Introduction
Gastric cancer (GC) is one of the major malignancies
worldwide (1). The overall 5-year
survival rate of patients with GC remains poor (2). The poor prognosis of patients with this
disease results from late diagnosis and a poor response to
available therapies. Cisplatin (DDP)-based chemotherapy is the main
treatment strategy for GC (3).
Although combined drug therapy pre- and post-surgery has increased
overall survival, drug resistance continues to be a major clinical
obstacle to therapeutic efficacy (4). Drug resistance is categorized into two
types: i) Intrinsic resistance, in which tumors are resistant to
the drug prior to treatment, thereby preventing the drug from
effectively treating the tumor even with early diagnosis; ii)
acquired resistance, in which resistance to the drug is formed
after prolonged cycles of chemotherapy despite an initial positive
reaction. Cisplatin typically kills tumor cells by inducing
apoptosis; however, it has been revealed that many solid tumors are
resistant to drug-induced apoptosis (5). Therefore, it is essential to identify
novel molecular markers for improving the sensitivity of GC cells
to cisplatin.
MicroRNAs (miRNAs) are small non-coding RNAs (19–24
nucleotides in length) that regulate gene expression at the
post-transcriptional level through base pairing with the
3′-untranslated region (3′-UTR) of target mRNAs, resulting in
translational repression or degradation of the mRNAs (6,7). An
increasing number of miRNAs have been demonstrated to be implicated
in the mechanism of drug resistance and the genes or pathways
involved in drug resistance (8).
Upregulation of miR-101 has been revealed to enhance cellular
sensitivity to cisplatin treatment by promoting the
apoptosis-inducing effects of cisplatin in GC (9). Previous studies demonstrated that
miR-362-5p serves a critical role in malignant tumors. By
collecting seven publicly available and independent renal cell
carcinoma miRNA expression profiling datasets, Ying et al
(10) revealed that hsa-miR-362-5p
is downregulated in renal carcinoma. Ni et al (11) have demonstrated that miR-362-5p
targets the cylindromatosis gene, thereby promoting hepatocellular
carcinoma cell growth and metastasis. It has also been demonstrated
that upregulation of miR-362-5p significantly accelerates
proliferation, migration and invasion of human breast cancer MCF7
cells (12). However, the biological
role of miR-362-5p in SGC7901/DDP cells remains to be explored.
Suppressor of zeste 12 protein (SUZ12) is a core
component of polycomb repressive complex 2 (PRC2), which
epigenetically silences gene transcription (13). In addition to SUZ12, PRC2 contains
the embryonic ectoderm development protein and the catalytic
subunit enhancer of zeste 2 polycomb repressive complex 2 (14), which is involved in the pathogenesis
of GC (15). Amplification and
overexpression of SUZ12 have been observed in multiple tumor types,
such as GC, ovarian cancer and non-small cell lung cancer (16–18).
Moreover, there is evidence that SUZ12 serves an important role in
GC by acting as an oncogene (16).
However, the role of SUZ12 in the cisplatin resistance of GC cells
has yet to be investigated.
The present study investigated the potential
function of miR-362-5p in vitro and aimed to further
understand its underlying mechanism in cisplatin-resistant GC
cells. In addition, this study defined the molecular mechanism of
SUZ12. These findings may provide novel insights into the tumor
biology of GC.
Materials and methods
Cell lines and culture
The human GC cell line SGC7901 and the corresponding
cisplatin-resistant cell line SGC7901/DDP were obtained from
Nanjing KeyGen Biotechnology Company. Cells were cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.),
containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 100
U/ml penicillin and 100 mg/ml streptomycin. To maintain the
cisplatin-resistant phenotype of SGC7901/DDP cells, cisplatin (800
ng/ml; Jiangsu Hansoh Pharmaceutical Group Co., Ltd.) was added to
the medium. The cells were maintained in a humidified incubator
with an atmosphere of 5% CO2 at 37°C.
Total RNA extraction and quality
control
Total RNA was extracted from cells
(~1×107 cells) using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. RNA levels were measured using a Nanodrop
2000 spectrophotometer (NanoDrop Technologies; Thermo Fisher
Scientific, Inc.) at UV absorbances of 260, 280 and 230 nm. All RNA
samples used met pre-determined quality control standards
(A260/A230 >2.0; A260/A280 >1.8).
miRNA microarray analysis
miRNAs from ~1×107 cells were extracted
using the miRVana™ miRNA isolation kit (cat. no. AM1560, Ambion;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. The miRNAs extracted from three matched pairs of SGC7901
and SGC7901/DDP cells were labeled and hybridized with an
Affymetrix GeneChip miRNA 4.0 Array (Affymetrix; Thermo Fisher
Scientific, Inc.), which contained miRNAs from the miRBase v20
database (http://www.mirbase.org/). The microarray
was scanned by CapitalBio Technology Corporation. The data were
normalized and analyzed using GeneSpring 13.0 software (Agilent
Technologies, Inc.). Student's t-test was used for analysis between
two groups of data. Differential expression of miRNA was defined as
a difference of >2-fold in miRNA expression that was
statistically significant at P<0.05. Cluster analysis and
graphical presentation of the data were performed using Cluster 3.0
software (developed by Michael Eisen, updated by Michiel de Hoon,
University of Tokyo, Human Genome Center).
Prediction of miRNA target genes
The mature miRNA sequences were acquired from
miRBase (http://www.mirbase.org/index.shtml). Potential miRNA
targets were predicted using microRNA.org
(http://www.microrna.org/microrna/getMirnaForm.do)
and miRDB (http://mirdb.org).
Reverse transcription-quantitative PCR
(RT-qPCR)
To detect the expression of miR-362-5p, RT-qPCR was
performed using Hairpin-it™ microRNAs (GenePharma) and U6 snRNA
Normalization RT-qPCR Quantitation kits (GenePharma) according to
the manufacturer's instructions. U6 was selected as the internal
control for miRNA-362-5p expression. The primers of miR-362-5p and
U6 were purchased from GenePharma and the sequences were as
follows: miR-362-5p RT primer
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACACTCAC-3′, PCR
primers, forward 5′-GTCACGAAATCCTTGGAACCTAG-3′ and reverse
5′-TATGGTTGTTCTCGTCTCCTTCTC-3′; and U6 RT primer
5′-CGCTTCACGAATTTGCGTGTCAT-3′, PCR primers, forward
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse
5′-CGCTTCACGAATTTGCGTGTCAT-3′. The miRNA RT reactions were
performed as follows: 25°C for 30 min, 42°C for 30 min, 85°C for 5
min and save at 4°C The PCR reactions were performed as follows:
Prevariant at 95°C for 3 min, cyclic reaction at 95°C for 12 sec
and 62°C for 40 sec and for 40 cycles. For analysis of SUZ12 mRNA
expression, total RNA was reverse-transcribed using a PrimeScript™
RT Reagent kit (Takara Bio, Inc.). RT-qPCR was performed using
SYBR® Premix Ex Taq™ II (Takara Bio, Inc.).
β-actin was used as the internal control. The PCR primers of mRNA
were purchased from Sangon Biotech Co., Ltd. and the sequences of
primers were as follows: SUZ12, forward 5′-ATTCATCGCCAACCTGGATT-3′,
reverse 5′-TGGCCTGCACACAAGAATATG-3′; β-actin, forward
5′-ATTCCTATGTGGGCGACGAG-3′, reverse 5′-AGAGGCGTACAGGGATAGCA-3′. The
mRNA RT reactions were performed as follows: 37°C for 15 min, 85°C
for 5 sec and save at 4°C The PCR reactions were performed as
follows: Prevariant at 95°C for 30 sec, cyclic reaction at 95°C for
5 sec and 60°C for 30 sec and for 40 cycles. All reactions were
performed in triplicate on a LightCycler 96 System (Roche Applied
Science). The relative expression levels of miR-362-5p and SUZ12
mRNA were calculated using the 2−ΔΔCq method (19) and were normalized to the U6 and
β-actin expression levels, respectively.
