Introduction
Uterine sarcomas represent ~8% of uterine malignant
tumors and entail a high mortality rate, with an overall 5-year
survival rate of ~30% (1). Uterine
leiomyosarcoma (ULMS) is the most common uterine sarcoma,
characterized by poor prognosis, early metastasis and high rate of
recurrence (2–4). ULMS occurs mostly in women over the age
of 40, and its symptoms include abnormal vaginal bleeding, palpable
pelvic mass and pelvic pain (5).
These features are similar to the symptoms of uterine leiomyoma
(ULM) and therefore it is difficult to distinguish ULMS from ULM
prior to surgery. Currently, there is still no optimal treatment
for ULMS (5,6). There are few studies on ULMS and little
is known about its associated oncogenic pathways.
The human CKS family consists of two members, CKS1
and CKS2, which share >80% sequence identity (7). CKS1 is a specific cofactor that is
necessary for the degradation and ubiquitination of p27 by the
Skp1-Cullin-F-box protein complex of S-phase kinase-associated
protein 2 (8). CKS2 is also
important during early embryonic development, for the process of
somatic cell division (9). In
addition, it has been shown to be essential for the first
metaphase/anaphase transition of mammalian meiosis (10). Several reports have indicated that
CKS2 was upregulated in a number of types of tumors, including
breast cancer, esophageal carcinoma, gastric cancer, colorectal
cancer, hepatocellular carcinoma, cholangiocarcinoma and bladder
cancer (11–17). However, the underlying cellular
functions of CKS2 and the associated mechanism involved in its
carcinogenicity remain unclear.
To identify a candidate gene that may contribute to
the progression of ULMS, a search in the Gene Expression Omnibus
(GEO) database was conducted. Expression arrays from the GEO
datasets (GSE64763, GSE764 and GSE36610) showed distinct levels of
expression in ULMS and ULM of genes such as cyclin-dependent kinase
subunit (CKS)2, thymidylate synthase and putative tenascin-XA.
Since CKS2 was found to be expressed in several tumors and little
was known about its role in ULMS, further analyses were conducted.
The present study investigated CKS2 expression in ULM/ULMS and
determined the role of CKS2 in the development of ULMS.
Materials and methods
Clinical tissue samples
The specimens of 38 cases with ULMS and 38 cases
with ULM were collected between January 2005 and October 2015 at Qi
Lu Hospital of Shandong University (Jinan, China). At the beginning
of the study, 45 cases with ULMS were recruited and followed-up
over the telephone. Seven cases were lost during the follow-up and
38 remained, with the complete clinical and prognostic information.
By the end of the follow-up (March 2016), 19 patients with ULMS
were deceased. The median age of the patients with ULMS was 45
years (range, 29–76 years). The median age of the patients with ULM
was 38 years (range, 25–65 years). None of these patients had
received pre-operative chemotherapy or radiotherapy. All patients
provided informed consent, and the study was approved by the Ethics
Committee of Shandong University (approval no. 201302036). All
samples were assessed by two well-trained pathologists. The
diagnosis was confirmed according to the 2003 World Health
Organization criteria (18), and
pathological records were reviewed according to the 2009
International Federation of Obstetrics and Gynecology (FIGO)
staging for sarcomas (19).
Immunohistochemistry (IHC)
Tissues were fixed with formalin (10%, pH 7.2) at
room temperature for 24 h and then embedded in paraffin. IHC
analysis was performed on these formalin-fixed, paraffin-embedded
specimens (4-µm sections). The sections were rehydrated through
alcohol gradient as follows: Anhydrous ethanol I for 5 min;
anhydrous ethanol II for 5 min; 95% ethanol for 5 min; 85% ethanol
for 5 min; and 75% ethanol for 5 min. The sections were submerged
in sodium citrate buffer at 100°C for 5 min for antigenic
retrieval. The endogenous peroxidase activity was blocked with 3%
hydrogen peroxide for 10 min at room temperature. After blocking,
the sections were incubated with anti-CKS2 primary antibody (1:100;
cat. no. ab155078; Abcam) overnight at 4°C. A PV-9000 2-step
plus® Poly-HRP Anti-Mouse/Rabbit IgG Detection System
(Zhongshan Golden Bridge Biotechnology Company) was used for
immunohistochemical assay according to the manufacturer's
instructions. PBS was used as the negative control.
Values were assigned to the samples according to the
percentage of positive cells: 0, ≤5; 1, 6–25; 2, 26–50; 3, 51–75;
and 4, >75%. Similarly, the staining intensity scores were
assigned as follows: 0, no staining; 1, weak; 2, moderate; and 3,
strong staining. The product of the two integers was used to divide
the participants into two groups. Participants with a product ≥4
were designated to the ‘high expression’ group, and those with a
product <4 were designated to the ‘low expression’ group. The
images were captured with an Olympus BX53 optical microscope
(Olympus Corporation) at ×400 magnification. Two senior
pathologists independently assessed these scores.
Cell culture and transfection
Human ULMS SK-UT-1 and SK-UT-1B cell lines were
kindly provided by Dr Kong Beihua of the Qi Lu Hospital of Shandong
University. These cell lines originate from the uterus and have
been reported to differ markedly in terms of morphology and
karyotype (20). The cells were
maintained in minimal essential medium (MEM; cat. no. 11095080;
Gibco; Thermo Fisher Scientific, Inc.), supplemented with 10% fetal
bovine serum (FBS; Biological Industries), 1X solution of
non-essential amino acids (Thermo Fisher Scientific, Inc.), 1 mM
sodium pyruvate and 100 U/ml penicillin and streptomycin in 5%
CO2 at 37°C.
