Introduction
Colorectal cancer (CRC) is among the leading
malignancies in terms of incidence rate and mortality rate
worldwide (1). Despite the overall
5-year survival rate of patients with CRC increasing from 51 to
65%, ~30% of patients with stage I–III disease develop recurrent
disease following initial treatment. In addition, ≤65% of patients
with stage IV CRC relapse following curative treatment (2,3). For
patients with metastatic CRC, there is a need for novel and
individualized therapies in the third-line setting and beyond.
Despite recent advances in the development of novel targeted
therapies, there remains an unmet need to exploit oncogenic drivers
of CRC and overcome acquired resistance. In addition, more
efficient prognostic markers should be established, which can
inform treatment decisions to avoid under- or over-treatment and to
guide the intensity of patient follow-up.
Long non-coding RNAs (lncRNAs) belong to a large
group of non-coding RNAs, and are generally classified as
transcripts that are 200 nt to 100 kb long, which lack the
open-reading frame (4). lncRNA
molecules are involved in diverse biological processes, including
epigenetic regulation via molecular scaffolding, regulation of mRNA
processing, molecular decoying and lncRNA-derived peptides through
diverse mechanisms (5,6). Notably, lncRNAs with highly specific
expression patterns in certain tissues and diseases, including
various types of cancer, have gained increasing interest as
molecular targets for therapy (7,8). Bladder
cancer-associated transcript 2 (BLACAT2) is a novel lncRNA that was
identified by He et al (9),
who demonstrated that BLACAT2 is markedly upregulated in lymph node
metastatic bladder cancer and that BLACAT2 is correlated with lymph
node metastasis. Additionally, overexpression of BLACAT2 promotes
bladder cancer-associated lymphangiogenesis and lymphatic
metastasis in cultured bladder cancer cell lines and mouse models
(9). However, the expression pattern
and functional role of BLACAT2 in other types of cancer, including
CRC, remain unknown.
The present study aimed to detect the expression of
BLACAT2 in CRC cells and tissues. The clinical significance of
BLACAT2 in CRC was explored. Additionally, the prognostic value of
BLACAT2 was investigated. Functionally, the role of BLACAT2 in
migration, invasion, proliferation and chemoresistance of CRC cells
was investigated.
Materials and methods
Cell culture and transfection
A human intestinal epithelial cell line (HIEC-6) and
CRC cell lines, including LOVO, SW620, HCT15, HCT116, SW480 and
DLD1, were purchased from American Type Culture Collection
(Manassas, VA, USA). LOVO and HCT15 cells were cultivated in
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), and SW620, HCT116, SW480 and
DLD1 cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.). Media were supplemented with 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 mg/ml streptomycin and cells were cultured at
37°C with 5% CO2. HIEC-6 cells were cultured in
OPTIMEM-GlutaMAX (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 5% FBS, 10 mM HEPES (Invitrogen; Thermo Fisher
Scientific, Inc.) and 5 ng/ml epidermal growth factor (BD
Biosciences, San Jose, CA, USA) at 37°C with 5% CO2.
BLACAT2 interference plasmids [pcDNA3.1-short hairpin RNA
(sh)BLACAT2#1 and pcDNA3.1-shBLACAT2#2], a BLACAT2 overexpression
plasmid (pcDNA3.1-BLACAT2) and control plasmids (pcDNA3.1-shControl
and pcDNA3.1-Vector) were synthesized by Shanghai OE Biotech Co.,
Ltd. (Shanghai, China). Cells at 30–50% confluence were transfected
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) in accordance with the manufacturer's protocols.
The mass of plasmids was 0.8 µg. Cells were cultured for 48 h after
transfection and harvested to determine transfection efficiency by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR), and then subjected to subsequent experimentation.
Clinical specimens
CRC tissues and paired normal colorectal tissues
(n=127) were collected between February 2012 and May 2016 at The
Affiliated Hospital of North Sichuan Medical College (Nanchong,
China). Normal tissues were located ≥2 cm away from the cancer
tissues. All patients received surgical resection without
neoadjuvant therapies. Patient characteristics are presented in
Table I. Tumor-Node-Metastasis (TNM)
staging was performed according to the American Joint Committee on
Cancer guidelines (10). Tissue
specimens were immediately snap-frozen and stored in liquid
nitrogen until RNA extraction. Written informed consent was
obtained from all patients. The present study was approved by the
Ethics Committee of The Affiliated Hospital of North Sichuan
Medical College. Follow-up was carried out regularly at the
outpatient clinic. The end of the follow-up period was December
2017.
 | Table I.Association between BLACAT2 expression
and clinicopathological characteristics of patients with colorectal
cancer. |
Table I.
Association between BLACAT2 expression
and clinicopathological characteristics of patients with colorectal
cancer.
