Introduction
Esophageal squamous cell carcinoma (ESCC) is a
common tumor of the digestive tract. The incidence and mortality of
ESCC varies among countries, and China has a high ESCC risk
(1,2). Thanks to the development of genetic and
molecular biology, pathogenesis of ESCC is now better understood.
Particularly, the association between microRNAs (miRs) and cancer
has been investigated. Recent studies have demonstrated that
abnormal expression of miRs is detected in the serum of patients
with ESCC (3–5). These findings may provide new direction
for further study of the progression of ESCC and to determine novel
potential therapeutic targets.
miRs are single-stranded non-coding small RNAs that
are typically 18–25 nucleotides in length. They mainly bind at a
post-transcriptional level to the target mRNAs, which are
subsequently degraded and not translated into protein. miRs can
precisely regulate certain biological processes, including cell
proliferation, apoptosis, migration and immune responses (6,7). It has
been reported that dysregulated miRs might have a role in of tumors
development (8). Notably, miR-490-3p
has been reported to be associated with the incidence of various
types of cancer (1,9–16). In
the present study, analysis of miRs expression in ESCC tissues and
adjacent tissues from The Cancer Genome Atlas (TCGA) database
demonstrated that miR-490-3p was significantly lower in ESCC
tissues; however, the role of miR-490-3p in ESCC remains
unknown.
Mitogen-activated protein kinases (MAPKs) are a
family of serine/threonine kinases that can be activated by various
signals (17). Their functions have
remained highly evolutionarily conserved (18). MAPK1, also known as ERK2, is a ~42 kD
protein and is mediates one of the most important signaling
pathways in vivo, with a crucial role in cell growth,
differentiation, proliferation, survival and in the inflammatory
response (19). A previous study
reported that the MAPK1 signaling pathway is crucial in most cells
and is regulated at numerous levels. It can respond to external
physiological and pathological stimuli, and to extracellular
stimuli of the nucleus (17,20,21).
The present study aimed to investigate the effect of
miR-490-3p on the proliferation and apoptosis of the ESCC TE1 and
ECA109 cell lines, and to determine whether miRNA-490-3p exerts its
biological function through regulation of MAPK1 expression.
Materials and methods
Data acquisition and analysis
The clinical data and the corresponding genome-wide
expression profile (TCGA-ESCA) using RNA-sequencing of 185 patients
with ESCC were obtained from TCGA database (https://cancergenome.nih.gov). A total of 100 cases
were subsequently randomly selected for further analysis. The TCGA
data was normalization by the MD Anderson Cancer Center. Two-way
hierarchical clustering of 88 tumor tissues and 12 adjacent normal
mucosa with the 2,114 differentially expressed RNAs were analyzed
using Euclidean distance and average linkage clustering. P<0.05
and |log fold change|>1 were defined as cut-off values.
Gene enrichment analysis
Gene Set Enrichment Analysis (GSEA) version 2.2.3
software (http://software.broadinstitute.org/gsea/index.jsp;
Broad Institute, Inc.) was used for data analysis. According to the
expression level of miR-490-3p, all cases were divided into the
miR-490-3p high expression group and low expression group, using
the aforementioned cut off values. In the present study, the
dataset of C2.CP.KEGG.v6.0 was obtained from the MSigDB database of
the GSEA website. The enrichment analysis was performed according
to the default weighted enrichment statistics method, and the
number of random combinations was set to 1,000 times.
Patients and sample collection
New ESCC tissues and paracarcinoma tissues were
obtained from 40 patients, including 18 women and 22 men, aged from
48–69 years old, diagnosed with ESCC and received surgical
treatment between July 2016 and September 2017 at The Affiliated
Tumor Hospital of Xinjiang Medical University. Patients had not
received any treatment prior to surgery and had no family history
of ESCC. Patients were diagnosed with ESCC by pathological analysis
of the tumors. All patients voluntarily participated in the study
and provided signed informed consent. The present study was
approved by The Affiliated Tumor Hospital of Xinjiang Medical
University Ethics Committee. After radical esophagectomy, tumor
tissues tissue did not contain any necrotic area, and adjacent
normal tissues that were resected at >5 cm from cancer tissue,
were stored in liquid nitrogen.
Cell culture
The poorly differentiated ESCC cell line TE1, the
highly differentiated ESCC cell line ECA109 and the control cell
line HEEC were purchased from Shanghai American Type Culture
Collection. Cells were cultured in Dulbecco's modified eagle medium
containing 10% FBS, 1% penicillin and 1% streptomycin and placed at
37°C in a humidified incubator containing 5% CO2 (Gibco;
Thermo Fisher Scientific, Inc.). After 48 h incubation, culture
medium was replaced every other day. Once cells had reached 80–90%
confluence, they were harvested with 0.25% trypsin and passaged at
a ratio of 1:2 or 1:3.
Cell transfection
Cells in the logarithmic growth phase were selected
for seeding into 24-well plates at 1×105 cells/well.
Transfection was performed using the Lipofectamine™ 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). When the cell density
reached 70–80%, they were transfected with miR-490-3p mimics (20
µM), miR-490-3p inhibitors (20 µM), pcDNA-MAPK1 (5 µg) and the
relative negative controls (Shanghai GenePharma Co., Ltd.). The
sequences were as follows: miR-490-3p mimics forward,
5′-CAACCUGGAGGACUCCAUGCCG-3′ and reverse,
5′-AGACCGUCGAUUGGGCCAGUUG-3′; miR-490-3p inhibitors forward,
5′-UUUAGCUGGUACCGACUGUACG-3′ and reverse,
5′-AUUCGGUAGUUUCAGGGCAUAG-3′; negative controls forward
5′-ACCUUCUCAGGCUUGACGUAGA-3′ and reverse,
5′-CCGUAUUAUCUGCCAAGUACGU-3′. At 48 h after transfection, cells
were collected for further experiments.