Small interfering RNA (siRNA) targeting CKS2
(si-CKS2 sense, 5′-GGAGACUUGGUGUCCAACATT-3′; and si-CKS2 antisense,
5′-UGUUGGACACCAAGUCUCCTC-3′) and negative control siRNA (si-Ctrl
sense, 5′-UUCUCCGAACGUGUCACGUTT-3′; and si-Ctrl antisense,
5′-ACGUGACACGUUCGGAGAATT-3′) were obtained from Shanghai GenePharma
Co., Ltd. The siRNA oligomer was diluted in Opti-MEM (Thermo Fisher
Scientific, Inc.). The diluted siRNA oligomer was mixed with
diluted Lipofectamine® 2000 (Thermo Fisher Scientific,
Inc.) and incubated for 20 min at room temperature to allow the
siRNA-lipid complexes to form. The complexes were then added to
each well of 6-well plates, giving a final concentration of siRNA
of 50 pmol/ml. After incubation in 5% CO2 at 37°C for
4–6 h, the cells were washed three times with Opti-MEM, and
returned for incubation for the following procedures.
Cell Counting Kit-8 (CCK-8) cell
proliferation assay
Cell proliferation rates were measured using the
CCK-8 (Beyotime Institute of Biotechnology). Following the
transfection of cells with si-CKS2 and si-Ctrl (after 24 h), the
cells were seeded at a density of 3×103 per well in a
96-well plate. The cells were incubated for 24, 48, 72 or 96 h and
10 µl CCK-8 was subsequently added into each well. The absorbance
was measured at 450 nm by a microplate reader (Bio-Rad
Laboratories, Inc.), following incubation in 5% CO2 at
37°C for 2 h.
Colony formation
Following 24 h from transfection with si-CKS2 or
si-Ctrl, SK-UT-1 or SK-UT-1B cells (500 cells/well) were seeded in
6-well plates and incubated for 14 days to form colonies. The
colonies were fixed with 4% paraformaldehyde at room temperature
for 20 min and stained with 0.1% crystal violet at room temperature
for 20 min. Images of colonies were captured by a digital camera
and the number of foci containing >50 cells was counted. The
mean number of foci formed by si-Ctrl was normalized to 100% and
compared with the number of si-CKS2 colonies was compared with
it.
Cell migration and invasion
assays
The migration assays were carried out using
Transwell inserts (Corning Inc.), according to the manufacturer's
instructions. For the invasion assays, the membranes of Transwell
inserts were coated with Matrigel matrix (BD Biosciences). The
Matrigel matrix was diluted by Opti-MEM (1:3; Thermo Fisher
Scientific, Inc.) and incubated at 37°C for 3 h to form the
Matrigel coating. Following 24 h transfection with si-RNA, cells
(1×105) in 100 µl serum-free medium were placed in the
upper chamber, and the lower chamber was filled with 700 µl culture
medium with 10% FBS. The cells were incubated for 24 h at 37°C. The
non-migrated/-invaded cells that remained on the upper chamber were
removed. The cells on the lower side of the Transwell membrane were
fixed with 4% paraformaldehyde at room temperature for 20 min and
stained with 0.1% crystal violet at room temperature for 20 min.
The number of migrated and invaded cells was counted under a light
microscope at 200× magnification in six random fields.
Cell cycle
The effects of silencing CKS2 on cell cycle
progression were assessed using propidium iodide staining and
analyzed by flow cytometry. Cells were seeded in 6-well plates
(2×105 per well) and transfected with si-CKS2 and
si-Ctrl. After 48 h, cells were washed with PBS, harvested and
fixed in 70% ethanol at 4°C overnight. Cells were treated with
DNase-free RNase and stained with propidium iodide (400 µl from 50
µg/ml stock solution) at 4°C for 30 min, and the cell cycle was
analyzed by flow cytometry based on the DNA content of cell
populations. Finally, the distribution of cells within the
G0/G1, S, and G2/M phases was
measured by using ModFit LT 5.0 (Verity Software House, Inc.).
Western blot analysis
Following 48 h transfection with si-RNA, total
protein was extracted from the sample with RIPA lysis buffer and
protein concentrations were detected by the BCA protein assay kit
(both from Beyotime Institute of Biotechnology). Equal amounts of
protein (30 µg) were applied to 15% sodium dodecyl sulfate
polyacrylamide gel electrophoresis gel and electroblotted to a
0.22-µm polyvinylidene difluoride membrane. The membranes were
blocked by QuickBlock™ Blocking Buffer for Western Blot (Beyotime
Institute of Biotechnology) at room temperature for 10 min. After
the blocking step, the membranes were incubated with rabbit
anti-human CKS2 monoclonal antibody (1:1,000; cat. no. ab155078;
Abcam), anti-beta actin antibody (1:1,000; cat. no. ab8227; Abcam),
anti-claudin-1 antibody (1:1,000; cat. no. ab15098; Abcam),
anti-p38 antibody (1:1,000; cat. no. ab195049; Abcam) and anti-bax
antibody (1:1,000; cat. no. ab32503; Abcam) at 4°C overnight and
then with horseradish peroxidase-labeled goat anti-rabbit IgG
(1:5,000; cat. no. ab 6721; Abcam) at 37°C for 30 min. Finally, the
membranes were analyzed using an electrochemiluminescence system
(EMD Millipore).
