Introduction
Pancreatic cancer is the seventh leading cause of
cancer-associated mortality in males and females worldwide, with
high incidence rates in both sexes (1,2).
Curative therapy comprises surgical resection; however, the
five-year survival rate for patients remains <5% (3). In addition, the current first-line
chemotherapeutic treatment, gemcitabine, has not improved the low
overall survival rate (3).
Therefore, there is an urgent requirement for novel therapeutic
targets and strategies for the treatment of patients with
pancreatic cancer.
Neddylation is a post-translational modification of
proteins (4). Neural precursor
cell-expressed, developmentally downregulated 8 (NEDD8) is a
ubiquitin-like protein that activates the largest member of the
ubiquitin E3 ligase family, cullin ring ligase (CRL) (5). Substrate-bound NEDD8 is unable to
degrade proteins; however, it is able to modulate their
conformation, stability, localization and function (6). The neddylation pathway has been
observed to be overactivated in multiple cancer types, including
liver (7), lung (8) and colon cancer (9). These previous studies suggested that
neddylation serves an important role in cancer. MLN4924 (also known
as pevonedistat) is a specific neddylation inhibitor that binds
irreversibly to NEDD8 activating enzyme E1 subunit 1 (NAE1)
(10). MLN4924 has shown promising
efficacy against a variety of cancers, including liver cancer
(11), lung cancer (8), cholangiocarcinoma (12) and glioblastoma (13). In addition, MLN4924 has been
demonstrated to enhance the effects of autophagy inhibitors in
liver cancer (14), as well as
sensitizing pancreatic cells to radiotherapy by promoting DNA
damage (15). Notably, MLN4924 was
revealed to enhance the therapeutic effects of gemcitabine in
pancreatic cancer by increasing cancer cell apoptosis (16). However, its role in modulating
neddylation and its efficacy in combination regimens require
further study.
In the present study, the effect of combined MLN4924
and cisplatin treatment in pancreatic cancer was investigated, and
the expression profile of neddylation pathway members was examined.
The results demonstrated that molecules involved in the neddylation
pathway were overexpressed at the protein, but not the mRNA level,
and only NAE1 overexpression was a predictor of poor prognosis in
patients with pancreatic cancer. Combined MLN4924 and cisplatin
treatment exhibited a marked therapeutic effect on pancreatic
cancer cells both in vivo and in vitro. These
mechanistic studies revealed that combined therapy suppressed
pancreatic cancer cell growth by promoting DNA damage.
Materials and methods
Cell lines, culture conditions and
reagents
The human pancreatic cancer cell lines BXPC3 and
SW1990 (purchased from the Cell Bank of Typical Culture
Preservation Commission of the Chinese Academy of Sciences) were
cultured in DMEM (HyClone; GE Healthcare Life Sciences) containing
10% FBS (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin solution at 37°C (5% CO2).
MLN4924 and cisplatin were purchased from MedChemExpress. For in
vitro studies, an MLN4924 stock solution (50 mM) was prepared
in DMSO, aliquoted and stored at −20°C until use at a final
concentration of 0.3 µM. For in vivo studies, MLN4924 was
dissolved in 15% 2-hydroxypropyl-β-cyclodextrin (HPBCD); the
solution was freshly prepared each week and stored in the dark at
4°C prior to use at 20 mg/kg. Cisplatin was used at a concentration
of 0.25 µM for in vitro, and 3 mg/kg for in vivo
studies.
Patients and specimens
The pancreatic cancer tissue samples used in the
present study were derived from patients that underwent primary
curative tumor resection at the Department of Hepatobiliary Surgery
(Yongchuan Hospital, Chongqing, China) between October 2015 and
February 2016. A total of 12 paired tumor and normal adjacent
samples, from nine males and three females with an average age of
46 years, were used for western blotting analysis, and 42 paired
tumor and normal adjacent tissues (including the 12 paired samples
used for western blotting analysis), from 32 males and 10 females
with an average age of 51 years, were used for reverse
transcription-quantitative PCR (RT-qPCR) analysis. Patients had not
received any radio- or chemotherapeutic treatment prior to surgery.
Ethical approval was obtained from the Ethics Committee of
Yongchuan Hospital of Chongqing Medical University (Chongqing,
China) and written informed consent for tissue collection and the
use of patient data for scientific purposes was obtained from all
participants.
Immunohistochemical tissue
staining
Subcutaneous tumor tissues from mice were
immunohistochemically stained with anti-cleaved (c)-caspase3
(1:100; cat. no. 9664S, Cell Signaling Technology, Inc.) and
anti-Ki-67 (1:100; cat. no. 9449S, Cell Signaling Technology, Inc.)
primary antibodies. Tissue sections (5-µm) were formalin-fixed in
10% formaldehyde at 20°C for 24 h, paraffin-embedded, dehydrated
and subjected to peroxidase blocking using an immunohistochemistry
kit (cat. no. GK500705; Dako; Agilent Technologies, Inc.) for 5 min
at room temperature according to the manufacturer's protocol.
Primary antibodies were added and the sections were incubated at
room temperature for 30 min. The sections were incubated for
another 30 min at room temperature with the no. 2 solution of the
EnVision Detection kit (cat. no. GK500705; Dako; Agilent
Technologies, Inc.). Subsequently, the sections were incubated in
prepared 3,3′-diaminobenzidine from the kit at room temperature for
10 min. The slides were then counterstained with hematoxylin (cat.
no. S3309; Dako; Agilent Technologies, Inc.) for 30 sec at room
temperature. The stained slides were observed under a light
microscope (Leica DMI3000 M; magnification, ×40) and images were
captured. The percentage of Ki-67-positive cells was calculated
using ImageJ software (version 1.50i; National Institutes of
Health).
The Cancer Genome Atlas (TGCA)
database
TCGA (https://cancergenome.nih.gov/) database was used for
the purposes of this study. The dataset was analyzed using UALCAN
(www.ualcan.path.uab.edu).