|
|
| BLACAT2 |
|
|---|
|
|
|
|
|
|---|
| Parameters | Number (n=127) | Low (n=66) | High (n=61) | P-value |
|---|
| Age |
|
|
| 0.396 |
| <60
years | 57 | 32 | 25 |
|
| ≥60
years | 70 | 34 | 36 |
|
| Sex |
|
|
| 0.122 |
| Male | 68 | 31 | 37 |
|
|
Female | 59 | 35 | 24 |
|
| CEA |
|
|
| 0.803 |
| <4.5
µg/ml | 61 | 31 | 30 |
|
| >4.5
µg/ml | 66 | 35 | 31 |
|
| Tumor location |
|
|
| 0.369 |
|
Colon | 49 | 23 | 26 |
|
|
Rectum | 78 | 43 | 35 |
|
| Differentiation
grade |
|
|
| 0.146 |
| Well +
Moderate | 75 | 43 | 32 |
|
| Poor +
Undifferentiated | 52 | 23 | 29 |
|
| Tumor size |
|
|
| 0.029 |
| <5
cm | 50 | 32 | 18 |
|
| ≥5
cm | 77 | 34 | 43 |
|
| T stage |
|
|
| 0.674 |
|
T1+T2 | 67 | 36 | 31 |
|
|
T3+T4 | 60 | 30 | 30 |
|
| N stage |
|
|
| <0.001 |
| N0 | 76 | 51 | 27 |
|
| N1 | 51 | 15 | 34 |
|
| M stage |
|
|
| 0.028 |
| M0 | 108 | 65 | 53 |
|
| M1 | 9 | 1 | 8 |
|
| TNM stage |
|
|
| <0.001 |
|
I+II | 74 | 50 | 24 |
|
|
III+IV | 53 | 16 | 37 |
|
RT-qPCR assay
Total RNA from CRC cells and tissues was isolated
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. Total
RNA (1 µg) was used as a template for cDNA synthesis using a
PrimeScript RT Reagent kit with gDNA Eraser (Takara Biotechnology
Co., Ltd., Dalian, China). The temperature protocol for RT was as
follows: 25°C for 5 min, followed by 42°C for 60 min and 70°C for 5
min. qPCR was performed using SYBR Premix Ex Taq (Takara
Biotechnology Co., Ltd.). Briefly, qPCR was performed under the
following conditions: 95°C for 5 min, followed by 40 cycles of
denaturation at 95°C for 30 sec, annealing at 60°C for 1 min and
extension at 70°C for 1 min. All RT-qPCR assays were performed on
an ABI 7900 system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). GAPDH was utilized as an internal control. The primer
sequences used were as follows: BLACAT2, forward
5′-GAGAGGGATCCACTCATCCA-3′, reverse 5′-GGAAACAAAGGCAGAGCTT-3′; and
GAPDH, forward 5′-CACCCACTCCTCCACCTTTG-3′ and reverse
5′-CCACCACCCTGTTGCTGTAG-3′. The 2−ΔΔCq method was used
to calculate the relative expression levels of BLACAT2 (11). Each experiment for the detection of
mRNA expression levels in cells was repeated three times
independently with triplicates.
Transwell migration and invasion
assays
CRC cells (DLD1 and SW620) with BLACAT2 knockdown or
overexpression and corresponding control cells were subjected to
Transwell migration and invasion assays, to evaluate the effect of
BLACAT2 on modulating migratory and invasive abilities. Briefly,
cells (3×104) were suspended in serum-free RPMI-1640
medium (200 µl) and seeded in the upper Transwell chamber (8 µm
pore size; BD Biosciences, San Jose, CA, USA) with (for invasion
assay) or without (for migration assay) Matrigel (BD Biosciences)
precoating. Medium containing 20% FBS (700 µl) was added to the
lower chamber to serve as a chemoattractant. Subsequently, the
cells were incubated for 36 h at 37°C. The cells in the upper
chamber were then removed, and the migratory or invasive cells in
the lower chamber were fixed with methanol for 20 min at room
temperature, stained with 0.1% crystal violet solution for 20 min
at room temperature and images were captured using a light
microscope. Cells in five random fields were counted and the
average number of migratory/invasive cells was utilized for
comparison. Each experiment was repeated three times independently
with triplicates.
Colony formation assay
Following transfection, CRC cells (500 cells/well)
were seeded in 6-well plates and incubated for 10 days.
Subsequently, colonies were fixed with methanol for 15 min and
stained with 0.1% crystal violet for 15 min at room temperature.
Colonies with diameters >1 mm were counted under a light
microscope (Olympus Corporation, Tokyo, Japan). Each experiment was
repeated three times independently with triplicates.
MTT assay
An MTT assay was used for cell proliferation and
cell inhibition rate analysis. CRC cells (3,000 cells/well) were
plated in 96-well plates and transfection was performed.
Subsequently, MTT (20 µl; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) was added to each well at different time points (0, 24,
48, 72 and 96 h) and plates were incubated for an additional 4 h in
a humidified incubator at 37°C. Dimethyl sulfoxide (100 µl/well)
was added to each well to dissolve the formazan crystals followed
by agitation of the plates for 10 min at room temperature. Optical
density was measured at 492 nm using a microplate spectrophotometer
(BioTek Instruments, Inc., Winooski, VT, USA). The cell inhibition
rates were generated based on the cell viability following
culturing of cells for 72 h with different concentrations (0, 10,
20, 30, 40 and 50 µg/ml) of 5-FU. Each experiment was repeated
three times independently with triplicates.
Establishment of 5-fluorouracil
(5-FU)-resistant cell lines
DLD1 and SW620 CRC cells at a confluence of 70% were
treated with 5-FU (5 µg/ml; Abcam, Cambridge, MA, USA) at room
temperature. The surviving cells were cultured until they reached
80% confluence and were passaged twice. Sequentially increasing
concentrations of 5-FU (5 µg/ml) were used to select 5-FU-resistant
cells, and the concentration was finally maintained at 30 µg/ml for
2 months. 5-FU-resistant cells were used to evaluate the expression
levels of BLACAT2 by RT-qPCR.
Statistical analysis
Data are expressed as the means ± standard deviation
and were analyzed using SPSS v19.0 software (IBM Corp., Armonk, NY,
USA). Each experiment was performed in triplicate. A χ2
test was used to explore the associations between classified
variables. The differences between two independent groups were
analyzed with a Student's t-test. One-way analysis of variance
followed by a least significance difference post hoc test was used
to examine differences among three or more groups. To determine the
cut-off value for distinguishing the specimens with high or low
BLACAT2 expression, a receiver operating characteristic (ROC) curve
was used. The point on the curve with the shortest distance to the
coordinate (0,1) was selected as the threshold value to classify
cases as high or low expression. The Kaplan-Meier method was used
to depict the overall survival (OS) curves, and the difference
between the two groups was examined using the log-rank test.