Reverse transcription-quantitative PCR
(qPCR)
Following 24 h transfection with si-RNA, total RNA
was extracted from cells using TRIzol reagent (Thermo Fisher
Scientific, Inc.) and was then transcribed into cDNA with ReverTra
Ace qPCR RT kit (cat. no. FSQ-101; Toyobo Life Science). The
thermocycling conditions were as follows: 37°C for 15 min and 98°C
for 5 min. Quantification of the cDNA template was performed on a
Real-time Thermo Cycler (cat. no. C1000; Bio-Rad Laboratories,
Inc.), using SYBR-Green Real-time PCR Master mix (cat. no. QPK-201;
Toyobo Life Science). The thermocycling conditions were as follows:
Initial denaturation at 95°C for 10 min, and 40 cycles of
denaturation at 95°C for 10 sec, annealing at 60°C for 1 min, and
extension at 65°C for 5 sec. The primers were as follows: CKS2
forward, 5′-TTCGACGAACACTACGAGTACC-3′; CKS2 reverse,
5′-GGACACCAAGTCTCCTCCAC-3′; GAPDH forward:
5′-TGAAGGTCGGAGTCAACGGA-3′; and GAPDH reverse,
5′-CCTGGAAGATGGTGATGGGAT-3′. The dissociation curve analysis was
performed in order to guarantee the specificity of the qPCR. The
relative expression level of CKS2 was normalized to GAPDH and
analyzed by the 2−ΔΔCt method (21).
Statistical analysis
Data analysis was carried out using the SPSS 22.0
software (IBM Corp.). The association between CKS2 expression and
clinicopathological factors was analyzed by Pearson's χ2
test. The overall survival (OS) rate was analyzed according to the
Kaplan-Meier method and the generalized log-rank test was applied
to analyze the survival curves. Prognostic factors were evaluated
by univariate and multivariate analyses (Cox proportional hazard
regression model). Other data are expressed as the mean ± SD and
analyzed with Student's t-test. All assays were performed in
triplicate. P<0.05 was considered to indicate a statistically
significant difference.
Results
CKS2 is overexpressed in ULMS and
predicts poor prognosis
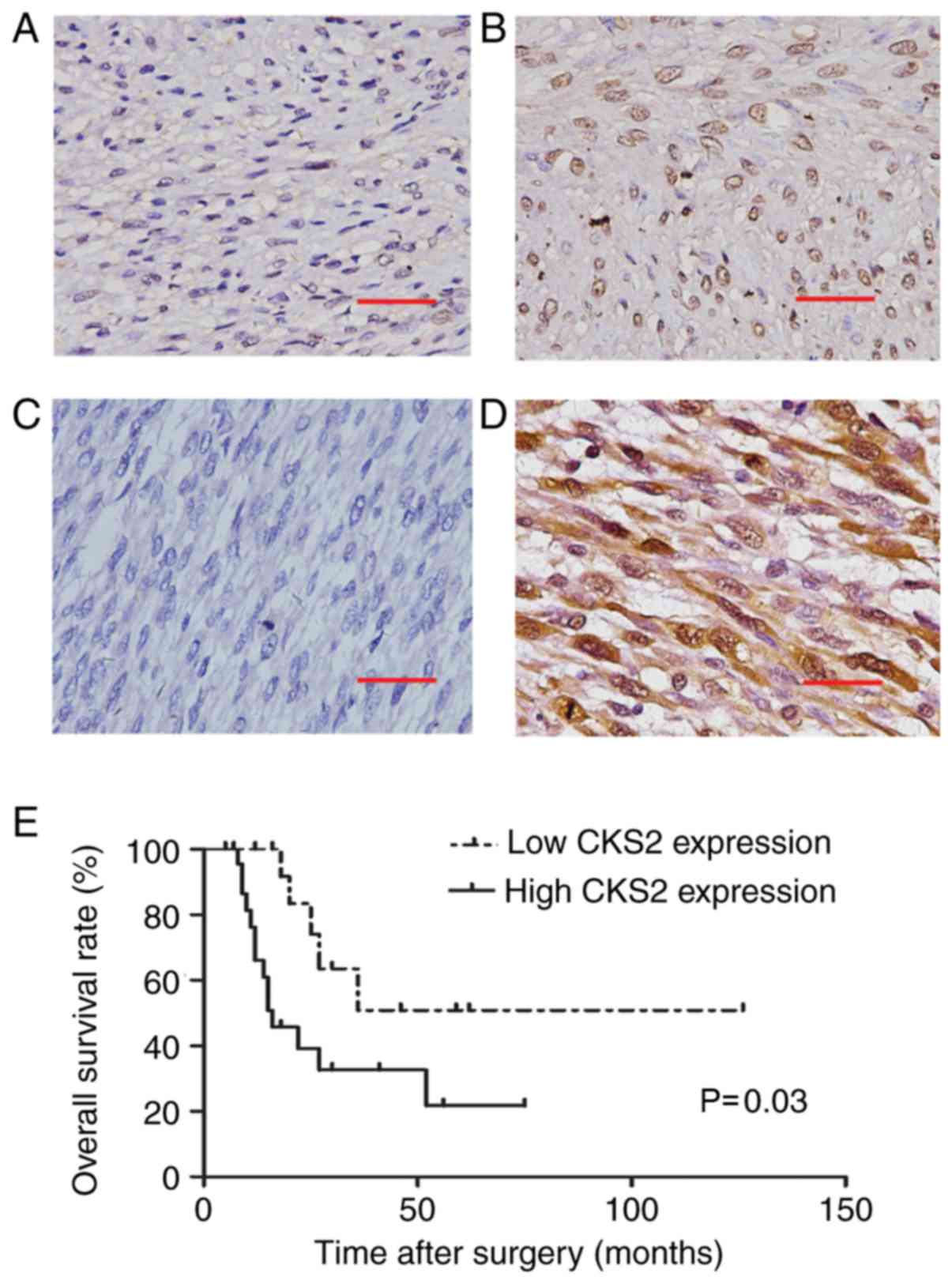
To determine the expression of CKS2 in ULMS, its
protein expression was analyzed by IHC. The results indicated that
the staining of CKS2 was significantly stronger in ULMS tissues
than that in ULM. CKS2 expression was high in 63.2% (24 of 38
cases) of the ULMS tissues, whereas only 18.4% (7 of 38 cases) of
ULM tissues had high CKS2 expression (P<0.001; Table I; Fig.
