Introduction
With the increase of aging population, the
acceleration of industrialization, and the intensification of
environmental pollution, the incidence and mortality of lung cancer
has increased rapidly (1). Lung
cancer has become one of the most rapidly increasing cancers in
China, and will be the leading cause of death for a long time
(2). Therefore, the prevention and
early detection of lung cancer is expected to become one of the key
points of tumor control in China. Moreover, non-small cell lung
cancer (NSCLC) accounts for approximately 84% of all lung cancers,
which can be transferred to the whole body through lymphoids and
blood. NSCLC is metastatic but not infectious (3). It should be emphasized that the
clinical staging before the treatment of NSCLC is crucial. Accurate
clinical staging is not only helpful for doctors to make scientific
and reasonable treatment program for patients with NSCLC, but also
can better help doctors to judge the prognosis of NSCLC and modify
the auxiliary treatment strategies after surgery (4). However, there is a great disparity in
clinical diagnosis and treatment due to the non-standard diagnosis
and treatment of NSCLC.
Carcinogenesis of NSCLC involves in multistep
process which is regulated by activating oncogenes or inactivating
tumor-suppressing genes (5).
Recently, several microRNAs (miRNAs) have been reported to
participate in tumorigenesis of NSCLC which play a carcinogenic or
suppressive role in the progression of NSCLC. For example, miR-520a
inhibited cell growth and metastasis of NSCLC through PI3K/AKT/mTOR
signaling pathway (6). On the
contrary, miR-96 promoted invasion and metastasis by targeting GPC3
in NSCLC cells (7). The function of
microRNA-545 (miR-545)in human disease and cancers has drawn
increased attention. The downregulation of miR-545 has been
identified in pancreatic ductal adenocarcinoma which inhibited
tumor growth through targeting RIG1 (8). In addition, miR-545 can be used as an
early biomarker of Alzheimer's disease (9). Moreover, a previous study reported the
role of miR-545 in lung cancer. miR-545 arrested cell cycle and
induced cell apoptosis in lung cancer through regulating cyclin D1
and CDK4 (10). However, the effect
of miR-545 on the development of NSCLC is still unknown.
Zinc finger E-box-binding homeobox 2 (ZEB2) has been
found to participate in pathogenesis of human cancers (11). ZEB2 has been documented to be
correlated with metastasis and invasion in several types of
malignant tumors. Chua et al (12) demonstrated that ZEB2 repressed
E-cadherin expression and enhanced epithelial to mesenchymal
transition (EMT) of mammary epithelial cells. Furthermore, miR-132
suppressed the migration and invasion of lung cancer cells via
targeting the EMT regulator ZEB2 (13). However, the relationship of miR-545
with ZEB2 still needs to be investigated in NSCLC. ZEB2 has been
revealed to be involved in the Wnt/β-catenin pathway to regulate
the development of gastric adenocarcinoma (14). In lung cancer, miR-130b targeted PTEN
to induce resistance to cisplatin by activating Wnt/β-catenin
pathway (15). However, no
interaction between ZEB2 and Wnt/β-catenin pathway in NSCLC has
been proposed.
The main objective of this research was to observe
the aberrant expression, as well as the function of miR-545 in the
progression of NSCLC with the aim to provide a theoretical basis
for illuminating the regulatory mechanism of miR-545 in NSCLC.
Patients and methods
Clinical tissues
In total, 84 human NSCLC tissues and adjacent normal
lung tissues were acquired from the The Third People's Hospital of
Qingdao (Qingdao, China). The patients with NSCLC did not receive
any treatment before the operation. The tissues were frozen in
liquid nitrogen and then stored at −80°C in a refrigerator. All the
patients provided informed consent and this experiment was approved
by the Institutional Ethics Committee of The Third People's
Hospital of Qingdao.
Cell culture
H522 [cat. no. ZK156 (XR)], A549 [cat. no ZK0930
(XR)], H650 [cat. no.ZK1573(XR)] cell lines and human normal
bronchial epithelial cells (HBECs) (ZKCC-X2676) were used for this
experiment. The cell lines were acquired from Shanghai Maisha
Biotechnology. The cells were seeded in DMEM medium with 10% fetal
bovine serum (FBS) and cultured at 37°C with 5% CO2.
Cell transfection
The miR-545 mimics or inhibitor and negative control
(NC) were obtained from RiboBio. ZEB2 siRNA or negative control
siRNA was purchased from Genechem. They were transferred into A549
cells respectively by using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) based on the manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was applied to extract total RNA in NSCLC. The
synthesis of cDNA was performed using RevertAid First Strand cDNA
kit (cat. no. K1622; Thermo Fisher Scientific, Inc.). RT-qPCR was
conducted using SYBR-Green Reagent (cat. no. 4385612; Thermo Fisher
Scientific, Inc.) on 7500 Fast Real-Time PCR system (Thermo Fisher
Scientific, Inc.). The thermocycling parameters were as follows:
95°C for 3 min and 38 cycles of 95°C for 15 sec followed by 60°C
for 30 sec The temperature conditions of the reverse transcription
were as follows: 37°C for 15 min and 85°C for 5 sec. U6 or GAPDH
was used as internal control for miR-545 or ZEB2. Their expression
levels were calculated using the 2−∆∆Cq method (16).
Western blot analysis
The protein samples were obtained using RIPA lysis
buffer. Protein determination method was calculated using
bicinchoninic acid (BCA). Then equal amounts of proteins (30 µg)
were separated through a 10% SDS-PAGE and transferred into a
polyvinylidene difluoride (PVDF) membrane (Thermo Fisher
Scientific, Inc.). Then, the membranes were blocked with 5% non-fat
milk for 1 h at room temperature. Next the membranes were incubated
overnight at 4°C with EMT markers [(E-cadherin (rabbit monoclonal
antibody; dilution, 1:1,000; cat. no. ab1416; Abcam), N-cadherin
(rabbit polyclonal antibody; dilution, 1:1,000; cat. no. 18203;
Abcam), vimentin (rabbit polyclonal antibody; dilution, 1:1,000;
cat. no. 137321; Abcam), Wnt/β-catenin pathway markers (β-catenin),
rabbit polyclonal antibody; dilution, 1:1,000; cat. no. 6302;
Abcam), ZEB2 (rabbit polyclonal antibody; dilution, 1:1,000; cat.
no. 138222; Abcam)] and GAPDH primary, rabbit monoclonal antibodies
(dilution, 1:1,000; cat. no. ab181602; Abcam). After washing, they
were incubated with the corresponding secondary mouse anti-rabbit
(dilution, 1:2,000; cat. no. 3678; Abcam) or goat anti-mouse
(dilution, 1:2,000; cat. no. 58802; Abcam) antibodies conjugated
with horseradish peroxidase (Cell Signaling Technology, Inc.) for 2
h at room temperature. Then, protein expression levels were
measured by ECL (Pierce; Thermo Fisher Scientific, Inc.).