Tissues and cellular RNA
extraction
1×106 Cells or 50 mg tissues were
collected and lysed by adding 1 ml TRIzol® (Invitrogen;
Thermo Fisher Scientific, Inc.). Cells and tissues were then
incubated with 250 µl chloroform, mixed for 30 sec and centrifuged
at 10,000 × g at 4°C for 10 min. The aqueous phase was discarded
and same volume of pre-cooled isopropanol was added. The
precipitate was gently cleaned with 75% ethanol after
centrifugation at 10,000 × g for 10 min at 4°C. Diethyl
pyrocarbonate (20 µl) water was used to dissolve RNAs. The RNA
concentration was measured using a spectrophotometer and stored at
−80°C.
Reverse transcription-quantitative PCR
(RT-qPCR)
RT was performed using the PrimeScript RT Reagent
kit (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocol. RT was performed on ice and produced
cDNAs. qPCR was performed according to the miScript SYBR Green PCR
kit instructions (Takara Bio, Inc.) for miR-490-3p, MAPK and GAPDH.
The PCR amplification conditions were as follows: Pre-denaturation
at 94°C for 5 min, followed by 94°C for 30 sec, 55°C for 30 sec,
and 72°C for 1 min and 30 sec for 40 cycles.
The primers were designed as follows: miR-490-3p,
forward 5′-CGGCGGTCAACCTGGAGGACTCC-3′, reverse
5′-CCAGTGCAGGGTCCGAGGTAT-3′; MAPK1, forward
5′-CGACGCGTCGTTGTAATAAAGCCTCCAG-3′, reverse
5′-GACTAGTCGTTTTCATTTCAATCGTAG-3′; GAPDH, forward
5′-AGCCACATCGCTCAGACAC-3′, reverse 5′-GCCCAATACGACCAAATCC-3′; and
U6, forward 5′-CTCGCTTCGGCAGCAGCACATATA-3′, and reverse
5′-AAATATGGAACGCTTCACGA-3′. The 2−ΔΔCq method was used
to determine the relative expression levels (22). The level of mRNA expression was
normalized to GAPDH, while miRNA expression levels were normalized
to U6.
Cell counting kit-8 (CCK-8) assay
At 48 h after transfection, TE1 and ECA109 cells
were plated in 96-well plates (1×104 cells/100 µl/well).
Five replicates were set for each sample. Cells were cultured at
37°C and 5% CO2 for 0, 24, 48 72 and 96 h. CCK-8 reagent
(10 µl; Dojindo Molecular Technologies, Inc.) was then added to
cells and incubated for 2 h at 37°C. Absorbance was measured at 450
nm using a microplate reader.
Plate cloning experiment
TE1 and ECA109 cells in the logarithmic growth phase
were harvested, and the cell suspension concentration was adjusted
to 1×104/ml. Cells were seeded in a 6-well plate at the
density of 2,000 cell/well. Following 3–4 days culture at 37°C,
cells were fixed with 4% paraformaldehyde for 30 min and stained
with 0.1% crystal violet for 30 min at 25°C. The numbers of clones
per well were counted and imaged.
Flow cytometry
Cells were harvested with trypsin and suspended with
100 µl 1X Annexin V binding buffer (AmyJet Scientific, Inc.) at a
minimum density of 1×105 cells/ml. Annexin V-FITC was
used for cell labeling. Annexin V (5 µl) and propidium iodide (1
µl) were added to the cell suspension, mixed, and incubated at room
temperature in the dark for 15 min. 1X Annexin V binding buffer
(400 µl) was then added, and cell apoptosis was measured on a flow
cytometer. Each sample was analyzed in triplicate and the
experiment was repeated three times.
Bioinformatics
The target genes for miR-490-3p were determined
through bioinformatics prior to performing functional analysis. The
basic information of base sequence, chromosome location and species
conservativeness of miR-490-3p was obtained by using miR Base
(http://www.mirbase.org/index.shtml).
The target genes of miR-490-3p were predicted and the intersection
was taken as the gene set for further analysis by four methods,
including TargetScan (http://www.targetscan.org/), MicroRanda (http://www.microrna.org/), RNA hybrid (version 2.1.2)
and DIANAmT from MicroWalk (umm.uni-heidelberg.de/apps/zmf/mirwalk/) integrated
database. Cytoscape software (version 3.7.1; http://cytoscape.org/) and its plug-in Bingo was
applied for function enrichment analysis.
Dual luciferase reporter assay
The 3′untranslated region (3′UTR) sequence of MAPK1
was downloaded from the NCBI website (https://www.ncbi.nlm.nih.gov/) to construct the
wild-type MAPK1 (MAPK1 WT 3′UTR) and the mutant (MAPK1 MUT 3′UTR)
pGL3-plasmids (Hanbio Biotechnology Co., Ltd.). pRL-TK
Renilla plasmid (Promega Corporation) was used as the
control luciferase for normalization. Cells were plated in 96-well
plates at 1×104 cells/well and co-transfected with 50
pmol/l miR-490-3p mimics or negative controls with 80 ng MAPK1 WT
3′UTR or MAPK1 MUT 3′UTR plasmid using Lipofectamine™ 2000. After
48 h, Dual Luciferase® reporter assay system (Promega
Corporation) was used to detect fluorescence intensity.
