Introduction
Malignant gliomas, which are classified by the World
Health Organization as grade III–IV tumors, are extremely lethal
primary brain tumors with an incidence of 3–5/100,000 each year
between 2000 and 2004 (1). At
present, the maximal therapy for malignant gliomas comprises
surgical resection followed by radiotherapy and concomitant
adjuvant temozolomide (1). However,
patients with malignant glioma exhibit a dismal prognosis (2), with a 5-year survival rate of <3%
(3). This poor prognosis is due to
therapeutic resistance and tumor recurrence following the combined
treatment approach (4). Previously,
numerous studies have identified that the resistance of malignant
gliomas to current therapy may reside in a relatively small
population of glioma stem cells (GSCs), which are hypothesized to
substantially influence tumor initiation and serve a critical role
in tumor propagation (5–9). The majority of GSCs have been revealed
to divide through symmetric cell division and not through
asymmetric cell division (10).
Recent reports have determined that the frequency of asymmetric
division of cancer stem cells is negatively correlated with their
proliferative capacity (11,12). A previous study in mouse models have
further demonstrated that decreased asymmetry in normal stem cells
is associated with neoplastic transformation, while therapies that
increase asymmetric division result in decreased numbers of
resistant GSCs (13).
All-trans retinoic acid (ATRA)-based differentiation
treatment has been observed to target the GSC population and induce
therapy-sensitizing and antitumor effects (14,15).
Although it has been demonstrated that ATRA-treatment alone is able
to induce antitumor effects (5), to
the best of our knowledge, it remains unknown whether and how the
GSC division mode is affected by treatment with ATRA. Therefore,
the present study used a single-cell-based method to observe the
differentiation-inducing effects of ATRA on the cell division
features of GSCs from the U87MG cell line.
Materials and methods
Culture of U87MG cells
The U87MG cell line (HTB-14; glioblastoma of unknown
origin) was purchased from the American Type Culture Collection
(Manassas, VA, USA). The cells were maintained in Dulbecco's
modified Eagle's medium (DMEM; Invitrogen: Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc.) at
37°C in 5% CO2. The identity of the U87MG cell line was
confirmed through comparison with the short tandem repeat profile
(Amelogenin, CSF1PO, D13S317, D7S820, D5S818, D16S539, vWA, THO1
and TPOX). This U87MG cells were used in subsequent experiments up
to the fifth passage.
Culture of U87MG GSCs
GSCs were isolated as described previously (16). U87MG cells were seeded onto plates at
a density of 2×104 cells/well. The serum-free culture
medium (termed tumor stem cell medium) consisted of DMEM/F12
(Gibco; Thermo Fisher Scientific, Inc.), B27 (1×; Gibco; Thermo
Fisher Scientific, Inc.), recombinant human epidermal growth factor
(20 ng/ml; Sigma-Aldrich, Merck, KGaA, Darmstadt, Germany) and
basic fibroblast growth factor (20 ng/ml; Upstate Biotechnology,
Inc., Lake Placid, NY, USA) without vincristine. Cultures were
incubated in 5% CO2 at 37°C and half of the medium was
replaced with DMEM every 2 days. When primary tumor spheres with
diameters of 70–100 µm were visible under a light microscope, all
of the culture medium was discarded and the wells were filled with
1 ml fresh serum-free tumor stem cell medium.
The primary tumor spheres were dissociated
enzymatically using Accutase (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol, into single cells and
seeded into a 96-well plate (2–3×103 cells/well) with
tumor stem cell medium (0.1 ml/well). The culture medium was
changed every 3 days. The clonogenic efficacy was examined by
calculating the percentage of the wells that contained spheres with
>50 cells on day 14. Five fields of vision under a light
microscope (×20 magnification) were randomly selected to count the
number of spheres. Adherent non-sphere-forming cells and parental
U87MG cells were used as controls for the capacity to form tumor
cell clones. The morphology of the secondary spheres was examined
microscopically. Five fields were randomly selected to examine the
morphology of the secondary spheres under a light microscope (×100
magnification).
Immunostaining
GSCs were plated onto poly-L-lysine (Sigma-Aldrich;
Merck, KGaA)-coated glass coverslips for 40 h at 37°C in DMEM
containing 10% FBS. The cells were fixed with 4% paraformaldehyde
(PFA) for 30 min at 4°C and then incubated with primary antibodies
against nestin (mouse monoclonal IgG1; 1:200; cat. no. 60091;
Stemcell Technologies, Inc., Vancouver, BC, Canada), β-tubulin III
(mouse monoclonal IgG1; 1:1,000; cat. no. 60052; Stemcell
Technologies, Inc.), glial fibrillary acidic protein (GFAP, rabbit
polyclonal; 1:400; cat. no. Z0334; Dako; Agilent Technologies,
Inc., Santa Clara, CA, USA), galactosylceramidase (GALC, rabbit
polyclonal; 1:20; cat. no. AB142; EMD Millipore, Billerica, MA,
USA) or Numb (rabbit polyclonal; 1:100; cat. no. ab155415; Abcam,
Cambridge, UK) overnight at 4°C. Appropriate secondary antibodies
(Alexa Fluor® 594 donkey anti-mouse; 1:1,000; cat. no.
A-21207; Alexa Fluor® 488 donkey anti-rabbit; 1:500;
cat. no. A-21202; Alexa Fluor® 488 donkey anti-mouse;
1:400; cat. no. A-21206; and Alexa Fluor® 594 donkey
anti-rabbit; 1:500; cat. no. A-21203; Molecular Probes; Thermo
Fisher Scientific, Inc.) were then selected for section incubation
at room temperature for 2 h. The cell nuclei were counterstained
with 4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich; Merck
KGaA) for 5 min at room temperature for fluorescence microscopy
(×200 magnification).
