Introduction
Epithelial ovarian cancer (EOC) accounted for ~90%
of ovarian cancer cases worldwide in 2012 (1). EOC remains the most lethal
gynecological malignancy due to the heterogeneous nature of the
disease, and the majority of cases are diagnosed at a late stage
(2). The principal treatments for
ovarian cancer are complete surgical staging and maximal resection
with cisplatin-based chemotherapy (3). However, chemoresistance against
platinum-based therapies can decrease chemotherapeutic efficacy and
culminate in tumor recurrence (4).
Therefore, it is imperative to explore the mechanisms of
chemoresistance and to develop effective therapeutics for the
treatment of ovarian cancer.
Cisplatin was introduced into clinical trials in
1971, and is widely used in the treatment of solid tumors, such as
ovarian cancer (5). This compound is
the core drug used in combination with other chemotherapeutic
agents in the treatment of ovarian cancer in the clinical setting
(6). However, primary or acquired
drug resistance is a major challenge and decreases the treatment
efficiency (7). The potential
mechanisms underlying platinum-based chemotherapy involve
pharmacological, biochemical, apoptosis and microenvironment
resistance (8). Emerging evidences
have also indicated that autophagy may be involved in cell survival
and chemoresistance mechanisms (9–12).
Macroautophagy (autophagy) is a conserved catabolic pathway in
which cytosolic contents, such as damaged organelles, misfolded
proteins and bacteria, are transported into lysosomes for
degradation (13,14). Cytotoxic drugs often trigger
autophagy, particularly in apoptosis-defective cells (15); however, how the tumor cells utilize
autophagy to prevent chemotherapy-mediated cell death remains
elusive.
The Bcl-2-associated athanogene 3 (BAG3) protein is
a member of the BAG family (16).
The human BAG3 protein contains a C-terminal heat shock protein
70-binding BAG domain, which inhibits its chaperone activity
(17), a WW domain, a proline-rich
(PXXP) domain, and two conserved IPV motifs, one located centrally
and the other closer to the N terminus (18). BAG3 not only interacts with Bcl-2 to
inhibit apoptosis (16,19), but also participates in the
regulation of autophagy by interacting with heat shock protein
family proteins and heat shock protein β-8, to facilitate the
removal of misfolded and degraded proteins (20). The human BAG3 gene is located on the
long arm of chromosome 10 26.11 and is expressed constitutively in
myocytes, certain other normal cell types, such as breast and
thyroid cell lines, and several primary tumors or tumor cell lines,
including ovarian cancer cells (21). Numerous types of cell factors can
induce the expression of BAG3. For example, stress, heavy metals
and HIV infection can upregulate BAG3 expression (22). BAG3 dysfunction has been implicated
in numerous disorders, including cancer, myopathies and
neurodegeneration. Previous evidence has suggested that the BAG3
protein exerts a function in regulating the balance between cell
survival and death (16). A previous
study reported that BAG3-mediated selective macroautophagy is an
adaptive mechanism that maintains cellular homeostasis under stress
conditions and during aging, making BAG3 a potential target for
future pharmacological interventions (16). However, whether BAG3 participates in
cisplatin resistance in ovarian cancer cells, and the underlying
mechanisms remain unclear.
The present study utilized the cisplatin-resistant
ovarian cancer cell line SKOV3/DDP and the parental SKOV3 cell line
to identify: i) The roles of BAG3 in cisplatin-induced autophagy
associated with the drug resistance and ii) the relevant mechanisms
for regulating the process. The present findings support BAG3 as a
potential therapeutic target for enhancing the efficacy of
cisplatin in ovarian cancer.
Materials and methods
Cell lines and reagents
The human EOC cell lines SKOV3 (parental) and
SKOV3/DDP (cisplatin-resistant) were obtained from the Cell Bank of
the Chinese Academy of Sciences (Beijing, China). The cells were
cultured in RPMI-1640 medium supplemented with 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and 100 U/ml streptomycin/penicillin at 37°C in a humidified
atmosphere containing 95% air and 5% CO2. The SKOV3/DDP
cells were additionally cultured with 1 µM cisplatin to maintain
drug-resistance. Antibodies against microtubule-associated protein
1 light chain 3 (LC3; cat. no. 2775), sequestosome 1 (p62/SQSTM1;
cat. no. 5114), caspase-3 (cat. no. 9662), cleaved caspase-3 (cat.
no. 9664), poly [ADP-ribose] polymerase (PARP; cat. no. 6542) and
cleaved PARP (cat. no. 5625) were obtained from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Antibodies against GAPDH (cat.
no. 10494-1-AP) and BAG3 (cat. no. 10599-1-AP) were purchased from
Protein Technology (Chicago, IL, USA). Cisplatin (cat. no. BP809,
15663-27-1), chloroquine disphosphate (CQ; cat. no. C6628, 50-63-5)
and the secondary antibody (anti-mouse immunoglobulin G; cat. no.
A2304) were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt,
Germany). The cell culture medium was purchased from Gibco (Thermo
Fisher Scientific, Inc., Waltham, MA, USA).
Cell viability assay
SKOV3 (1×104) and SKOV3/DDP
(6×103) cells were seeded into 96-well plates and
cultured in the aforementioned conditions for 24 h. Cells were
permitted to adhere, and were then pretreated at 37°C with or
without 10 µM CQ for 2 h, followed by the indicated concentrations
of cisplatin (20 µM), CQ (10 µM) or a combination (20 µM cisplatin
+ 10 µM CQ) for the indicated times. DMSO was used as the untreated
control. In brief, 10 µl Cell Counting Kit-8 (CCK8) reagents
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China) were added to each well and cultured at 37°C for ~1 h, and
the absorbance at 450 nm was then calculated using a microplate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Cell
survival was calculated as: Cell survival rate (%)=(experimental
group OD value-blank group OD value)/(control group OD value-blank
group OD value). The concentration of cisplatin necessary to cause
50% inhibition (IC50) was calculated by non-linear
regression using GraphPad Prism version 5.0 (GraphPad Software,
Inc., La Jolla, CA, USA). All samples were performed in triplicate
for each group.
Green fluorescent protein (GFP)-LC3
plasmid transfection and autophagic flux assay
The SKOV3 and SKOV3/DDP cells were transfected with
a GFP-LC3 expression plasmid (500 ng/µl, BioVector, Ltd, Preston,
UK) at 70–80% confluence using Lipofectamine 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.). The time interval between
transfection and subsequent experimentation was 48 h. After 48 h,
the cells were fixed with 3.7% formaldehyde for 15 min at room
temperature and washed three times with cold PBS, and the
distribution and fluorescence of GFP-LC3 were visualized by
fluorescence microscopy (magnification, ×200). The numbers of
GFP-LC3 puncta were counted manually in 20 cells per group, in 10
randomly selected fields of view as described previously (23).