Lentiviral transfection, stable cell
line establishment and siRNA transfection
The lentiviral vector carrying miR-362-5p
mimic-green fluorescent protein (GFP)-puromycin, the negative
control-GFP-puromycin or miR-362-5p inhibitor small interfering RNA
(siRNA)-GFP-puromycin and the corresponding viruses
[(1×108 plaques forming units (pfu)] were constructed by
GenePharma. Prior to transfection, ~2×105 cells were
plated into six-well plates. When the cells were in the logarithmic
growth phase, they were then infected with lentivirus with a
multiplicity of infection of 10 pfu/cell. RPMI-1640 medium without
penicillin and streptomycin was added to the wells for cell
culture. Cells were transfected for 24 h at 37°C, and the culture
medium containing lentivirus was removed from the wells and fresh
complete medium was added for continued cell culture. At 72 h, the
expression of GFP was observed under a fluorescence microscope to
determine transfection efficiency. As the lentiviral vectors also
carried the gene expressing a puromycin resistance protein,
purified cells were obtained by screening with puromycin. RT-qPCR
was performed to verify the efficiency of miR-362-5p upregulation
or downregulation. The siRNA specific for SUZ12 (si-SUZ12) and
negative control siRNA (si-NC) were synthesized by GenePharma. The
sequences of siRNAs were as follows: si-SUZ12
5′-CUGCCUCCAUUCGAAACAUTTAUGUUUCGAAUGGAGGCAGTT-3′ and si-NC
5′-UUCUCCGAACGUGUCACGUTTACGUGACACGUUCGGAGAATT-3′. Prior to
transfection, ~5×105 cells were seeded in 6-well plates.
After 24 h incubation at 37°C, cells were transfected with si-SUZ12
(100 pmol) or si-NC (100 pmol) using 5 µl Lipofectamine®
2000 (Thermo Fisher Scientific, Inc.) and maintained at 37°C in a
humidified incubator containing 5% CO2 for 48 h.
Western blotting
Cells (~1×107) were washed twice with PBS
and lysed in RIPA buffer (Thermo Fisher Scientific, Inc.)
supplemented with protease inhibitors (Thermo Fisher Scientific,
Inc.) on ice for 30 min. Cell lysates were centrifuged for 15 min
(13,000 × g; 4°C), and the supernatant was collected. Protein
concentrations were quantified by the bicinchoninic acid assay
method (Thermo Fisher Scientific, Inc.). Proteins (60 µg) were
separated by 10% SDS-PAGE. Subsequently, the proteins were
transferred to a PVDF membrane (Merck KGaA) and blocked with 5%
skimmed milk for 4 h at 4°C The membrane was incubated with
antibodies against SUZ12 (cat. no. ab175187; 1:1,000; Abcam),
caspase-3 (cat. no ab184787; 1:1,000; Abcam),
cleaved-poly-(ADP-ribose) polymerase (PARP; cat. no 5625S; 1:1,000;
Cell Signaling Technology, Inc.), NF-κB (cat. no. 8242S; 1:1,000;
Cell Signaling Technology, Inc.), β-actin (cat. no. 4970S; 1:1,000;
Cell Signaling Technology, Inc.) or GAPDH (cat. no. 5174S; 1:1,000;
Cell Signaling Technology, Inc.) at 4°C overnight. Following three
washes with TBS + Tween-20 (0.05%, v/v), the membrane was incubated
with horseradish peroxidase-conjugated secondary antibodies at 4°C
for 4 h. Protein bands were visualized with an enhanced
chemiluminescence reagent kit (Thermo Fisher Scientific, Inc.)
using Fine Do X6 imaging system (Tanon Science and Technology Co.,
Ltd.). Protein expression levels were semi-quantified using the
Tanon 3500/3500R gel imaging system (Tanon Science and Technology
Co., Ltd.) and normalized to the expression of the internal control
GAPDH or β-actin.
Cell viability assay
Cells were plated into 96-well plates
(5×103 cells/well). At 24 h post-transfection, the cells
were treated with a range of concentrations of cisplatin at 37°C.
At 48 h post-transfection, 20 µl MTT (5 mg/ml; Sigma-Aldrich; Merck
KGaA) was added to each well, followed by culturing for 4 h in an
incubator with an atmosphere of 5% CO2 at 37°C. To
dissolve the crystals, 150 µl DMSO (Sigma-Aldrich; Merck KGaA) was
added to each well, and the plates were agitated lightly for 10
min. The absorbance of the samples at 490 nm was measured with an
ELx800 spectrophotometer (BioTek Instruments, Inc.). The median
inhibitory concentration (IC50) for cisplatin was
estimated based on the cell viability using SPSS 16.0 software. All
reactions were performed in triplicate.
Apoptosis assay
The apoptotic rates of SGC7901/DDP and SGC7901 cells
were evaluated by flow cytometry (FCM) with an Annexin
V-FITC/propidium iodide (PI) Cell Apoptosis Detection kit
(BestBio). SGC7901/DDP and SGC7901 cells (~1×106 cells)
were treated with cisplatin at a final concentration of 0.8 and 0.3
µg/ml, respectively. Following 48 h of treatment at 37°C the cells
were collected and washed twice with ice-cold PBS. Cells were
resuspended with 400 µl of 1X binding buffer and maintained at a
final density of approximately 1×106 cells/ml. Annexin
V-FITC (5 µl) was added to the suspension, which was incubated in
the dark for 15 min at room temperature. Following the addition of
PI (10 µl) to the suspension, cells were incubated for an
additional 5 min in the dark at room temperature. Subsequently,
cell apoptosis was assessed (early + late) using a Gallios flow
cytometer (Beckman Coulter, Inc.).
Statistical analysis
All experiments were repeated at least three times.
Statistical analyses were performed using SPSS 16.0 software (SPSS,
Inc.). Data are expressed as the mean ± standard deviation.
Comparison between two groups was analyzed by t-test. ANOVA was
used for comparison between multiple groups followed by the Least
Significant Difference post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
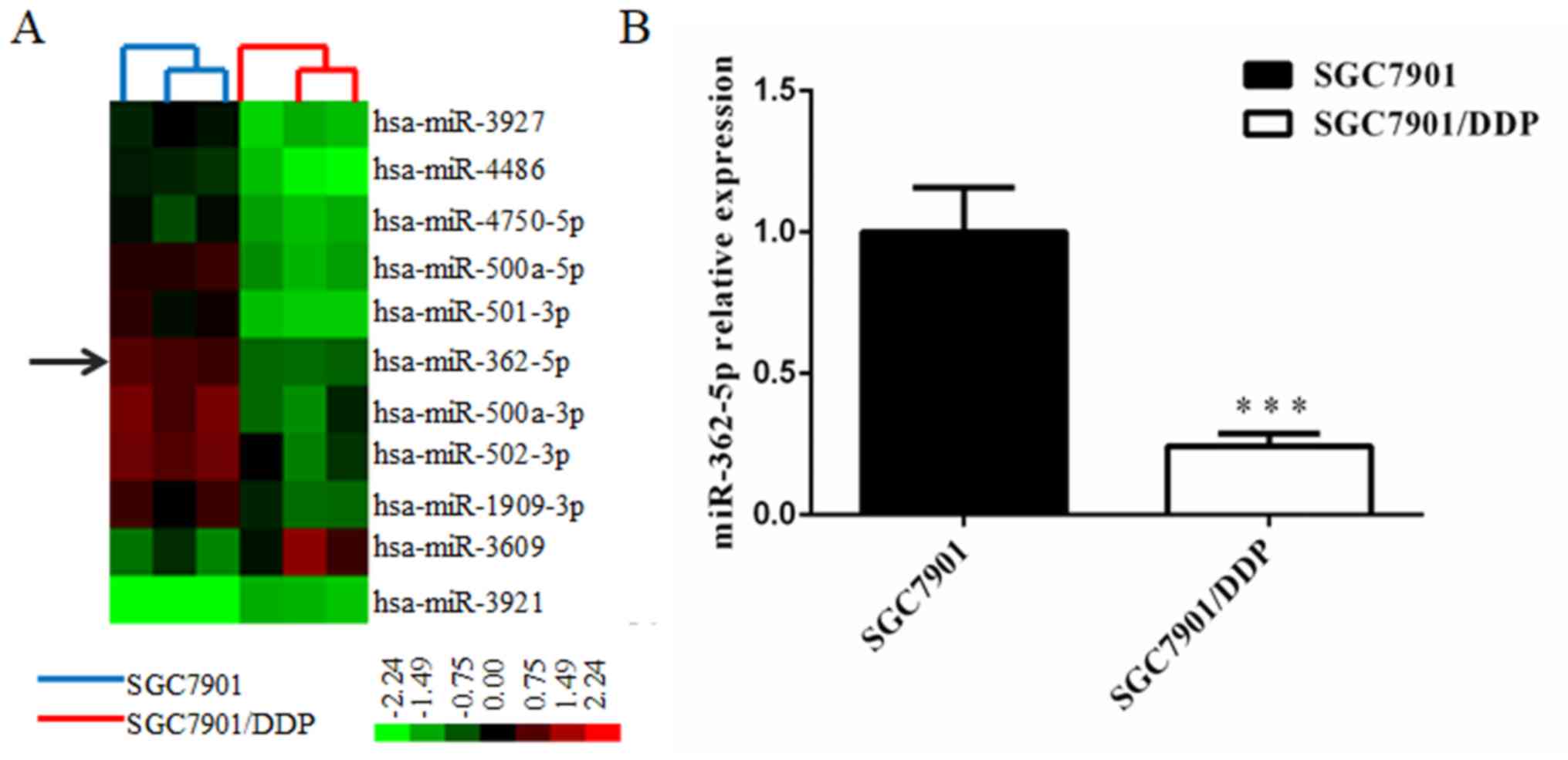
miR-362-5p screening in
cisplatin-sensitive and cisplatin-resistant GC cells
The differential miRNA expression profiles between
SGC7901/DDP cells and SGC7901 cells were obtained by miRNA
microarray analysis. The Affymetrix miRNA GeneChip 4.0 was used to
scan and quantify the signal intensity of probes of 1,316 human
mature miRNAs on the chips for the two cell lines. The results
indicated that 48 miRNAs were significantly differentially
expressed (>2-fold) in SGC7901/DDP cells relative to the
parental cells, including 19 upregulated miRNAs and 29
downregulated miRNAs. miR-362-5p was revealed to be downregulated
in SGC7901/DDP cells and was selected for further research to
determine its function in the cisplatin resistance of GC cells.