MTT assay
MTT
(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)
assay was used to assess cell viability. The 5×104 A549
cells with transfection were incubated in a 96-well plate at 37°C
for 48 h. Next, the cells were incubated at 37°C in a serum-free
medium with 20 µl MTT for 4 h. Finally, they were dissolved in 100
µl of dimethyl sulfoxide after removing the supernatant and
absorbance was read at 490 nm on a microplate spectrophotometer
(Thermo Labsystems).
Transwell assay
Transwell chambers (8 µm pore size; Millipore) were
applied to evaluate the migratory and invasive ability of A549
cells in 24-well plates. A549 cells (3×104) without
serum were put in the upper chamber on the non-coated membrane, and
the lower chamber was filled with 10% FBS to induce A549 cells to
migrate or invade through the membrane. The cells were put in the
upper chamber with 100 µl Matrigel (dilution, 1:8 in DMEM medium)
for invasion assay. The cells on the lower surface of the membrane
were fixed with 4% paraformaldehyde, stained with 1% crystal violet
(Solarbio) for 5 min and subsequently counted in 3 randomly
selected fields under an inverted microscope (Leica Microsystems
GmbH) at magnification ×200.
Dual luciferase assay
The wild or mutant type of 3′-UTR of ZEB2 was
inserted into pcDNA3.1 plasmid vector (Promega Corporation) to
perform luciferase reporter experiments. Then, wild or mutant type
of 3′-UTR of ZEB2 and miR-545 mimic were transfected into A549
cells. Subsequently, the luciferase activity was measured through
dual luciferase assay system (Promega Corporation).
Statistical analysis
The data are shown as mean ± SD. SPSS 19.0 or
Graphpad Prism 6 (GraphPad Software, Inc.) was employed to analyze
these data. The difference between the groups was calculated
through one-way ANOVA followed by the Bonferroni post hoc test. The
correlation of miR-545 with clinicopathological characteristics of
NSCLC was calculated through the Chi-squared test. The overall
survival rates and survival differences were detected through
univariate Kaplan-Meier method followed by log-rank test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Expression of miR-545 is decreased in
NSCLC tissues
Alteration of miR-545 expression was examined in
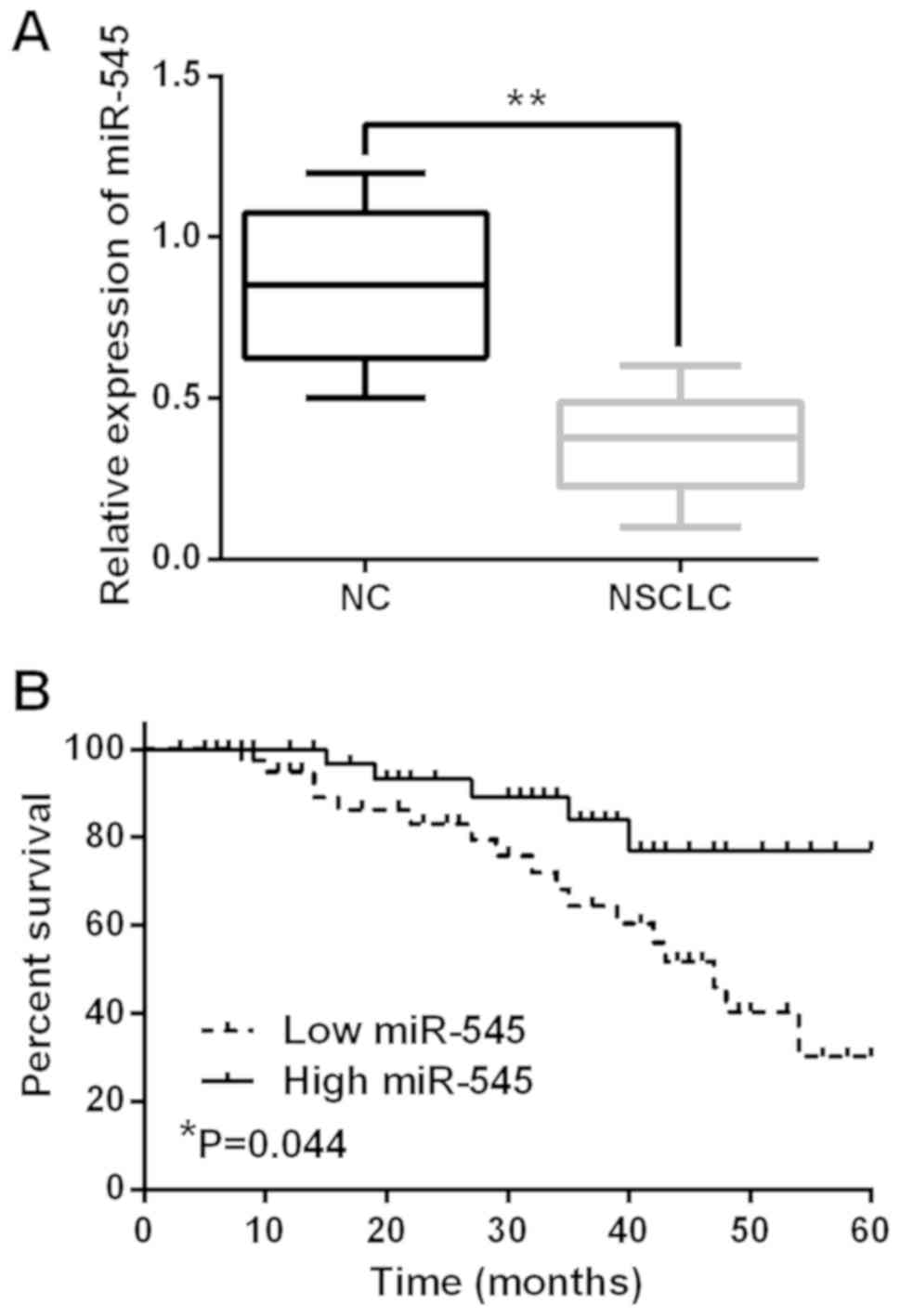
NSCLC tissues by RT-qPCR assay. miR-545 was found to be
downregulated in NSCLC tissues in comparison with the normal
tissues (P<0.01, Fig. 1A).