BrdU incorporation
Cells were cultured in DMEM/F12 medium containing 20
ng/ml EGF (Invitrogen; Thermo Fisher Scientific, Inc.). Once pairs
were observed, 10 µg/ml bromodeoxyuridine (BrdU; Invitrogen; Thermo
Fisher Scientific, Inc.) was added to the wells for 8–10 h. The
washed cells were fixed in 4% PFA at 4°C for 30 min, stained for
Numb (rabbit polyclonal; cat. no. ab155415; Abcam) overnight at
4°C, and then stained with anti-BrdU antibody (cat. no. ab6326;
1:250; Abcam). The cell nuclei were counterstained with DAPI for 5
min at room temperature for fluorescence microscopy (×200
magnification).
Flow cytometric analysis
For flow cytometric analysis, expanded GSCs were
collected and evaluated on an FC500 flow cytometer (Beckman
Coulter, Inc., Brea, CA, USA). The GSCs were stained with
PE-conjugated CD133 clone AC133 antibody (cat. no. 130-113-670;
Miltenyi Biotec, GmbH, Bergisch Gladbach, Germany) at a dilution of
1:50 in PBS with 0.5% BSA containing 20% FcR Blocking Reagent
(Miltenyi Biotec, GmbH) at 4°C for 10 min. The control was the
non-stained cells in the same buffer. Data were analyzed using
FlowJo software version 7.6.1 (Tree Star, Inc., Ashland, OR,
USA).
Mitotic image analysis
The fluorescence immunostaining intensity was
evaluated for each daughter cell to determine the degree of
asymmetric distribution of detected molecules between the two
daughter cells. The condensed DNA was revealed via staining with
DAPI and the morphology of the phase-contrasted image was used to
identify the dividing daughter cells. Using Image-Pro Plus v. 6.2
(Media Cybernetics, Inc., Rockville, MD, USA) with the polygon
tool, the integrated fluorescent signals of each defined daughter
cell in a dividing pair were determined for the stained markers and
DNA.
Differentiation assay
U87MG GSC spheres and adherent non-sphere-forming