Cell viability assessment
BXPC3 and SW1990 cells were seeded in triplicate
into 96-well plates at a density of 1×103 cells/well.
The cells were subsequently treated with DMSO, MLN4924, cisplatin
or MLN4924 + cisplatin at the indicated doses (0.3 µM MLN; 0.5 µM
cisplatin) for 48 h. Cell viability was assessed using the Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.) and
the ATPlite™ kit (PerkinElmer, Inc.) according to the
manufacturer's instructions.
Clonogenic assay
BXPC3 and SW1990 cells were trypsinized and seeded
in triplicate in 6-well plates (300 cells/well). Following
overnight culture at 37°C (5% CO2), the cells were
incubated with DMSO (dilution, 1:1,000), MLN4924, cisplatin or
MLN4924 + cisplatin (0.3 µM MLN; 0.5 µM cisplatin) for 12 days.
Colonies were fixed with 4% paraformaldehyde at room temperature
for 30 min and stained with 0.4% crystal violet solution in
methanol for 30 min at room temperature, images were captured using
a camera, and those with >30 cells were counted by three
independent investigators. Representative results of 3 independent
experiments are presented.
RT-qPCR analysis
Total RNA was isolated from cells and tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions, and all samples
were then treated with RNase-free DNase (Ambion; Thermo Fisher
Scientific, Inc.). The RT reaction was performed using 2 µg total
RNA/sample with the PrimeScript RT Reagent kit (Takara
Biotechnology Co., Ltd.) according to the manufacturer's protocol.
PCR amplification was performed using the Power SYBR®
Green PCR Master Mix (Applied Biosystems; Thermo Fisher Scientific,
Inc.) and the ABI 7900 thermocycler (Applied Biosystems; Thermo
Fisher Scientific, Inc.) following the manufacturer's protocol. The
thermocycling conditions were as follows: 95°C for 2 min; followed
by 40 cycles of 95°C for 10 sec, 60°C for 30 sec and 72°C for 30
sec; dissociation curve (95°C for 15 sec, 60°C for 15 sec and 95°C
for 15 sec). The primer sequences were as follows: Human NAE1,
forward, 5′-GCTGTTGTCATACTTCTC-3′, and reverse,
5′-TCGTGGACATAATCATCT-3′; human ubiquitin like modifier activating
enzyme 3 (UBA3), forward, 5′-TGGTGTTGGTGCTTGTAA-3′, and reverse,
5′-GTTGTATGCTTCCTCCTCTAA-3′; NEDD8-conjugating enzyme Ubc12
(UBC12), forward, 5′-CAGAAGAAGGAGGAGGAGTC-3′, and reverse,
5′-GAAGTTGAGGAGGTCGTCTG-3′; β-actin, forward,
5′-CATGTACGTTGCTATCCAGGC-3′, and reverse,
5′-CTCCTTAATGTCACGCACGAT-3′. The data were quantified using the
2−ΔΔCq method (17) and
the results are presented as relative expression levels.
Western blot analysis
Cell and tissue lysates were prepared using RIPA
buffer (Beyotime Institute of Biotechnology), and protein
concentrations were determined using an enhanced bicinchoninic acid
protein assay kit (Beyotime Institute of Biotechnology). Protein
samples (50 µg/lane) were separated using SDS-PAGE on a 12% gel,
then proteins were transferred to PVDF membranes, blocked with 5%
BSA (cat. no. ST023; Beyotime Institute of Biotechnology) in TBS
with Tween-20 (dilution, 1:1,000) for 2 h at room temperature, and
probed using primary antibodies against histone γ-H2AX (1:1,000;
cat. no. 9718; Cell Signaling Technology, Inc.), chromatin
licensing and DNA replication factor 1 (CDT1; 1:1,000; cat. no.
8064S; Cell Signaling Technology, Inc.), origin recognition complex
subunit 1 (ORC1; 1:1,000; cat. no. 4731S; Cell Signaling
Technology, Inc.), phosphorylated (p)-IκBα (S32; 1:1,000; cat. no.
D155066-0100; Sangon Biotech Co., Ltd.), NAE1 (1:1,000; cat. no.
ab187142; Abcam), UBA3 (1:1,000; cat. no. ab124728; Abcam), UBC12
(1:1,000; cat. no. ab109507; Abcam), p21 (1:1,000; cat. no.
ab109520; Abcam), p27 (1:1,000; cat. no. ab32034; Abcam), GAPDH
(1:2,000; cat. no. ab181602, Abcam), p-checkpoint kinase (CHK)1
(1:1,000; cat. no. ab58567; Abcam), p-CHK2 (1:1,000; cat. no.
ab32148; Abcam), c-poly (ADP-ribose) polymerase 1 (PARP; 1:1,000;
cat. no. ab32561; Abcam), c-caspase-3 (1:1,000; cat. no. 9664; Cell
Signaling Technology, Inc.), CHK1 (1:1,000; cat. no. ab40866;
Abcam), CHK2 (1:1,000; cat. no. ab47433; Abcam), H2AX (1:1,000;
cat. no. ab11175; Abcam), IκBα (1:1,000; cat. no. ab32518; Abcam)
and cullin 1 (1:1,000; cat. no. sc-135874; Santa Cruz
Biotechnology, Inc.) at 4°C overnight. Subsequently, membranes were
incubated with horseradish peroxidase-conjugated secondary
antibodies (1:1,000; cat. nos. 7077, 7074 and 7076; Cell Signaling
Technology, Inc.) at room temperature for 2 h. All membranes were
imaged using the LAS400 system (GE Healthcare Life Sciences) and an
enhanced chemiluminescent kit (cat. no. 32106; Thermo Fisher
Scientific, Inc.). The results were quantified using ImageJ
software (version 1.50i; National Institutes of Health) with GAPDH
as the loading control. All the bands presented together possessed
the same loading control.