1A-D). The expression of CKS2 was mainly located in the
nucleus.
 | Table I.Expression of CKS2 in ULMS and ULM
tissues. |
Table I.
Expression of CKS2 in ULMS and ULM
tissues.
|
| CKS2
expression |
|
|---|
|
|
|
|
|---|
| Tissue | High, n | Low, n | P-value |
|---|
| Tissue type |
|
| <0.001 |
|
ULMS | 24 | 14 |
|
|
ULM | 7 | 31 |
|
To investigate the potential roles of CKS2 in ULMS,
patients were divided into two groups according to CKS2 expression.
The statistical analysis demonstrated that high expression of CKS2
in patients was associated with larger tumor size and low
expression of progesterone receptor (PR); whereas no association of
CKS2 expression was observed with other clinicopathological
features, such as age, FIGO stage and estrogen receptor expression
(Table II).
| Table II.Association between CKS2 expression
and clinical features of uterine leiomyosarcoma. |
Table II.
Association between CKS2 expression
and clinical features of uterine leiomyosarcoma.
|
|
| CKS2
expression |
|
|---|
|
|
|
|
|
|---|
| Parameter | Total cases, n | Low (n=14), n | High (n=24), n | P-value |
|---|
| Age, years |
|
|
| 0.076 |
|
≤45 | 20 | 10 | 10 |
|
|
>45 | 18 | 4 | 14 |
|
| Tumor size, cm |
|
|
| 0.014a |
| ≤5 | 12 | 8 | 4 |
|
|
>5 | 26 | 6 | 20 |
|
| FIGO stage |
|
|
| 0.826 |
| I | 26 | 11 | 15 |
|
| II | 3 | 1 | 2 |
|
|
III | 5 | 1 | 4 |
|
| IV | 4 | 1 | 3 |
|
| ER expression |
|
|
| 0.391 |
|
Positive | 13 | 6 | 7 |
|
|
Negative | 25 | 8 | 17 |
|
| PR expression |
|
|
| 0.048a |
|
Positive | 14 | 8 | 6 |
|
|
Negative | 24 | 6 | 18 |
|
The association between CKS2 expression and
prognosis was determined by analyzing the OS rates of 38 patients
with ULMS. As shown in Fig. 1E,
patients in the CKS2 high-expression group had a markedly poorer
survival rate than those in the low-expression group (P=0.03).
Univariate analysis revealed that CKS2 expression (P=0.040), tumor
size (P=0.045) and FIGO stage (P=0.001) were significant risk
factors for OS. In multivariate analysis, CKS2 expression (P=0.036)
and FIGO stage (P=0.001) were independent predictors of OS with
ULMS (Table III). Taken together,
the above results indicate that CKS2 expression is upregulated in
ULMS, which predicts poor prognosis in ULMS.
| Table III.Univariate and multivariate analysis
for prognostic factors of survival of uterine leiomyosarcoma. |
Table III.
Univariate and multivariate analysis
for prognostic factors of survival of uterine leiomyosarcoma.
|
|
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|
|
|---|
| Parameter | Category | Cases, n | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| CKS2
expression | Low vs. high | 14 vs. 24 | 2.940
(1.052–8.012) | 0.040a | 3.124
(1.079–9.048) | 0.036a |
| Age, years | ≤45 vs. >45 | 20 vs. 18 | 1.144
(0.463–2.825) | 0.771 |
|
|
| Tumor size, cm | ≤5 vs. >5 | 12 vs. 26 | 3.572
(1.028–12.414) | 0.045a |
|
|
| FIGO stage | I vs. II vs. III
vs. IV | 26 vs. 3 vs. 5 vs.
4 | 1.949
(1.312–2.896) | 0.001b | 2.087
(1.341–3.247) | 0.001b |
| ER expression | Positive vs.
negative | 13 vs. 25 | 0.374
(0.122–1.143) | 0.084 |
|
|
| PR expression | Positive vs.