Transmission electron microscopy (TEM)
assay
Following treatment, cells were washed with 0.1×
cacodylate buffer (pH 7.4) and were fixed with 2% glutaraldehyde in
PBS for 24 h at 4°C. The rest of the process was conducted as
described previously (24). Sections
were examined using a Zeiss transmission electron microscope
(magnification, ×10,000; Zeiss GmbH, Jena, Germany).
Flow cytometry assay
The degree of apoptosis was assessed using an
Annexin V-FITC/propidium iodide (PI) apoptosis detection kit (BD
Pharmingen; BD Biosciences, San Jose, CA, USA). Following the
aforementioned treatments, cells were harvested in 0.25% trypsin
and washed three times with PBS, and 1×105 cells were
then suspended in 500 µl binding buffer and incubated with 5 µl
Annexin V and 5 µl PI for 15 min in the dark. Analysis of apoptotic
cells was performed using a flow cytometer (Cytomics™ FC 500;
Beckman Coulter, Inc., Brea, CA, USA; CXP. Software version 1.0 CXP
Cytometer; Beckman Coulter, Inc.).
TUNEL assay
A total of 3×105 SKOV3 and SKOV3/DDP
cells per well were seeded on six-well plates and fixed with 3.7%
paraformaldehyde at room temperature for 1 h. The cells were
permitted to adhere. The cells were washed three times with PBS for
5 min between each step, which was followed by the addition of 100
µl 0.1% Triton X-100. The cells were then placed on ice for 2 min
and 50 µl TUNEL (Beyotime Institute of Biotechnology, Shanghai,
China) reaction mixture (2 µl TdT enzyme and 45 µl fluorescence
labeling) was added to the cells, followed by incubation at 37°C
for 1 h. Cell nuclei were stained with 1 ml 10 µg/ml DAPI (Beyotime
Institute of Biotechnology) for 5 min at room temperature in the
dark and observed under a fluorescence microscope for TUNEL and
DAPI staining (EVOS™ Thermo Fisher Scientific, Inc.). Cells with
red fluorescence were defined as apoptotic cells, and the number of
apoptotic cells was counted in 5 randomly selected fields of view
at ×200 magnification.
cDNA constructs, short hairpin (sh)RNA
and transfection
Cells were transfected with a GFP-tagged LC3 cDNA
expression construct or BAG3 shRNA (GenePharma, Shanghai, China)
using Lipofectamine 3000, according to the manufacturer's protocol.
The sequence of BAG3 shRNA was 5′-GATACACGAGCAGAACGTTAC-3′, the
concentration of BAG3 shRNA used for transfection was 500 ng/µl.
The time interval between transfection and subsequent
experimentation was 48 h.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from SKOV3 and SKOV3/DDP
cells using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) and RT was performed using the PrimeScript™ RT
Master Mix (Takara Bio, Inc., Shiga, Japan). The temperature
protocol used for RT was, 37°C for 15 min, 85°C for 5 sec followed
by a 4°C holding step. The qPCR reactions were performed using SYBR
Green reagents (Takara Bio Inc.) on an Agilent Mx3005P system
(Agilent Technologies, Inc., Santa Clara, CA, USA). The
thermocycling conditions were: 95°C for 30 sec; 35 cycles of 95°C
for 30 sec; 60°C for 30 sec; 72°C for 15 sec, fluorescent signals
are collected here; 95°C for 5 sec; and 65°C for 59 sec; and 95°C
for 30 sec. Each sample was run in triplicate. The primers used
were as follows: Human BAG3 forward, 5′-CTCCATTCCGGTGATACACGA-3′
and reverse, 5′-TGGTGGGTCTGGTACTCCC-3′; GAPDH forward,
5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse, 5′-GAAGATGGTGATGGGATTTC-3′.
The relative expression levels were calculated using the
2−ΔΔCq method (25).
Immunofluorescence assay
SKOV3 and SKOV3/DDP cells were cultured on
coverslips, fixed with 3.7% paraformaldehyde in PBS for 30 min at
room temperature, and permeabilized with 0.1% Triton X-100 in PBS
for 20 min. Following blocking with 10% FBS for 30 min at room
temperature, cells were incubated with primary antibodies for 2 h,
washed three times with PBS, and incubated with secondary
antibodies for 1 h at room temperature in the dark. The nuclei were
then visualized using DAPI for 5 min at room temperature in the
dark (Beyotime Institute of Biotechnology) and the cells were
observed using a Leica TCS SP5 laser confocal microscope
(magnification, ×800; Leica Microsystems GmbH, Wetzlar,
Germany).
Western blot analysis
Following the aforementioned treatments, SKOV3 and
SKOV3/DDP cells were washed with ice-cold PBS and lysed in RIPA
buffer (Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China), and protein concentrations were subsequently
determined by a bicinchoninic acid protein assay (Beyotime
Biotechnology Institute of Biotechnology). The protein lysates (40
µg) were separated by 12% SDS-PAGE or 15% SDS-PAGE and transferred
to PVDF membranes (EMD Millipore Corp., Billerica, MA, USA). The
membranes were blocked with 5% skimmed milk for 2 h at room
temperature with slow agitation, incubated with primary antibodies
(1:1,000) overnight at 4°C, and were subsequently incubated with
horseradish peroxidase-conjugated secondary antibodies (1:2,000)
for 1 h at room temperature. The membranes were then visualized
with ECL reagents (Applygen, Beijing, China). Densitometric
analysis was performed using the ImageJ software version 6.0
(National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All the data are presented as the mean ± standard
deviation. Data were analyzed and visualized using GraphPad Prism
version 5.0. Two-sided Student's t-test was used to assess for
statistically significant differences between two groups. A one-way
analysis of variance followed by Tukey's post-hoc test was used to
determine statistical differences among multiple treatment groups.
All experiments were independently repeated in triplicate.
P<0.05 was considered to indicate a statistically significant
difference; all tests were two-sided and no corrections were
applied for multiple significance testing.