Moreover, the close association between the decreased expression of
miR-545 and lymph node metastasis (P=0.016) or tumor stage (P=0.02)
was identified in NSCLC patients (Table
I). Low miR-545 expression predicted worse prognosis and was
associated with shorter overall survival of NSCLC patients
(P=0.044, Fig. 1B). Thus, it was
implied that miR-545 was involved in the tumorigenesis and
prognosis of NSCLC.
 | Table I.Relationship of miR-545 expression and
their clinicopathological characteristics of NSCLC patients. |
Table I.
Relationship of miR-545 expression and
their clinicopathological characteristics of NSCLC patients.
|
|
| miR-545 |
|
|---|
|
|
|
|
|
|---|
| Characteristics | Cases | High | Low | P-value |
|---|
| Age (years) |
|
|
| 0.072 |
| ≥60 | 54 | 20 | 34 |
|
|
<60 | 30 | 12 | 18 |
|
| Sex |
|
|
| 0.21 |
|
Male | 50 | 21 | 29 |
|
|
Female | 34 | 16 | 18 |
|
| Tumor size
(mm) |
|
|
| 0.144 |
| ≤3 | 48 | 22 | 26 |
|
|
>3 | 36 | 15 | 21 |
|
| Lymph node
metastasis |
|
|
| 0.016a |
|
Yes | 19 | 7 | 12 |
|
| No | 65 | 25 | 40 |
|
| Tumor stage |
|
|
| 0.02a |
|
I–II | 60 | 24 | 36 |
|
|
III–IV | 24 | 10 | 14 |
|
Overexpression of miR-545 inhibits
progression of NSCLC
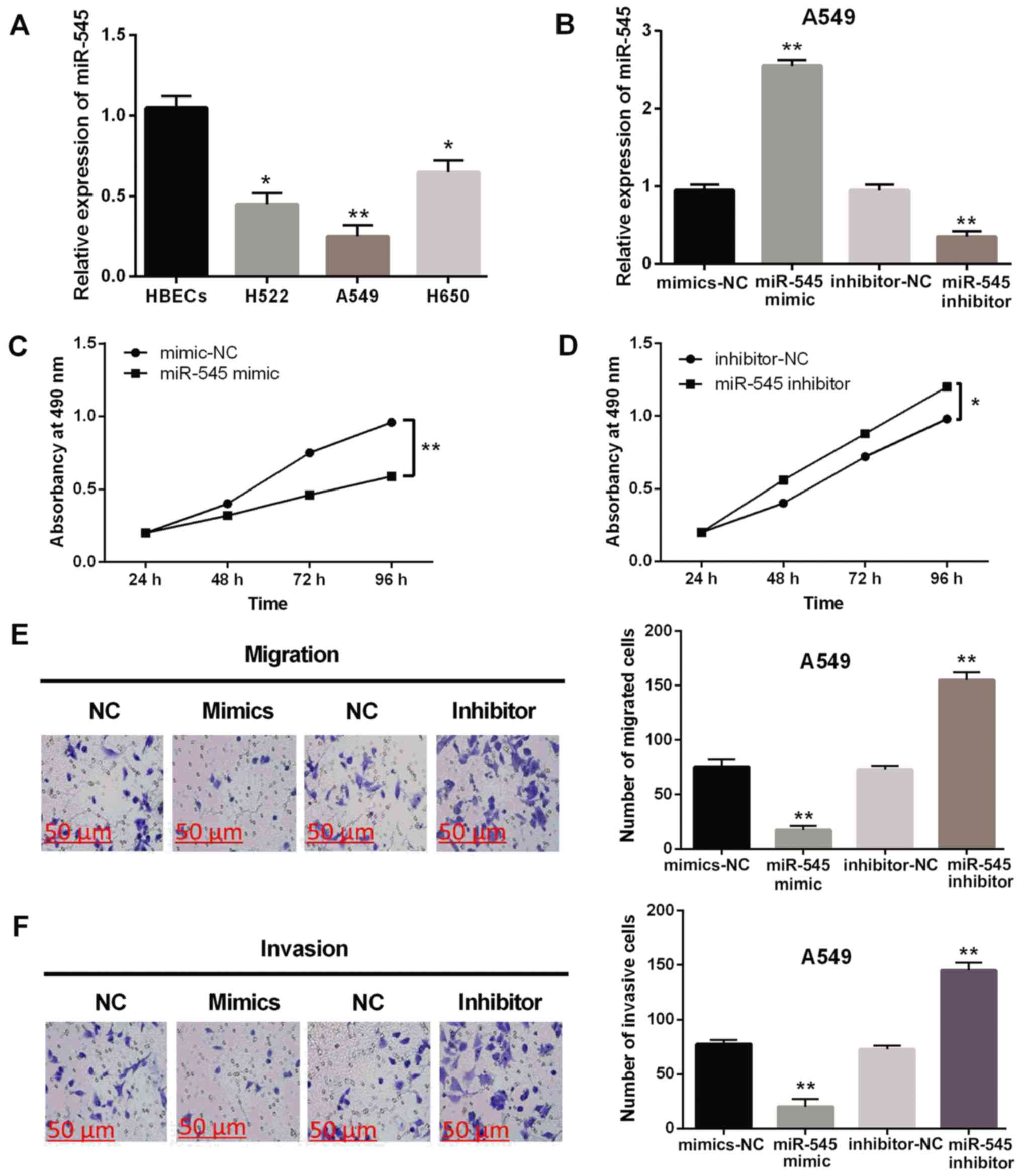
The expression levels of miR-545 were observed in
H522, A549, H650 and HBECs cell lines. Decreased miR-545 expression
was identified in H522, A549 and H650 cell lines compared with HBEC
cells (P<0.05 or 0.01, Fig. 2A).