Univariate and multivariate analyses were conducted to analyze the
risk factors for poor prognosis. P<0.05 was considered to
indicate a statistically significant difference.
Results
Overexpression of BLACAT2 is
associated with CRC clinical progression
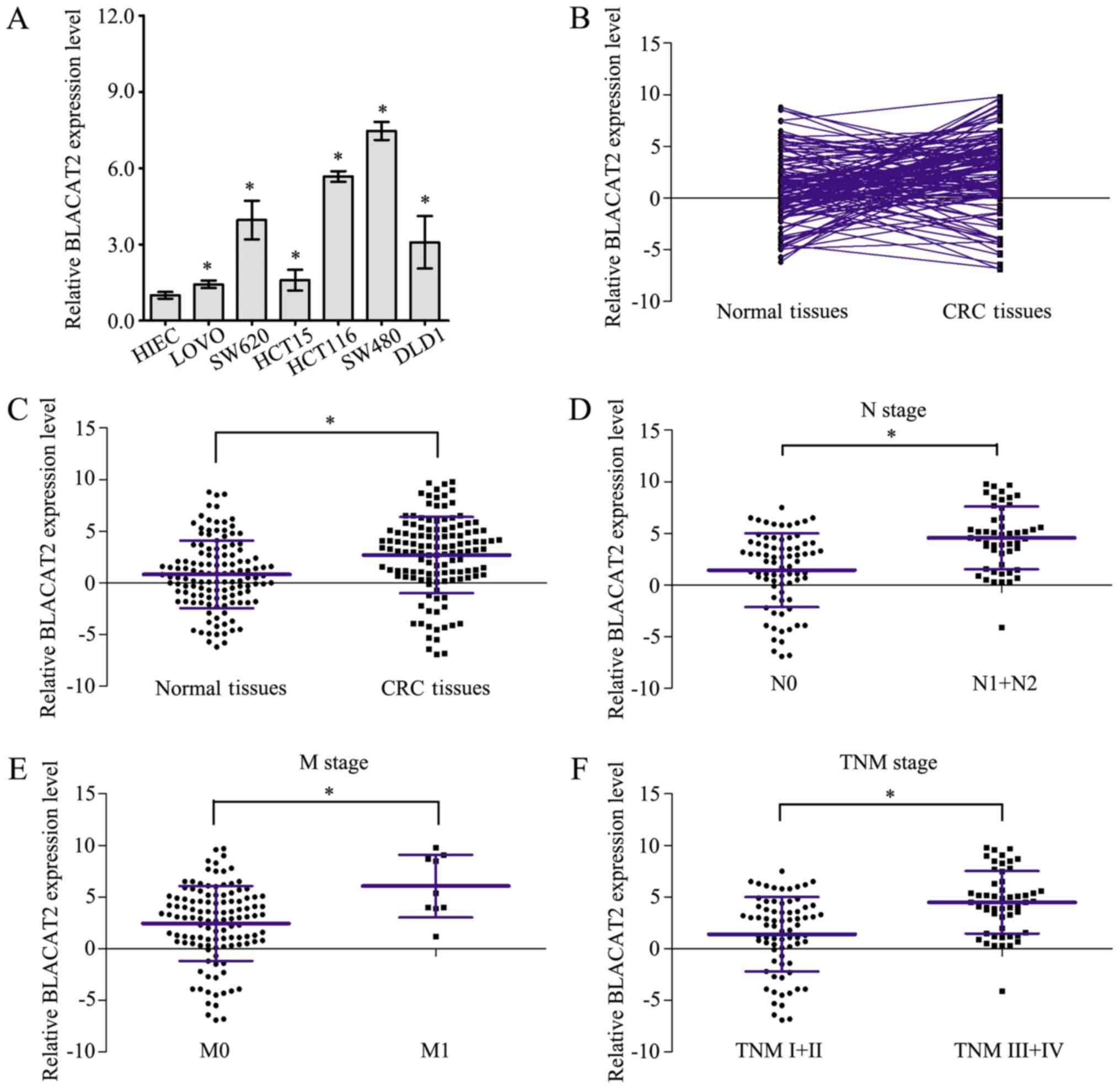
Measurement of BLACAT2 expression in CRC cells by
RT-qPCR revealed that BLACAT2 was markedly upregulated in six CRC
cell lines compared with in HIEC cells (Fig. 1A). Furthermore, expression of BLACAT2
was detected by RT-qPCR in 127 CRC tissues and paired normal
tissues (Fig. 1B). As shown in
Fig. 1C, CRC tissues exhibited
higher BLACAT2 expression than paired normal tissues. In addition,
BLACAT2 expression levels were statistically compared in CRC
tissues of different clinical stages, and the results confirmed
that CRC tissues of node (N)1/2 stage, metastasis (M)1 stage and
TNM III/IV stage exhibited higher BLACAT2 expression than CRC
tissues of N0 stage (Fig. 1D), M0
stage (Fig. 1E) and TNM I/II stage
(Fig. 1F), respectively. These
results demonstrated that BLACAT2 was overexpressed in CRC and may
be associated with CRC clinical progression.