Cluster analysis was performed with Cluster 3.0 software (Fig. 1A). To study the potential effects of
miRNA-362-5p on the cisplatin resistance of GC, the difference in
miR-362-5p expression between SGC7901 cells and SGC7901/DDP cells
was determined by RT-qPCR. The results revealed that the expression
level of miR-362-5p was significantly lower in SGC7901/DDP cells
compared with expression in SGC7901cells (P<0.01; Fig. 1B). These results suggested that
reduced expression of miR-362-5p may be associated with cisplatin
resistance in SGC7901/DDP cells.
Overexpression of miR-362-5p reverses
cisplatin resistance in SGC7901/DDP cells
To further determine the association between
cisplatin resistance and the regulation of miR-362-5p expression,
the effects of miR-362-5p upregulation on cisplatin resistance in
cisplatin-resistant SGC7901/DDP cell lines were investigated.
Lentiviral plasmids carrying miR-362-5p mimic or negative control
miRNA were transfected into SGC7901/DDP cells. After screening with
puromycin, the GFP-positive rate of the cells subjected to
lentiviral transfection was >90% based on fluorescence
microscopy (Fig. 2A and B). RT-qPCR
was performed to verify the transfection efficiency in the
constructed cell lines, and the results demonstrated that
miR-362-5p expression levels were significantly increased
(P<0.01; Fig. 2C). After stable
cell line establishment, the cells were treated with different
concentrations of cisplatin for 48 h. The IC50 for
cisplatin was estimated based on cell viability. The SGC7901/DDP
cells transfected with miR-362-5p mimic exhibited a notably lower
survival status compared with control SGC7901/DDP-NC and
SGC7901/DDP cells (IC50, 1.609±0.332 vs. 5.133±0.569 and
5.161±0.641 µg/ml, respectively; P<0.01; Fig. 2D). These results indicated that
upregulation of miR-362-5p may enhance the cisplatin sensitivity of
cisplatin-resistant GC cells.
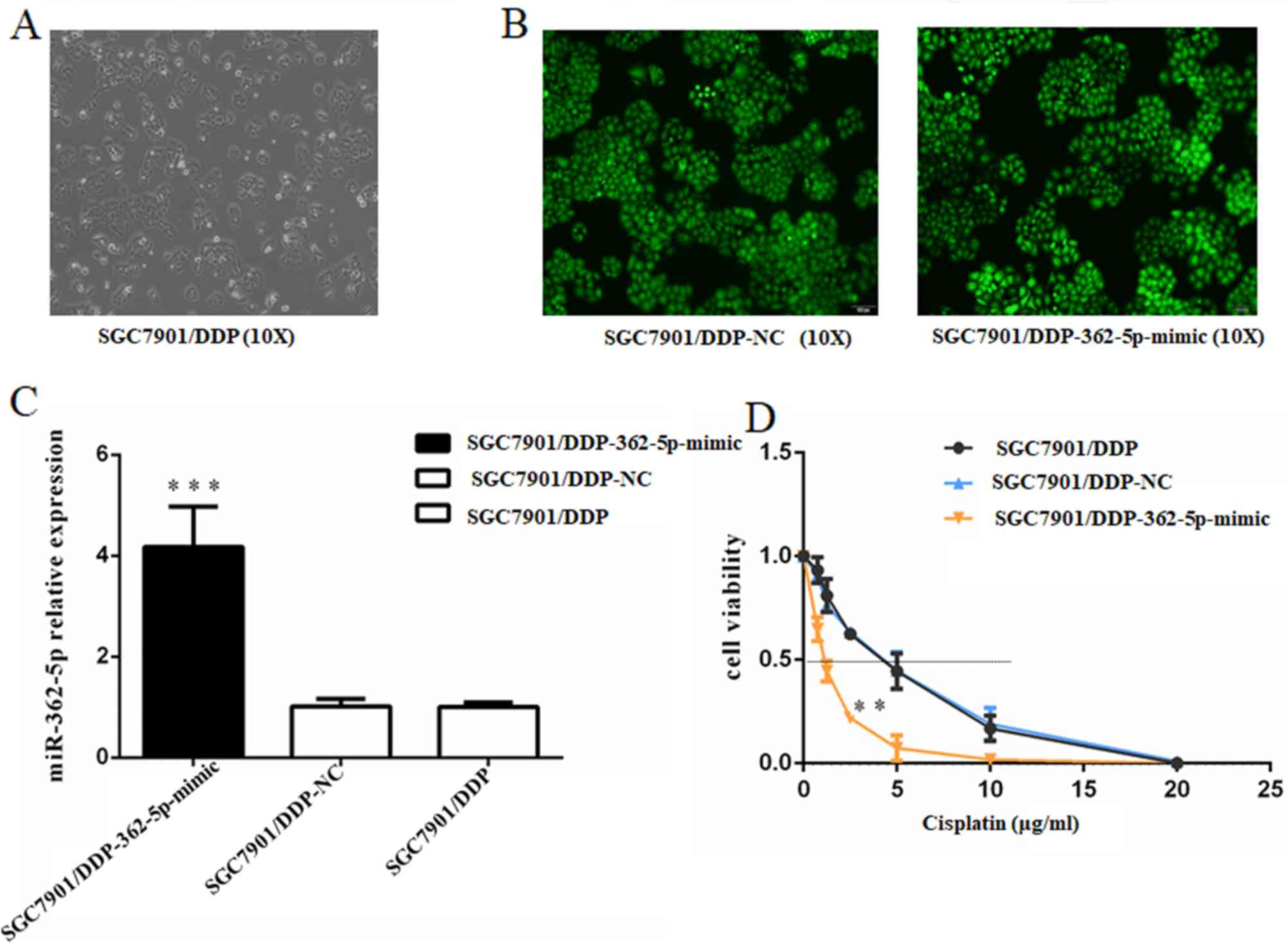 | Figure 2.Upregulation of miR-362-5p reverses
the cisplatin resistance of SGC7901/DDP cells. (A) Light microscopy
(magnification, ×100) and (B) lentiviral transfection assay of
SGC7901/DDP cells (magnification, ×100). Cells that were
successfully transfected expressed green fluorescent protein. (C)
Reverse transcription-quantitative PCR analysis demonstrated that
the expression level of miR-362-5p was markedly increased in
SGC7901/DDP cells transfected with miR-362-5p mimic compared with
NC-transfected and untransfected cells. (D) Viability of cells
treated with cisplatin for 48 h. MTT assays were conducted on
SGC7901/DDP cells treated with various concentrations of cisplatin
(0, 0.75, 1.25, 2.5, 5, 10 or 20 µg/ml). The IC50 for
cisplatin was estimated based on the cell viability. The
IC50 for cisplatin of the miR-362-5p mimic-transfected
cells was notably lower compared with the controls. n=3;