cells were plated on a sterile 24-well glass slide coated with
poly-L-lysine (Sigma-Aldrich, Merck KGaA) in DMEM/F12 neurosphere
culture medium. These cells were fed with FBS-supplemented culture
medium, and coverslips were stained for GFAP, GALC and β-tubulin
III as described above on day 2. The nuclei were visualized using
DAPI. Quantification of the positive cells stained with each
antibody was evaluated as a percentage of the total number of cells
counted under five random fluorescence microscope fields (×200
magnification).
ATRA-treatment
The cells in the culture medium were fed with 10%
FBS (vehicle group) or FBS combined with ATRA (Sigma-Aldrich;
dissolved in DMSO) at 10 nmol/l (ATRA group) (5).
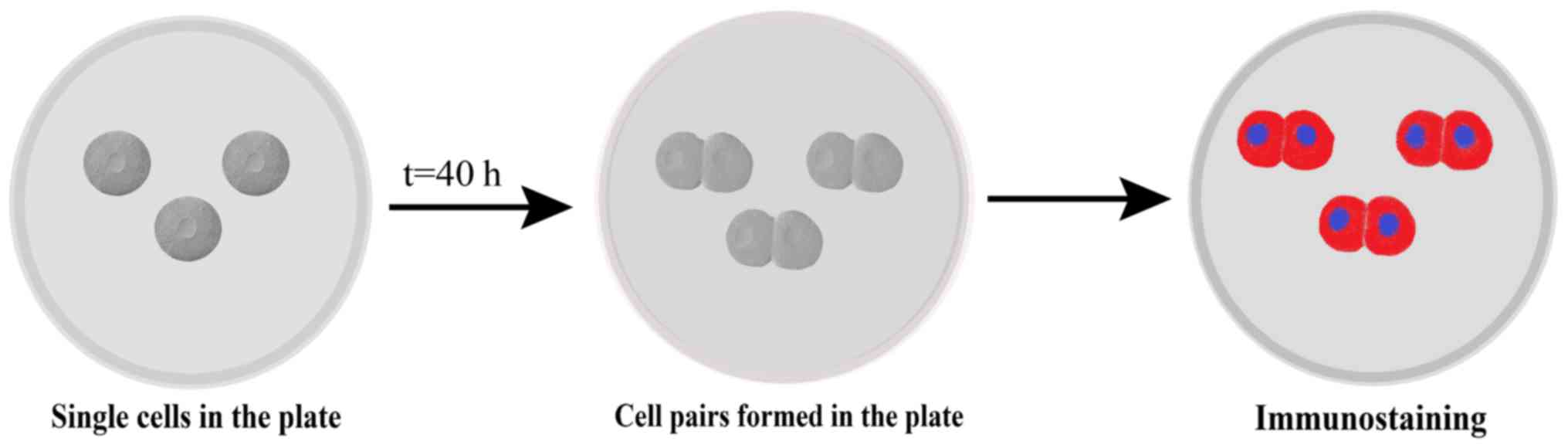
Pair assay
Cells were subjected to a pair assay as described
previously (17). In brief, cells
were plated at a density of 2,000–3,000 cells/well for 40 h and the
cell pairs were observed using time-lapse microscopy, fixed with 4%
PFA at 4°C for 30 min, and subjected to immunostaining with Numb
primary and secondary antibodies as described above. Nuclei were
stained with DAPI, and a total of 200 cell pairs were analyzed
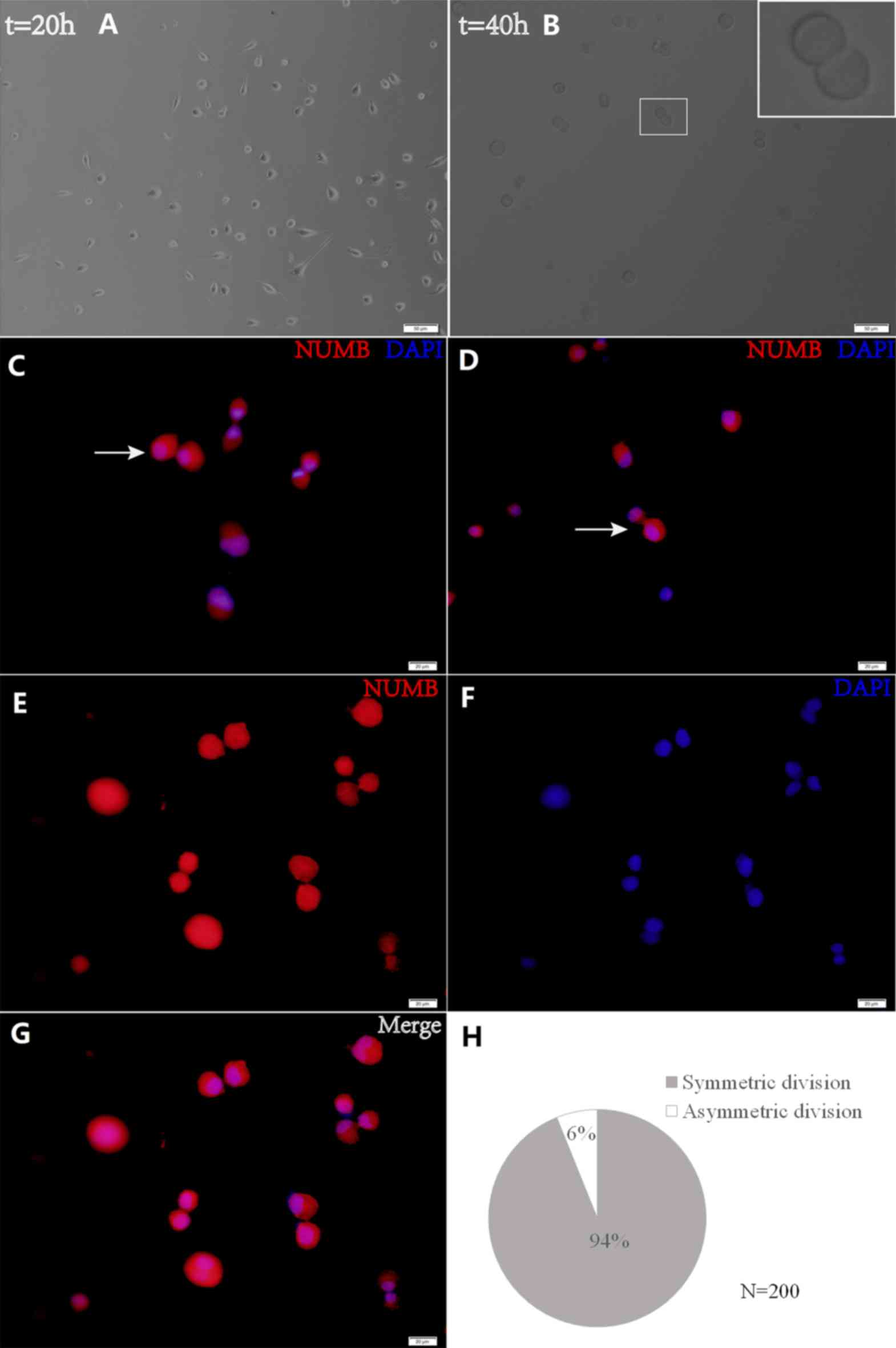
using a fluorescence microscope (×200 magnification; Fig. 1).
Statistical analysis
Since paired daughter cells should have equal DAPI,
the percentage deviation of the DAPI distribution was calculated in
order to quantify the asymmetry, according to the protocol outlined
in a previous study (12). For a
minimum of 100 cells, the mean distribution of DAPI was calculated
to be 6.848% and the standard deviation was 5.501. Then, the
asymmetry cutoff was set to be a difference of >21% between
paired daughter cells, based on the >99% confidence interval for
DAPI. Based on the 21% cutoff value, each cell division was defined
as either symmetrical or asymmetrical.