Results
Sensitivity of EOC cell lines to
cisplatin treatment
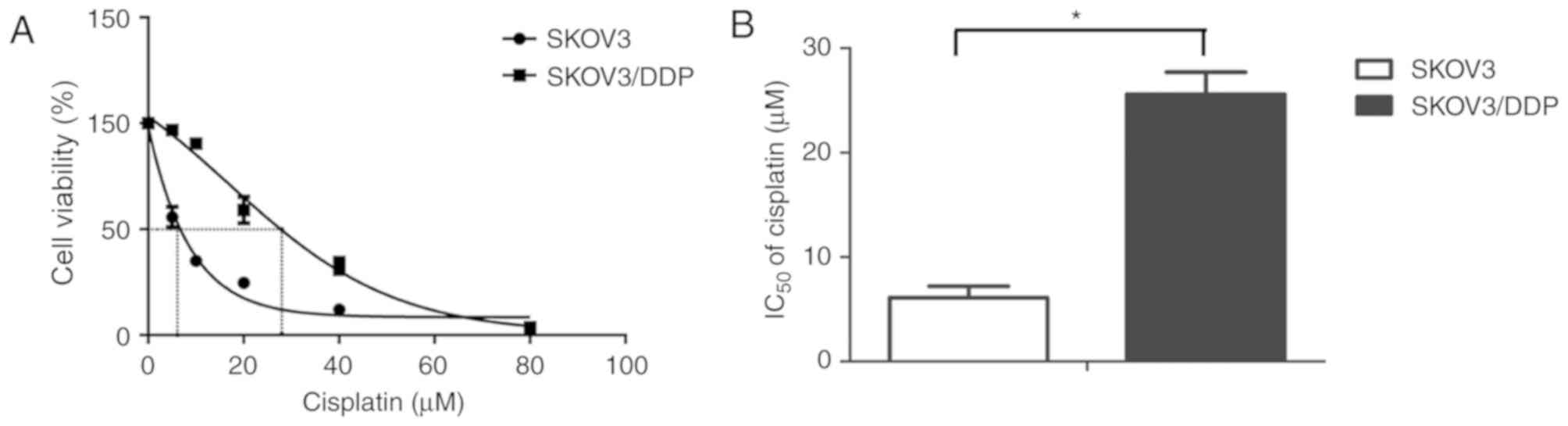
The present study detected cisplatin sensitivity of
the SKOV3/DDP cells compared with their parental SKOV3 cells using
the CCK8 assay after the cells were exposed to various
concentrations of cisplatin for 48 h. The percentage of surviving
cells decreased in a dose-dependent manner in the SKOV3 and
SKOV3/DDP cells (Fig. 1A). The 48 h
IC50 values of cisplatin in SKOV3 and SKOV3/DDP cells
were 6.12±1.09 and 25.59±2.116 µM, respectively (Fig. 1B). Based on these results, 20 µM
cisplatin was selected for use in the subsequent experiments.
Cisplatin induces cytoprotective
autophagy in EOC cells
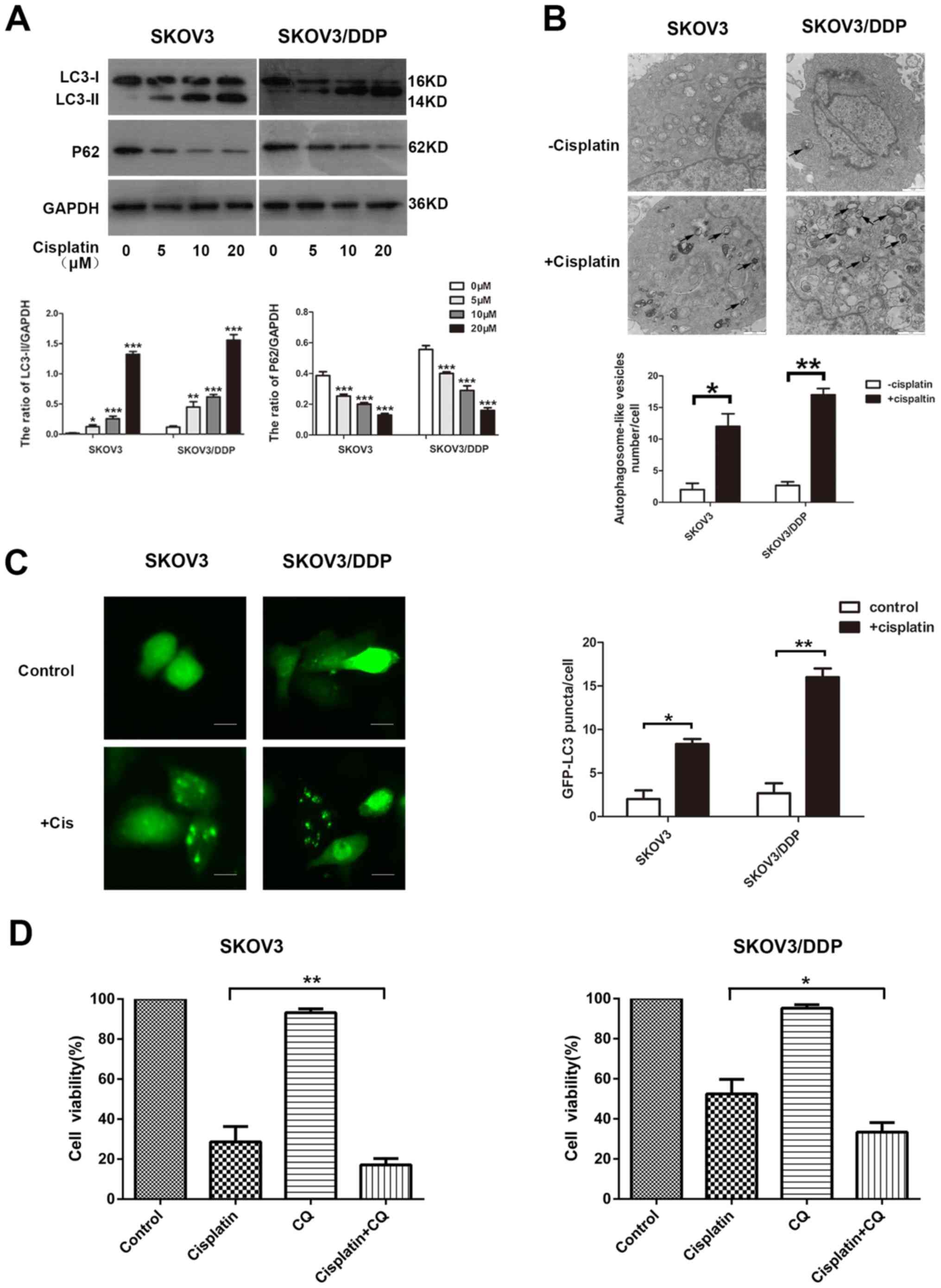
The ability of cisplatin to induce autophagy in EOC
cells was investigated. The detection of LC3, a mammalian ortholog
of yeast Atg8, is a common way to monitor autophagy. During
autophagy induction, the non-lipidated form of LC3 (LC3-I) is
conjugated with phosphatidylethanolamine, converted into the
lipidated form (LC3-II), which is associated with autophagosome
biogenesis. The p62 protein links with LC3-II or ubiquitinated
substrates, which are degraded during autophagic flux (26). In the present study, SKOV3 and
SOV3/DPP cells were exposed to concentrations of cisplatin ranging
from 0 to 20 µM, and the drug was observed to activate autophagy.
The treatment increased the level of LC3-II and decreased p62
expression in the SKOV3 and SKOV3/DDP cells, as demonstrated by
western blotting (Fig. 2A). Similar
results were observed with ultrastructural analysis of
autophagosomes using TEM (Fig. 2B).