***P<0.001 vs. SGC7901/DDP-NC and SGC7901/DDP; **P<0.01 vs.
SGC7901/DDP-NC and SGC7901/DDP; IC50, median inhibitory
concentration; miR-362-5p, microRNA-362-5p; NC, negative
control. |
Inhibition of miR-362-5p enhances the
cisplatin resistance of SGC7901 cells
Based on the aforementioned results, the effects of
miR-362-5p inhibition on cisplatin-induced cytotoxicity in
cisplatin-sensitive SGC7901 cell line was further investigated.
SGC7901 cells were transfected with lentiviral plasmids carrying
miR-362-5p inhibitor or its negative control. The transfected cells
were screened with puromycin and then observed for fluorescence
(Fig. 3A and B). The data from
RT-qPCR revealed that the miR-362-5p inhibitor significantly
decreased the expression level of miR-362-5p in SGC7901 cells
compare with negative control and blank control (P<0.001;
Fig. 3C), which confirmed that the
cells were successfully transfected. Subsequently, a range of
concentrations of cisplatin was applied to the cultures and the
IC50 for cisplatin was estimated based on cell
viability. Cisplatin sensitivity was significantly attenuated in
SGC7901 cells transfected with the miR-362-5p inhibitor compared
with the control SGC7901-NC and SGC7901 cells (IC50,
0.676±0.042 vs. 0.286±0.025 and 0.300±0.009 µg/ml, respectively;
P<0.01; Fig. 3D). These data
verified that downregulation of miR-362-5p enhances the cisplatin
resistance of cisplatin-sensitive GC cell lines.
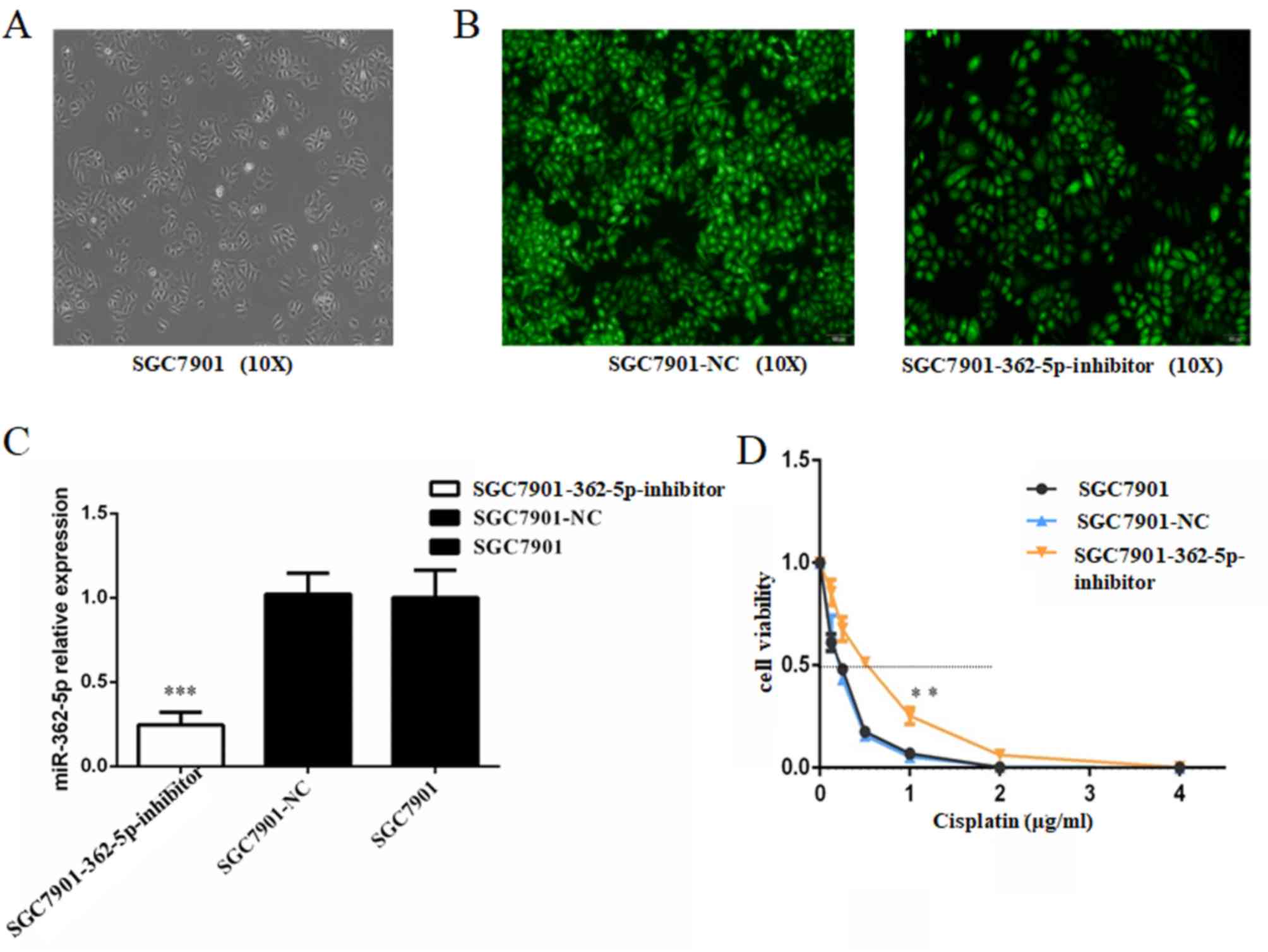 | Figure 3.Inhibition of miR-362-5p reduces the
cisplatin sensitivity of SGC7901 cells. (A) Light microscopy
(magnification, ×100) and (B) lentiviral plasmid transfection assay
of SGC7901 cells (magnification, ×100). Cells that were
successfully transfected expressed green fluorescent protein. (C)
Reverse transcription-quantitative PCR analysis demonstrated that
the expression level of miR-362-5p was markedly decreased in
SGC7901 cells transfected with miR-362-5p inhibitor compared with