All experiments were repeated in triplicate, and the
data are presented as the mean ± standard deviation. The data were
processed using SPSS 22.0 statistical software (IBM Corp., Armonk,
NY, USA). Unpaired Student's t-test was used to determine the
significance between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
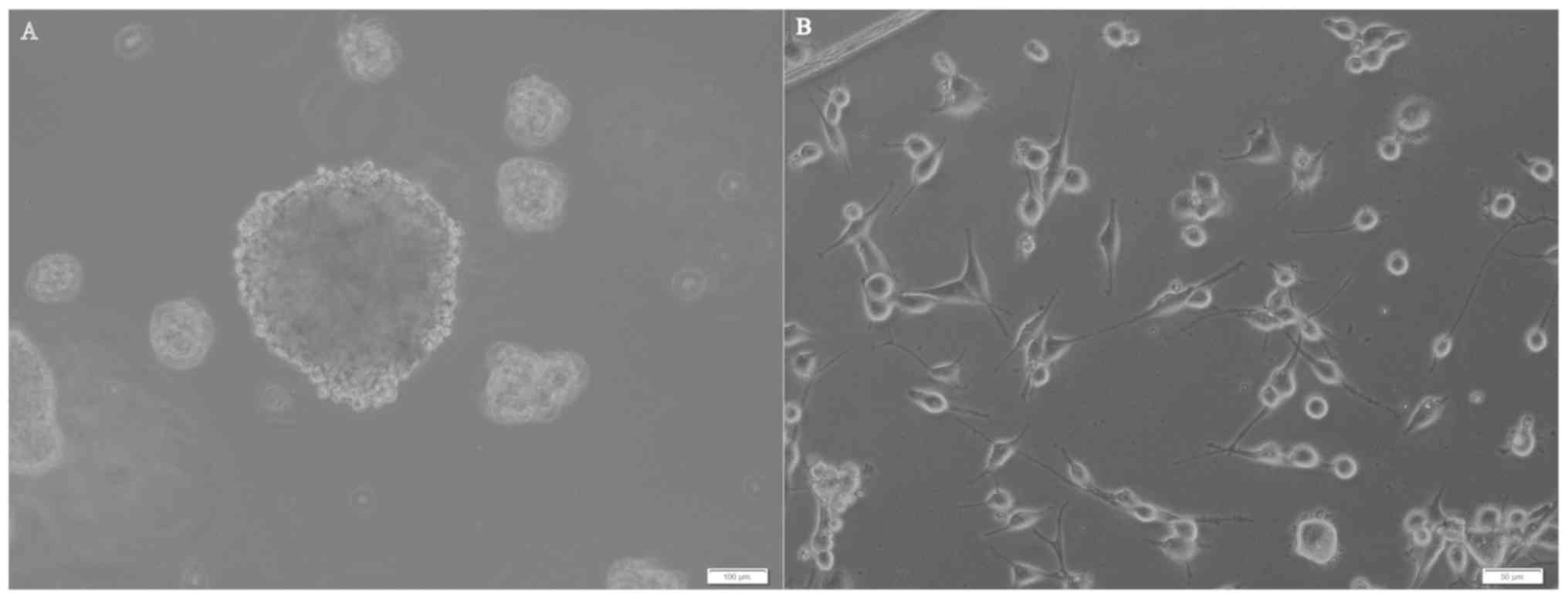
Detection of tumor sphere forming
cells with self-renewal capability
U87MG cells were seeded on plates at a density of
2×104/well with progressive increases in the serum-free
neural stem cell culture medium. After 5–6 days, 30–50 small
spheres per well (each containing 10–20 cells) were observed. At 2
weeks, these spheres had increased their diameters by 10 to 30-fold
(Fig. 2A). Numerous adherent
non-sphere-forming cells were observed in the bottoms of the wells
(Fig. 2B), and remained viable
during an 8-week period; however, they did not generate new
spheres. Cells in tumor spheres could be passaged for many
generations in serum-free neural stem cell culture medium.
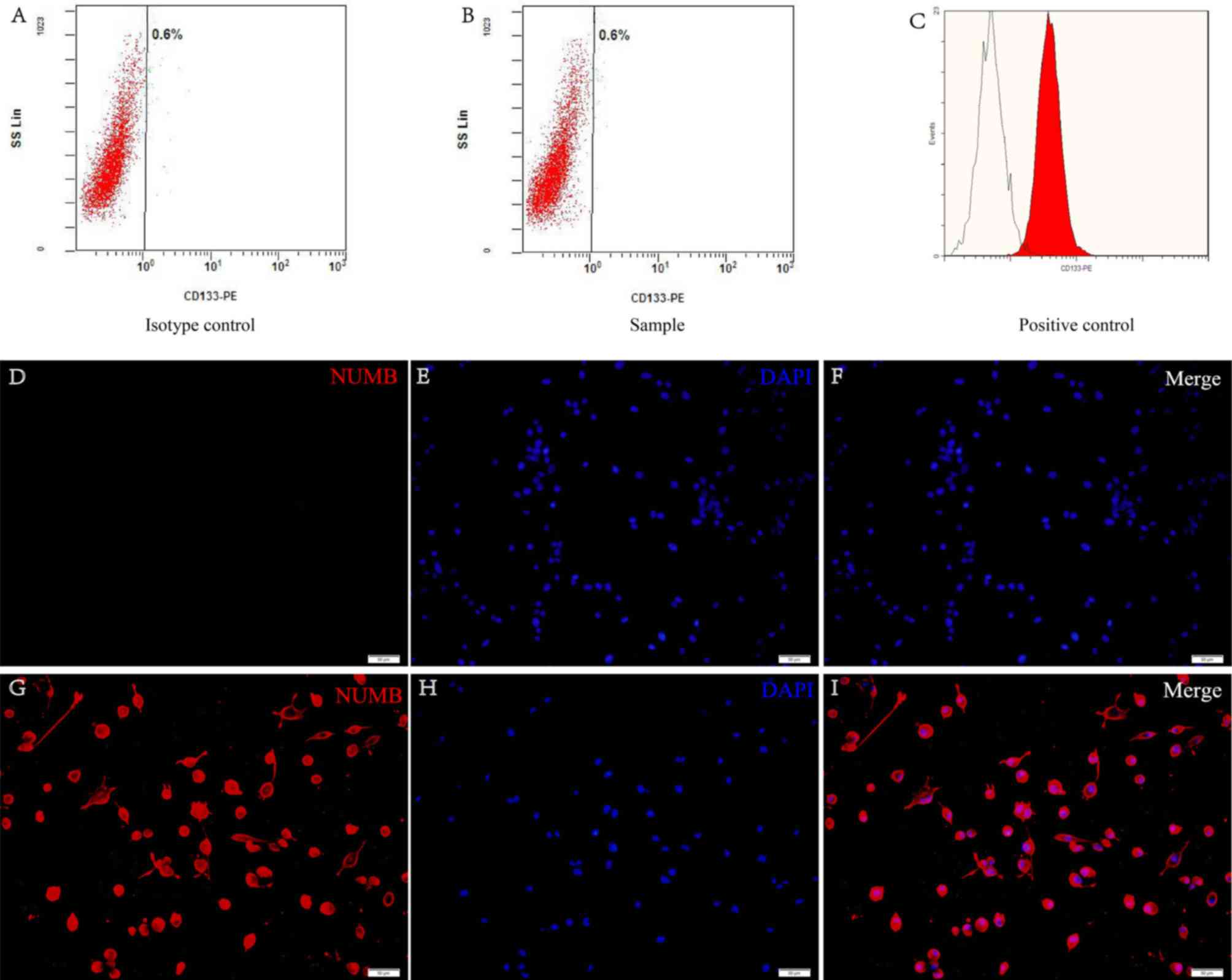
Expression of neural stem cell
markers
CD133 and nestin are cell surface markers for neural
stem cells and are the widely accepted markers for brain GSCs
(11). Flow cytometric analysis
revealed that these cells had no presence of CD133 on the surface
(Fig. 3A and B). The retinoblastoma
cell line was used as a positive control (Fig. 3C). The data demonstrated that nestin
was positive in the cells from the tumor secondary spheres
(Fig. 3D-I).