Western blot analysis
Cells were lysed by radioimmunoprecipitation assay
buffer (Yeasen), sonicated, centrifuged at 5,000 × g for 10 min at
4°C after which the supernatant was collected. After adding
bromophenol blue (9.5 to 0.5 ml), supernatant was boiled for 10 min
and stored at −20°C. Proteins (10 µg) were separated by 12%
SDS-PAGE and transferred onto polyvinylidene fluoride membranes
(Roche Diagnostics). Membranes were then blocked using 5% skimmed
milk at 25°C for 1 h. Membranes were incubated with specific
primary antibodies at 4°C overnight, which included MAPK1 (1:500;
cat. no. ab241580; Abcam) and GAPDH (1:500; cat. no. ab8245;
Abcam). Then the membranes were incubated with secondary antibody
goat anti-rabbit IgG H&L (1:1,000; cat. no. ab7090; Abcam) the
next day at 25°C for 2 h. Protein bands were visualized following a
3-min incubation with enhanced chemiluminescence reagent (Thermo
Fisher Scientific, Inc.).
Statistical analysis
SPSS 19.0 (IBM Corp.) was used for statistical
analysis. χ2 test was used for classification data and
Student's t-test was used for data comparison between two groups.
Comparison between multiple groups was performed using One-way
ANOVA test followed by the least significant difference post hoc
test. Data were expressed as the mean ± standard deviation.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-490-3p expression level is lower
in ESCC tissues from TCGA database
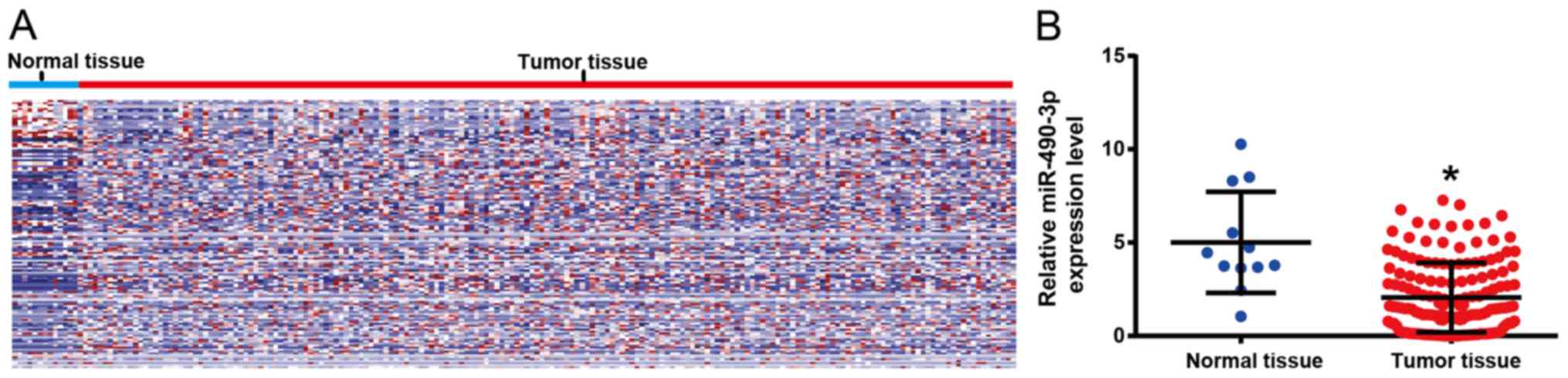
The differentially expressed miRs in ESCC tissues
and adjacent tissues from the TCGA database were analyzed. Two-way
hierarchical clustering of 988 tumor tissues and 12 adjacent normal
mucosa with the 2,114 differentially expressed RNAs were analyzed
using Euclidean distance and average linkage clustering (Fig. 1A). P<0.05 and |log fold
change|>1 were defined as cut-off values. The results
demonstrated that miR-490-3p was significantly lower in ESCC
tissues compared with adjacent tissues (Fig. 1B) and chosen for further analysis, as
it was the most significant.