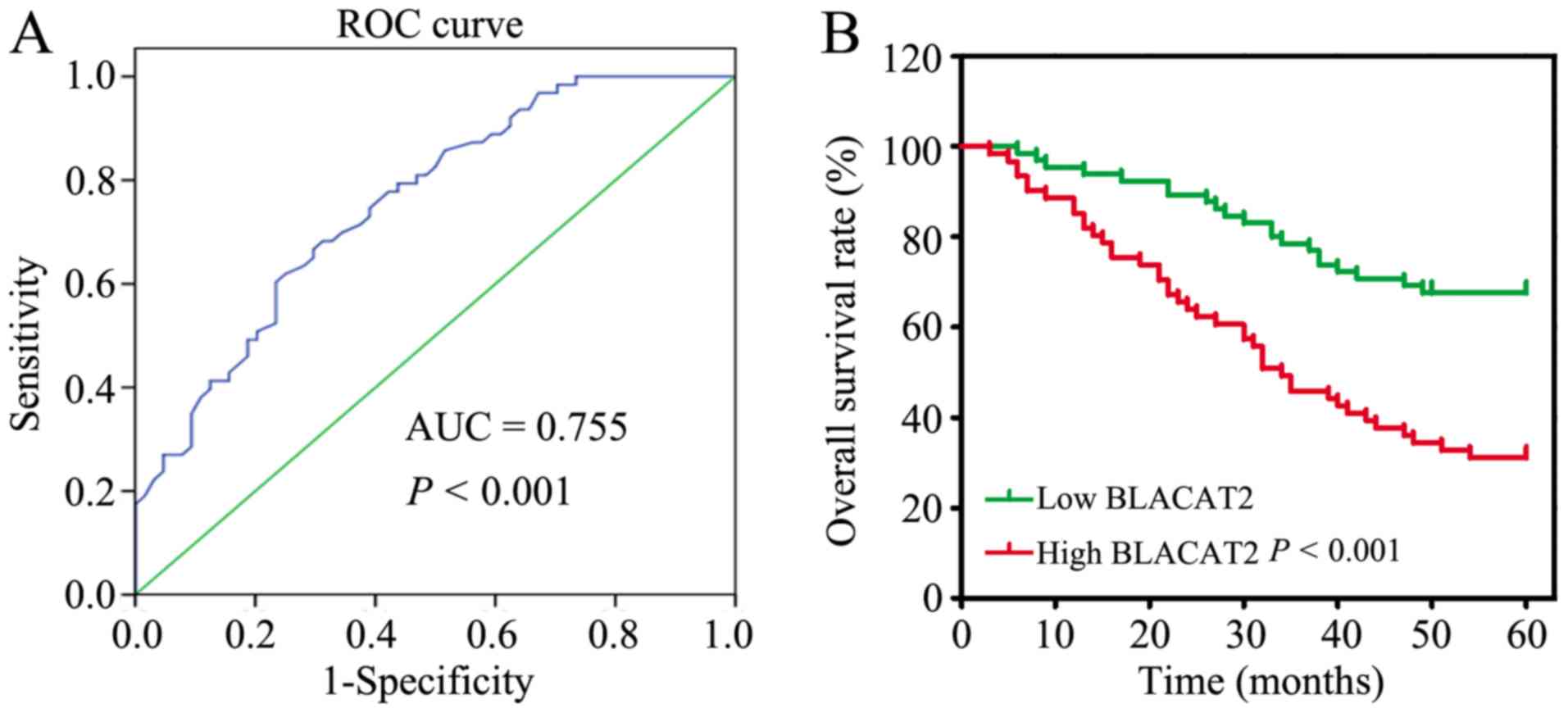
High BLACAT2 expression predicts poor
OS in patients with CRC
To further verify the clinical significance of
BLACAT2 in CRC, the 127 patients with CRC were divided into two
groups: A low BLACAT2 expression group (n=66) and a high BLACAT2
expression group (n=61), based on the cutoff value (3.35)
determined by ROC curve analysis (Fig.
2A). Subsequently, the associations between the
clinicopathological features and BLACAT2 expression were
statistically analyzed. The results revealed that high BLACAT2
expression was significantly associated with larger tumor size
(P=0.029) and more advanced N stage (P<0.001), M stage (P=0.028)
and TNM stage (P<0.001; Table I).
In addition, the association between BLACAT2 expression and the OS
rate was analyzed, which demonstrated that patients with high
BLACAT2 expression had a poor OS rate compared with patients with
low BLACAT2 expression (P<0.001; Fig.
2B).
The risk factors for poor prognosis of CRC were
identified by univariate analysis, and it was revealed that
advanced N stage (P<0.001), M stage (P<0.001), TNM stage
(P<0.001) and high BLACAT2 expression were risk factors for poor
CRC prognosis (Table II). Further
analysis of these factors with multivariate analysis identified
that high BLACAT2 expression was an independent risk factor for CRC
OS (hazard ratio=1.164, 95% confidence interval=1.062–1.277,
P=0.001; data not shown). In summary, BLACAT2 may be a promising
prognostic marker for CRC.
| Table II.Statistical analysis of risk factors
for overall survival of patients with colorectal cancer. |
Table II.
Statistical analysis of risk factors
for overall survival of patients with colorectal cancer.
| Parameters | Hazard ratio | 95% Confidence
interval | P-value |
|---|
| Age (<60 years
vs. ≥60 years) | 0.800 | 0.488–1.313 | 0.378 |
| Sex (male vs.
female) | 0.951 | 0.580–1.560 | 0.844 |
| CEA level (<4.5
µg/ml vs. >4.5 µg/ml) | 1.631 | 0.987–2.695 | 0.056 |
| Tumor location
(colon vs. rectum) | 1.013 | 0.609–1.686 | 0.959 |
| Differentiation
(Poor + Undifferentiated vs. Well + Moderate) | 1.530 | 0.933–2.508 | 0.092 |
| Tumor size (≥5 cm
vs. 5 cm) | 0.743 | 0.442–1.248 | 0.261 |
| T stage (T1+T2 vs.
T3+T4) | 0.998 | 0.609–1.637 | 0.994 |
| N stage (N1 vs.
N0) | 4.917 | 2.918–8.285 | <0.001 |
| M stage (M1 vs.