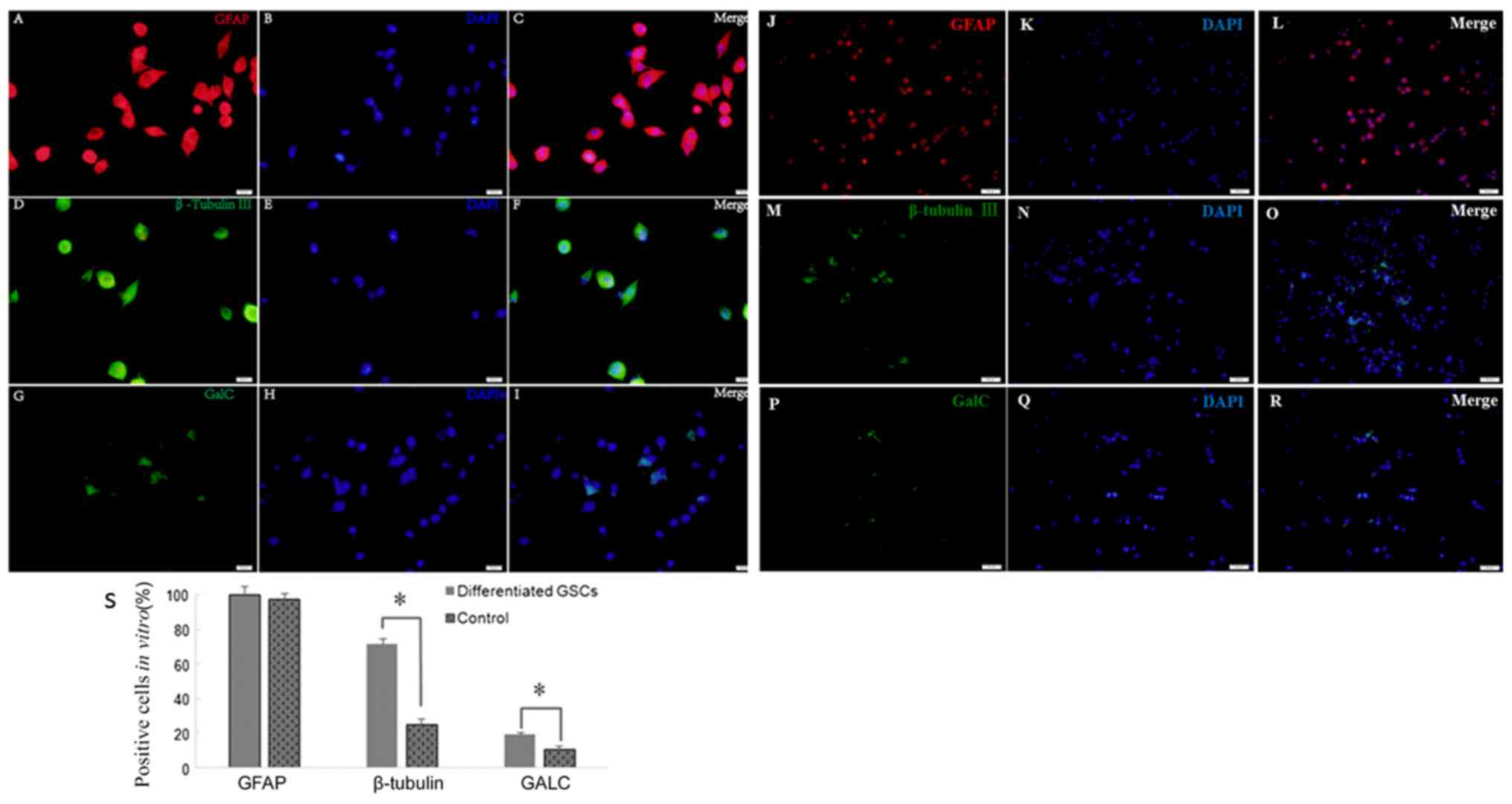
Multipotency in the differentiation of
secondary spheres
An important feature of GSCs are their
differentiation capability. Following culture in medium with 10%
FBS for 2 weeks, the cells differentiated from tumor secondary
spheres were stained positive for GFAP (99.65±7.42 vs. 97.37±6.06%;
P>0.05), β-tubulin III (71.25±3.45 vs. 25.33±3.18%; P<0.05)
and GALC (19.23±1.41 vs. 10.87±2.01%; P<0.05) compared with the
control cells (Fig. 4). Therefore,
secondary spheres derived from the single parental cells of primary
tumor spheres were able to differentiate into three neural cell
lineages.