Assessment of the antitumor effects of
MLN4924 and cisplatin in vivo
Male athymic nude mice (n=32; weight, 17–19 g; age,
5 weeks) were purchased from the Shanghai Experimental Animal
Center. The housing conditions were as follows: Temperature,
18–29°C; daily temperature difference, ≤3°C; relative humidity,
40–70%; fresh air ventilation, 10 times/h; airflow speed, ≤0.18
m/sec; pressure difference, 25 Pa; cleanliness level, 10,000;
ammonia concentration l5 mg/m3; noise, ≤60 dB;
illumination, 150–300 Lux; light/dark cycle, 12 h/12 h; and free
access to food and water. BXPC3 cells were trypsinized, resuspended
in PBS and subcutaneously injected into the groins of the mice
(5×106 cells/injection). At 1 week post-injection, the
tumor-bearing mice were randomly divided into 4 groups (n=8
mice/group). When the tumors had reached an average size of 250
mm3, mice were treated by subcutaneous injection with
15% HPBCD, MLN4924 (20 mg/kg), cisplatin (2.5 mg/kg) or MLN4924 +
cisplatin twice a week for 4 weeks (the largest tumor volume was
1,854 mm3, the longest tumor length was 15.94 mm). Tumor
size was measured weekly using vernier calipers and tumor volume
was calculated using the following formula: Volume=π/6 (length ×
width × height) (18). At the end of
the study, the mice were sacrificed using CO2 followed
by cervical dislocation (the flow rate of CO2 was 20% of
the volume of the chamber/min), and tumor tissues and blood were
collected. Tumor tissues were fixed with 10% formaldehyde at 20°C
for 24 h. Paraffin-embedded tissues were sectioned (5-µm thick) for
immunohistochemical analysis of c-caspase-3 and Ki-67. Blood
samples were extracted from the tail vein and centrifuged at 500 ×
g for 10 min at room temperature. Serum samples were collected for
the biochemical assays of aspartate aminotransferase (AST) and
alanine aminotransferase (ALT) levels using ELISA kits (cat. nos.
MBS016898 and MBS775731; MyBioSource, Inc.) according to the
manufacturer's protocols and the Modular DDP Analyzer (Roche
Applied Science) according to the manufacturer's instructions.
Ethical approval was obtained from the Ethics Committee of
Yongchuan Hospital of Chongqing Medical University, and all
procedures were performed in accordance with the National
Institutes of Health Guide for the Care and Use of Laboratory
Animals.
Statistical analysis
The results from three independent experiments are
presented as the mean ± standard deviation. Paired Student's
t-tests were used for the comparison of parameters between two
groups, and multiple comparisons were performed using one-way
analysis of variance followed by the Student-Newman-Keuls post hoc
test. SPSS software (version 17.0; SPSS, Inc.) was used for
statistical analyses.
Results
Neddylation is overactivated in
pancreatic cancer, and high NAE1 expression is an indicator of poor
patient prognosis
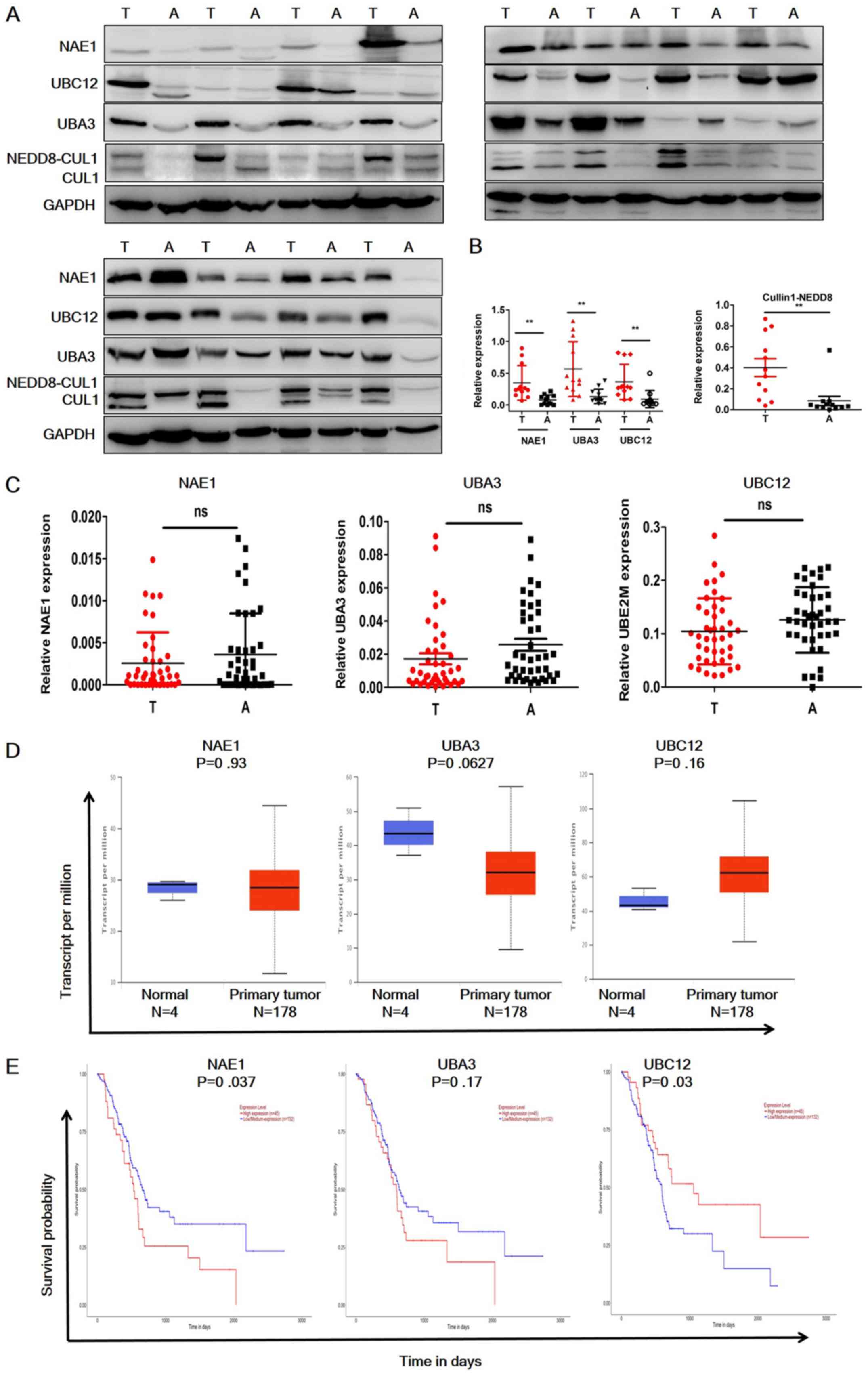
To evaluate the status of the neddylation pathway in
pancreatic cancer, the protein expression levels of NAE1, UBA3
(subunits of E1), UBE12 (E2) and cullin 1 in 12 paired-pancreatic
cancer and adjacent normal tissues were analyzed (Fig. 1A and B). NAE1, UBA3 and UBE12 were