negative | 14 vs. 24 | 0.407
(0.135–1.231) | 0.111 |
|
|
Silencing of CKS2 inhibits cell
proliferation
CKS2 may contribute to the malignant behaviors of
ULMS cells, because an association was found between CKS2
expression and tumor size. To determine the role of CKS2 in ULMS,
two cell lines (SK-UT-1 and SK-UT-1B), were selected for further
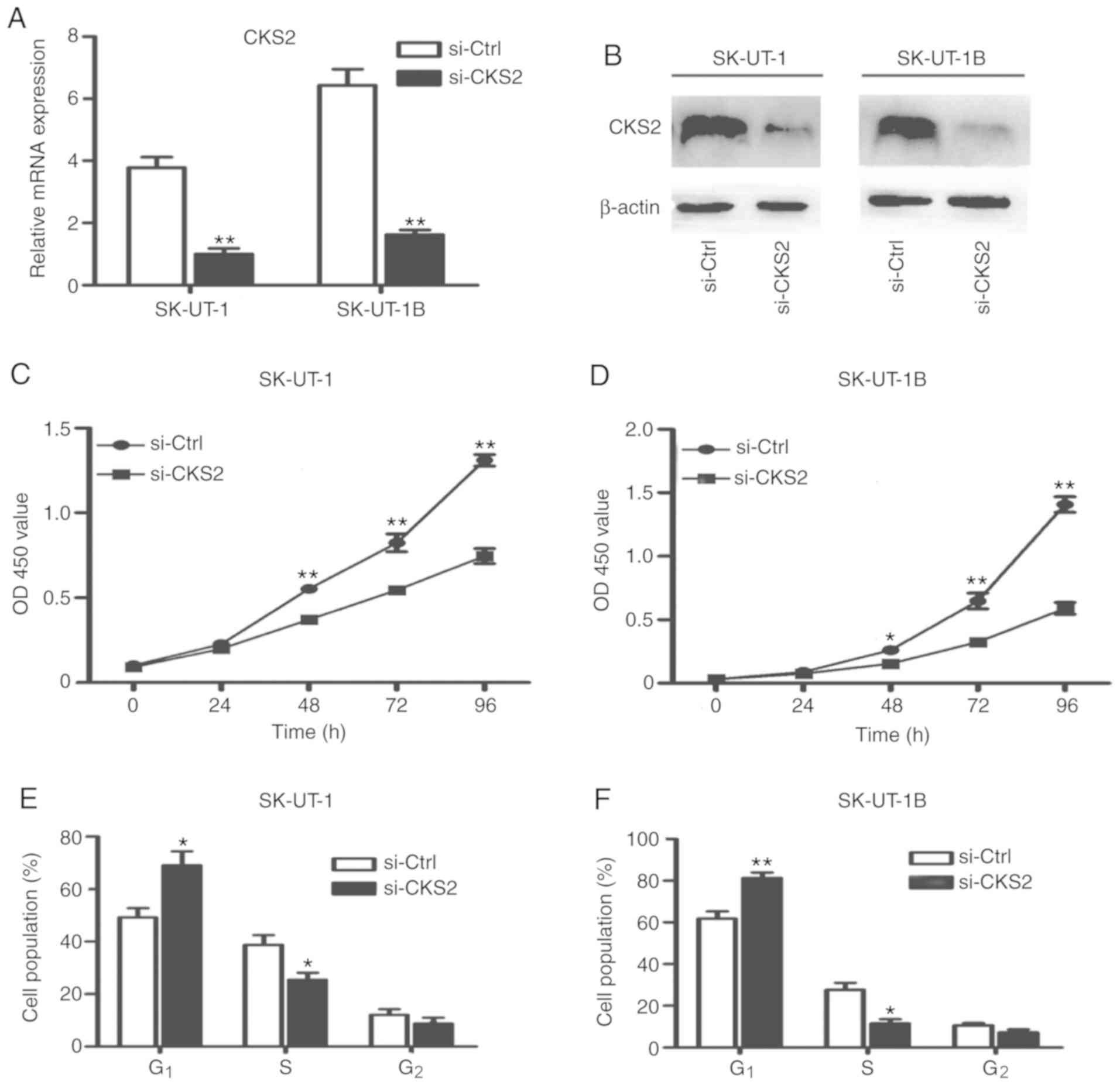
investigation. As shown in Fig. 2A and
B, the expression of CKS2 significantly decreased, following
the transfection with si-CKS2 (both P<0.01). The CCK-8 assay was
used to determine the change in cell viability, as shown in
Fig. 2C and D, where knockdown of
CKS2 decreased the viability of SK-UT-1 and SK-UT-1B cells. These
results indicate that silencing of CKS2 may inhibit cell
proliferation in ULMS cell lines.
Silencing of CKS2 inhibits cell cycle
progression
As CKS2 is a member of the cell cycle-dependent
protein kinase subunits family, cell cycle analysis was performed.
The effect of CKS2 knockdown on the distribution of cells at the
different phases of the cell cycle was observed. The silencing of
CKS2 increased the population of cells at the G1 phase
and decreased the cells at S phase (Fig.
2E and F). These results suggest that the silencing of CKS2 can
cause cell cycle arrest at the G1/S transition phase in
ULMS cells.