the control groups. (D) Viability of cells treated with cisplatin
for 48 h. MTT assays were conducted on SGC7901 cells treated with
various concentrations of cisplatin (0, 0.125, 0.25, 0.5, 1.0, 2.0
or 4.0 µg/ml). The IC50 for cisplatin was estimated
based on the cell viability. The IC50 for cisplatin of
SGC7901 cells transfected with miR-362-5p inhibitor was notably
higher compared with the control groups (P<0.01). n=3;
**P<0.01, ***P<0.001; IC50, median inhibitory
concentration; miR-362-5p, microRNA-362-5p; NC, negative
control. |
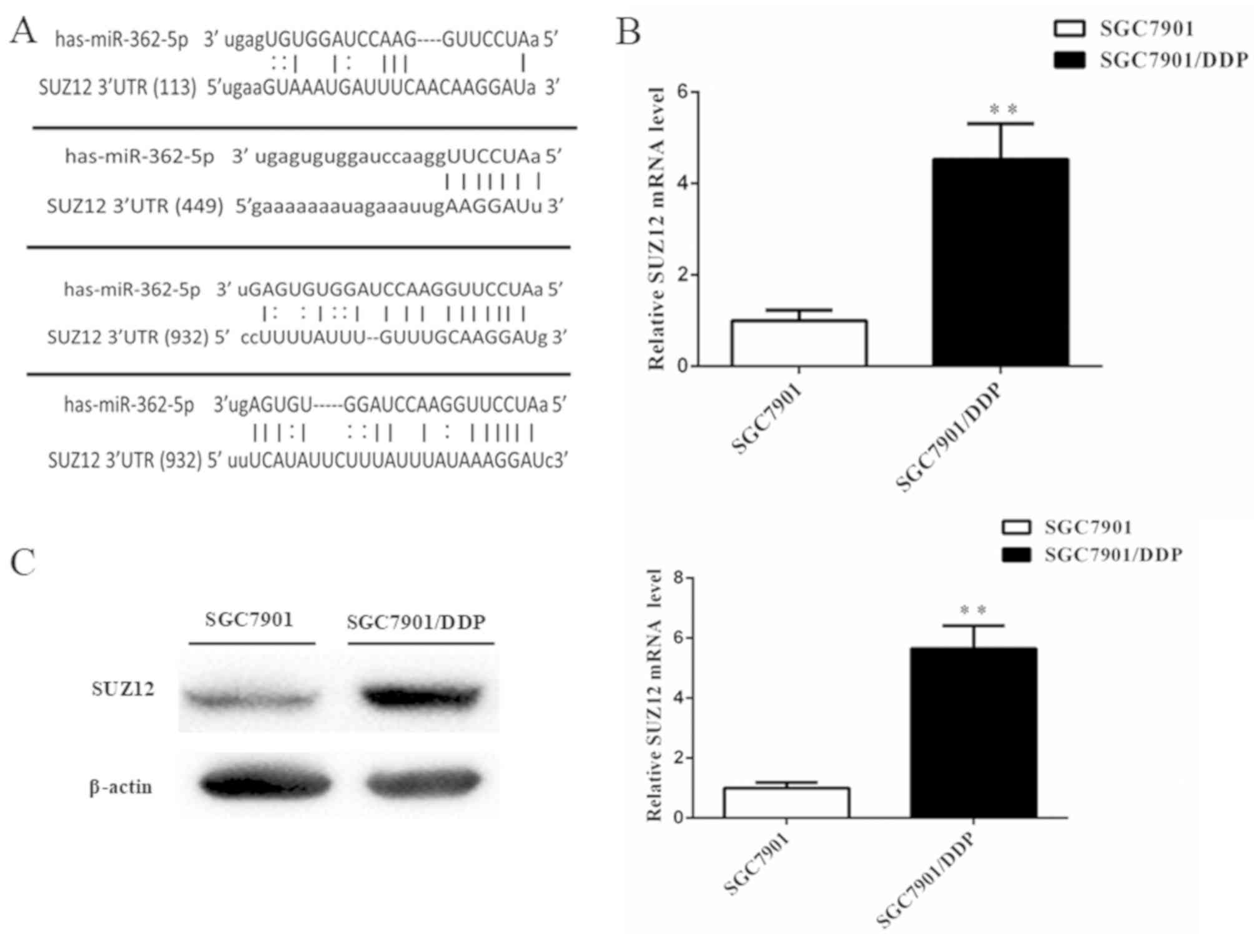
miR-362-5p modulates cisplatin
resistance by targeting SUZ12 in GC cells
To explore the mechanism of miR-362-5p-mediated
cisplatin resistance in human GC cells, the bioinformatics
algorithms in the microRNA.org and miRDB databases were
used to search for possible targets of miR-362-5p. The databases
predicted SUZ12 as a target of miR-362-5p. The 3′-UTR of SUZ12 mRNA
contains four miR-362-5p target sites at nucleotide positions
113–139, 449–472, 932–954 and 1,196-1,222 (Fig. 4A). A previous study has demonstrated
that SUZ12 acts as an oncogene in GC cells (16). However, whether SUZ12 is involved in
the cisplatin resistance of GC cells is not fully understood. The
mRNA and protein expression levels of SUZ12 in SGC7901/DDP cells
were markedly higher compared with the SGC7901 cells based on
RT-qPCR and western blot analysis (Fig.
4B and C), which suggested that there may be an association
between miRNA-362-5p and SUZ12 in cisplatin-resistant GC.
To study the role of SUZ12, si-SUZ12 was used to
knock down SUZ12 expression in SGC7901/DDP cells, and the protein
expression level of SUZ12 was significantly suppressed (Fig. 5A). MTT assays confirmed that
knockdown of SUZ12 in SGC7901/DDP cells treated with cisplatin for
48 h significantly decreased cell survival status (IC50,
2.569±0.479 µg/ml in SGC7901/DDP-SUZ12-siRNA cells vs. 5.097±0.629
µg/ml in SGC7901/DDP cells; P<0.01; Fig. 5B). Therefore, downregulation of SUZ12