M0) | 5.751 | 2.731–12.110 | <0.001 |
| TNM stage (III+IV
vs. I+II) | 5.422 | 3.179–9.247 | <0.001 |
| BLACAT2 (high vs.
low) | 1.273 | 1.165–1.391 | <0.001 |
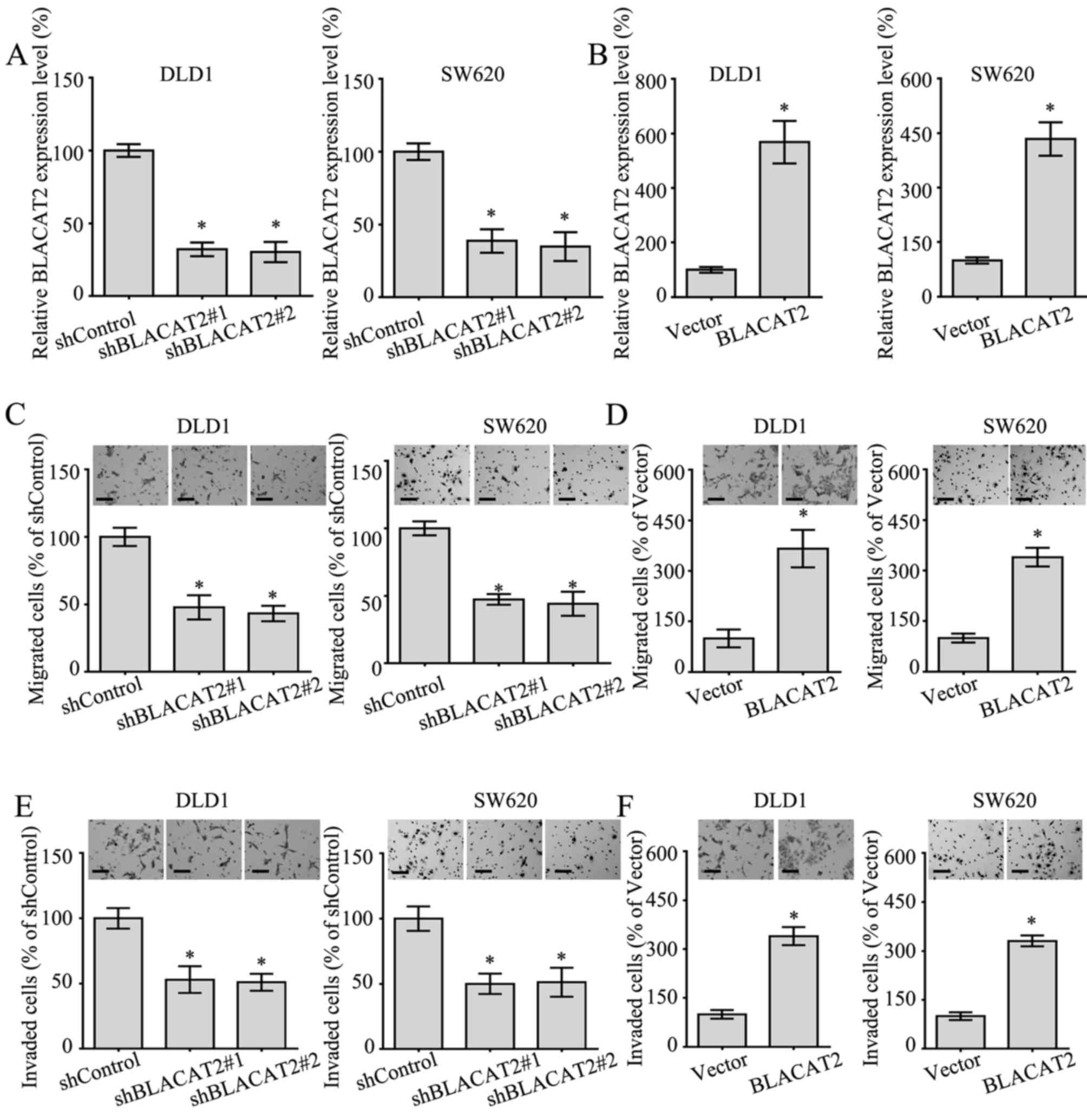
lncRNA BLACAT2 accelerates the
migration and invasion of CRC cells
To better understand the functional role of BLACAT2
in CRC, BLACAT2 was silenced or ectopically overexpressed in DLD1
and SW620 cells (Fig. 3A and B),
since the expression levels of BLACAT2 in these two cells lines
were relatively moderate (Fig. 1A).
In addition, the migratory and invasive abilities of CRC cells
overexpressing BLACAT2, or in which BLACAT2 was silenced, were
detected by Transwell migration and invasion assays. As expected,
the interference of BLACAT2 in DLD1 and SW620 cells notably
suppressed migration in the Transwell assay (Fig. 3C). Accordingly, the migration of DLD1
and SW620 cells with BLACAT2 overexpression was markedly increased
(Fig. 3D). In addition, the invasive
ability of DLD1 cells and SW620 cells exhibited a positive
association with BLACAT2 levels (Fig. 3E
and F). Overall, BLACAT2 promoted the migration and invasion of
CRC cells.