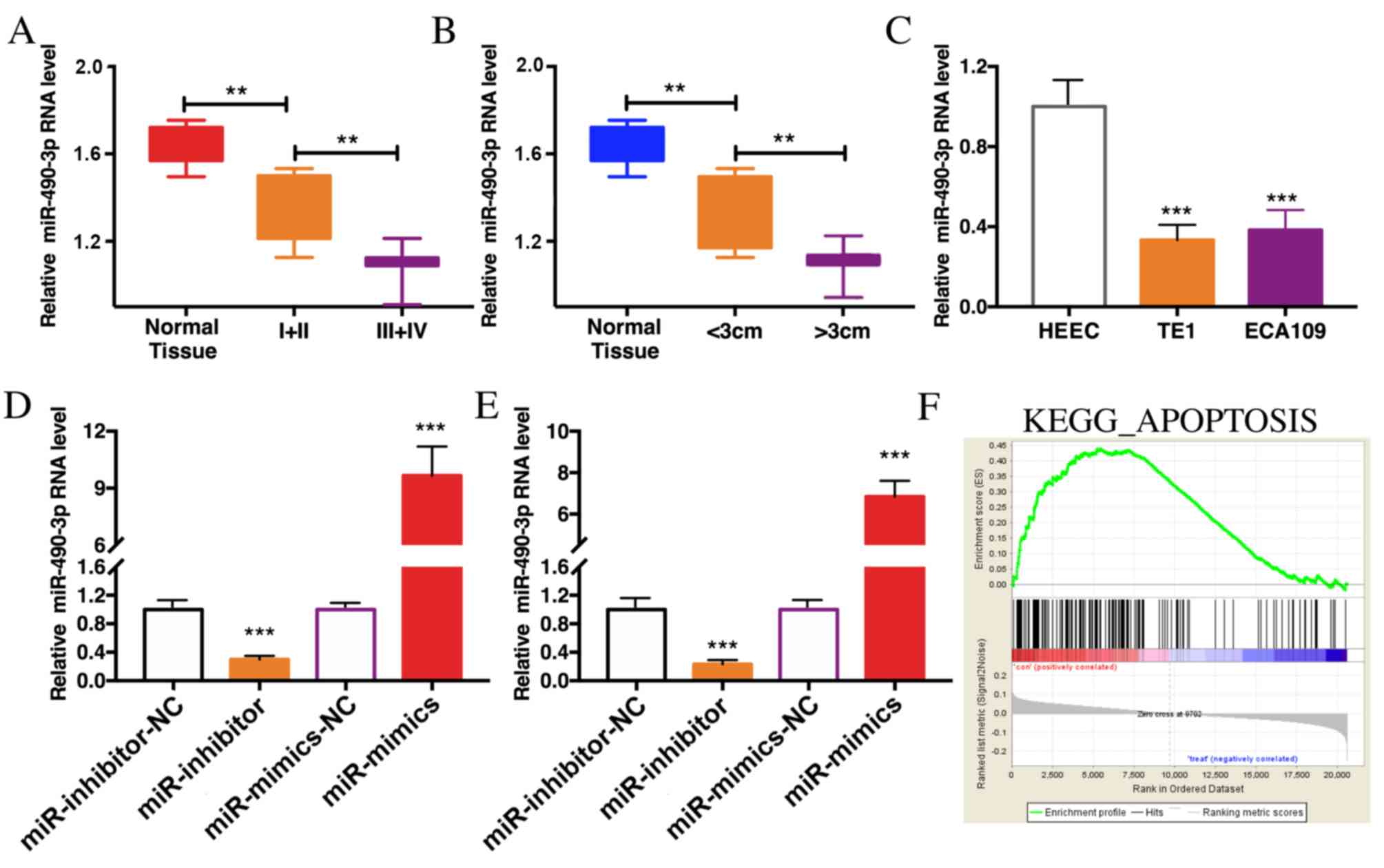
Expression of miR-490-3p is reduced in
ESCC
The expression of miR-490-3p in 40 cases of ESCC
tissues and paracancerous tissues were determined by RT-qPCR. The
results demonstrated that miR-490-3p was expression was lower in
ESCC tissues than in paracancerous tissues. In advanced ESCC
tissues with tumors >3 cm, miR-490-3p expression level was even
lower (Fig. 2A and B). In addition,
miR-490-3p expression level was significantly reduced in TE1 and
ECA109 cells compared with HEEC cells (Fig. 2C). To further investigate the role of
miR-490-3p in ESCC, cells were transfected with miR-490-3p mimics
or miR-490-3p inhibitors to overexpress or knockdown miR-490-3p,
respectively. The transfection efficiency was verified in TE1 and
ECA109 cells (Fig. 2D and E). In
addition, GSEA analysis reported that miR-490-3p was associated
with in apoptosis regulation in vivo (Fig. 2F).
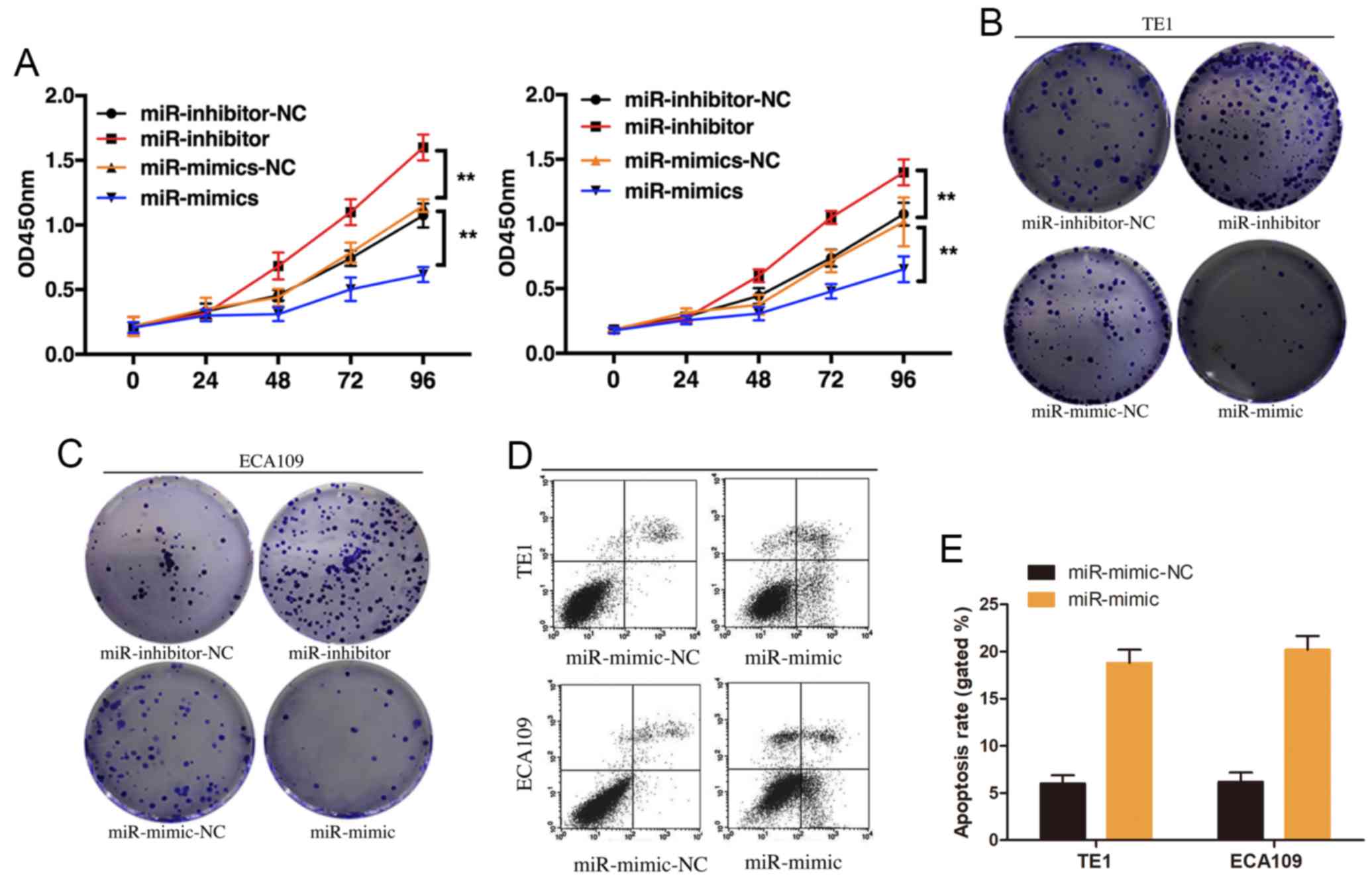
miR-490-3p induces apoptosis and
suppresses proliferation in ESCC cells