enhanced the cisplatin sensitivity of SGC7901/DDP cells. SUZ12 has
been demonstrated to act as a positive regulator of NF-κB signaling
(20). In the present study, western
blot analysis revealed that the NF-κB/p65 protein levels were
significantly decreased following SUZ12-knockdown compared with the
control group (Fig. 5C).
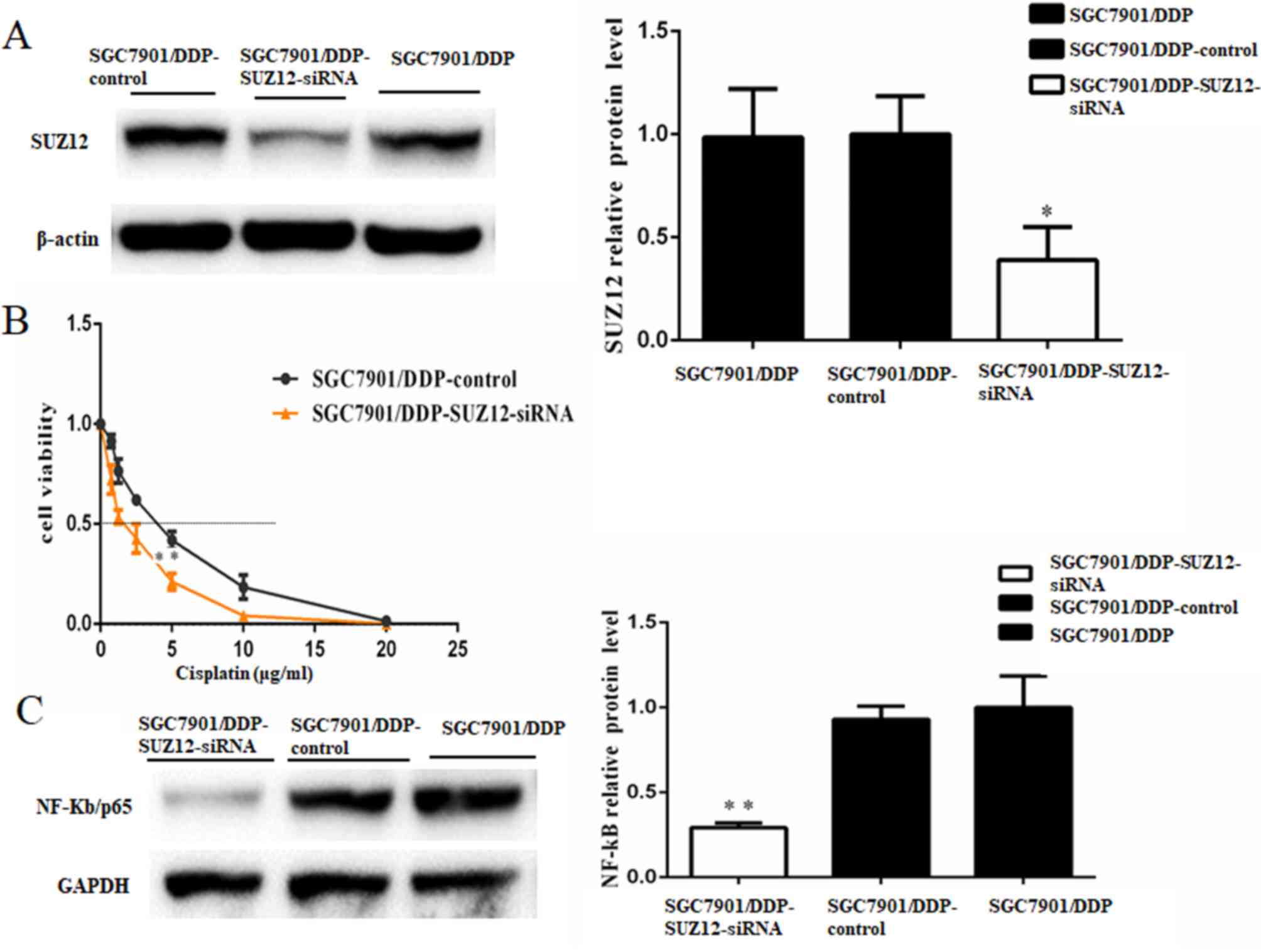 | Figure 5.SUZ12 acts as a promoter of cisplatin
resistance in GC cells. (A) Western blotting demonstrated that the
protein expression level of SUZ12 was significantly reduced by
si-SUZ12 transfection; β-actin was used as a loading control. (B)
Viability of cells treated with cisplatin for 48 h. MTT assays were
conducted on SGC7901/DDP cells treated with various concentrations
of cisplatin (0, 0.75, 1.25, 2.5, 5, 10 or 20 µg/ml). The
IC50 for cisplatin was estimated based on the cell
viability. The IC50 for cisplatin of the SGC7901/DDP
cells transfected with SUZ12-siRNA was notably lower compared with
the control cells (P<0.01). (C) Western blotting demonstrated
that NF-κB/p65 protein level was decreased in SGC7901/DDP cells
transfected with SUZ12-siRNA compared with control cells; GADPH was
used as a loading control. n=3; *P<0.05; **P<0.01;
IC50, median inhibitory concentration; miR-362-5p,
microRNA-362-5p; NC, negative control; siRNA, short interfering
RNA; SUZ12, suppressor of zeste 12 protein. |
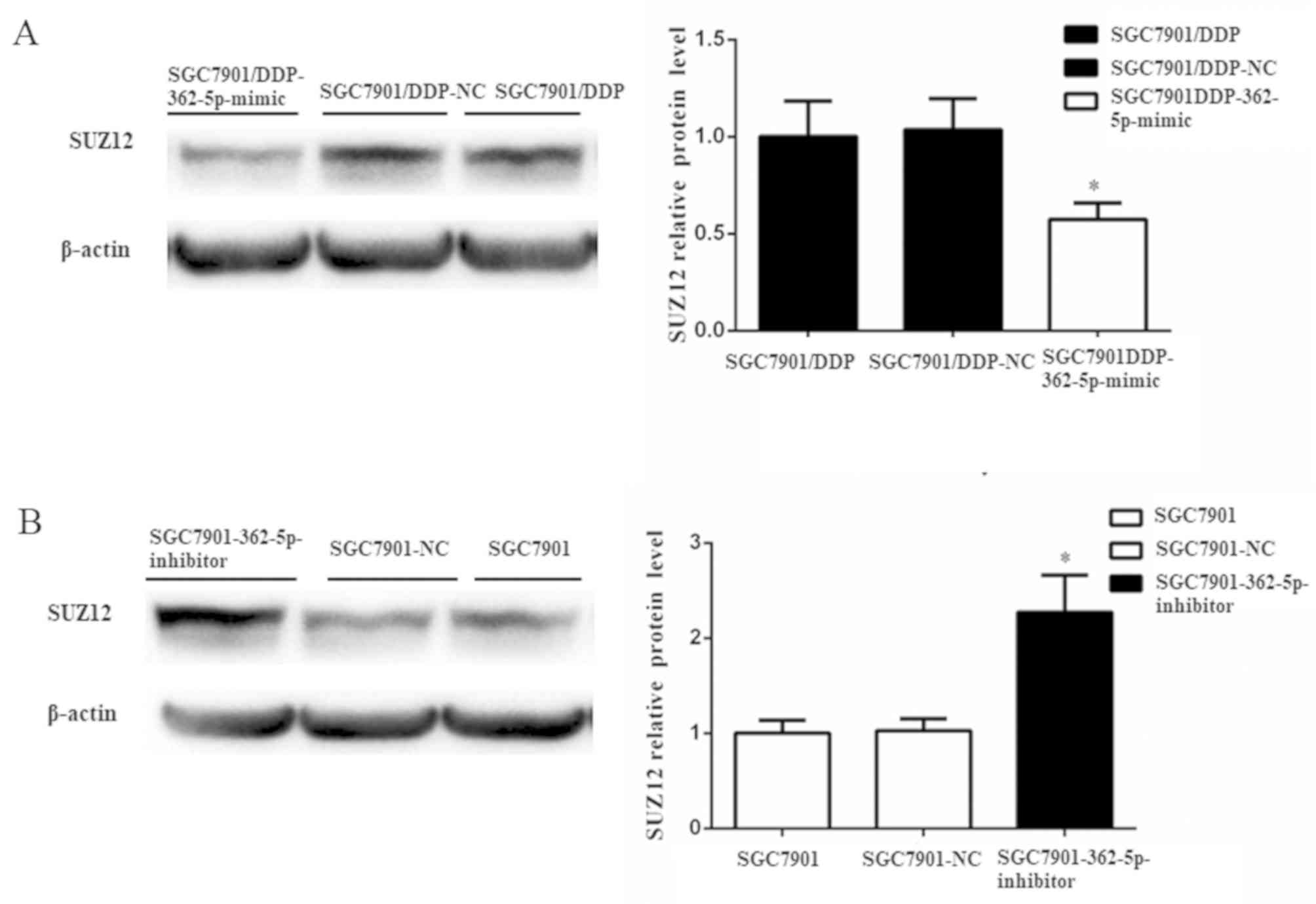
Subsequently, the changes in the expression of SUZ12
protein were further evaluated by western blot analysis after the
modulation of miR-362-5p expression. SUZ12 protein expression was
significantly reduced in SGC7901/DDP cells transfected with
miR-362-5p mimic compared with those transfected with its negative
control (Fig. 6A). Additionally,
elevated expression of SUZ12 protein was detected following the
downregulation of miR-362-5p in SGC7901 cells (Fig. 6B). Taken together, the results of the
present study demonstrated that miR-362-5p may modulate cisplatin
resistance by targeting SUZ12.