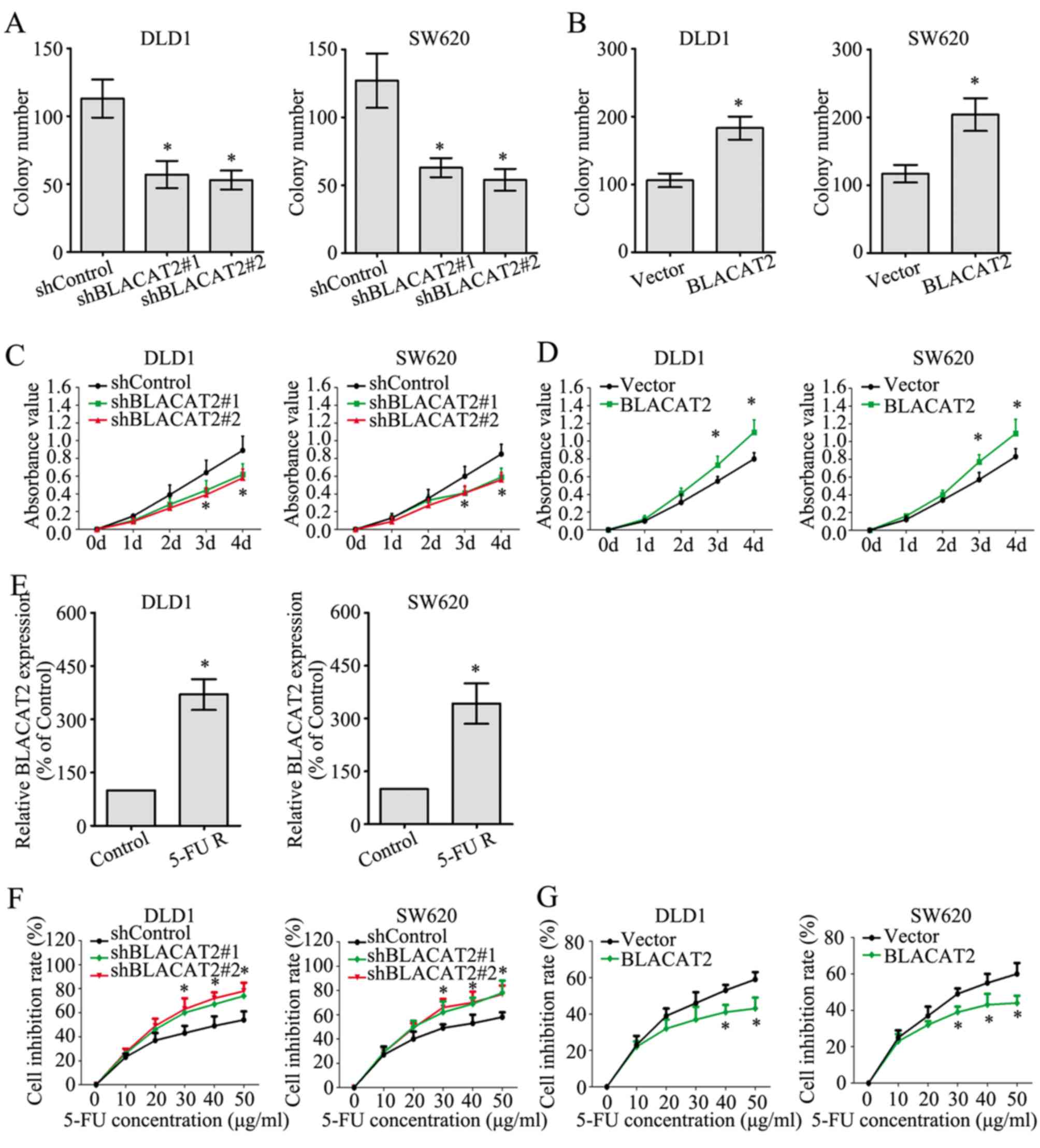
lncRNA BLACAT2 contributes to the
proliferation and chemoresistance of CRC cells
The effect of BLACAT2 on CRC cell proliferation was
investigated by colony formation and MTT assays. The colony numbers
were greatly decreased in DLD1 and SW620 cells with BLACAT2
deficiency (Fig. 4A), whereas the
colony numbers were markedly increased following BLACAT2
overexpression (Fig. 4B). In
accordance with the colony formation assay, the MTT assay also
demonstrated the role of BLACAT2 in promoting the proliferation of
CRC cells (Fig. 4C and D).
Chemoresistance is a primary factor influencing the
prognosis of patients with CRC, and the potential role of BLACAT2
in CRC chemoresistance was determined in the present study. First,
5-FU-resistant DLD1 and SW620 cells were established. In addition,
the 5-FU-resistant DLD1 and SW620 cells exhibited much higher
BLACAT2 expression than the untreated control DLD1 and SW620 cells,
respectively. (Fig. 4E).
Additionally, interference of BLACAT2 in DLD1 and SW620 cells
significantly increased the cell inhibition rate as the
concentration of 5-FU increased when cells were treated with 5-FU
for 2 days (Fig. 4F). By contrast,
the cell inhibition rates were reduced in CRC cells overexpressing
BLACAT2 when treated with 5-FU for 2 days (Fig. 4G). These findings suggested that
BLACAT2 contributed to the proliferation and chemoresistance of CRC
cells.
Discussion
The function of lncRNAs has been extensively
investigated in recent years, and lncRNAs are emerging from the
‘desert region’ of the genome as a novel source of biomarkers to
characterize disease recurrence and progression, since a number of
lncRNAs have restricted species-specific and cancer-specific
expression patterns (12). The first
CRC-associated lncRNA was identified in 1996 when Hibi et al
(13) reported that the endogenous
H19 imprinted maternally expressed transcript (H19) gene is
frequently abundant in CRC specimens, and overexpression of H19
serves an important role in CRC development. A recent study
demonstrated that lncRNAs serve pivotal roles in invasion,
metastasis, early diagnosis, prognosis, chemoresistance and
radioresistance of CRC (14). Since
the 1950s, 5-FU-based chemotherapy has been a fundamental method
for the treatment of patients with CRC (15). However, a previous study revealed
that almost half of all patients with metastatic CRC are resistant
to 5-FU-based chemotherapy (16).
Therefore, further investigations to reveal the chemoresistant
mechanisms in CRC are required. The results of the present study
revealed a potential role of BLACAT2 in contributing to
chemoresistance of CRC cells.
