, Shan C, Kang HB, Ji Q, Zhou
L, Hitosugi T, Zhang L, Zhang S, Seo JH, et al: 6-Phosphogluconate
dehydrogenase links oxidative PPP, lipogenesis and tumour growth by
inhibiting LKB1-AMPK signalling. Nat Cell Biol. 17:1484–1496. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Buhman KF, Accad M and Farese RV:
Mammalian acyl-CoA:cholesterol acyltransferases. Biochim Biophys
Acta. 1529:142–154. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Behjati S, Tarpey PS, Haase K, Ye H, Young
MD, Alexandrov LB, Farndon SJ, Collord G, Wedge DC, Martincorena I,
et al: Recurrent mutation of IGF signalling genes and distinct
patterns of genomic rearrangement in osteosarcoma. Nat Commun.
8:159362017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Obr AE, Kumar S, Chang YJ, Bulatowicz JJ,
Barnes BJ, Birge RB, Lazzarino DA, Gallagher E, LeRoith D and Wood
TL: Insulin-like growth factor receptor signaling in breast tumor
epithelium protects cells from endoplasmic reticulum stress and
regulates the tumor microenvironment. Breast Cancer Res.
20:1382018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cao HY, Guo XF, Zhu XF, Li SS and Zhen YS:
A ligand-based and enediyne-energized bispecific fusion protein
targeting epidermal growth factor receptor and insulin-like growth
factor-1 receptor shows potent antitumor efficacy against
esophageal cancer. Oncol Rep. 37:3329–3340. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mano SS, Uto K and Ebara M:
Material-induced Senescence (MIS): Fluidity induces senescent type
cell death of lung cancer cells via insulin-like growth factor
binding protein 5. Theranostics. 7:4658–4670. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mochizuki S, Shimoda M, Abe H, Miyamae Y,
Kuramoto J, Aramaki-Hattori N, Ishii K, Ueno H, Miyakoshi A, Kojoh
K, et al: Selective inhibition of ADAM28 suppresses lung carcinoma
cell growth and metastasis. Mol Cancer Ther. 17:2427–2438. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang H, Su X, Fang J, Xin X, Zhao X, Gaur
U, Wen Q, Xu J, Little PJ and Zheng W: Tanshinone IIA attenuates
insulin like growth factor 1-induced cell proliferation in PC12
cells through the PI3K/Akt and MEK/ERK pathways. Int J Mol Sci.
19(pii): E27192018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen X, Song Q, Xia L and Xu X: Synergy of
dendritic cell vaccines and avasimibe in treatment of head and neck
cancer in mice. Med Sci Monith. 23:4471–4476. 2017. View Article : Google Scholar
|
|
37
|
Bernard NJ: Cholesterol test for T cells.
Nat Rev Rheumatol. 15:1892019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li M, Yang Y, Wei J, Cun X, Lu Z, Qiu Y,
Zhang Z and He Q: Enhanced chemo-immunotherapy against melanoma by
inhibition of cholesterol esterification in CD8+ T
cells. Nanomedicine. 14:2541–2550. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kidani Y, Elsaesser H, Hock MB, Vergnes L,
Williams KJ, Argus JP, Marbois BN, Komisopoulou E, Wilson EB,
Osborne TF, et al: Sterol regulatory element-binding proteins are
essential for the metabolic programming of effector T cells and
adaptive immunity. Nat Immunol. 14:489–499. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Molnár E, Swamy M, Holzer M, Beck-García
K, Worch R, Thiele C, Guigas G, Boye K, Luescher IF, Schwille P, et
al: Cholesterol and sphingomyelin drive ligand-independent T-cell
antigen receptor nanoclustering. J Biol Chem. 287:42664–42674.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao L, Li J, Liu Y, Kang L, Chen H, Jin
Y, Zhao F, Feng J, Fang C, Zhu B, et al: Cholesterol esterification
enzyme inhibition enhances antitumor effects of human chimeric
antigen receptors modified T cells. J Immunother. 41:45–52.
2018.PubMed/NCBI
|
|
42
|
Yang W, Bai Y, Xiong Y, Zhang J, Chen S,
Zheng X, Meng X, Li L, Wang J, Xu C, et al: Potentiating the
antitumour response of CD8(+) T cells by modulating cholesterol
metabolism. Nature. 531:651–655. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang H, Yang S, Wang Y, Jiang T, Li S and
Lv Z: Immunoenhancement effects of glycosaminoglycan from
Apostichopus japonicus: In vitro and in
cyclophosphamide-induced immunosuppressed mice studies. Mar Drugs.
15(pii): E3472017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhu G, Jiang Y, Yao Y, Wu N, Luo J, Hu M,
Tu Y and Xu M: Ovotransferrin ameliorates the dysbiosis of
immunomodulatory function and intestinal microbiota induced by
cyclophosphamide. Food Funct. 10:1109–1122. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cheng D, Wan Z, Zhang X, Li J, Li H and
Wang C: Dietary chlorella vulgaris ameliorates altered
immunomodulatory functions in cyclophosphamide-induced
immunosuppressive mice. Nutrients. 9(pii): E7082017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chang TY, Li BL, Chang CC and Urano Y:
Acyl-coenzyme A: Cholesterol acyltransferases. Am J Physiol
Endocrinol Metab. 297:E1–E9. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Pal P, Gandhi H, Giridhar R and Yadav MR:
ACAT inhibitors: The search for novel cholesterol lowering agents.
Mini Rev Med Chem. 13:1195–1219. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sistigu A, Viaud S, Chaput N, Bracci L,
Proietti E and Zitvogel L: Immunomodulatory effects of
cyclophosphamide and implementations for vaccine design. Semin
Immunopathol. 33:369–383. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ganz PA, Romond EH, Cecchini RS, Rastogi
P, Geyer CE Jr, Swain SM, Jeong JH, Fehrenbacher L, Gross HM,
Brufsky AM, et al: Long-Term Follow-up of cardiac function and
quality of life for patients in NSABP protocol B-31/NRG oncology: A
randomized trial comparing the safety and efficacy of doxorubicin
and cyclophosphamide (AC) followed by paclitaxel with ac followed
by paclitaxel and trastuzumab in patients with node-positive breast
cancer with tumors overexpressing human epidermal growth factor
receptor 2. J Clin Oncol. 35:3942–3948. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Garderet L, Kuhnowski F, Berge B, Roussel
M, Escoffre-Barbe M, Lafon I, Facon T, Leleu X, Karlin L, Perrot A,
et al: Pomalidomide, cyclophosphamide, and dexamethasone for
relapsed multiple myeloma. Blood. 132:2555–2563. 2018. View Article : Google Scholar : PubMed/NCBI
|