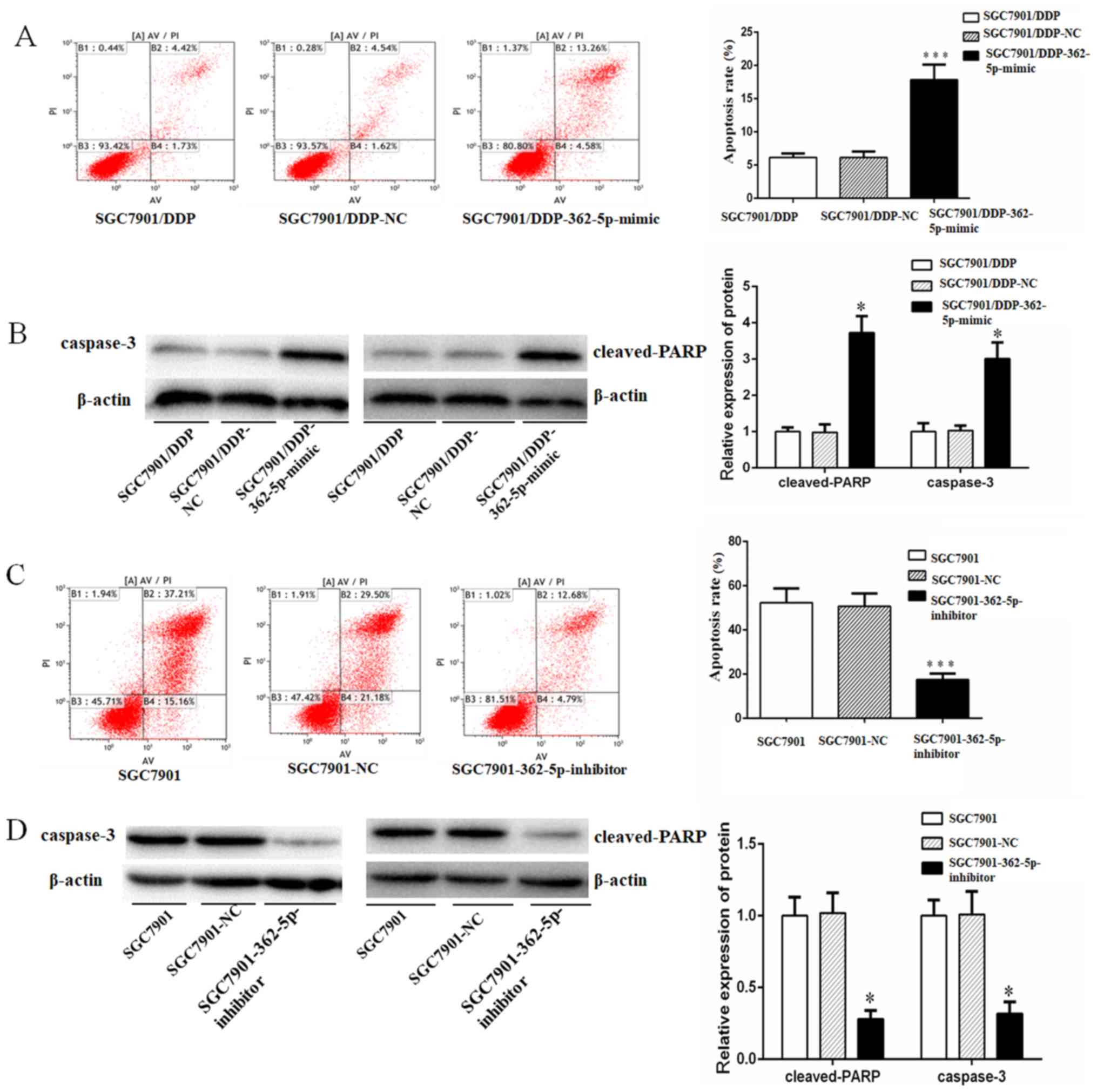
miR-362-5p sensitizes GC cells to
cisplatin-induced apoptosis
A previous study has demonstrated that the
development of chemotherapy resistance in a variety of cancer types
is associated with decreased susceptibility to drug-induced
apoptosis (21). Therefore, it was
hypothesized that miR-362-5p may also serve a role in the cisplatin
resistance of GC cells through the modulation of apoptosis. To test
this hypothesis, cisplatin-induced apoptosis in SGC7901 and
SGC7901/DDP cells transfected with miR-362-5p negative control,
mimic or inhibitor, as well as untransfected cells, was quantified
by flow cytometry. Additionally, the protein expression of
apoptotic markers caspase-3 and cleaved PARP was determined by
western blotting. The activation of caspase-3 and its downstream
target PARP mediate apoptosis by chromatin condensation and DNA
fragmentation (22). Following 48-h
treatment with cisplatin at a final concentration of 0.8 µg/ml, the
SGC7901/DDP cells overexpression miR-362-5p exhibited significantly
higher apoptotic rates and expression levels of caspase-3 and
cleaved-PARP compared with the control cells (Fig. 7A and B). Furthermore, following 48-h
treatment with cisplatin at a final concentration of 0.3 µg/ml, the
SGC7901 cells transfected with miR-362-5p inhibitor displayed
significantly lower cisplatin-induced apoptosis, and expression
levels of caspase-3 and cleaved-PARP compared with the control
cells (Fig. 7C and D). Taken
together, the results of the present study demonstrated that
miR-362-5p may modulate the cisplatin resistance of human GC in
part by sensitizing cells to cisplatin-induced apoptosis.
Discussion
GC is one of the most common types of malignancy
worldwide (1). To date, surgery and
chemotherapy have been the major methods of GC treatment (23). For post-operative patients with GC or
patients with unresectable GC, chemotherapy may increase survival
rates (3). Cisplatin-based
chemotherapy, as the most common chemotherapy regimen for GC,
induces tumor cell death through DNA damage (24). However, its therapeutic effectiveness
is limited, as demonstrated by the occurrence of cisplatin
resistance, which is the primary cause of disease recurrence and
metastasis, and which results in GC treatment failure (4). Numerous studies (25–27) have
indicated various mechanisms of drug resistance; for instance,
mutations of target genes, increases in drug efflux, enhancement of
DNA repair activity or dysfunction of pro-apoptotic proteins, the
mechanisms of cancer cell drug resistance are still not clearly
understood. Therefore, the identification of novel molecular
mechanisms of cisplatin resistance is urgently needed to improve
the survival status of patients with GC.
miRNAs have been demonstrated to regulate the
progression of various types of cancer (28,29). It
has been extensively reported that miRNAs can modulate cisplatin
resistance in tumor cells. For instance, it has been reported that
miR-130a targeting copper transporter protein 1 (30) and miR-1284 targeting high mobility
group box 1 (31) can regulate
cisplatin resistance in cervical cancer. Furthermore, miR-144-3p
targeting nuclear factor erythroid 2-related factor 2 (32) and microRNA-133b targeting epidermal
growth factor receptor (33) also
regulate lung cancer resistance to cisplatin. To explore novel
molecular mechanisms of cisplatin resistance, the differential
miRNA expression profiles among GC cells (cisplatin-resistant
SGC7901/DDP cells vs. cisplatin-sensitive SGC7901 cells) were
determined by miRNA microarray analysis. A total of 48 miRNAs,
including miR-362-5p, were identified as significantly
differentially expressed (by >2-fold). miR-362-5p, which was
identified by the miRNA microarray screen and confirmed by RT-qPCR
analysis, was notably downregulated in human GC SGC7901/DDP cells
compared with SGC7901 cells. Therefore, it was hypothesized that
miR-362-5p may be involved in the cisplatin resistance of GC cells.
Though miR-362-5p has been reported to contribute to the regulation
of cancer processes (11,12), its role in SGC7901/DDP cells is not
fully understood. Results from the present study demonstrated the
regulation of miR-362-5p in GC SGC7901 and SGC7901/DDP cells. The
results also revealed that upregulation of miR-362-5p in GC cells
increased cisplatin sensitivity and cisplatin-induced apoptosis,
whereas downregulation of miR-362-5p in GC cells attenuated
cisplatin sensitivity and cisplatin-induced apoptosis. These
results revealed that miR-362-5p may act as a novel effector
molecule that serves crucial roles in the regulation of the
cisplatin resistance of GC cells.
Generally, miRNAs bind to a target mRNA at the
3′-UTR to promote target mRNA degradation or block target mRNA
translation (6,7). Bioinformatics analysis predicted that
SUZ12 may be a target of miR-362-5p. SUZ12 is a carcinogenic factor
and a catalyst of migration and invasion in various cancer types
(16–18). The expression of SUZ12 is
significantly increased in GC tissues, and increased SUZ12
expression promotes GC cell proliferation and metastasis (16). Furthermore, SUZ12 is involved in long
non-coding RNA (lncRNA)-mediated promotion of cancer progression
(34). However, whether SUZ12 is
involved in the cisplatin resistance of GC cells is still unknown.
To verify whether the cisplatin resistance induced by miR-362-5p
was mediated through SUZ12, further experiments are required. The
results of the present study demonstrated that the SUZ12 expression
level in SGC7901/DDP cells was markedly higher compared with
expression in SGC7901 cells and that knockdown of SUZ12 enhanced
the cisplatin sensitivity of SGC7901/DDP cells. These results
implied that SUZ12 may act as a promoter of cisplatin resistance in
GC. Notably, a previous study has confirmed that SUZ12 acts as
positive regulator for NF-κB signaling (20). The NF-κB signaling pathway is
involved in anti-apoptosis, inflammation and immunity. The present
study demonstrated that the NF-κB/p65 protein levels were
significantly decreased following knockdown of SUZ12. Therefore,
SUZ12 may inhibit the cisplatin-induced apoptosis of GC cells by
increasing the expression of NF-κB/p65. Notably, SUZ12 protein
expression was increased in SGC7901 cells transfected with
miR-362-5p inhibitor. By contrast, the expression levels of SUZ12
protein were reduced in SGC7901/DDP cells with upregulated
miR-362-5p expression. Therefore, SUZ12 may be a target of
miR-362-5p in GC cells.