The effect of miR-490-3p on ESCC proliferation was
investigated by overexpressing or knocking down miR-490-3p in TE1
and ECA109 cell lines. CCK-8 results demonstrated that miR-490-3p
overexpression significantly inhibited ESCC cell proliferation,
whereas miR-490-3p knockdown promoted ESCC cell proliferation
(Fig. 3A). These results were
confirmed by the plate cloning formation experiment (Fig. 3B and C). The effect of miR-490-3p on
ESCC cell apoptosis was examined by flow cytometry. The results
demonstrated that miR-490-3p overexpression induced ESCC cell
apoptosis (Fig. 3D and E). These
results suggested that miR-490-3p may induce ESCC cell apoptosis
and suppress ESCC cell proliferation.
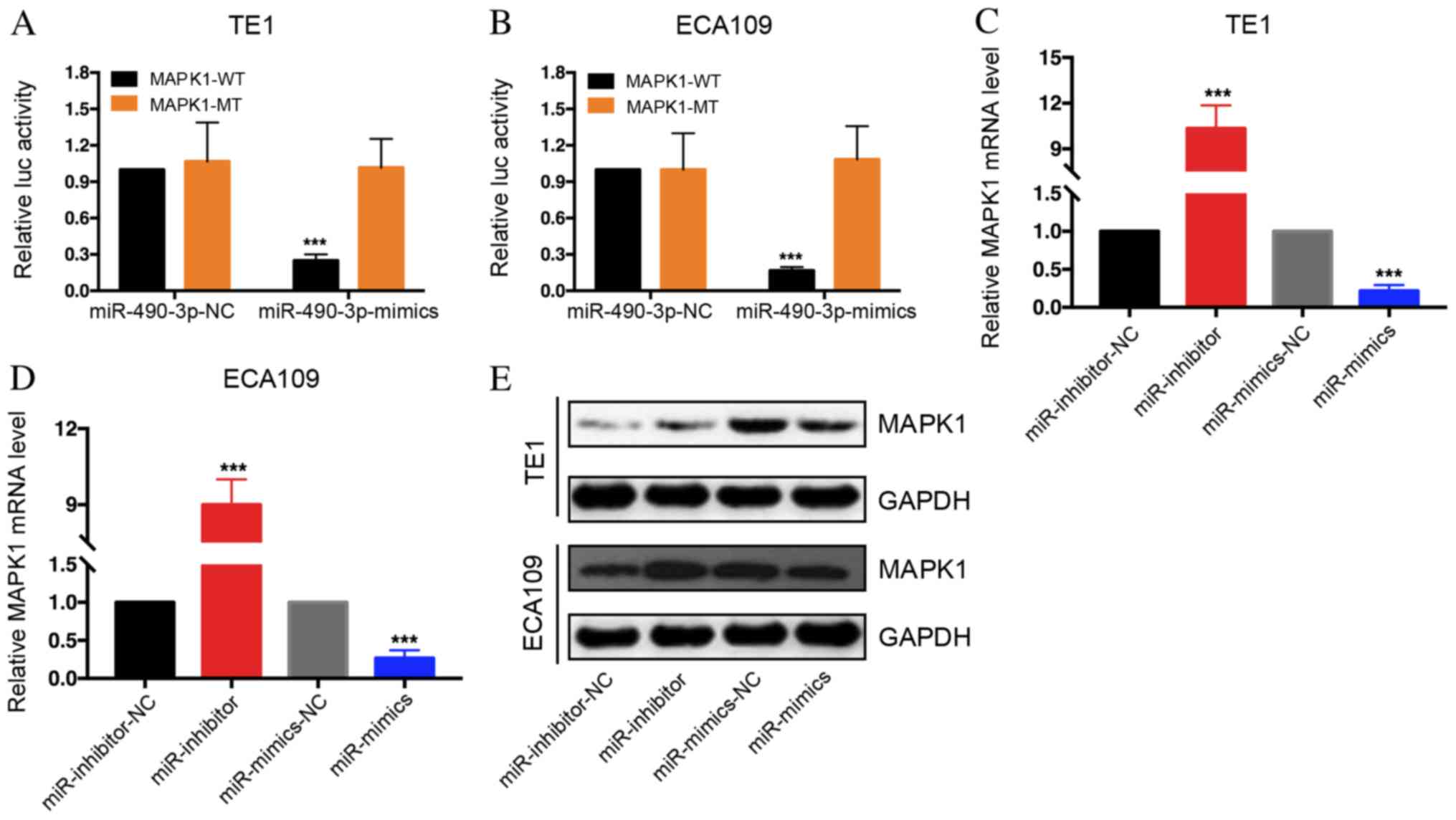
miR-490-3p may stimulate MAPK1
degradation
The target genes for miR-490-3p were determined
through bioinformatics prior to performing functional analysis. The
basic information of base sequence, chromosome location and species
conservativeness of miR-490-3p was obtained by using miR Base
(http://www.mirbase.org/index.shtml).