Asymmetric distribution of Numb is
associated with asymmetric GSC divisions
The present study utilized time-lapse lineage
tracing to monitor GSC cell divisions at a single-cell resolution
(Fig. 5). Cell divisions were
recorded using time-lapse microscopy, and cells were subsequently
fixed and stained for Numb. Following a 40-h culture, the present
study identified the cells that had divided once and generated a
cell pair (Fig. 5B). Through this
analysis, the Numb distribution modes were detected in the daughter
cells (Fig. 5C and D). Sister cells
with symmetric Numb usually appeared similar. In order to quantify
the asymmetry, asymmetrical division was defined as having occurred
when the percent deviation of the stained marker between daughter
cells was >21%. Using this criterion, it was discovered that the
majority of pairs (93.91±1.59 vs. 6.09±1.57%) exhibited symmetry in
the Numb distribution (Fig. 5H).
These data demonstrate that GSCs are primarily maintained by
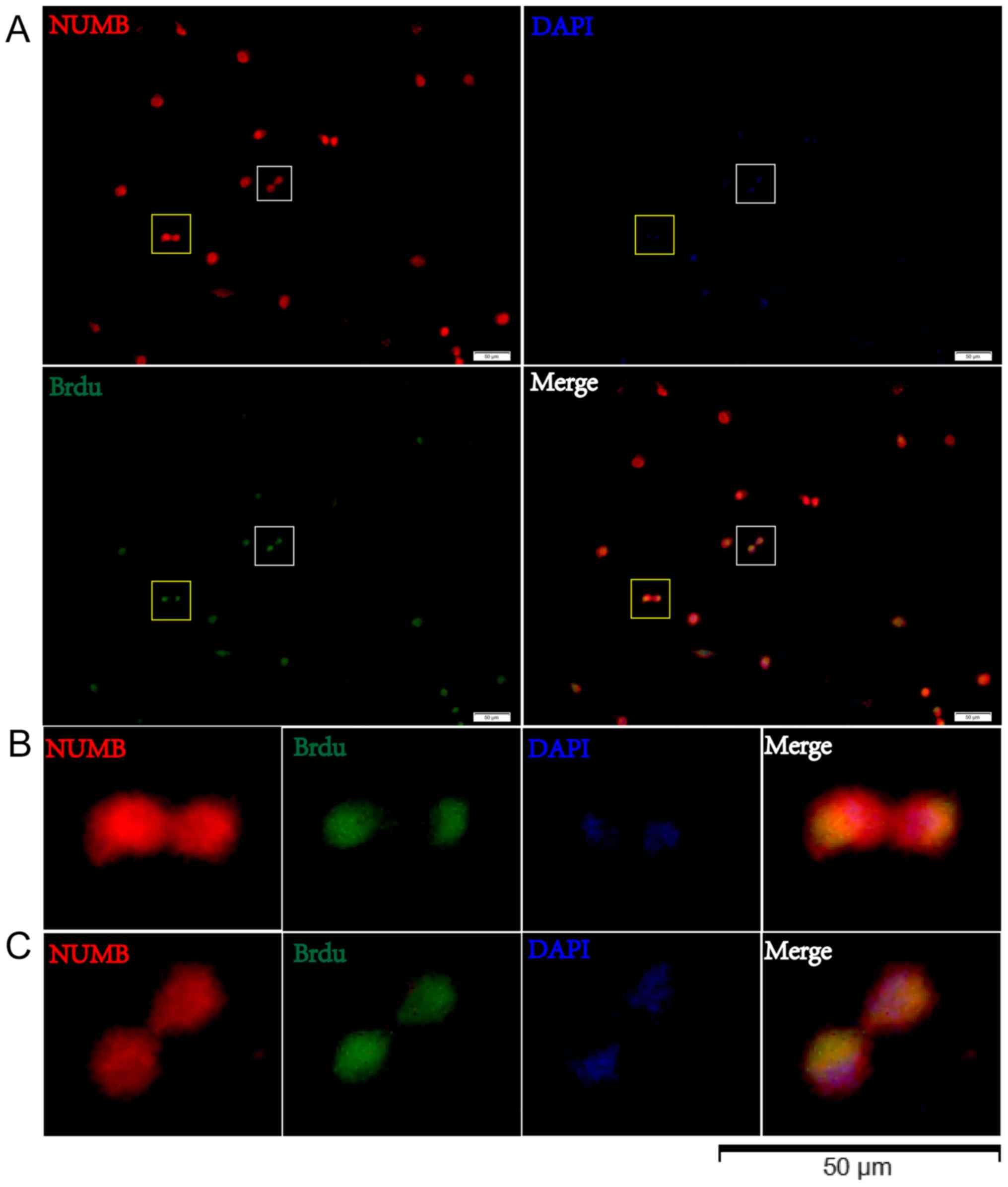
symmetric, as opposed to asymmetric, cell divisions (Fig. 5E-G). Cells were then treated with
BrdU for 8 h and the BrdU distribution was observed in relation to
Numb expression by immunostaining in pair assays. The data revealed
that BrdU incorporation in the two daughter cells was associated
with Numb asymmetry (Fig. 6).