The effect of cisplatin on autophagy was further confirmed by a
GFP-LC3 punctate-formation assay. SKOV3 and SKOV3/DDP cells were
transiently transfected with a GFP-LC3 plasmid to monitor
autophagosome formation through fluorescence microscopy, and the
number of GFP-LC3 puncta cells were increased in the examined cell
lines (Fig. 2C).
In order to determine whether cisplatin-induced
autophagy exerted a protective function in cisplatin treatment, an
autophagy inhibitor, CQ, was used. Treatment with CQ increases the
formation of autophagosomes by preventing autophagosome-lysosome
fusion, eventually inhibiting late-stage autophagy (27). Cells were treated with cisplatin in
the presence or absence of CQ, and cell viability was determined
using a CCK8 assay. Cells treated with cisplatin alone were more
resistant than those treated with cisplatin and CQ in the examined
cell lines (Fig. 2D). These results
suggest that cisplatin induces autophagy, serving a protective role
in ovarian cancer cells exposed to cisplatin.
Cisplatin promotes BAG3 expression in
EOC cells
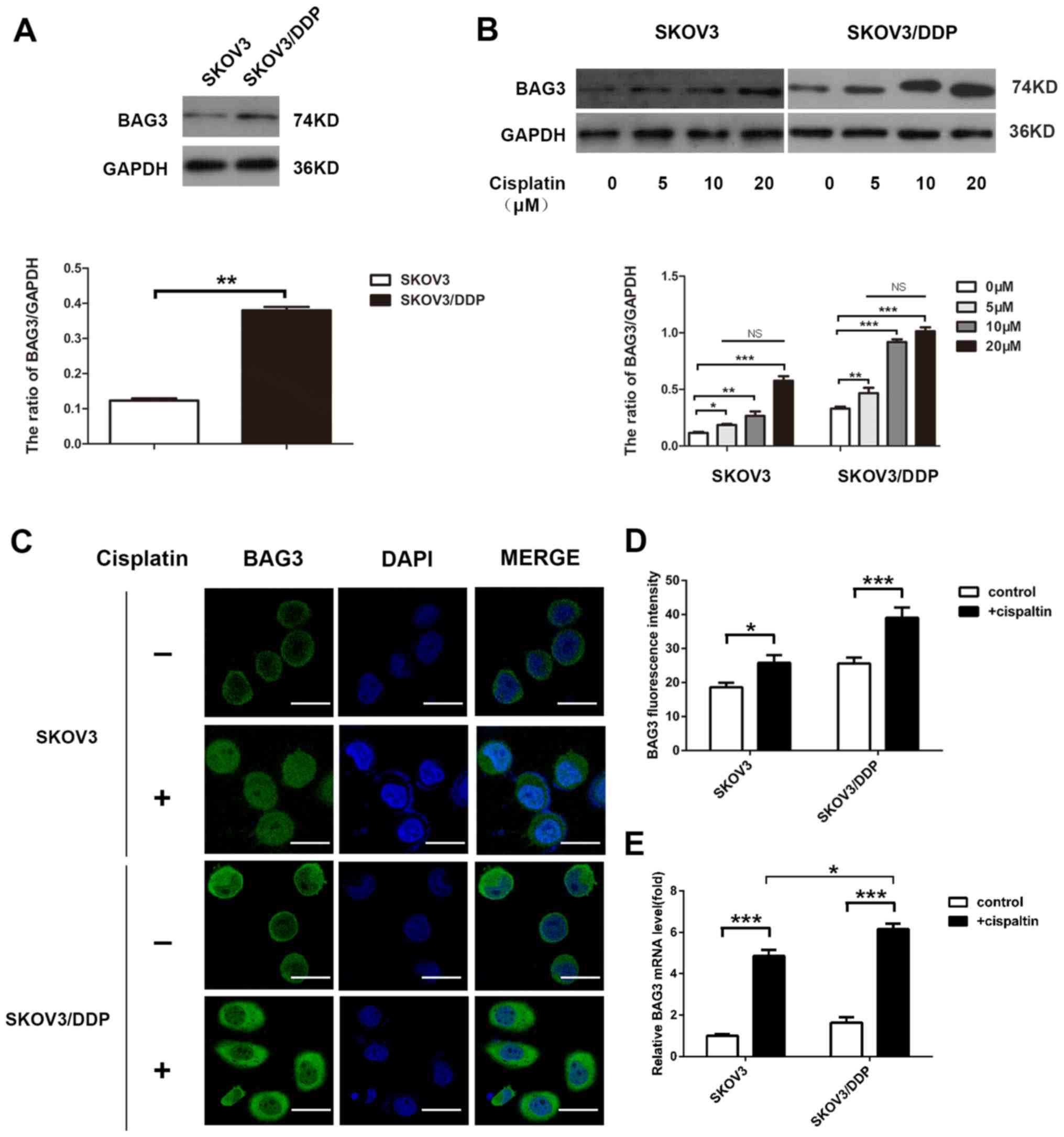
The present study investigated the potential role of
BAG3 in the regulation of cisplatin-induced autophagy. The
endogenous expression level of BAG3 was lower in the SKOV3 compared
with the SKOV3/DDP cells, according to western blot analysis
(Fig. 3A). The effect of 48-h
treatment with various concentrations of cisplatin on BAG3
expression was then determined. The treatment increased BAG3
protein levels in the cells and the effect was not
concentration-dependent. The relative BAG3 expression exhibited no
significant differences between different treatment concentration
groups (Fig. 3B). Furthermore,
confocal microscopy analysis revealed that the green fluorescence
of BAG3 accumulated primarily in the cytoplasm, with small amounts
in the nucleus (Fig. 3C), and the
fluorescence intensity was significantly increased following
treatment with 20-µM cisplatin (Fig.
3D). Consistent with the western blotting results, RT-qPCR
demonstrated that BAG3 mRNA levels increased following treatment
with cisplatin for 48 h, the increase in BAG3 mRNA expression
levels in SKOV3/DDP cells was larger than the increase in SKOV3
cells (Fig. 3E).
Downregulation of BAG3 modulates
cisplatin resistance in ovarian cancer cells via inhibition of
autophagy
The present study investigated the association
between BAG3 and cisplatin-induced autophagy in EOC cells via the
knockdown of BAG3 with shRNA. SKOV3/DDP and SKOV3 cells transfected
with BAG3 shRNA were treated with cisplatin (20 µM) for 48 h. BAG3
knockdown caused marked decreases in the protein level of BAG3 in
the examined cell lines, as demonstrated by western blot analysis.