In order to investigate the function of miR-545 in NSCLC, A549
cells were transfected with miR-545 mimics or inhibitor. The
RT-qPCR assay showed that miR-545 expression level was enhanced by
miR-545 mimics and inhibited by miR-545 inhibitor (P<0.01,
Fig. 2B). MTT assay suggested that
upregulation of miR-545 inhibits the proliferation of A549 cells
(P<0.01, Fig. 2C), whereas, cell
proliferation was promoted by the knockout of miR-545 in NSCLC
(P<0.05, Fig. 2D). Similarly,
cell migration was also suppressed by upregulation of miR-545 and
promoted by the knockout of miR-545 in A549 cells (P<0.01,
Fig. 2E). Cell invasion in A549
cells was also identified with similar results (P<0.01, Fig. 2F). These results indicate that
miR-545 plays a suppressive role in the progression of NSCLC.
ZEB2 is a direct target gene of
miR-545
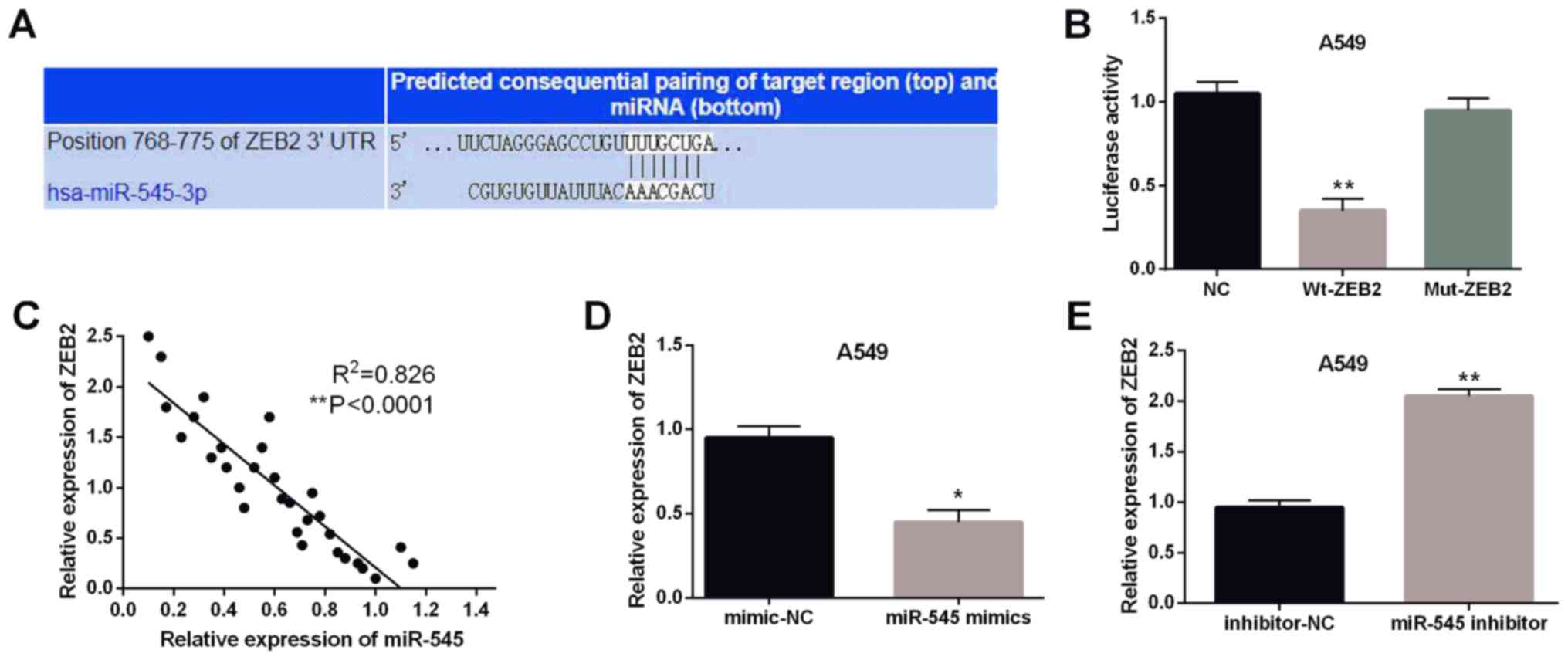
Furthermore, miR-545 was predicted to have binding
sites with the 3′-UTR of ZEB2 in the database of TargetScan
(http://www.targetscan.org/) (Fig. 3A). Then luciferase reporter assay was
performed to confirm the above prediction. As expected, the
luciferase activity of Wt-ZEB2 was obviously inhibited by miR-545
mimics (P<0.01, Fig. 3B).
However, the luciferase activity of Mut-ZEB2 was not affected by
miR-545 mimics. Moreover, the negative association between miR-545
and ZEB2 was detected in NSCLC tissues (P<0.0001,
R2=0.826; Fig. 3C). To
further confirm their relationship, ZEB2 expression was examined in
A549 cells with miR-545 mimics or inhibitor. Consistently, the
expression level of ZEB2 was declined by miR-545 mimics (P<0.05,
Fig. 3D) and was enhanced by miR-545
inhibitor (P<0.01, Fig. 3E).
These results confirmed that miR-495 directly targeted ZEB2 and had
negative association with ZEB2 expression in NSCLC tissues.