significantly overexpressed in pancreatic cancer samples, and
neddylated cullin 1 expression was significantly higher in tumor
tissues compared with the adjacent normal tissues. RT-qPCR analysis
was then performed to investigate the mRNA expression levels of
these molecules (Fig. 1C). In
contrast to the protein expression levels, the RT-qPCR results
demonstrated that NAE1, UBA3 and UBE12 mRNA expression levels were
not significantly elevated in tumor tissues. These data were
further confirmed by consulting TCGA datasets (Fig. 1D). UBC12 expression appeared to be
elevated in the TCGA dataset; however, this was not identified as
statistically significant. Notably, the TCGA datasets indicated
that NAE1 overexpression was a predictor of poor prognosis in
patients with pancreatic cancer, whereas high expression levels of
UBA3 were not a predictor of poor prognosis (Fig. 1E), and low UBC12 expression predicted
a poor prognosis (Fig. 1E). These
data indicated that neddylation pathway members are only
overexpressed at the protein level in pancreatic cancer, and high
NAE1 and low UBC12 expression may be indicators of poor patient
prognosis.
MLN4924 enhances the therapeutic
effect of cisplatin in pancreatic cancer cells in vitro
Overactivation of the neddylation pathway in
pancreatic cancer indicates that it may present a potential
therapeutic target. To evaluate the effects of combined MLN4924 and
cisplatin treatment in pancreatic cancer cells, inhibition of
neddylation using MLN4924 was first demonstrated (Fig. 2A). To investigate the effect of
combined MLN4924 and cisplatin treatment on the proliferation of
pancreatic cancer cells, BXPC3 and SW1990 cells were then treated
with MLN4924, cisplatin or MLN4934 + cisplatin at the indicated
doses (0.3 µM MLN; 0.5 µM cisplatin) for 48 h, and analyzed using
ATPlite and CCK-8 cell viability assays. As presented in Fig. 2B, MLN4924 or cisplatin treatment
alone significantly suppressed the proliferation of both cell
lines, and the combined administration of these agents
significantly further suppressed proliferation compared with
monotherapy. Cell clonogenic survival analysis revealed comparable
results (Fig. 2C and D), indicating
that MLN4924 suppresses the proliferation of pancreatic cancer
cells in vitro, and that combined MLN4924 + cisplatin
treatment demonstrates a greater effect than treatment with each
agent alone.
 | Figure 2.MLN4924 + cisplatin treatment
suppresses pancreatic cancer proliferation in vitro. (A)
MLN4924 inhibited the neddylation of CUL1 in pancreatic cancer
cells. Cells were treated with MLN4924 for 24 h and analyzed by
western blotting. (B) Effects of MLN4924 + cisplatin treatment on
the viability of BXPC3 and SW1990 cells in vitro. Cells were
treated with DMSO, MLN4924, cisplatin or MLN4924 + cisplatin for 48
h (0.3 µM MLN; 0.5 µM cisplatin) and viability was assessed using
ATPlite and Cell Counting Kit-8 assays, respectively. (C and D)
Effects of MLN4924 + cisplatin treatment on clonogenic survival.
BXPC3 and SW1990 cells were treated with DMSO, MLN4924, cisplatin
or MLN4924 + cisplatin for 12 days (0.3 µM MLN; 0.5 µM cisplatin).