Protein level detection also revealed that the downregulation of
BAG3 prevented the formation of LC3-II and the degradation of p62,
and enhanced the expression of apoptosis-associated proteins
cleaved-PARP and cleaved-caspase-3, following exposure to
cisplatin. Furthermore, BAG3 downregulation inhibited autophagy
activity more markedly in the SKOV3/DDP cells compared with SKOV3
(Fig. 4A). Subsequently, cell
viability was examined using a CCK8 assay. BAG3 knockdown
significantly enhanced the sensitivity of the cells to cisplatin
compared with that in cells transfected with control shRNA
(Fig. 4B). The formation of GFP-LC3
punctate fluorescent dots following treatment with cisplatin were
also decreased in SKOV3/DDP cells transfected with BAG3 shRNA,
compared with the vector control group (Fig. 4C). Taken together, these results
demonstrated that BAG3 serves an important role in the regulation
of cisplatin-induced autophagy in ovarian cancer cells.
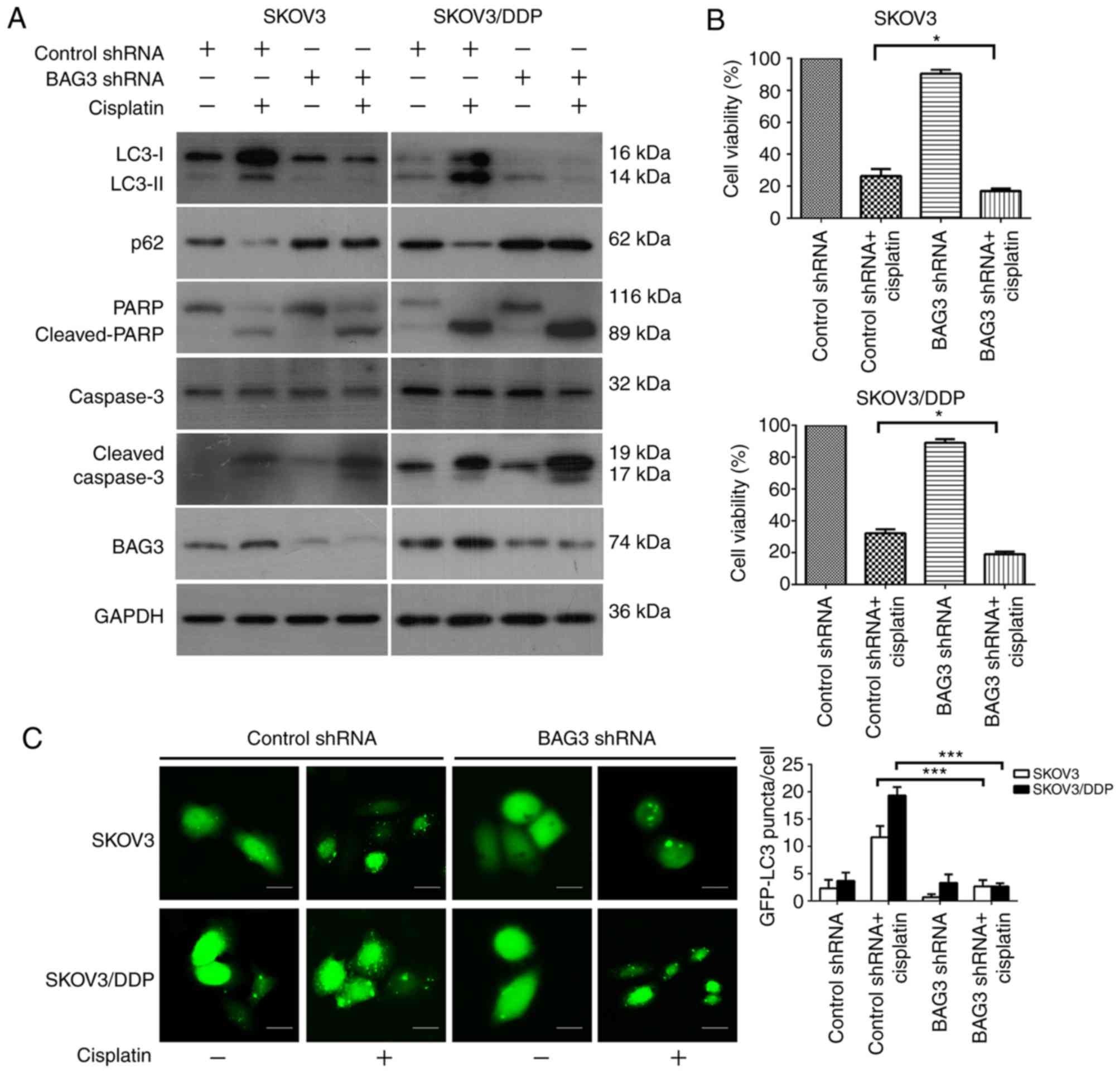 | Figure 4.Downregulation of BAG3 modulates
cisplatin resistance in ovarian cancer cells via the inhibition of
autophagy. SKOV3 and SKOV3/DDP cells were transfected with control
RNA or shRNA targeting BAG3, followed by treatment with 20 µM
cisplatin for 48 h. (A) Whole-cell lysates were subjected to
western blot analysis to detect LC3, p62, PARP, cleaved-PARP,
caspase-3, cleaved-caspase-3, BAG3 and GAPDH (as a loading
control). (B) Cell viability was analyzed using a Cell Counting
Kit-8. The data are presented as the mean ± standard deviation from
three independent experiments. *P<0.05. (C) SKOV3 and SKOV3/DDP
cells were co-transfected with either control or BAG3 shRNA and
GFP-LC3 plasmid followed by treatment with cisplatin (20 µM) for an
additional 48 h. Scale bar, 200 µm. The formation of GFP-LC3 puncta
were examined using immunofluorescence and quantified. The data are
presented as the mean ± standard deviation from three independent
experiments. ***P<0.001. BAG3, Bcl-2-associated athanogene 3;
LC3, microtubule-associated protein 1 light chain 3; p62,
sequestosome 1; PARP, poly [ADP-ribose] polymerase; shRNA, short
hairpin RNA; GFP, green fluorescent protein. |
BAG3 enhances cisplatin-induced
apoptosis in EOC cells
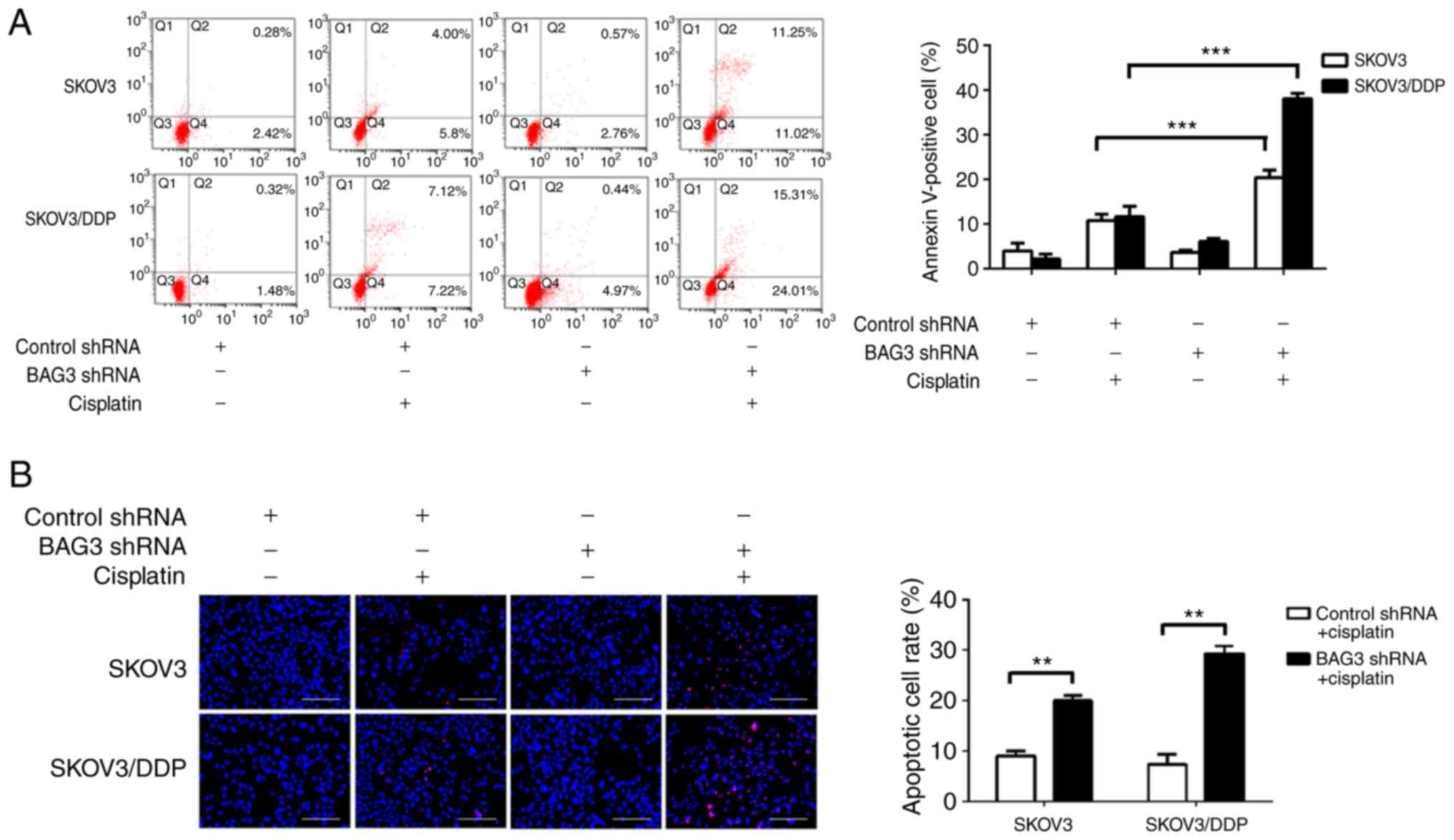
To clarify the role of BAG3 in the regulation of EOC
cell apoptosis following cisplatin treatment, shRNA targeting BAG3
was transfected into SKOV3 and SKOV3/DDP cells. Cisplatin induced
apoptotic cell death to a great extent after BAG3 knockdown, as
revealed by an increase of Annexin V-positive cells by 20.41±1.36
and 38.06±0.99% in SKOV3 and SKOV3/DDP cells, respectively
(Fig. 5A). Consistent with the flow
cytometry results, the TUNEL assay revealed that cisplatin induced
apoptotic cell death to a great degree following BAG3 knockdown in
the SKOV3/DDP cells (**P<0.01; Fig.
5B). These data indicate that downregulation of BAG3 enhances
cisplatin-induced apoptosis, confirming its role in the resistance
of ovarian cancer cell to cisplatin.