Silencing of CKS2 inhibits ULMS cell
colony formation
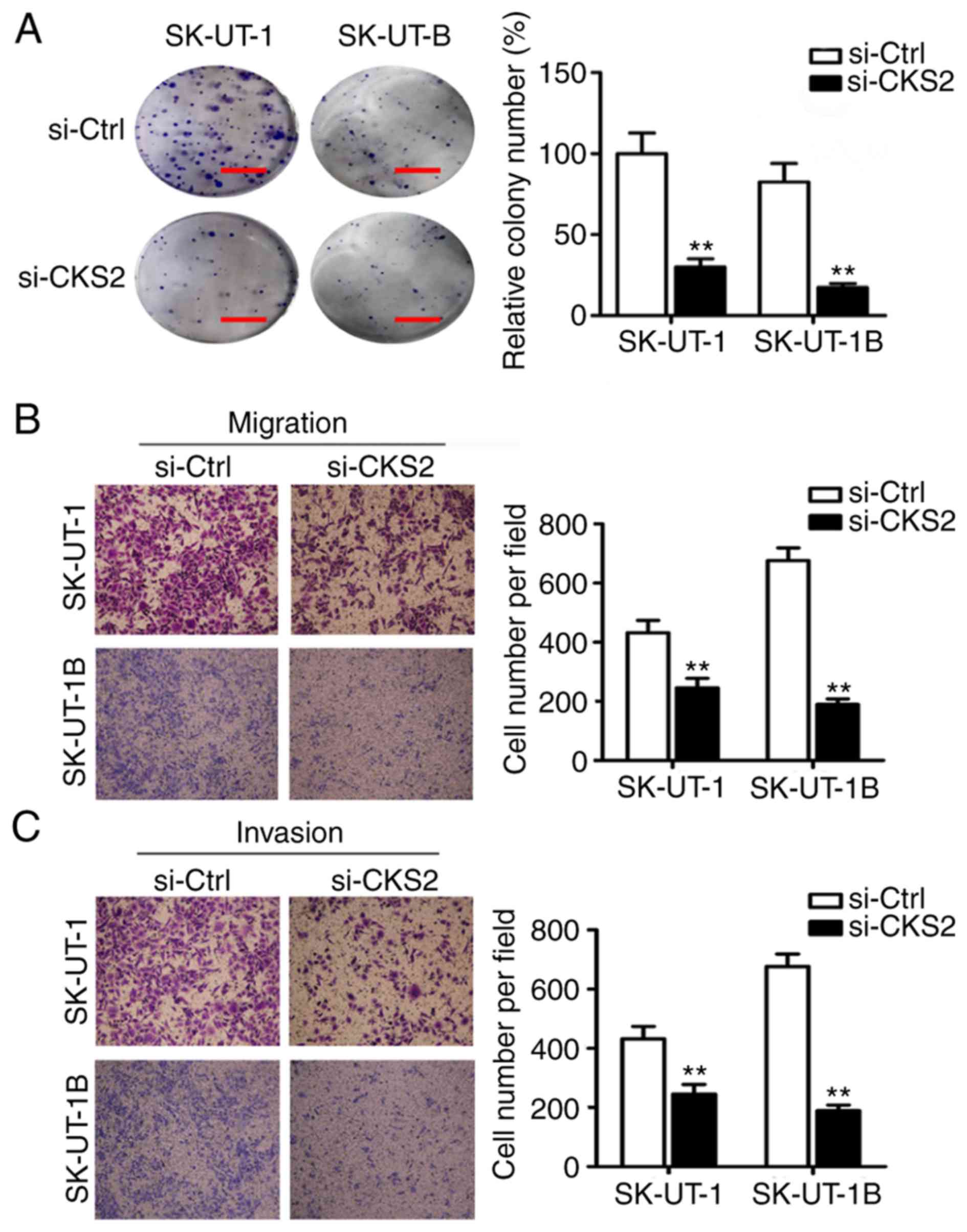
The role of CKS2 in cell transformation was
subsequently examined by colony-formation assay, to analyze the
oncogenic potential of CKS2 in vitro. Both of the ULMS cell
lines showed diminished ability to form foci when CKS2 was
downregulated by si-CKS2 (Fig. 3A).
This indicated that CKS2 contributes to promoting the clonogenic
survival of ULMS cells.
Silencing of CKS2 inhibits cell
migration and invasion
To further determine the role of CKS2 in the
metastasis of ULMS, the migratory and invasive potential of SK-UT-1
and SK-UT-1B cells was assessed. Cells were treated with si-CKS2 or
si-Ctrl and investigated in in vitro migration and Matrigel
invasion assays. As shown in Fig. 3B and
C, the migrating and invading cells in the si-CKS2 group were
significantly decreased compared with those in the si-Ctrl group in
both ULMS cell lines. These results indicate that CKS2 affects ULMS
cell migration and invasion.
Furthermore, the effect of si-CKS2 on the apoptosis
of ULMS cells was investigated, by measuring the apoptotic rates of
si-Ctrl and si-CKS2 by Annexin V-FITC/PI double-staining assay.
However, the apoptotic rate of si-CKS2 cells (8.2%) was not
significantly higher than si-Ctrl cells (5.9%). These results
suggest that CKS2 may have no effect on ULMS cell apoptosis (data
not shown).
To investigate the molecular mechanisms of the
effect of CKS2 in ULMS, western blot analysis was performed to
measure changes in the levels of claudin-1, p-p38 and Bax, which
were reported to be modulated by CKS2 in other tumors (13,14,16,22).
However, there were no significant changes following the silencing
of CKS2 in ULMS cells (data not shown). These results suggest that
CKS2 may exert its oncogenic function in ULMS through different
pathways and further research is required.
Discussion
ULMS, the most common uterine sarcoma, has a poor
outcome and a high recurrence rate, according to a survey from 2009
in Norway (4). Furthermore, it
remains a clinical challenge to distinguish benign ULM from ULMS
(23). Previous studies have shown
that ULMS is characteristic of excessive activation of cell
proliferation pathways and loss of chromosomal fragments that
contain tumor suppressor genes (24,25). The
Cancer Genome Atlas found that the most commonly mutated genes of
ULMS were TP53, retinoblastoma transcriptional corepressor
1, and alpha thalassemia/mental retardation syndrome X-linked
(26). However, the exact
pathophysiological mechanism of ULMS is still unclear.