All experiments were repeated 3 times and the results are presented
as the mean ± standard deviation. ***P<0.001. DMSO, dimethyl
sulfoxide; CON, control; MLN, MLN4924; CIS, cisplatin; NEDD8,
Neural precursor cell-expressed, developmentally downregulated 8;
CULl, cullin 1. |
MLN4924 enhances the therapeutic
effect of cisplatin in pancreatic cancer in vivo
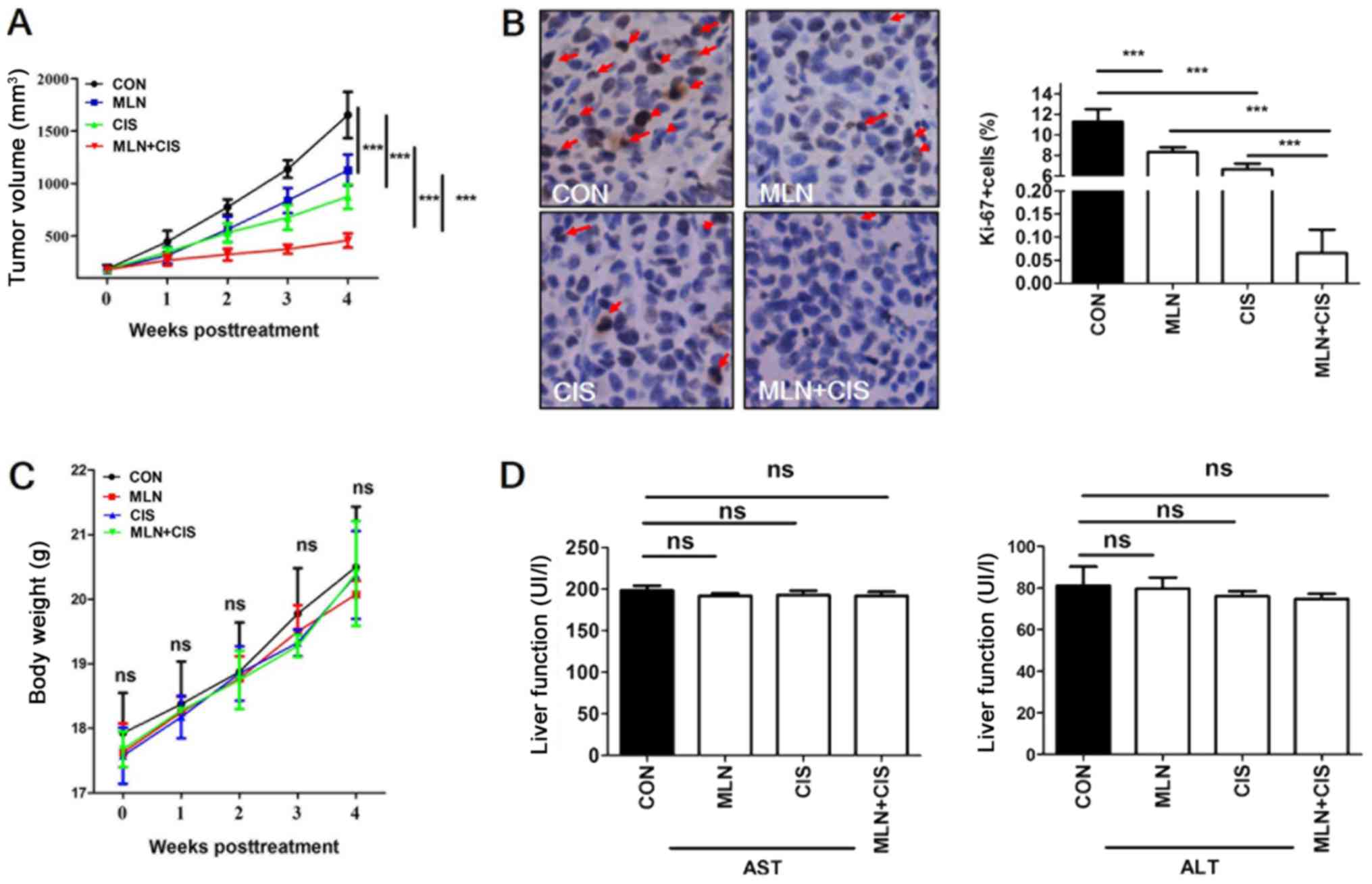
To evaluate the antitumor effects of MLN4924 +
cisplatin treatment in vivo, a mouse BXPC3 ×enograft tumor
model was established and mice were treated with MLN4924, cisplatin
or MLN4924 + cisplatin. Compared with monotherapy, MLN4924 +
cisplatin significantly inhibited tumor growth (Fig. 3A). In addition, positive Ki-67
staining was significantly decreased in the MLN4924 + cisplatin
group compared with the monotherapy and control groups (Fig. 3B). These results indicate that the
proliferation of pancreatic tumor cells in the MLN4924 +
cisplatin-treated group was significantly suppressed. During the
treatment period, no obvious side effects, such as weight loss or
reduced liver function were observed (Fig. 3C and D). These results demonstrated
that MLN4924 + cisplatin suppressed pancreatic tumor growth in
vivo.
MLN4924 enhances the tumor-suppressive
effects of cisplatin by inducing DNA damage and apoptosis
MLN4924 is a promising anticancer agent as it
induces the accumulation of CRL substrates that promote DNA damage,
cell apoptosis and autophagy (6).
Cisplatin also demonstrates anticancer effects via the induction of
DNA damage. It was therefore hypothesized that the enhanced
anticancer effects of MLN4924 combined with cisplatin may be due to
increased levels of DNA damage resulting from the accumulation of
CRL substrates. To test this hypothesis, pancreatic cancer cells
were treated with MLN4924, cisplatin or MLN4924 + cisplatin. The
DNA damage markers p-CHK 1, p-CHK2 and γ-H2AX were subsequently
analyzed. As indicated in Fig. 4A and
C, p-CHK1, p-CHK2 and γ-H2AX expression levels were markedly
increased in the MLN4924 + cisplatin-treated group compared with
the monotherapy and control groups. These results indicated that
MLN4924 + cisplatin treatment enhanced DNA damage in pancreatic
cancer cells.