Discussion
The platinum-based cytotoxic compound cisplatin has
been commonly used in ovarian cancer treatment for almost three
decades, as it induces apoptosis in cancer cells due to lethal DNA
damage (28). However, the intrinsic
and acquired resistance to cisplatin in cancer cells remains a
major challenge. Multiple molecular mechanisms contribute to
cisplatin-resistance, including a defective DNA repair system,
enhanced drug clearance and detoxification, epigenetic regulation
and abnormal signaling pathways (29). Growing evidence indicates a role of
autophagy as a pro-survival and resistance mechanism to
chemotherapy. A recent study demonstrated that cisplatin can induce
autophagy in ovarian cancer cells (5). Herein, a new mechanism is described in
which the downregulation of BAG3 mediates cisplatin-induced
autophagy in cisplatin-resistant ovarian cancer cells.
Autophagy is an evolutionarily conserved process
whereby cytoplasmic cargo sequestered inside double-membrane
vesicles is delivered to the lysosome for degradation (30). Autophagy exerts cytoprotective
functions and may also lead programmed cell death (31). An increasing amount of evidence
suggests that autophagy process may promote cell survival following
treatment with anticancer drugs, and may therefore be associated
with chemotherapy resistance (32).
The induction of autophagy has been demonstrated to promote
resistance of ovarian cancer to cisplatin (33). Zhang et al (34) demonstrated that thioredoxin domain
containing 17 promoted paclitaxel resistance by inducing autophagy
in ovarian cancer. The present study investigated cisplatin
resistance in ovarian cancer SKOV3 and SKOV3/DDP cell lines, and
the IC50 of cisplatin in the SKOV3/DDP was ~4-fold
higher than the parental SKOV3 cells. Cisplatin induced autophagy
in a concentration-dependent manner in the examined cells, as
demonstrated by the western blot results for the autophagy markers
LC3-I, LC3-II and p62. This compound has previously been
demonstrated to induce autophagy in various types of cancer cells,
such as ovarian cancer cells and osteosarcoma cancer cells
(35,36), and another study indicated that
autophagy serves a protective role in cisplatin resistance
(34). The results of the present
study demonstrated that the blockade of autophagy promoted
cisplatin-induced cell death in SKOV3 and SKOV3/DDP cells, and
partially re-sensitized cisplatin-resistant SKOV3/DDP cells.
BAG3 has been reported to function as a novel
modulator of autophagy in cancer cells by regulating key
autophagy-related proteins (37).
The functions of BAG3 in ovarian cancer have also been partly
investigated. A previous study reported that increased BAG3
expression was significantly associated with poor overall survival
in patients with primary ovarian tumors (38). BAG3 was also revealed to increase the
invasiveness of uterine corpus and ovarian carcinomas (39,40).
Furthermore, it also induced resistance to paclitaxel in ovarian
clear cell carcinoma cells (41).