CKS2, a member of the CKS family, was reported to be
highly expressed in various malignant tumors, including breast
cancer, esophageal carcinoma and gastric cancer (11–13). It
was found to regulate the cell cycle and promote cancer invasion
and metastasis (27). Nevertheless,
the role of CKS2 in ULMS is not clearly understood.
In the present study, it was demonstrated that CKS2
was upregulated in ULMS tissues compared with ULM tissues. This was
consistent with the results reported in studies by Wang et
al (12) and Shen et al
(15), where CKS2 was found to be
upregulated in esophageal carcinoma and hepatocellular carcinoma.
Moreover, Tanaka et al (13)
also found that overexpression of CKS2 was correlated with tumor
size, serosal invasion, lymph node metastasis and distant
metastasis in gastric cancer. Furthermore, Wang et al
(11) found that CKS2 was
upregulated in breast cancer and associated with large tumor size,
poor tumor differentiation and survival. In the present study, in
addition to the association with large tumor size and poor
prognosis in ULMS, CKS2 overexpression was also associated with
lack of PR expression. Due to the low incidence rate of ULMS (only
0.4 out of 100,000 women each year, according to a survey from 2012
in Nordic countries) (28), the
sample size is a limitation of the present study and will be
addressed in future studies.
To further determine the role of CKS2 in ULMS, its
expression was silenced using siRNA. The CCK-8 and colony-formation
assays demonstrated that the silencing of CKS2 decreased cell
viability and weakened the colony-forming ability of ULMS cells.
These results indicate that CKS2 promotes cell proliferation and
the clonogenic survival of ULMS cells.
Flow cytometry analysis indicated that the silencing
of CKS2 in ULMS cells increased the cells at the G1
phase and decreased the cells at the S phase. Yu et al
(14) demonstrated that
downregulation of CKS2 resulted in cell cycle arrest in
G1/S transition in colorectal cancer, whereas Shen et
al (16) found that silencing of
CKS2 increased the number of cells at the G2/M phase and
decreased the number at G1 and S phase in
cholangiocarcinoma. CKS2 was reported to be associated with somatic
cell division and meiosis (9,10);
however, the effect of CKS2 on the cell cycle is still unclear. In
the present study, the results demonstrated that CKS2 serves as a
cell cycle checkpoint protein for G1/S transition, which
may be how CKS2 contributes to ULMS progression.
CKS2 does not only regulate the cell cycle in ULMS
cells, but also has a notable effect on the capacity for migration
and invasion. Specifically, the results showed that the knockdown
of CKS2 inhibited ULMS cell migration and invasion in vitro.
Similarly, Tanaka et al (13)
and Yu et al (14)
demonstrated that downregulation of CKS2 weakened the capacity of
migration and invasion in gastric cancer and colorectal cancer.
These findings indicate that CKS2 promotes the migration and
invasion of ULMS cells, and therefore promotes metastasis. This may
explain the occurrence of early metastasis in patients with ULMS.
However, further study is required to elucidate the potential
mechanisms.
In conclusion, this is the first study to
investigate the function of CKS2 in ULMS. The findings of this
study indicate that CKS2 plays an important role in ULMS and may
serve as a useful marker for the differential diagnosis of ULMS and
ULM. It is postulated that CKS2 exerts its oncogenic effects by
promoting G1/S transition, proliferation, colony
formation, migration and invasion in ULMS cells. It may also act as
an independent prognostic factor in patients with ULMS and serve as
a novel target for ULMS therapy.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81372810).
Availability of data and materials
The datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
TZ conceived the experiments. YD and QH carried out
the experiments and drafted the manuscript. SM, HL and FY collected
the clinical data and tissue samples. JW and SG performed the
statistical analysis. XJ and HX helped performing the experiments
and writing the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Shandong University (approval no. 201302036). All
patients provided informed written consent prior to the
investigation.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CKS2
|
cyclin-dependent kinase subunit 2
|
|
ULMS
|
uterine leiomyosarcoma
|
|
ULM
|
uterine leiomyoma
|
|
siRNA
|
small interfering RNA
|
|
PR
|
progesterone receptor
|
References
|
1
|
Hensley ML, Barrette BA, Baumann K,
Gaffney D, Hamilton AL, Kim JW, Maenpaa JU, Pautier P, Siddiqui NA,
Westermann AM and Ray-Coquard I: Gynecologic cancer intergroup
(GCIG) consensus review: Uterine and ovarian leiomyosarcomas. Int J
Gynecol Cancer. 24 (Suppl 3):S61–S66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Giuntoli RL II, Metzinger DS, DiMarco CS,
Cha SS, Sloan JA, Keeney GL and Gostout BS: Retrospective review of
208 patients with leiomyosarcoma of the uterus: Prognostic
indicators, surgical management, and adjuvant therapy. Gynecol
Oncol. 89:460–469. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kapp DS, Shin JY and Chan JK: Prognostic
factors and survival in 1,396 patients with uterine
leiomyosarcomas: Emphasis on impact of lymphadenectomy and
oophorectomy. Cancer. 112:820–830. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Abeler VM, Røyne O, Thoresen S, Danielsen
HE, Nesland JM and Kristensen GB: Uterine sarcomas in Norway. A
histopathological and prognostic survey of a total population from
1970 to 2000 including 419 patients. Histopathology. 54:355–364.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gockley AA, Rauh-Hain JA and del Carmen
MG: Uterine leiomyosarcoma: A review article. Int J Gynecol Cancer.