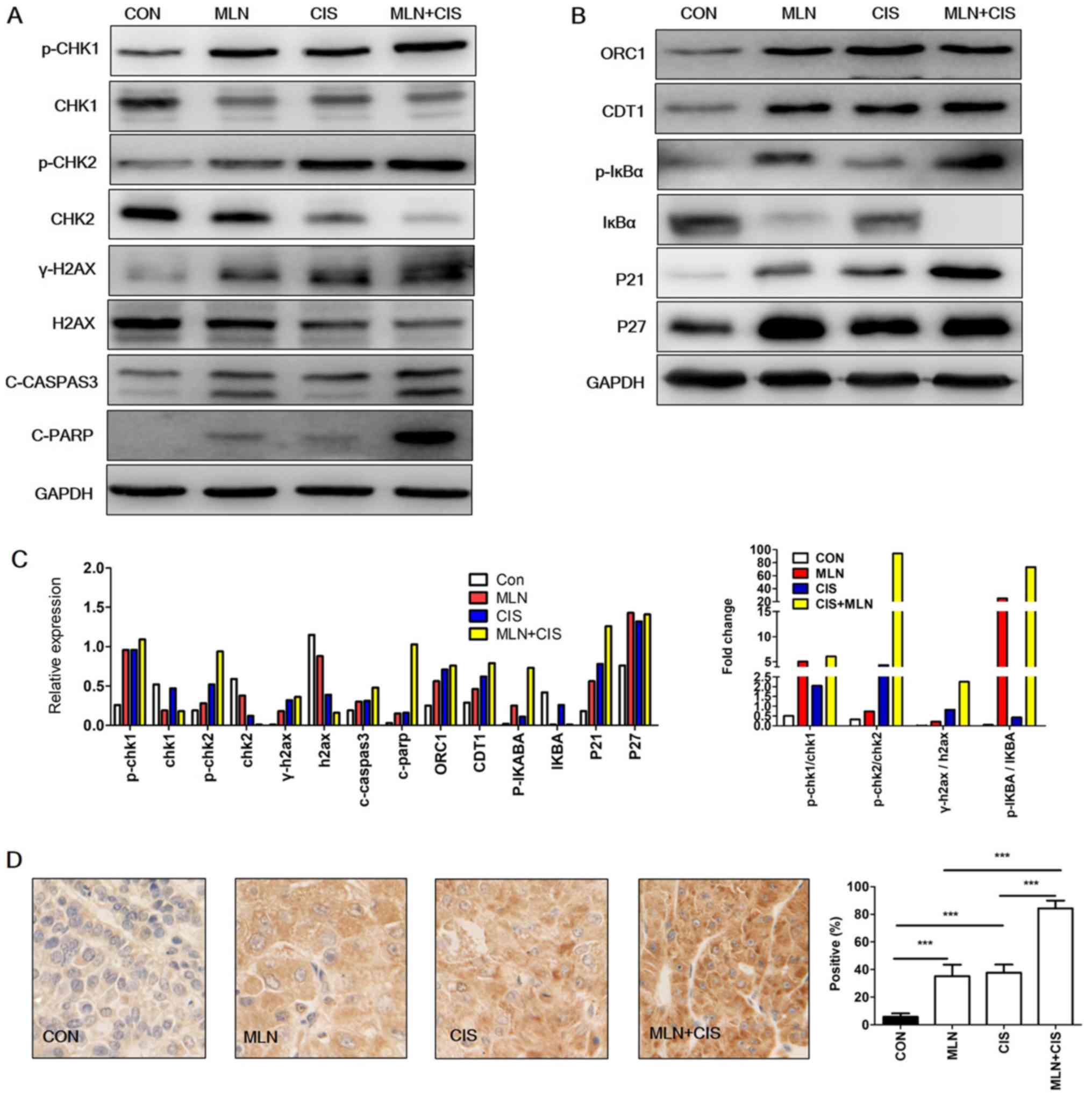 | Figure 4.Enhanced DNA damage and accumulation
of CRL substrates results in cell apoptosis. (A and B) Western blot
analysis of BXPC3 cells treated with MLN4924 + cisplatin for 24 h.
(C) Quantification of western blotting and the ratio of p-/total
protein. Each column represents a single quantification. (D)
Representative immunohistochemical staining images of C-caspase-3
expression in xenograft tumors (magnification, ×400). The results
are presented as the mean ± standard deviation. ***P<0.001.
C-caspase-3, cleaved caspase-3; CRL, cullin ring ligase; CON,
control; MLN, MLN4924; CIS, cisplatin; CHK, checkpoint kinase;
γ-H2AX, phospho-histone H2AX; C-PARP, cleaved-poly (ADP-ribose)
polymerase 1; ORC1, origin recognition complex subunit 1; p-,
phosphorylated; CDT1, chromatin licensing and DNA replication
factor 1. |
In order to investigate the mechanisms underlying
increased DNA damage following MLN4924 + cisplatin treatment, the
levels of CRL substrates (known to induce DNA damage) were
determined. As presented in Fig. 4B and
C, accumulation of the DNA replication license proteins CDT1
and ORC1 was observed in the MLN4924 + cisplatin group compared
with in the control group. Similarly, a marked increase in the CRL
substrates p27, p-IκBα and p21 was observed in the MLN4924 +
cisplatin treatment group. These results indicated that the
enhanced DNA damage caused by MLN4924 + cisplatin treatment may be
due to the increased accumulation of CRL substrates.
It has been reported that DNA damage induces
apoptosis (19). Given that MLN4924
+ cisplatin enhanced DNA damage in pancreatic cancer cells in the
present study, it was hypothesized that this treatment may also
lead to increased apoptosis. As shown in Fig. 4A and C, an increase in the expression
level of apoptosis markers c-caspase-3 and C-PARP was observed,
compared with the monotherapy and the control groups. Similarly,
immunohistochemical staining analysis of c-caspase-3 expression in
mouse xenograft tumors revealed that the levels of this
apoptosis-associated protein were also significantly increased
(Fig. 4D). These results indicate
that MLN4924 + cisplatin enhanced DNA damage and induced the
expression of apoptosis-associated proteins.
Discussion
Pancreatic cancer is a human malignancy associated
with high mortality rates. Despite recommended treatment with
gemcitabine in the clinic, the 5-year survival rates for patients
remain low worldwide (1).
Overactivation of the neddylation pathway has been observed in
specific malignancies and indicates poor prognosis (6,8,12,13).