The target genes of miR-490-3p were predicted and the intersection
was taken as the gene set for further analysis by four methods,
including TargetScan, MicroRanda, RNA hybrid and DIANAmT from
MicroWalk (umm.uni-heidelberg.de/apps/zmf/mirwalk/) integrated
database. Cytoscape software (version 3.7.1; http://cytoscape.org/) and its plug-in Bingo was
applied for function enrichment analysis. As a result, it was
revealed that miR-490-3p may be able to target MAPK1 degradation.
Furthermore, luciferase activity of MAPK1-WT 3′UTR group was
decreased following miR-490-3p mimics transfection of TE1 and
ECA109 cells, whereas no difference was observed in the MAPK1-MUT
3′UTR group (Fig. 4A and B). These
results suggested that MAPK1 may bind to miR-490-3p. Subsequently,
MAPK1 mRNA expression level was determined following TE1 and ECA109
cell transfection with miR-490-3p mimics and miR-490-3p inhibitors.
The results demonstrated that MAPK1 expression level was decreased
in the mimics group but increased in the inhibitors group (Fig. 4C and D). Western blot analysis
revealed that MAPK1 expression was decreased in
miR-490-3p-overexpressing cells; however, MAPK1 expression was
increased in miR-490-3p-depleted cells (Fig. 4E). These results indicated that
miR-490-3p may regulate MAPK1 expression.
MAPK overexpression reverses the
effect of miR-490-3p on TE1 cell apoptosis
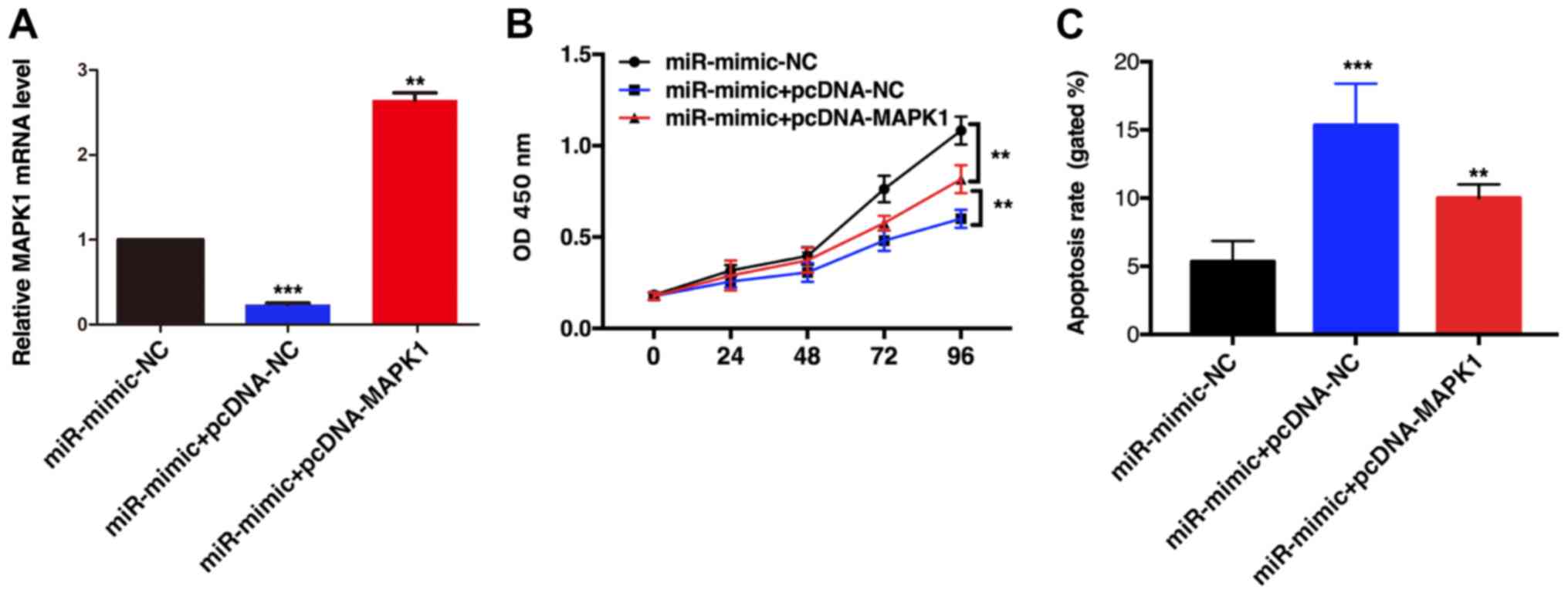
miR-490-3p overexpression significantly inhibited
TE1 cell proliferation; however, when MAPK1 and miR-490-3p were
simultaneously overexpressed, cell proliferation was increased,
although it was still lower than in the control group (Fig. 5A and B). In addition, MAPK1
overexpression partially reversed the miR-490-3p-induced TE1 cell
apoptosis (Fig. 5C). These results
indicated that miR-490-3p may inhibit TE1 cell proliferation and
promote ESCC cell apoptosis by inhibiting MAPK1 expression.