The knockout of ZEB2 suppresses the
development of NSCLC
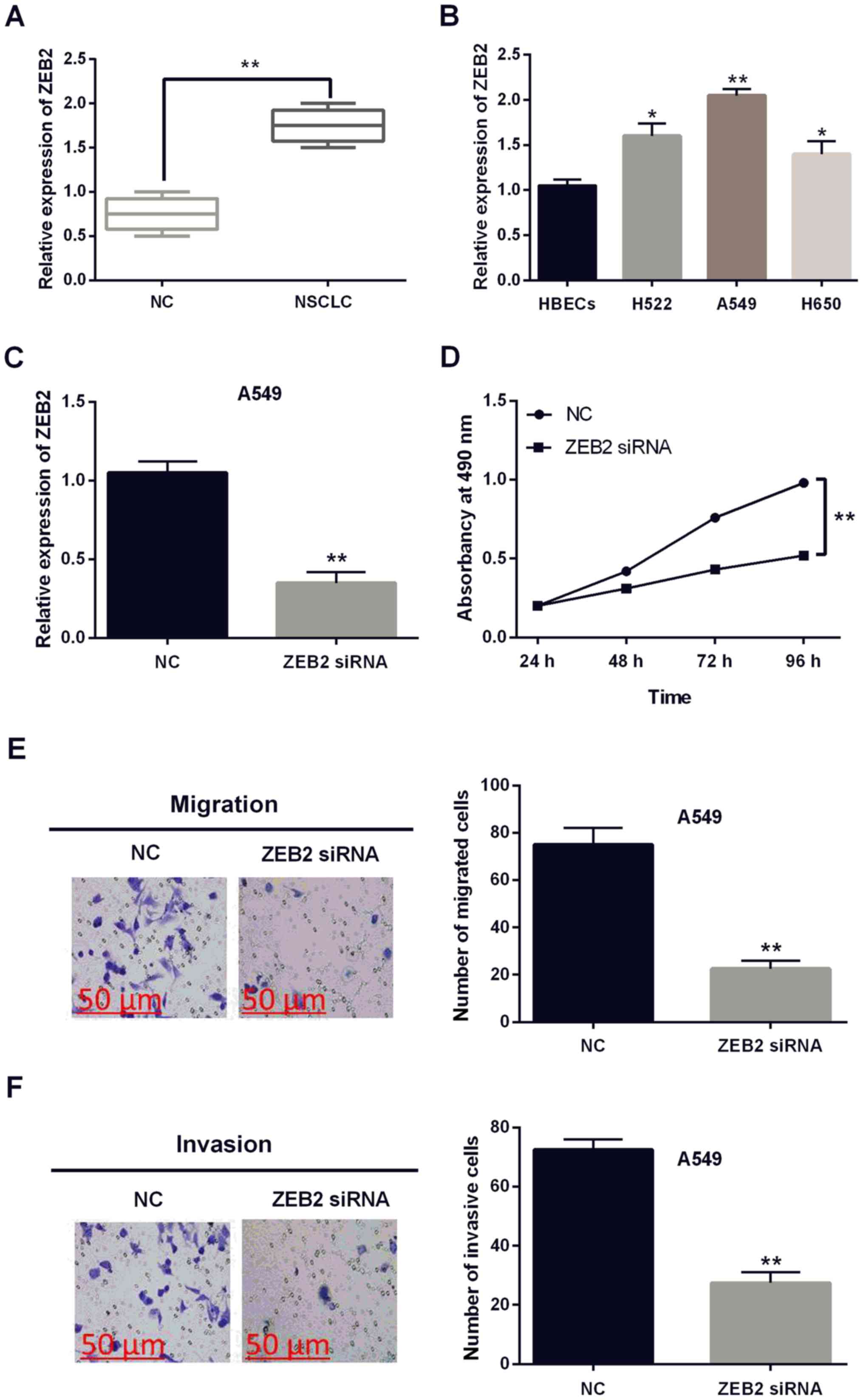
Subsequently, the expression of ZEB2 was identified
in NSCLC tissues and cell lines. We found that ZEB2 was upregulated
in NSCLC tissues in contrast to the normal tissues (P<0.01,
Fig. 4A). Increased ZEB2 expression
was also identified in H522, A549 and H650 cell lines compared with
HBECs cells (P<0.05 or 0.01, Fig.
4B). ZEB2 siRNA was transfected into A549 cells to investigate
its role in NSCLC. The expression of ZEB2 was significantly
declined by ZEB2 siRNA (P<0.01, Fig.
4C). Furthermore, knockout of ZEB2 suppressed the proliferation
of A549 cells (P<0.01, Fig. 4D).
Moreover, cell migration and invasion were also found to be
inhibited by ZEB2 siRNA (P<0.01, Fig.
4E and F). These findings revealed that ZEB2 had carcinogenic
effect on NSCLC.
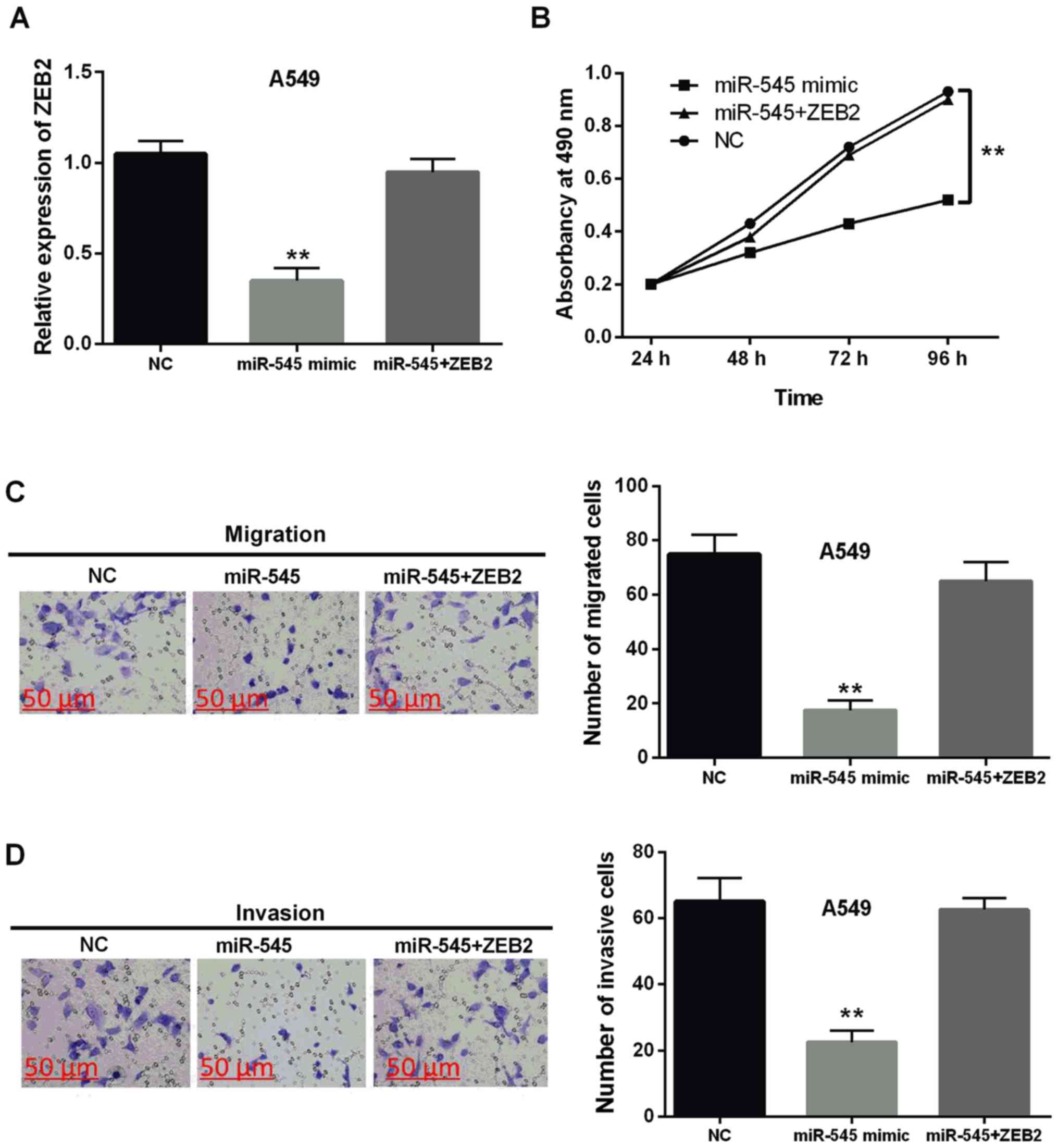
miR-545 inhibits progression of NSCLC
through targeting ZEB2
miR-545 mimics and ZEB2 vector were co-transfected
into A549 cells to evaluate their interaction. It was found that
the decreased ZEB2 expression induced by miR-545 mimics was
restored by ZEB2 vector in A549 cells (P<0.01, Fig. 5A). Similarly, the inhibitory effect