However, to the best of our knowledge, its mRNA expression in
pancreatic cancer had not been previously studied. A previous study
demonstrated that neddylation pathway-associated molecules, such as
NAE1, UBA3 and UBC12 were overexpressed at the protein level in
pancreatic cancer tissues when compared with adjacent normal
tissues (8). In addition, the levels
of neddylated cullin 1, a known neddylation substrate (8) were observed to be markedly increased in
pancreatic cancer tissue samples when compared with adjacent normal
tissues. However, the results of the present study demonstrated
that no differences in the mRNA expression levels of these
molecules were observed in pancreatic cancer tissues compared with
adjacent normal tissues. UBC12 appeared to be elevated in the TCGA
dataset; however, this was not statistically significant. In
addition, by consulting TCGA database, only NAE1 overexpression was
demonstrated to be a predictor of poor patient prognosis in
pancreatic cancer. This contradicts previous studies involving lung
cancer and cholangiocarcinoma (8,12), but
is consistent with the results presented by Li et al
(16). However, low UBC12 expression
predicted a poor prognosis, which was inconsistent with a previous
study (8). UBC12 may not only
function as an E2 of the neddylation process. The potential
mechanism requires further investigation in the future.
Neddylation inhibitors have demonstrated promising
anticancer effects in certain malignancies by inducing DNA damage,
apoptosis, cell cycle arrest, cell senescence and autophagy
(11,20–23).
Combined MLN4924 and gemcitabine treatment has been demonstrated to
significantly suppress pancreatic cancer growth by inducing
apoptosis, via the elevation of NADPH oxidase activator 1
expression in vitro and in vivo (16). In addition, combined MLN4924 and
radiotherapy treatment was observed to significantly inhibit
pancreatic cancer growth by promoting DNA damage (15). As MLN4924 and cisplatin are both
known to induce DNA damage and are used in clinical practice for
the treatment of specific malignancies, the effects of combined
MLN4924 and cisplatin treatment on pancreatic cancer cells were
investigated in the present study. This hypothesis was verified in
esophageal squamous cell carcinoma (24). The results demonstrated that MLN4924
+ cisplatin treatment significantly suppressed pancreatic cancer
growth both in vitro and in vivo, and that combined
treatment was well tolerated by the mice.
As MLN4924 + cisplatin therapy induced significant
effects on the proliferation of pancreatic cancer cells,
mechanistic studies were subsequently performed. The results
demonstrated that MLN4924 + cisplatin-mediated suppression of cell
proliferation may be related to enhanced DNA damage and increased
levels of apoptosis-associated proteins. This was consistent with a
previous study involving combined MLN4924 and radiotherapy
treatment (15). Additionally,
inhibition of CHK1 (which is involved in the DNA damage response)
using a specific inhibitor (SCH 900776) was also observed to
increase the anticancer activity of MLN4924 in pancreatic cancer
cells (25). MLN4924 induces DNA
damage by promoting the accumulation of the DNA replication license
proteins CDT1 and ORC1, as well as additional CRL substrates
(6). Consistent with these
observations, a significant accumulation of CDT1, ORC1, p27, p-IκBα
and p21 following MLN4924 + cisplatin treatment was observed in
pancreatic cancer cells.
The constitutive photomorphogenesis 9 (COP9)
signalosome (CSN) complex was revealed to be responsible for the
de-neddylation of Cul family proteins (26). CSN complex subunit 5 is a key
component of the CSN complex (28),
which was found to be overexpressed in a number of cancer types,
including pancreatic cancer (27,28). As
neddylation and de-neddylation enzymes are both overexpressed in
cancer, it was speculated that the neddylation dynamics were
tightly regulated. Hua et al (13) demonstrated that the neddylation
pathway was overactivated in glioblastoma and that MLN4924
suppressed GBM cell (U251 and A172) growth significantly; however,
neddylated cul1 is expressed to a greater degree in U251 cells than
A172 cells. In the present study, the levels of neddylated Cul1
varied between BXPC3 and SW1990 cells; however, this did not alter
the effects of MLN4924. These results were consistent with another
glioblastoma study (13).
In conclusion, the results of the present study
indicated that the neddylation pathway is overactivated in
pancreatic cancer, and that the expression levels of key molecules
within this pathway are upregulated. In addition, MLN4924 +
cisplatin treatment significantly suppressed pancreatic cancer cell
growth in vitro and in vivo. MLN4924 + cisplatin
treatment may suppress pancreatic cancer cell proliferation by
promoting DNA damage and increasing the expression of
apoptosis-associated proteins via enhanced CDT1, ORC1, p27, p-IκBα
and p21 accumulation.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Nature Science
Foundation of Chongqing Yongchuan Science and Technology Commission
(grant no. Ycstc-2017nc5004).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YZ, YSI and HL contributed to the study design,
analysis and interpretation of the data. HL conceived the study.
YZ, YSI, QHP, and YGZ performed the experiments, and YZ, YSI and
YGZ performed the statistical analyses. HL drafted the manuscript
and supervised the study. All of the authors approved the final
manuscript.
Ethics approval and consent to
participate
Ethical approval was obtained from the Research
Ethics Committee of Chongqing Medical University, and written
informed consent regarding tissue collection and the use of patient
data for scientific purposes was obtained from all recruited
patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kamisawa T, Wood LD, Itoi T and Takaori K:
Pancreatic cancer. Lancet. 388:73–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rabut G and Peter M: Function and
regulation of protein neddylation. ‘Protein modifications: Beyond
the usual suspects’ review series. EMBO Rep. 9:969–976. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Enchev RI, Schulman BA and Peter M:
Protein neddylation: Beyond cullin-RING ligases. Nat Rev Mol Cell
Biol. 16:30–44. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao Y, Morgan MA and Sun Y: Targeting
neddylation pathways to inactivate cullin-RING ligases for
anticancer therapy. Antioxid Redox Signal. 21:2383–2400. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu J, Li L, Yu G, Ying W, Gao Q, Zhang W,
Li X, Ding C, Jiang Y, Wei D, et al: The neddylation-cullin 2-RBX1
E3 ligase axis targets tumor suppressor RhoB for degradation in
liver cancer. Mol Cell Proteomics. 14:499–509. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li L, Wang M, Yu G, Chen P, Li H, Wei D,
Zhu J, Xie L, Jia H, Shi J, et al: Overactivated neddylation
pathway as a therapeutic target in lung cancer. J Natl Cancer Inst.