Discussion
miRs are small, non-coding RNAs that regulate the
expression of their corresponding proteins mainly at a
post-transcriptional level. Numerous studies have reported that
abnormally expressed miRs in tumors are associated with
proto-oncogenes or tumor suppressor genes expression, and with
patient sensitivity to chemotherapy and radiotherapy (5–7,23). The determination of tumor-specific
miRs and their downstream target genes is therefore crucial to
understand the role of miRs in tumor growth, and to develop novel
targeted therapeutic strategies. Previous studies reported that
miR-490-3p can act as a tumor suppressor gene and regulate various
molecular signaling pathways involved in tumor proliferation,
migration and invasion (13,15). For instance, miR-490-3p is
downregulated in human cancerous gastric tissues, and miR-490-3p
silencing can increase SWI/SNF-related matrix-associated
actin-dependent regulator of chromatin subfamily D member 1
expression and promote gastric cancer (11,24).
Chen et al (10) reported
that miR-490-3p was downregulated in human epithelial ovarian
cancer compared with paracancerous tissues. In addition, miR-490-3p
is lowly expressed in metastatic ovarian epithelial cancer.
Furthermore, miR-490-3p can inhibit tumor cell proliferation by
directly downregulating CDK1 expression (25). miR-490-3p was also decreased in
osteosarcoma tissues, which promoted proliferation, inhibited
apoptosis and cycle progression of osteosarcoma cells by targeting
the high mobility group AT-hook 2 gene (14). However, the functional role of
miR-490-3p in ESCC pathogenesis has been rarely reported.
The present study aimed to explore the role of
miR-490-3p in ESCC pathogenesis and to explore its underlying
mechanism. The results demonstrated that miR-490-3p expression was
significantly decreased in ESCC tissues and in ESCC cell lines.
These results suggested that miR-490-3p may be associated with the
biological function of tumor cells. Based on previous studies TE1
and ECA109 cell lines were selected for miR-490-3p functional
studies (26,27). The results demonstrated that
miR-490-3p depletion increased the proliferation and decreased the
apoptosis of TE1 and ECA109 cell lines.
In order to elucidate the underlying mechanism of
miR-490-3p at regulating tumor cell biological function, one
downstream target of miR-490-3p, MAPK1, was selected. The process
of tumor development is closely associated with tumor cell
proliferation, differentiation, apoptosis and metastasis (28). The molecular mechanism involves many
changes in signaling pathways (28).
The MAPK pathway is a crucial signaling pathway that communicates
signal from a receptor at the cell surface into the cell nucleus
(20). The MAPK pathway can
therefore affect numerous biological processes, including cell
proliferation, differentiation and apoptosis by directly affecting
the transcription and regulation of the associated genes.
Bioinformatics analyses demonstrated that MAPK1 was a target gene
of miR-490-3p. Furthermore, a luciferase reporter gene assay
revealed that miR-490-3p could directly bind to the MAPK1 3′UTR. In
addition, miR-490-3p overexpression or depletion induced a decrease
or increase in MAPK1 mRNA level and protein expression,
respectively. MAPK1 stimulation could also restore the effect of
miR-490-3p on proliferation and apoptosis of ESCC cells, which
suggested that miR-490-3p may affect ESCC cell proliferation and
apoptosis by targeting MAPK1.
In conclusion, the present study demonstrated that
miR-490-3p expression was decreased in ESCC tissues. Furthermore,
miR-490-3p depletion may promote the proliferation and inhibit the
cell apoptosis of ESCC. In addition, MAPK1 may be considered as a
target gene of miR-490-3p. This study provided novel insights into
the role of miR-490-3p in the pathogenesis of ESCC and may
facilitate the development of specific diagnostics or
treatments.
Acknowledgements
Not applicable.
Funding
This study was supported by the Xinjiang Autonomous
Region Natural Fund (grant. no. 2016D01C356).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BZ and AS designed the study and performed the
experiments. BZ, MY and YL collected the data and MY and YL
analyzed the data. BZ and AS wrote the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the ethics committee of
The Third Clinical Medical College (Affiliated Tumor Hospital) of
Xinjiang Medical University. Signed written informed consent was
obtained from all participants before the study.
Patient consent for publication
Patients or their guardians provided written
informed consent for publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mao A: Interventional therapy of
Esophageal Cancer. Gastrointest Tumors. 3:59–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shaib WL, Nammour JP, Gill H, Mody M and
Saba NF: The future prospects of immune therapy in gastric and
esophageal adenocarcinoma. J Clin Med. 5(pii): E1002016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang K, Wu X, Wang J, Lopez J, Zhou W,
Yang L, Wang SE, Raz DJ and Kim JY: Circulating miRNA profile in
esophageal adenocarcinoma. Am J Cancer Res. 6:2713–2721.
2016.PubMed/NCBI
|
|
4
|
Malhotra A, Sharma U, Puhan S, Chandra
Bandari N, Kharab A, Arifa PP, Thakur L, Prakash H, Vasquez KM and
Jain A: Stabilization of miRNAs in esophageal cancer contributes to
radioresistance and limits efficacy of therapy. Biochimie.