of miR-545 for cell proliferation was impaired by ZEB2 vector
(P<0.01, Fig. 5B). Moreover, the
diminished role of miR-545 for cell migration and invasion was also
impeded by ZEB2 vector in NSCLC (P<0.01, Fig. 5C and D). Collectively, miR-545
inhibited the progression of NSCLC through targeting ZEB2.
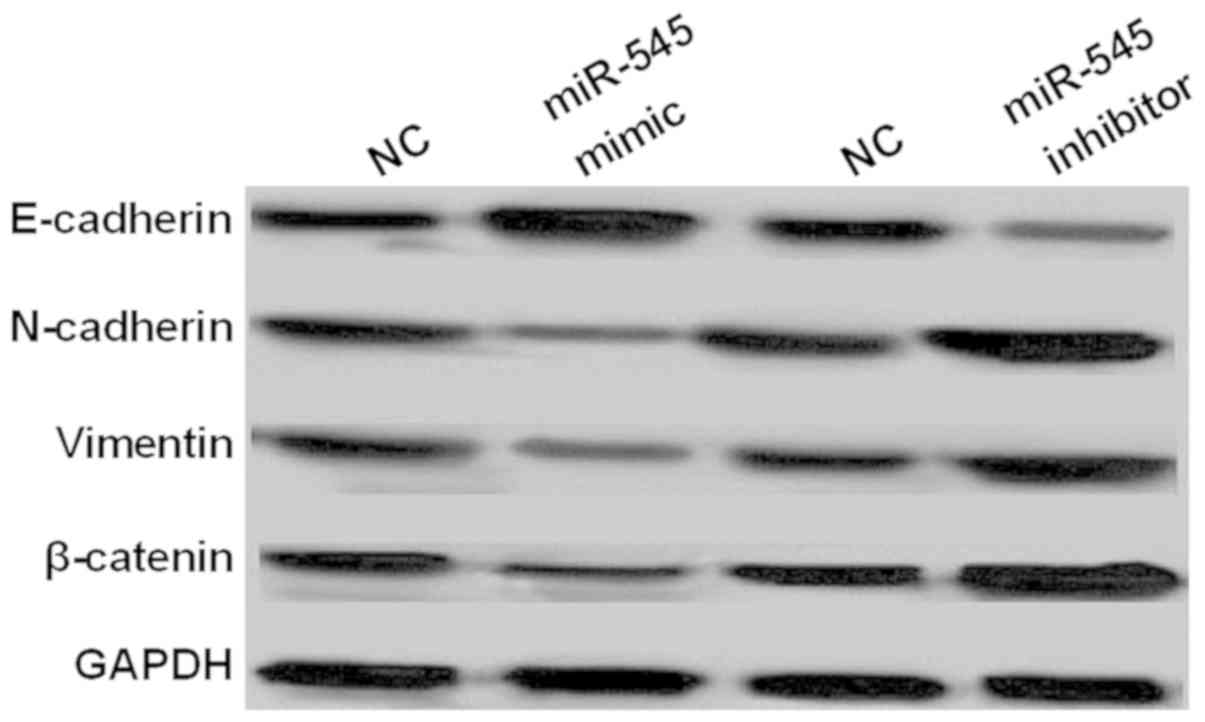
miR-545 blocks EMT and Wnt/β-catenin
pathway in NSCLC
In order to further confirm the effect of miR-545 on
cell survival and metastasis, EMT and Wnt/β-catenin pathway was
investigated in NSCLC. It was found that the expression levels of
N-cadherin and vimentin were inhibited by overexpression of miR-545
which promoted E-cadherin expression in A549 cells (Fig. 6). Whereas, the knockout of miR-545
had opposite effect on the expression levels of the three markers
(Fig. 6). The results suggested that
miR-545 inhibited the metastasis of NSCLC cells through blocking
EMT. Moreover, expression of β-catenin involved in Wnt/β-catenin
pathway was also inhibited by overexpression of miR-545 (Fig. 6), while the knockout of miR-545
promoted β-catenin expression (Fig.
6). In total, miR-545 repressed cell survival and metastasis
through blocking EMT and Wnt/β-catenin pathway in NSCLC.