Recent evidence has suggested that BAG3 exerts a function in
adjusting apoptosis and modulating cisplatin resistance (42). The present findings support a role
for autophagy activation in chemoresistance in cancer cells, and
the downregulation of autophagy was revealed to sensitize the
examined cancer cells to cisplatin. Few previous studies have
investigated the association between BAG3 and autophagy in ovarian
cancer cisplatin resistance, although the current study
demonstrated that cisplatin treatment upregulates BAG3 expression.
Regulating autophagy to induce cell death, inhibiting protective
autophagy, and promoting crosstalk with tissue-specific apoptosis
may be promising avenues for novel anticancer chemotherapeutic
strategies (43). Therefore, the
present study evaluated the role of BAG3 in regulating autophagy in
ovarian cancer. The knockdown of BAG3 by shRNA led to the
suppression of autophagy, which was measured by a decrease in the
level of LC3-II, GFP-LC3 puncta formation and p62 degradation, when
compared with the control group, particularly in SKOV3/DDP cells.
The downregulation of BAG3 also markedly increased the sensitivity
to cisplatin in SKOV3/DDP cells compared with that in SKOV3, as
determined by a CCK8 assay. The present study further investigated
the association between autophagy and apoptosis with BAG3
downregulation. The knockdown of BAG3 significantly augmented
cisplatin-induced apoptosis, as indicated by an increase in the
expression of cleaved caspase-3 and PARP. In addition, cisplatin
induced apoptotic cell death to a greater degree following BAG3
knockdown, as revealed by Annexin V/PI and TUNEL staining. These
results suggest that the downregulation of BAG3 attenuates
cisplatin resistance by inhibiting autophagy in ovarian cancer
cells. However, the detailed molecular mechanisms underlying the
regulation of autophagy via BAG3 may be complex, and further
studies are required in order to clarify these.
In conclusion, the present study identified BAG3 as
a novel regulator of autophagy and demonstrated its involvement in
the modulation of cisplatin resistance in ovarian cancer cells.
BAG3 also affected apoptosis, therefore the downregulation of BAG3
can enhance the sensitivity of ovarian cancer cells to cisplatin by
regulating autophagy and apoptosis, particularly in the
cisplatin-resistant SKOV3/DDP cells. Consequently, BAG3 may
represent a novel therapeutic target for preventing chemoresistance
in cancer cells, and the knockdown of BAG3 may be a useful strategy
to overcome chemoresistance by preventing cytoprotective autophagy
in ovarian cancer cells.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SQ conceived and designed the study. SQ and LS
performed the whole experiment, analyzed the data and wrote the
article. YZ and SH analyzed the data and proofread the article.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
EOC
|
epithelial ovarian cancer
|
|
BAG3
|
Bcl-2-associated athanogene 3
|
|
LC3
|
microtubule-associated protein 1 light
chain 3
|
|
p62/SQSTM1
|
sequestosome 1
|
|
PARP
|
poly [ADP-ribose] polymerase
|
|
FBS
|
fetal bovine serum
|
|
CQ
|
chloroquine disphosphate
|
|
PBS
|
phosphate-buffered saline
|
|
DMSO
|
dimethylsulfoxide
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN, 2012. Int J cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jeon SY, Hwang KA and Choi KC: Effect of
steroid hormones, estrogen and progesterone, on epithelial
mesenchymal transition in ovarian cancer development. J Steroid
Biochem Mol Biol. 158:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim A, Ueda Y, Naka T and Enomoto T:
Therapeutic strategies in epithelial ovarian cancer. J Exp Clin
Cancer Res. 31:142012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang P, Zhang P, Shi B, Zhou M, Jiang H,
Zhang H, Pan X, Gao H, Sun H and Li Z: Galectin-1 overexpression
promotes progression and chemoresistance to cisplatin in epithelial
ovarian cancer. Cell Death Dis. 5:e9912014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhu J, Zheng Y, Zhang H, Zhu J and Sun H:
Low concentration of chloroquine enhanced efficacy of cisplatin in
the treatment of human ovarian cancer dependent on autophagy. Am J
Transl Res. 9:4046–4058. 2017.PubMed/NCBI
|
|
6
|
Maurmann L, Belkacemi L, Adams NR,
Majmudar PM, Moghaddas S and Bose RN: A novel cisplatin mediated
apoptosis pathway is associated with acid sphingomyelinase and FAS
proapoptotic protein activation in ovarian cancer. Apoptosis.
20:960–974. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cornelison R, Llaneza DC and Landen CN:
Emerging therapeutics to overcome chemoresistance in epithelial
ovarian cancer: A mini-review. Int J Mol Sci. 18:E21712017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ren F, Shen J, Shi H, Hornicek FJ, Kan Q
and Duan Z: Novel mechanisms and approaches to overcome multidrug
resistance in the treatment of ovarian cancer. Biochim Biophys
Acta. 1866:266–275. 2016.PubMed/NCBI
|
|
9
|
Lou JS, Bi WC, Chan GKL, Jin Y, Wong CW,
Zhou ZY, Wang HY, Yao P, Dong TTX and Tsim KWK: Ginkgetin induces
autophagic cell death through p62/SQSTM1-mediated autolysosome
formation and redox setting in non-small cell lung cancer.
Oncotarget. 8:93131–93148. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Naik PP, Mukhopadhyay S, Panda PK, Sinha
N, Das CK, Mishra R, Patil S and Bhutia SK: Autophagy regulates
cisplatin-induced stemness and chemoresistance via the upregulation
of CD44, ABCB1 and ADAM17 in oral squamous cell carcinoma. Cell
Prolif. 51:2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pistollato F, Calderón Iglesias R, Ruiz R,
Aparicio S, Crespo J, Dzul Lopez L, Giampieri F and Battino M: The
use of natural compounds for the targeting and chemoprevention of
ovarian cancer. Cancer Lett. 14:191–200. 2017. View Article : Google Scholar
|
|
12
|
Zhou WJ, Chang KK, Wu K, Yang HL, Mei J,
Xie F, Li DJ and Li MQ: Rapamycin synergizes with cisplatin in
antiendometrial cancer activation by improving IL-27-stimulated
cytotoxicity of NK cells. Neoplasia. 20:69–79. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rubinsztein DC: The roles of intracellular
protein-degradation pathways in neurodegeneration. Nature.
443:780–786. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu X, Feng X, Zhao X, Ma F, Liu N, Guo H,
Li C, Du H and Zhang B: Role of beclin-1-mediated autophagy in the
survival of pediatric leukemia cells. Cell Physiol Biochem.
39:1827–1836. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu G, Pei F, Yang F, Li L, Amin AD, Liu
S, Buchan JR and Cho WC: Role of autophagy and apoptosis in
non-small-cell lung cancer. Int J Mol Sci. 18:E3672017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Behl C: Breaking BAG: The Co-Chaperone
BAG3 in health and disease. Trends Pharmacol Sci. 37:672–688. 2017.