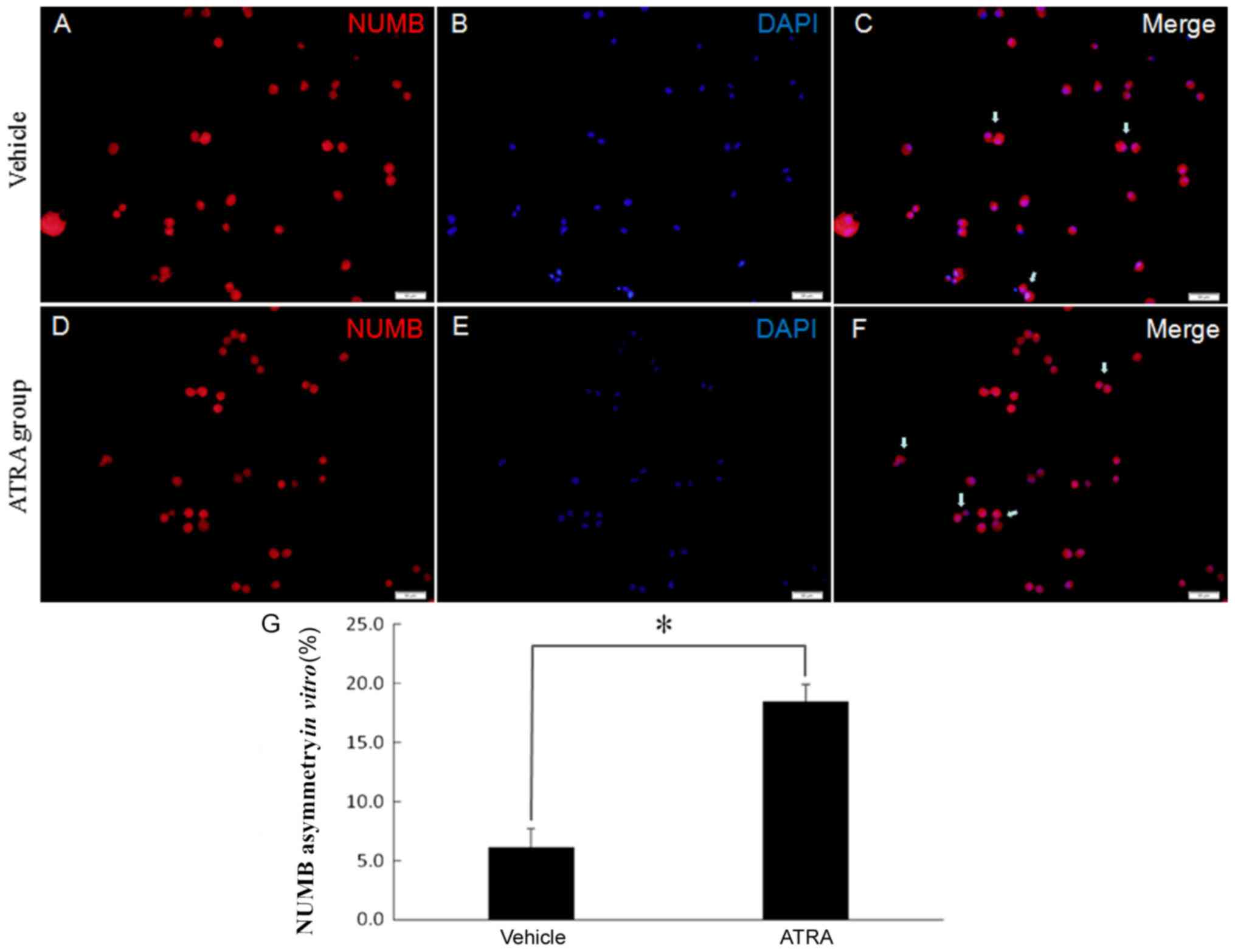
ATRA increased the asymmetric cell
division of GSCs
To investigate whether symmetric divisions are
affected when ATRA is administered, the present study observed the
division mode when ATRA was added to the culture. Neurospheres were
grown in DMEM containing 10% FBS and 10 nmol/l ATRA to induce
differentiation. Cell divisions were recorded for 40 h using
time-lapse video microscopy. Pairs of daughter cells from single
progenitor cells were identified, fixed and stained for Numb. There
was a decrease in symmetrical stem cell expansion division
incidence to 18.30±1.43% of the events in the ATRA-treated cells
(Fig. 7). This suggests that the
cancer stem cells were impaired in their ability to undergo a
symmetric division.
Discussion
The results of the present study demonstrated that
the GSCs isolated from the U87MG cell line did not express CD133.
Using single-cell-based analytical methods and quantitative
immunofluorescence, the present study revealed that Numb was widely
expressed in GSCs and symmetrically segregated into two daughter
cells during the division of GSCs. The present study, to the best
of our knowledge, is the first to demonstrate, that the Numb
symmetrical distribution proportion was decreased following
ATRA-based differentiation treatment.
GSCs have been identified in numerous primary
culture GBM cells and malignant glioma cell lines (5,18–21). A
previous study has confirmed that GSCs are the source of new tumor
cells following the administration of temozolomide (22). The universal GSC marker remains a
controversial topic due to high interpatient as well as
intratumoral variability (8).
Various surface markers, including CD133, nestin, Musashi-1, Nanog
and platelet-derived growth factor receptor, have been reported to
successfully identify GSC populations in vitro and in
vivo (8,14). Additional stem cell marker detection
would further support our conclusions. The present study analyzed
CD133 expression using flow cytometry and identified that CD133 was
negative in glioma cell spheres cells cultured from the U87MG cell
line. This result differs from that of previous research, which
reported that the majority of U87MG cells in the spheres were
positive for CD133 (20). Further
results using immunofluorescence revealed that the CD133-negative
cell populations expressed nestin. In addition, the cell
populations of the cultured tumor spheres were able to
differentiate into cells positive for GFAP, β-tubulin III and GALC,
which are representative markers of neuronal, astroglial and
oligodendroglial cells (23). These
results suggested successful induction of GSCs from the U87MG cell
line. However, the lack of an exact evaluation of
stemness/differentiation marker expression levels is a limitation
of the present study. Although CD133 has been defined as a marker
of glioma stem cells, an increasing amount of evidence has
demonstrated that the use of CD133 as a unique glioma stem cell
marker is insufficient to tag all GSCs. For example, fresh human
glioma and gliomasphere cultures express CD133 at low and sometimes
barely detectable levels (21).