106:dju0832014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xie P, Zhang M, He S, Lu K, Chen Y, Xing
G, Lu Y, Liu P, Li Y, Wang S, et al: The covalent modifier Nedd8 is
critical for the activation of Smurf1 ubiquitin ligase in
tumorigenesis. Nat Commun. 5:37332014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Soucy TA, Smith PG, Milhollen MA, Berger
AJ, Gavin JM, Adhikari S, Brownell JE, Burke KE, Cardin DP,
Critchley S, et al: An inhibitor of NEDD8-activating enzyme as a
new approach to treat cancer. Nature. 458:732–736. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo Z, Yu G, Lee HW, Li L, Wang L, Yang D,
Pan Y, Ding C, Qian J, Wu L, et al: The Nedd8-activating enzyme
inhibitor MLN4924 induces autophagy and apoptosis to suppress liver
cancer cell growth. Cancer Res. 72:3360–3371. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao Q, Yu GY, Shi JY, Li LH, Zhang WJ,
Wang ZC, Yang LX, Duan M, Zhao H, Wang XY, et al: Neddylation
pathway is up-regulated in human intrahepatic cholangiocarcinoma
and serves as a potential therapeutic target. Oncotarget.
5:7820–7832. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hua W, Li C, Yang Z, Li L, Jiang Y, Yu G,
Zhu W, Liu Z, Duan S, Chu Y, et al: Suppression of glioblastoma by
targeting the overactivated protein neddylation pathway. Neuro
Oncol. 17:1333–1343. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen P, Hu T, Liang Y, Jiang Y, Pan Y, Li
C, Zhang P, Wei D, Li P, Jeong LS, et al: Synergistic inhibition of
autophagy and neddylation pathways as a novel therapeutic approach
for targeting liver cancer. Oncotarget. 6:9002–9017.
2015.PubMed/NCBI
|
|
15
|
Wei D, Li H, Yu J, Sebolt JT, Zhao L,
Lawrence TS, Smith PG, Morgan MA and Sun Y: Radiosensitization of
human pancreatic cancer cells by MLN4924, an investigational
NEDD8-activating enzyme inhibitor. Cancer Res. 72:282–293. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li H, Zhou W, Li L, Wu J, Liu X, Zhao L,
Jia L and Sun Y: Inhibition of neddylation modification sensitizes
pancreatic cancer cells to gemcitabine. Neoplasia. 19:509–518.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feldman JP, Goldwasser R, Mark S, Schwartz
J and Orion I: A mathematical model for tumor volume evaluation
using two-dimensions. J Appl Quant Methods. 4:455–462. 2009.
|
|
19
|
Roos WP and Kaina B: DNA damage-induced
cell death by apoptosis. Trends Mol Med. 12:440–450. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Milhollen MA, Traore T, Adams-Duffy J,
Thomas MP, Berger AJ, Dang L, Dick LR, Garnsey JJ, Koenig E,
Langston SP, et al: MLN4924, a NEDD8-activating enzyme inhibitor,
is active in diffuse large B-cell lymphoma models: Rationale for
treatment of NF-{kappa}B-dependent lymphoma. Blood. 116:1515–1523.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin JJ, Milhollen MA, Smith PG, Narayanan
U and Dutta A: NEDD8-targeting drug MLN4924 elicits DNA
rereplication by stabilizing Cdt1 in S phase, triggering checkpoint
activation, apoptosis, and senescence in cancer cells. Cancer Res.
70:10310–10320. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao Y, Xiong X, Jia L and Sun Y:
Targeting Cullin-RING ligases by MLN4924 induces autophagy via
modulating the HIF1-REDD1-TSC1-mTORC1-DEPTOR axis. Cell Death Dis.
3:e3862012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Blank JL, Liu XJ, Cosmopoulos K, Bouck DC,
Garcia K, Bernard H, Tayber O, Hather G, Liu R, Narayanan U, et al:
Novel DNA damage checkpoints mediating cell death induced by the
NEDD8-activating enzyme inhibitor MLN4924. Cancer Res. 73:225–234.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin S, Shang Z, Li S, Gao P, Zhang Y, Hou
S, Qin P, Dong Z, Hu T and Chen P: Neddylation inhibitor MLN4924
induces G2 cell cycle arrest, DNA damage and sensitizes
esophageal squamous cell carcinoma cells to cisplatin. Oncol Lett.
15:2583–2589. 2018.PubMed/NCBI
|
|
25
|
Li JA, Song C, Rong Y, Kuang T, Wang D, Xu
X, Yuan J, Luo K, Qin B, Nowsheen S, et al: Chk1 inhibitor SCH
900776 enhances the antitumor activity of MLN4924 on pancreatic
cancer. Cell Cycle. 17:191–199. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cope GA and Deshaies RJ: COP9 signalosome:
A multifunctional regulator of SCF and other cullin-based ubiquitin
ligases. Cell. 114:663–671. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fukumoto A, Ikeda N, Sho M, Tomoda K,
Kanehiro H, Hisanaga M, Tsurui Y, Tsutsumi M, Kato JY and Nakajima
Y: Prognostic significance of localized p27Kip1 and potential role
of Jab1/CSN5 in pancreatic cancer. Oncol Rep. 11:277–284.
2004.PubMed/NCBI
|
|
28
|
Pan Y and Claret FX: Targeting Jab1/CSN5
in nasopharyngeal carcinoma. Cancer Lett. 326:155–160. 2012.
View Article : Google Scholar : PubMed/NCBI
|