Mechanistically, lncRNAs affect CRC progression by
functioning as precursors of microRNAs or competing with endogenous
RNAs, interacting with proteins and regulating epigenetic
mechanisms (17). For example, the
lncRNA metastasis-associated lung adenocarcinoma transcript 1 can
competitively bind to the tumor suppressor splicing factor proline
and glutamine rich (SFPQ) gene and release SFPQ from the
SFPQ/polypyrimidine tract binding protein 2 complex, thus promoting
the growth and migration of CRC cells (18). The lncRNA colorectal neoplasia
differentially expressed may regulate the progression and
chemoresistance of CRC by binding microRNA (miRNA)-181a-5p and
modulating its expression and subsequently mediating the activity
of the Wnt/β-catenin signaling pathway (19). The lncRNA ubiquitin-like with plant
homeodomain and really interesting new gene finger domains 1
protein associated transcript may interact with and stabilize the
epigenetic factor ubiquitin like with PHD and ring finger domains 1
by interfering with its β-transducin repeat-containing protein
mediated ubiquitination, which promotes the survival and
tumorigenicity of CRC cells (20).
It has been verified that BLACAT2 is upregulated in
bladder cancer, is correlated with lymph node metastasis, and
induces lymphangiogenesis in vitro and in vivo
(9). In addition, BLACAT2 was
demonstrated to epigenetically increase vascular epidermal growth
factor C (VEGF-C) expression by directly associating with WD repeat
domain 5 (WDR5), a core subunit of the human H3K4 methyltransferase
complexes (9). Therefore, expression
of VEGF-C and WDR5 could be further investigated in CRC cells with
BLACAT2 overexpression or knockdown to elucidate the possible
mechanisms of BLACAT2. In addition, numerous mechanisms have been
reported in the field of lncRNA-mediated cancer biology, including
influencing chromatin state and methylation, and the stability of
proteins and complexes or by acting as a sponge for miRNA
inhibition (21). Epigenetic
remodeling is frequently achieved through the interaction of a
lncRNA with polycomb repressive complex 2, a protein from the
polycomb complex that induces chromatin inactivation by
establishing inhibitory H3K427me3 histone signals (22,23).
Certain lncRNAs recruit DNA methyltransferases directly to modify
chromatin conformation, or they modify nucleosome positioning. In
addition, numerous lncRNAs exert their oncogenic function through
direct interaction with proteins or protein complexes as scaffolds
or allosteric activators/inhibitors (24–28).
Furthermore, some lncRNAs have recently been revealed to act as
competing endogenous RNAs by binding miRNAs (‘sponging’), and
reducing their inhibitory effect on their natural targets (29–31).
Potential targeting miRNAs could be predicted with bioinformatics
methods, and further experiments, such as a dual luciferase assay,
are required to verify the potential targets. Information regarding
BLACAT2 is limited since it is a novel lncRNA; therefore, further
investigations are required to determine its significance.
The present study aimed to define the clinical
significance and functional role of BLACAT2 in CRC. Notably,
BLACAT2 may function as an oncogene in CRC and may be associated
with CRC clinicopathological progression as reported in bladder
cancer (9). To determine the cutoff
value of the BLACAT2 expression level, a ROC curve was utilized.
Notably, the area under the curve was 0.755, with a P-value
<0.001, which suggested that BLACAT2 may be a promising
diagnostic marker for CRC. Based on the cutoff value obtained from
ROC curve analysis, statistical analysis of clinicopathological
features was carried out to compare the high and low BLACAT2
expression groups. Notably, high BLACAT2 expression was associated
with larger tumor size (P=0.029), more advanced N stage
(P<0.001), M stage (P=0.028), TNM stage (P<0.001) and poor OS
(P<0.001), all of which are fundamental factors in evaluating
the clinical progression of CRC. Additionally, univariate and
multivariate analyses further identified high BLACAT2 expression as
an independent survival risk factor for patients with CRC
(P=0.001), which suggested a prognostic value of BLACAT2 in
patients with CRC. Furthermore, BLACAT2 was demonstrated to promote
the proliferation, metastasis and chemoresistance of CRC cells,
which further confirmed the pivotal role of BLACAT2 in CRC
biology.
In conclusion, the present study verified the
oncogenic role of BLACAT2 in CRC. However, the detailed mechanisms
by which BLACAT2 promoted CRC progression and chemoresistance
require further exploration, and more evidence is required to
confirm the results of the present study. The results suggested
that BLACAT2 may be a potential prognostic marker and therapeutic
target in CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Nanchong
Natural Science Fund Project (grant no. 16YFZJ0054) and the Sichuan
Provincial Natural Science Foundation (grant no. 17ZB0169).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YR designed the study, measured the expression of
BLACAT2 in tissues, performed some of the in vitro
experiments and revised the manuscript. CZ and YH collected the
tissues, conducted some of the in vitro assays and drafted
the manuscript. HX and XM performed the statistical analysis of the
data and conducted the follow-up of the patients. All authors
approved the final manuscript.
Ethics approval and consent to
participate
Written informed consent was obtained from all
patients. The present study was approved by the Ethics Committee of
The Affiliated Hospital of North Sichuan Medical College.
Patient consent for publication
Written informed consent was obtained from all
patients.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Esin E and Yalcin S: Maintenance strategy
in metastatic colorectal cancer: A systematic review. Cancer Treat
Rev. 42:82–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
van der Stok EP, Spaander MCW, Grunhagen
DJ, Verhoef C and Kuipers EJ: Surveillance after curative treatment
for colorectal cancer. Nat Rev Clin Oncol. 14:297–315. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jarroux J, Morillon A and Pinskaya M:
History, discovery, and classification of lncRNAs. Adv Exp Med
Biol. 1008:1–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rinn JL and Chang HY: Genome regulation by
long noncoding RNAs. Annu Rev Biochem. 81:145–166. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kwok ZH and Tay Y: Long noncoding RNAs:
Lincs between human health and disease. Biochem Soc Trans.