In summary, the results of the present study
revealed that miR-362-5p may sensitize SGC7901/DDP cells to
cisplatin by regulating the SUZ12/NF-κB/p65 pathway. miR-362-5p may
be a potential therapeutic target in combination with anti-GC
chemotherapies. One potential adjunct therapy may be treatment with
mimics that reinforce the expression and function of miR-362-5p.
However, miR-362-5p may also non-specifically bind to other
functional mRNAs to cause adverse side effects. In addition, it
should be noted that experiments were conducted in only one type of
GC cells. Further investigation on miR-362-5p function is therefore
required.
Acknowledgements
Not applicable.
Funding
The present study was funded by The Anhui Provincial
Science and Technology Plan Project (grant no. 1604b0602027).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW, MeG, YA and KG performed the experiments and
wrote the manuscript. KG made substantial contributions to
conception and design of the manuscript. XW, MeG and YA were
responsible for the design of the experiments. MiG, WL, YZ, XX, QZ
and HW analyzed the experimental data. KG assisted with the
statistical analysis. KG critically revised the manuscript and
provided final approval, and made substantial contributions to
conception and design of the study. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Petrillo A, Pompella L, Tirino G,
Pappalardo A, Laterza MM, Caterino M, Orditura M, Ciardiello F,
Lieto E, Galizia G, et al: Perioperative treatment in resectable
gastric cancer: Current perspectives and future directions. Cancers
(Basel). 11(pii): E3992019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marin JJ, Al-Abdulla R, Lozano E, Briz O,
Bujanda L, Banales JM and Macias RI: Mechanisms of resistance to
chemotherapy in gastric cancer. Anticancer Agents Med Chem.
16:318–334. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Orditura M, Galizia G, Sforza V,
Gambardella V, Fabozzi A, Laterza MM, Andreozzi F, Ventriglia J,
Savastano B, Mabilia A, et al: Treatment of gastric cancer. World J
Gastroenterol. 20:1635–1649. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kaufmann SH and Earnshaw WC: Induction of
apoptosis by cancer chemotherapy. Exp Cell Res. 256:42–49. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li C, Zou J, Zheng G and Chu J: MiR-30a
decreases multidrug resistance (MDR) of gastric cancer cells. Med
Sci Monit. 0:02016.PubMed/NCBI
|
|
9
|
Bao J, Xu Y, Wang Q, Zhang J, Li Z, Li D
and Li J: miR-101 alleviates chemoresistance of gastric cancer
cells by targeting ANXA2. Biomed Pharmacother. 92:1030–1037. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ying G, Wu R, Xia M, Fei X, He QE, Zha C
and Wu F: Identification of eight key miRNAs associated with renal
cell carcinoma: A meta-analysis. Oncol Lett. 16:5847–5855.
2018.PubMed/NCBI
|
|
11
|
Ni F, Zhao H, Cui H, Wu Z, Chen L, Hu Z,
Guo C, Liu Y, Chen Z, Wang X, et al: MicroRNA-362-5p promotes tumor
growth and metastasis by targeting CYLD in hepatocellular
carcinoma. Cancer Lett. 356:809–818. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ni F, Gui Z, Guo Q, Hu Z, Wang X, Chen D
and Wang S: Downregulation of miR-362-5p inhibits proliferation,
migration and invasion of human breast cancer MCF7 cells. Oncol
Lett. 11:1155–1160. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cao R, Wang L, Wang H, Xia L,
Erdjument-Bromage H, Tempst P, Jones RS and Zhang Y: Role of
histone H3 lysine 27 methylation in Polycomb-group silencing.
Science. 298:1039–1043. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Margueron R and Reinberg D: The Polycomb
complex PRC2 and its mark in life. Nature. 469:343–349. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He LJ, Cai MY, Xu GL, Li JJ, Weng ZJ, Xu
DZ, Luo GY, Zhu SL and Xie D: Prognostic significance of
overexpression of EZH2 and H3k27me3 proteins in gastric cancer.
Asian Pac J Cancer Prev. 13:3173–3178. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia R, Jin FY, Lu K, Wan L, Xie M, Xu TP,
De W and Wang ZX: SUZ12 promotes gastric cancer cell proliferation
and metastasis by regulating KLF2 and E-cadherin. Tumour Biol.
36:5341–5351. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li H, Cai Q, Wu H, Vathipadiekal V, Dobbin
ZC, Li T, Hua X, Landen CN, Birrer MJ, Sánchez-Beato M and Zhang R:
SUZ12 promotes human epithelial ovarian cancer by suppressing
apoptosis via silencing HRK. Mol Cancer Res. 10:1462–1472. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu C, Shi X, Wang L, Wu Y, Jin F, Bai C
and Song Y: SUZ12 is involved in progression of non-small cell lung
cancer by promoting cell proliferation and metastasis. Tumour Biol.
35:6073–6082. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Su SK, Li CY, Lei PJ, Wang X, Zhao QY, Cai
Y, Wang Z, Li L and Wu M: The EZH1-SUZ12 complex positively
regulates the transcription of NF-kB target genes through
interaction with UXT. J Cell Sci. 129:2343–2353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhu W, Shan X, Wang T, Shu Y and Liu P:
miR-181b modulates multidrug resistance by targeting BCL2 in human
cancer cell lines. Int J Cancer. 127:2520–2529. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Petrelli F, Zaniboni A, Ghidini A, Ghidini
M, Turati L, Pizzo C, Ratti M, Libertini M and Tomasello G: Timing
of adjuvant chemotherapy and survival in colorectal, gastric, and
pancreatic cancer. A systematic review and meta-analysis. Cancers
(Basel). 11(pii): E5502019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu W, Chen Q, Wang Q, Sun Y, Wang S, Li A,
Xu S, Røe OD, Wang M, Zhang R, et al: JWA reverses cisplatin
resistance via the CK2-XRCC1 pathway in human gastric cancer cells.
Cell Death Dis. 5:e15512014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Galluzzi L, Senovilla L, Vitale I, Michels
J, Martins I, Kepp O, Castedo M and Kroemer G: Molecular mechanisms
of cisplatin resistance. Oncogene. 31:1869–1883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ghosh S: Cisplatin: The first metal based
anticancer drug. Bioorg Chem. 88:1029252019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rocha CRR, Silva MM, Quinet A, Cabral-Neto
JB and Menck CFM: DNA repair pathways and cisplatin resistance: An
intimate relationship. Clinics (Sao Paulo). 73 (Suppl 1):e478s2018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin Y, Gu Q, Sun Z, Sheng B, Qi C, Liu B,
Fu T, Liu C and Zhang Y: Upregulation of miR-3607 promotes lung
adenocarcinoma proliferation by suppressing APC expression. Biomed
Pharmacother. 95:497–503. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Herr I, Sähr H, Zhao Z, Yin L, Omlor G,
Lehner B and Fellenberg J: MiR-127 and miR-376a act as tumor
suppressors by in vivo targeting of COA1 and PDIA6 in giant cell
tumor of bone. Cancer Lett. 409:49–55. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Feng C, Ma F, Hu C, Ma JA, Wang J, Zhang
Y, Wu F, Hou T, Jiang S, Wang Y and Feng Y: SOX9/miR-130a/CTR1 axis
modulates DDP-resistance of cervical cancer cell. Cell Cycle.
17:448–458. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen J and Li G: MiR-1284 enhances
sensitivity of cervical cancer cells to cisplatin via
downregulating HMGB1. Biomed Pharmacother. 107:997–1003. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yin Y, Liu H, Xu J, Shi D, Zhai L, Liu B,
Wang L, Liu G and Qin J: miR-144-3p regulates the resistance of
lung cancer to cisplatin by targeting Nrf2. Oncol Rep.
40:3479–3488. 2018.PubMed/NCBI
|
|
33
|
Li B, Ding CM, Li YX, Peng JC, Geng N and
Qin WW: Over-regulation of microRNA-133b inhibits cell
proliferation of cisplatin-induced non-small cell lung cancer cells
through PI3K/Akt and JAK2/STAT3 signaling pathway by targeting
EGFR. Oncol Rep. 39:1227–1234. 2018.PubMed/NCBI
|
|
34
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|