24:1538–1542. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miller H, Ike C, Parma J, Masand RP, Mach
CM and Anderson ML: Molecular targets and emerging therapeutic
options for uterine leiomyosarcoma. Sarcoma. 2016:70181062016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Richardson HE, Stueland CS, Thomas J,
Russell P and Reed SI: Human cDNAs encoding homologs of the small
p34Cdc28/Cdc2-associated protein of Saccharomyces cerevisiae
and Schizosaccharomyces pombe. Genes Dev. 4:1332–1344. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ganoth D, Bornstein G, Ko TK, Larsen B,
Tyers M, Pagano M and Hershko A: The cell-cycle regulatory protein
Cks1 is required for SCF(Skp2)-mediated ubiquitinylation of p27.
Nat Cell Biol. 3:321–324. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Martinsson-Ahlzén HS, Liberal V,
Grünenfelder B, Chaves SR, Spruck CH and Reed SI: Cyclin-dependent
kinase-associated proteins Cks1 and Cks2 are essential during early
embryogenesis and for cell cycle progression in somatic cells. Mol
Cell Biol. 28:5698–5709. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Spruck CH, de Miguel MP, Smith AP, Ryan A,
Stein P, Schultz RM, Lincoln AJ, Donovan PJ and Reed SI:
Requirement of cks2 for the first metaphase/anaphase transition of
mammalian meiosis. Science. 300:647–650. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang J, Xu L, Liu Y, Chen J, Jiang H, Yang
S and Tan H: Expression of cyclin kinase subunit 2 in human breast
cancer and its prognostic significance. Int J Clin Exp Pathol.
7:8593–8601. 2014.PubMed/NCBI
|
|
12
|
Wang JJ, Fang ZX, Ye HM, You P, Cai MJ,
Duan HB, Wang F and Zhang ZY: Clinical significance of
overexpressed cyclin-dependent kinase subunits 1 and 2 in
esophageal carcinoma. Dis Esophagus. 26:729–736. 2013.PubMed/NCBI
|
|
13
|
Tanaka F, Matsuzaki S, Mimori K, Kita Y,
Inoue H and Mori M: Clinicopathological and biological significance
of CDC28 protein kinase regulatory subunit 2 overexpression in
human gastric cancer. Int J Oncol. 39:361–372. 2011.PubMed/NCBI
|
|
14
|
Yu MH, Luo Y, Qin SL, Wang ZS, Mu YF and
Zhong M: Up-regulated CKS2 promotes tumor progression and predicts
a poor prognosis in human colorectal cancer. Am J Cancer Res.
5:2708–2718. 2015.PubMed/NCBI
|
|
15
|
Shen DY, Fang ZX, You P, Liu PG, Wang F,
Huang CL, Yao XB, Chen ZX and Zhang ZY: Clinical significance and
expression of cyclin kinase subunits 1 and 2 in hepatocellular
carcinoma. Liver Int. 30:119–125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen DY, Zhan YH, Wang QM, Rui G and Zhang
ZM: Oncogenic potential of cyclin kinase subunit-2 in
cholangiocarcinoma. Liver Int. 33:137–148. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kawakami K, Enokida H, Tachiwada T,
Gotanda T, Tsuneyoshi K, Kubo H, Nishiyama K, Takiguchi M, Nakagawa
M and Seki N: Identification of differentially expressed genes in
human bladder cancer through genome-wide gene expression profiling.
Oncol Rep. 16:521–531. 2006.PubMed/NCBI
|
|
18
|
Walker RA: World Health Organization
classification of tumours. Pathology and genetics of tumours of the
breast and female genital organs. Histopathology. 46:229. 2005.
View Article : Google Scholar
|
|
19
|
Prat J: FIGO staging for uterine sarcomas.
Int J Gynaecol Obstet. 104:177–178. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Coley HM, Shotton CF, Kokkinos MI and
Thomas H: The effects of the CDK inhibitor seliciclib alone or in
combination with cisplatin in human uterine sarcoma cell lines.
Gynecol Oncol. 105:462–469. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi J, Yu Y, Akilli Öztürk Ö, Holland JD,
Besser D, Fritzmann J, Wulf-Goldenberg A, Eckert K, Fichtner I and
Birchmeier W: New Wnt/β-catenin target genes promote experimental
metastasis and migration of colorectal cancer cells through
different signals. Gut. 65:1690–1701. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cui RR, Wright JD and Hou JY: Uterine
leiomyosarcoma: A review of recent advances in molecular biology,
clinical management and outcome. BJOG. 124:1028–1037. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cuppens T, Tuyaerts S and Amant F:
Potential therapeutic targets in uterine sarcomas. Sarcoma.
2015:2432982015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Taylor BS, Barretina J, Maki RG, Antonescu
CR, Singer S and Ladanyi M: Advances in sarcoma genomics and new
therapeutic targets. Nat Rev Cancer. 11:541–557. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mäkinen N, Aavikko M, Heikkinen T, Taipale
M, Taipale J, Koivisto-Korander R, Bützow R and Vahteristo P: Exome
sequencing of uterine leiomyosarcomas identifies frequent mutations
in tp53, atrx, and med12. PLoS Genet. 12:e10058502016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
You H, Lin H and Zhang Z: CKS2 in human
cancers: Clinical roles and current perspectives (Review). Mol Clin
Oncol. 3:459–463. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Koivisto-Korander R, Martinsen JI,
Weiderpass E, Leminen A and Pukkala E: Incidence of uterine
leiomyosarcoma and endometrial stromal sarcoma in Nordic countries:
Results from NORDCAN and NOCCA databases. Maturitas. 72:56–60.
2012. View Article : Google Scholar : PubMed/NCBI
|