Discussion
Recently, exploiting and antagonizing microRNA
regulation have been reported in therapeutic and experimental
applications (17). Moreover, many
miRNAs have been found to contribute or attenuate the carcinogenic
effect of NSCLC, such as miR-101 (18) and miR-4443 (19). In the current study, we paid
attention to the abnormal expression and function of miR-545.
miR-545 was identified to be downregulated in NSCLC which was
associated with worse prognosis of NSCLC patients. Similarly, Liao
et al (20) found
downregulation of miR-545 in Lewis lung carcinoma. In addition,
miR-545 was related to poor prognosis of HCC patients shortening
the 5-year overall survival (OS) (21). MTT and Transwell assays were
performed to further explore the function of miR-545 in NSCLC. We
found that overexpression of miR-545 inhibited the proliferation,
migration and invasion of NSCLC cells. A previous study reported
the tumor suppressing role of miR-545 in epithelial ovarian cancer
(22). miR-545 negatively mediated
colorectal cancer cell proliferation through upregulating EGFR
expression (23). In the present
study, miR-545 inhibited the progression of NSCLC through targeting
ZEB2.
Previous studies have implied that ZEB2 was a vital
oncogene which contributed to the progression of multiple cancers
including NSCLC (24). Here,
upregulation of ZEB2 was identified in NSCLC and the knockout of
ZEB2 suppressed the development of NSCLC. Similar results of ZEB2
was also detected in osteosarcoma (25). miR-200c inhibited metastasis of NSCLC
cells by targeting ZEB2 (26) which
was consistent with our findings. Additionally, ZEB2 was reported
to induce EMT via suppressing E-cadherin and promoting
tumorigenesis (27). EMT is
recognized as a crucial biological process which is related to
metastasis of cancers. This study indicates that miR-545/ZEB2 axis
blocked EMT to metastasis of NSCLC. Shi et al (28) demonstrated that downregulation of
miR-218 contributed to EMT and tumor metastasis in lung cancer by
targeting ZEB2. ZEB2 has been reported to mediate multiple pathways
regulating cell proliferation, migration, invasion, and apoptosis
in glioma, including Wnt/β-catenin pathway (29). We also observed that miR-545/ZEB2
blocked Wnt/β-catenin pathway to regulate the progression of
NSCLC.
Accumulated evidence demonstrate that aberrant
activation of the Wnt/β-catenin pathway could result in tumor
formation (30). Previous studies
demonstrated that Wnt/β-catenin pathway was involved in
pathogenesis of NSCLC (31). Qi
et al (29) revealed that
downregulation of ZEB2 suppressed cell proliferation and reduced
β-catenin expression. It is well known that β-catenin is a key
regulator in Wnt/β-catenin pathway. In the present study,
overexpression of miR-545 declined β-catenin expression and miR-545
inhibited the development of NSCLC by targeting ZEB2 and
inactivating Wnt/β-catenin pathway. Similarly, miR-145 also
inhibited hepatic stellate cell proliferation by targeting ZEB2
through Wnt/β-catenin pathway (32).
Combined with these results, we considered that the suppressive
effect of miR-545 on the development of NSCLC was partly exhibited
through the abnormal ZEB2 expression involved in Wnt/β-catenin
pathway.
In conclusion, downregulation of miR-545 was
identified in NSCLC which is involved in tumorigenesis and
prognosis of NSCLC. Moreover, ZEB2 was confirmed as a direct target
gene of miR-545. miR-545 inhibited progression of NSCLC through
targeting ZEB2 and blocking EMT and Wnt/β-catenin pathway. These
results can potentially help in elucidation of the regulatory
mechanism of miR-545 in NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
JC contributed to the study design and drafted the
manuscript. GP was involved in data acquisition and revision of the
manuscript. QH was involved in data acquisition and analysis. LY
contributed to data analysis. RG contributed to data acquisition
and analysis and revised the manuscript. HB contributed to the
conception and design of the study. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
All the patients provided an informed consent and
this experiment was approved by the Institutional Ethics Committee
of The Third People's Hospital of Qingdao (Qingdao, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Field JK, Oudkerk M, Pedersen JH and Duffy
SW: Prospects for population screening and diagnosis of lung
cancer. Lancet. 382:732–741. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang L, Yu C, Liu Y, Wang J, Li C, Wang Q,
Wang P, Wu S and Zhang ZJ: Lung cancer mortality trends in China
from 1988 to 2013: New challenges and opportunities for the
government. Int J Environ Res Public Health. 13:E10522016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Reck M, Popat S, Reinmuth N, De Ruysscher
D, Kerr KM and Peters S; ESMO Guidelines Working Group, :
Metastatic non-small-cell lung cancer (NSCLC): ESMO Clinical
Practice Guidelines for diagnosis, treatment and follow-up. Ann
Oncol. 25 (Suppl 3):iii27–iii39. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pao W and Girard N: New driver mutations
in non-small-cell lung cancer. Lancet Oncol. 12:175–180. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Araujo LH and Carbone DP: Non-small cell
lung cancer genomics around the globe: Focus on ethnicity. J Thorac
Dis. 9:E392–E394. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lv X, Li CY, Han P and Xu XY:
MicroRNA-520a-3p inhibits cell growth and metastasis of non-small
cell lung cancer through PI3K/AKT/mTOR signaling pathway. Eur Rev
Med Pharmacol Sci. 22:2321–2327. 2018.PubMed/NCBI
|
|
7
|
Fei X, Zhang J, Zhao Y, Sun M, Zhao H and
Li S: miR-96 promotes invasion and metastasis by targeting GPC3 in
non-small cell lung cancer cells. Oncol Lett. 15:9081–9086.
2018.PubMed/NCBI
|
|
8
|
Song B, Ji W, Guo S, Liu A, Jing W, Shao
C, Li G and Jin G: miR-545 inhibited pancreatic ductal
adenocarcinoma growth by targeting RIG-I. FEBS Lett. 588:4375–4381.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cosín-Tomás M, Antonell A, Lladó A,
Alcolea D, Fortea J, Ezquerra M, Lleó A, Martí MJ, Pallàs M,
Sanchez-Valle R, et al: Plasma miR-34a-5p and miR-545-3p as early
biomarkers of Alzheimer's disease: Potential and limitations. Mol
Neurobiol. 54:5550–5562. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Du B, Wang Z, Zhang X, Feng S, Wang G, He
J and Zhang B: MicroRNA-545 suppresses cell proliferation by
targeting cyclin D1 and CDK4 in lung cancer cells. PLoS One.