View Article : Google Scholar
|
|
17
|
Takayama S, Xie Z and Reed JC: An
evolutionarily conserved family of Hsp70/Hsc70 molecular chaperone
regulators. J Biol Chem. 274:781–786. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rauch JN, Tse E, Freilich R, Mok SA,
Makley LN, Southworth DR and Gestwicki JE: Bag3 is a modular,
scaffolding protein that physically links heat shock protein 70
(Hsp70) to the small heat shock proteins. J Mol Biol. 429:128–141.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Knezevic T, Myers VD, Gordon J, Tilley DG,
Sharp TE III, Wang J, Khalili K, Cheung JY and Feldman AM: BAG3: A
new player in the heart failure paradigm. Heart Fail Rev.
20:423–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ganassi M, Mateju D, Bigi I, Mediani L,
Poser I, Lee HO, Seguin SJ, Morelli FF, Vinet J, Leo G, et al: A
surveillance function of the HSPB8-BAG3-HSP70 chaperone complex
ensures stress granule integrity and dynamism. Mol Cell.
63:796–810. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Felzen V, Hiebel C, Koziollek-Drechsler I,
Reißig S, Wolfrum U, Kögel D, Brandts C, Behl C and Morawe T:
Estrogen receptor alpha regulates non-canonical autophagy that
provides stress resistance to neuroblastoma and breast cancer cells
and involves BAG3 function. Cell Death Dis. 6:e18122015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim EM, Jung CH, Kim J, Hwang SG, Park JK
and Um HD: The p53/p21 complex regulates cancer cell invasion and
apoptosis by targeting Bcl-2 family proteins. Cancer Res.
77:3092–3100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen G, Ke Z, Xu M, Liao M, Wang X, Qi Y,
Zhang T, Frank JA, Bower KA, Shi X and Luo J: Autophagy is a
protective response to ethanol neurotoxicity. Autophagy.
8:1577–1589. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ylä-Anttila P, Vihinen H, Jokitalo E and
Eskelinen EL: Monitoring autophagy by electron microscopy in
mammalian cells. Methods Enzymol. 452:143–164. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pan B, Chen D, Huang J, Wang R, Feng B,
Song H and Chen L: HMGB1-mediated autophagy promotes docetaxel
resistance in human lung adenocarcinoma. Mol Cancer. 13:1652014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee SW, Kim HK, Lee NH, Yi HY, Kim HS,
Hong SH, Hong YK and Joe YA: The synergistic effect of combination
temozolomide and chloroquine treatment is dependent on autophagy
formation and p53 status in glioma cells. Cancer Lett. 360:195–204.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Damia G and Broggini M: Platinum
resistance in ovarian cancer: Role of DNA repair. Cancer (Basel).
11:E1192019. View Article : Google Scholar
|
|
29
|
Bookman MA: First-line chemotherapy in
epithelial ovarian cancer. Clin Obstet Gynecol. 55:96–113. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ahmad L, Mostowy S and Sancho-Shimizu V:
Autophagy-virus interplay: From cell biology to human disease.
Front Cell Dev Biol. 6:1552018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen N and Karantza V: Autophagy as a
therapeutic target in cancer. Cancer Biol Ther. 11:157–168. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He C, Dong X, Zhai B, Jiang X, Dong D, Li
B, Jiang H, Xu S and Sun X: MiR-21 mediates sorafenib resistance of
hepatocellular carcinoma cells by inhibiting autophagy via the
PTEN/Akt pathway. Oncotarget. 30:28867–28881. 2015.
|
|
33
|
Xiao L, Shi XY, Zhang Y, Zhu Y, Zhu L,
Tian W, Zhu BK and Wei ZL: YAP induces cisplatin resistance through
activation of autophagy in human ovarian carcinoma cells. Onco
Ther. 9:1105–1114. 2016.
|
|
34
|
Zhang SF, Wang XY, Fu ZQ, Peng QH, Zhang
JY, Ye F, Fu YF, Zhou CY, Lu WG, Cheng XD and Xie X: TXNDC17
promotes paclitaxel resistance via inducing autophagy in ovarian
cancer. Autophagy. 11:225–238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Y, Jiang W, Hu Y, Da Z, Zeng C, Tu M,
Deng Z and Xiao W: MicroRNA-199a-5p inhibits cisplatin-induced drug
resistance via inhibition of autophagy in osteosarcoma cells. Oncol
Lett. 12:4203–4208. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin WM and Li ZG: Blockage of
cisplatin-induced autophagy sensitizes cervical cancer cells to
cisplatin. Gene Mol Res. 14:16905–16912. 2015. View Article : Google Scholar
|
|
37
|
De Marco M, Basile A, Iorio V, Festa M,
Falco A, Ranieri B, Pascale M, Sala G, Remodelli P, Capunzo M, et
al: Role of BAG3 in cancer progression: A therapeutic opportunity.
Semin Cell Dev Biol. 78:85–92. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nymoen DA, Hetland Falkenthal TE, Holth A,
Ow GS, Ivshina AV, Tropé CG, Kuznetsov VA, Staff AC and Davidson B:
Expression and clinical role of chemoresponse-associated genes in
ovarian serous carcinoma. Gynecol Oncol. 139:30–39. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Suzuki M, Iwasaki M, Sugio A, Hishiya A,
Tanaka R, Endo T, Takayama S and Saito T: BAG3 (BCL2-associated
athanogene 3) interacts with MMP-2 to positively regulate invasion
by ovarian carcinoma cells. Cancer Lett. 303:65–71. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Habata S, Iwasaki M, Sugio A, Suzuki M,
Tamate M, Satohisa S, Tanaka R and Saito T: BAG3 increases the
invasiveness of uterine corpus carcinoma cells by suppressing
miR-29b and enhancing MMP2 expression. Oncol Rep. 33:2613–2621.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Habata S, Iwasaki M, Sugio A, Suzuki M,
Tamate M, Satohisa S, Tanaka R and Saito T: BAG3-mediated Mcl-1
stabilization contributes to drug resistance via interaction with
USP9X in ovarian cancer. Int J Oncol. 49:402–410. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wu K, Yang Y, Zhao J and Zhao S:
BAG3-mediated miRNA let-7g and let-7i inhibit proliferation and
enhance apoptosis of human esophageal carcinoma cells by targeting
the drug transporter ABCC10. Cancer Lett. 371:125–133. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Belaid A, Ndiaye PD, Filippakis H, Roux J,
Röttinger É, Graba Y, Brest P, Hofman P and Mograbi B: Autophagy:
Moving benchside promises to patient bedsides. Curr Cancer Drug
Targets. 15:684–702. 2015. View Article : Google Scholar : PubMed/NCBI
|