45:805–812. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rao AKDM, Rajkumar T and Mani S:
Perspectives of long non-coding RNAs in cancer. Mol Biol Rep.
44:203–218. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chandra Gupta S and Nandan Tripathi Y:
Potential of long non-coding RNAs in cancer patients: From
biomarkers to therapeutic targets. Int J Cancer. 140:1955–1967.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He W, Zhong G, Jiang N, Wang B, Fan X,
Chen C, Chen X, Huang J and Lin T: Long noncoding RNA BLACAT2
promotes bladder cancer-associated lymphangiogenesis and lymphatic
metastasis. J Clin Invest. 128:861–875. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Weiser MR: AJCC 8th edition: Colorectal
cancer. Ann Surg Oncol. 25:1454–1455. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Han D, Wang M, Ma N, Xu Y, Jiang Y and Gao
X: Long noncoding RNAs: Novel players in colorectal cancer. Cancer
Lett. 361:13–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hibi K, Nakamura H, Hirai A, Fujikake Y,
Kasai Y, Akiyama S, Ito K and Takagi H: Loss of H19 imprinting in
esophageal cancer. Cancer Res. 56:480–482. 1996.PubMed/NCBI
|
|
14
|
Luo J, Qu J, Wu DK, Lu ZL, Sun YS and Qu
Q: Long non-coding RNAs: A rising biotarget in colorectal cancer.
Oncotarget. 8:22187–22202. 2017.PubMed/NCBI
|
|
15
|
Showalter SL, Showalter TN, Witkiewicz A,
Havens R, Kennedy EP, Hucl T, Kern SE, Yeo CJ and Brody JR:
Evaluating the drug-target relationship between thymidylate
synthase expression and tumor response to 5-fluorouracil. Is it
time to move forward? Cancer Biol Ther. 7:986–994. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Douillard JY, Cunningham D, Roth AD,
Navarro M, James RD, Karasek P, Jandik P, Iveson T, Carmichael J,
Alakl M, et al: Irinotecan combined with fluorouracil compared with
fluorouracil alone as first-line treatment for metastatic
colorectal cancer: A multicentre randomised trial. Lancet.
355:1041–1047. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Weng M, Wu D, Yang C, Peng H, Wang G, Wang
T and Li X: Noncoding RNAs in the development, diagnosis, and
prognosis of colorectal cancer. Transl Res. 181:108–120. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ji Q, Zhang L, Liu X, Zhou L, Wang W, Han
Z, Sui H, Tang Y, Wang Y and Liu N: Long non-coding RNA MALAT1
promotes tumour growth and metastasis in colorectal cancer through
binding to SFPQ and releasing oncogene PTBP2 from SFPQ/PTBP2
complex. Br J Cancer. 111:736–748. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Taniue K, Kurimoto A, Sugimasa H, Nasu E,
Takeda Y, Iwasaki K, Nagashima T, Okada-Hatakeyama M, Oyama M,
Kozuka-Hata H, et al: Long noncoding RNA UPAT promotes colon
tumorigenesis by inhibiting degradation of UHRF1. Proc Natl Acad
Sci USA. 113:1273–1278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bartonicek N, Maag JL and Dinger ME: Long
noncoding RNAs in cancer: Mechanisms of action and technological
advancements. Mol Cancer. 15:432016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ringrose L, Ehret H and Paro R: Distinct
contributions of histone H3 lysine 9 and 27 methylation to
locus-specific stability of polycomb complexes. Mol Cell.
16:641–653. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Davidovich C and Cech TR: The recruitment
of chromatin modifiers by long noncoding RNAs: Lessons from PRC2.
RNA. 21:2007–2022. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takayama K, Horie-Inoue K, Katayama S,
Suzuki T, Tsutsumi S, Ikeda K, Urano T, Fujimura T, Takagi K,
Takahashi S, et al: Androgen-responsive long noncoding RNA CTBP1-AS
promotes prostate cancer. EMBO J. 32:1665–1680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li Y, Wang Z, Shi H, Li H, Li L, Fang R,
Cai X, Liu B, Zhang X and Ye L: HBXIP and LSD1 scaffolded by lncRNA
hotair mediate transcriptional activation by c-Myc. Cancer Res.
76:293–304. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rippe K and Luke B: TERRA and the state of
the telomere. Nat Struct Mol Biol. 22:853–858. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mourtada-Maarabouni M, Pickard MR, Hedge
VL, Farzaneh F and Williams GT: GAS5, a non-protein-coding RNA,
controls apoptosis and is downregulated in breast cancer. Oncogene.
28:195–208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang G, Lunardi A, Zhang J, Chen Z, Ala U,
Webster KA, Tay Y, Gonzalez-Billalabeitia E, Egia A, Shaffer DR, et
al: Zbtb7a suppresses prostate cancer through repression of a
Sox9-dependent pathway for cellular senescence bypass and tumor
invasion. Nat Genet. 45:739–746. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nie W, Ge HJ, Yang XQ, Sun X, Huang H, Tao
X, Chen WS and Li B: LncRNA-UCA1 exerts oncogenic functions in
non-small cell lung cancer by targeting miR-193a-3p. Cancer Lett.
371:99–106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou X, Gao Q, Wang J, Zhang X, Liu K and
Duan Z: Linc-RNA-RoR acts as a ‘sponge’ against mediation of the
differentiation of endometrial cancer stem cells by microRNA-145.
Gynecol Oncol. 133:333–339. 2014. View Article : Google Scholar : PubMed/NCBI
|