9:e880222014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gheldof A, Hulpiau P, van Roy F, De Craene
B and Berx G: Evolutionary functional analysis and molecular
regulation of the ZEB transcription factors. Cell Mol Life Sci.
69:2527–2541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chua HL, Bhat-Nakshatri P, Clare SE,
Morimiya A, Badve S and Nakshatri H: NF-kappaB represses E-cadherin
expression and enhances epithelial to mesenchymal transition of
mammary epithelial cells: Potential involvement of ZEB-1 and ZEB-2.
Oncogene. 26:711–724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
You J, Li Y, Fang N, Liu B, Zu L, Chang R,
Li X and Zhou Q: MiR-132 suppresses the migration and invasion of
lung cancer cells via targeting the EMT regulator ZEB2. PLoS One.
9:e918272014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cong N, Du P, Zhang A, Shen F, Su J, Pu P,
Wang T, Zjang J, Kang C and Zhang Q: Downregulated microRNA-200a
promotes EMT and tumor growth through the wnt/β-catenin pathway by
targeting the E-cadherin repressors ZEB1/ZEB2 in gastric
adenocarcinoma. Oncol Rep. 29:1579–1587. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Q, Zhang B, Sun L, Yan Q, Zhang Y,
Zhang Z, Su Y and Wang C: MicroRNA-130b targets PTEN to induce
resistance to cisplatin in lung cancer cells by activating
Wnt/β-catenin pathway. Cell Biochem Funct. 36:194–202. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brown BD and Naldini L: Exploiting and
antagonizing microRNA regulation for therapeutic and experimental
applications. Nat Rev Genet. 10:578–585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Han L, Chen W, Xia Y, Song Y, Zhao Z,
Cheng H and Jiang T: MiR-101 inhibits the proliferation and
metastasis of lung cancer by targeting zinc finger E-box binding
homeobox 1. Am J Transl Res. 10:1172–1183. 2018.PubMed/NCBI
|
|
19
|
Zhang W, Qiao B and Fan J: Overexpression
of miR-4443 promotes the resistance of non-small cell lung cancer
cells to epirubicin by targeting INPP4A and regulating the
activation of JAK2/STAT3 pathway. Pharmazie. 73:386–392.
2018.PubMed/NCBI
|
|
20
|
Liao C, Xiao W, Zhu N, Liu Z, Yang J, Wang
Y and Hong M: MicroR-545 enhanced radiosensitivity via suppressing
Ku70 expression in Lewis lung carcinoma xenograft model. Cancer
Cell Int. 15:562015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Z, Dou C, Yao B, Xu M, Ding L, Wang Y,
Jia Y, Li Q, Zhang H, Tu K, et al: Ftx non coding RNA-derived
miR-545 promotes cell proliferation by targeting RIG-I in
hepatocellular carcinoma. Oncotarget. 7:25350–25365.
2016.PubMed/NCBI
|
|
22
|
Jia X, Liu X, Li M, Zeng Y, Feng Z, Su X,
Huang Y, Chen M and Yang X: Potential tumor suppressing role of
microRNA-545 in epithelial ovarian cancer. Oncol Lett.
15:6386–6392. 2018.PubMed/NCBI
|
|
23
|
Huang X and Lu S: MicroR-545 mediates
colorectal cancer cells proliferation through up-regulating
epidermal growth factor receptor expression in HOTAIR long
non-coding RNA dependent. Mol Cell Biochem. 431:45–54. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin X, Yang Z, Zhang P, Liu Y and Shao G:
miR-154 inhibits migration and invasion of human non-small cell
lung cancer by targeting ZEB2. Oncol Lett. 12:301–306. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fei D, Zhao K, Yuan H, Xing J and Zhao D:
MicroRNA-187 exerts tumor-suppressing functions in osteosarcoma by
targeting ZEB2. Am J Cancer Res. 6:2859–2868. 2016.PubMed/NCBI
|
|
26
|
Jiao A, Sui M, Zhang L, Sun P, Geng D,
Zhang W, Wang X and Li J: MicroRNA-200c inhibits the metastasis of
non-small cell lung cancer cells by targeting ZEB2, an
epithelial-mesenchymal transition regulator. Mol Med Rep.
13:3349–3355. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Comijn J, Berx G, Vermassen P, Verschueren
K, van Grunsven L, Bruyneel E, Mareel M, Huylebroeck D and van Roy
F: The two-handed E box binding zinc finger protein SIP1
downregulates E-cadherin and induces invasion. Mol Cell.
7:1267–1278. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi ZM, Wang L, Shen H, Jiang CF, Ge X, Li
DM, Wen YY, Sun HR, Pan MH, Li W, et al: Downregulation of miR-218
contributes to epithelial-mesenchymal transition and tumor
metastasis in lung cancer by targeting Slug/ZEB2 signaling.
Oncogene. 36:2577–2588. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qi S, Song Y, Peng Y, Wang H, Long H, Yu
X, Li Z, Fang L, Wu A, Luo W, et al: ZEB2 mediates multiple
pathways regulating cell proliferation, migration, invasion, and
apoptosis in glioma. PLoS One. 7:e388422012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Polakis P: Wnt signaling in cancer. Cold
Spring Harb Perspect Biol. 4:a0080522012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang XH, Cui YX, Wang ZM and Liu J:
Down-regulation of FOXR2 inhibits non-small cell lung cancer cell
proliferation and invasion through the Wnt/β-catenin signaling
pathway. Biochem Biophys Res Commun. 500:229–235. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou DD, Wang X, Wang Y, Xiang XJ, Liang
ZC, Zhou Y, Xu A, Bi CH and Zhang L: MicroRNA-145 inhibits hepatic
stellate cell activation and proliferation by targeting ZEB2
through Wnt/β-catenin pathway. Mol Immunol. 75:151–160. 2016.
View Article : Google Scholar : PubMed/NCBI
|