156:148–157. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jamali L, Tofigh R, Tutunchi S, Panahi G,
Borhani F, Akhavan S, Nourmohammadi P, Ghaderian SMH, Rasouli M and
Mirzaei H: Circulating microRNAs as diagnostic and therapeutic
biomarkers in gastric and esophageal cancer. J Cell Physiol.
233:8538–8550. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shenouda SK and Alahari SK: MicroRNA
function in cancer: Oncogene or a tumor suppressor? Cancer
Metastasis Rev. 28:369–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou HQ, Chen QC, Qiu ZT, Tan WL, Mo CQ
and Gao SW: Integrative microRNA-mRNA and protein-protein
interaction analysis in pancreatic neuroendocrine tumors. Eur Rev
Med Pharmacol Sci. 20:2842–2852. 2016.PubMed/NCBI
|
|
8
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xuan Y, Yang H, Zhao L, Lau WB, Lau B, Ren
N, Hu Y, Yi T, Zhao X, Zhou S and Wei Y: MicroRNAs in colorectal
cancer: Small molecules with big functions. Cancer Lett.
360:89–105. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen S, Chen X, Xiu YL, Sun KX and Zhao Y:
MicroRNA-490-3P targets CDK1 and inhibits ovarian epithelial
carcinoma tumorigenesis and progression. Cancer Lett. 362:122–130.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen J, Xiao Z, Wu WK, Wang MH, To KF,
Chen Y, Yang W, Li MS, Shin VY, Tong JH, et al: Epigenetic
silencing of miR-490-3p reactivates the chromatin remodeler SMARCD1
to promote Helicobacter pylori-induced gastric carcinogenesis.
Cancer Res. 75:754–765. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li W, Guo F, Wang P, Hong S and Zhang C:
MiR-221/222 confers radioresistance in glioblastoma cells through
activating Akt independent of PTEN status. Curr Mol Med.
14:185–195. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang LY, Liu M, Li X and Tang H:
MiR-490-3p modulates cell growth and epithelial to mesenchymal
transition of hepatocellular carcinoma cells by targeting
endoplasmic reticulum-Golgi intermediate compartment protein 3
(ERGIC3). J Biol Chem. 288:4035–4047. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu W, Xu G, Liu H and Li T:
MicroRNA-490-3p regulates cell proliferation and apoptosis by
targeting HMGA2 in osteosarcoma. FEBS Lett. 589:3148–3153. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tian J, Xu YY, Li L and Hao Q: MiR-490-3p
sensitizes ovarian cancer cells to cisplatin by directly targeting
ABCC2. Am J Transl Res. 9:1127–1138. 2017.PubMed/NCBI
|
|
16
|
Li S, Xu X, Xu X, Hu Z, Wu J, Zhu Y, Chen
H, Mao Y, Lin Y, Luo J, et al: MicroRNA-490-5p inhibits
proliferation of bladder cancer by targeting c-Fos. Biochem Biophys
Res Commun. 441:976–981. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rezatabar S, Karimian A, Rameshknia V,
Parsian H, Majidinia M, Kopi TA, Bishayee A, Sadeghinia A, Yousefi
M, Monirialamdari M and Yousefi B: RAS/MAPK signaling functions in
oxidative stress, DNA damage response and cancer progression. J
Cell Physiol. Feb 27–2019;doi: 10.1002/jcp.28334 (Epub ahead of
print). PubMed/NCBI
|
|
18
|
Krysan PJ and Colcombet J: Cellular
complexity in MAPK signaling in plants: Questions and emerging
tools to answer them. Front Plant Sci. 9:16742018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chatterjee N, Das S, Bose D, Banerjee S,
Jha T and Das Saha K: Lipid from infective L. Donovani regulates
acute myeloid cell growth via mitochondria dependent MAPK pathway.
PLoS One. 10:e1205092015. View Article : Google Scholar
|
|
20
|
Chen SX, Zhao F and Huang XJ: MAPK
signaling pathway and erectile dysfunction. Zhonghua Nan Ke Xue.
24:442–446. 2018.(In Chinese). PubMed/NCBI
|
|
21
|
Furukawa T: Impacts of activation of the
mitogen-activated protein kinase pathway in pancreatic cancer.
Front Oncol. 5:232015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hummel R, Sie C, Watson DI, Wang T, Ansar
A, Michael MZ, Van der Hoek M, Haier J and Hussey DJ: MicroRNA
signatures in chemotherapy resistant esophageal cancer cell lines.
World J Gastroenterol. 20:14904–14912. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shah AA, Leidinger P, Backes C, Keller A,
Karpinski P, Sasiadek MM, Blin N and Meese E: A set of specific
miRNAs is connected with murine and human gastric cancer. Genes
Chromosomes Cancer. 52:237–249. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang LL, Sun KX, Wu DD, Xiu YL, Chen X,
Chen S, Zong ZH, Sang XB, Liu Y and Zhao Y: DLEU1 contributes to
ovarian carcinoma tumourigenesis and development by interacting
with miR-490-3p and altering CDK1 expression. J Cell Mol Med.
21:3055–3065. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu J, Pan X and Hu Z: MiR-502 mediates
esophageal cancer cell TE1 proliferation by promoting AKT
phosphorylation. Biochem Biophys Res Commun. 501:119–123. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liang N, Song X, Xie J, Xu D, Liu F, Yu X,
Tian Y, Liu Z, Qiao L and Zhang J: Effect of galectin-3 on the
behavior of Eca109 human esophageal cancer cells. Mol Med Rep.
11:896–902. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim EK and Choi EJ: Compromised MAPK
signaling in human diseases: An update. Arch Toxicol. 89:867–882.
2015. View Article : Google Scholar : PubMed/NCBI
|