Secondly, CD133-positive and CD133-negative GSCs from cell lines
and GBM tumors exhibited cancer stem cell properties (20,24).
Thirdly, neither the expression of stemness genes nor the long-term
self-renewal capacities of CD133-positive and CD133-negative cells
were significantly different (25).
Finally, CD133 negative cells were tumorigenic when implanted into
rat brains (26). A previous study
demonstrated that the levels of surface CD133 fluctuate during the
cell cycle in GSCs (27), indicating
that CD133 expression is likely a marker of certain stages of GSC
division, rather than a constitutive marker of GSCs.
Lathia et al (10) examined a variety of molecules in GSCs
and observed that only Numb and CD133 could be asymmetrically
segregated. Since the results of the present study demonstrated
that CD133 expression was negative in GSCs cultured from the U87MG
glioblastoma of unknown origin cell line, the present study used
Numb to analyze the GSC division mode. The data revealed that Numb
protein was expressed in 99% of GSCs from the U87MG cell line.
Using single-cell-based observations, the current study
demonstrated that the Numb distribution was predominantly symmetric
in the two daughter cells (94%) during GSC division. BrdU
incorporation indicates the proliferative ability of cells that
were actively replicating their DNA. The results of the present
study demonstrated that the BrdU distribution in the two daughter
cells was associated with Numb asymmetry. A limitation of the
present study is that the exact level of BrdU in paired cells was
not measured. In paraffin-embedded glioblastoma specimens, a
previous study indicated that 85% of cells exhibited a symmetric
pattern of Numb immunoreactivity (28). Numb is a so-called fate-determining
molecule that promotes the differentiation of neural stem cells
through antagonizing the notch and hedgehog signaling pathways
(29,30). The function of Numb is critical for
the occurrence of asymmetric cell division, and different
expressions of Numb may indicate cell fate divergence (31). Previous studies have suggested that
symmetric determinants exert pivotal functions in tumor initiation,
as defects in either the function of fate determinants and
regulators of asymmetric division, or the loss of asymmetric
division may lead to tumor development (13,32).
Although previous data demonstrated that the overexpression of Numb
did not induce either differentiation of U87MG cells or alter their
morphology, nor that the cell population doubling time was
significantly affected (33), until
now, numerous findings have demonstrated that Numb is associated
with the GSC markers SRY (sex determining region Y)-box 2 and
paired box protein Pax-6, as well as GSC survival, proliferation,
aggressiveness and therapeutic resistance (12,28,34).
The present results demonstrated a decrease in
symmetrical cell division incidence to 82% of the events in
ATRA-treated GSCs from the U87MG cell line. ATRA is a metabolite of
vitamin A that is able to induce complete remission in the majority
of acute promyelocytic leukemia cases when administered in
combination with light chemotherapy and/or arsenic trioxide
(35). To date, the efficacy of ATRA
to treat solid tumors remains poor (35). A previous study demonstrated that
ATRA could reduce the proliferation and invasiveness of U87MG
stem-like glioma cells in a dose-dependent manner and decrease tube
formation and vascular endothelial growth factor secretion
(36). When ATRA was administered in
combination with mTOR and PI3K inhibition, there was a synergistic
effect resulting in the minimum amount of cellular migration in
GSCs (15). It has been revealed
that ATRA-based differentiation targeted GSCs and induced GSCs to
differentiate, which was mediated by the activation of ERK1/2
(37). The results of the present
study demonstrated that differentiation treatment with ATRA induced
asymmetric division in GSCs from the U87MG cell line. The
underlying mechanism may be associated with the distribution of
Numb (12,28,31). In
a future study, we aim to investigate how ATRA affects the
distributions of p53, E-cadherin and Notch in the paired GSCs in
order to understand this underlying mechanism. Asymmetric division
is a feature more closely associated with cancer stem cells from
early-stage and well-differentiated tumors, while late stage tumors
suppress asymmetric division and increase symmetric division
(38). Therefore, the increase of
asymmetric division following treatment with ATRA may be associated
with the suppression of tumorigenesis in GSCs. However, from the
present evidence it cannot be concluded that ATRA can be used as a
glioblastoma treatment agent.
Although ATRA induces morphological differentiation
of GSCs in vitro, one of the primary limitations in the
present study is that it remains controversial as to whether the
cell line U87MG is representative of highly malignant glioma.
Therefore, further experiments on primary cell lines are required.
Secondly, ATRA-metabolizing enzymes (39) and loss of retinoic acid receptors
(40) may render cells irresponsive
to ATRA effects. Additional studies are necessary in order to
investigate how ATRA influences the GSC division mode in
vivo. In conclusion, the results of the present study
demonstrated that ATRA-induced asymmetric division of GSCs from the
U87MG glioblastoma cell line of unknown origin.
Acknowledgements
Not applicable.
Funding
This study was supported by the Shaanxi Province
Scientific and Technology Development Program (grant no.
2012SF2-03-05) and the Xi'an Jiaotong University Free Exploration
Program (grant no. xjj2013059).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
RW designed the study and performed the majority of
experiments. CL performed the GSC cultures and immunostaining. RW
was a major contributors to the writing of the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Second Hospital of Xi'an Jiaotong University
(Xi'an, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Johnson DR and O'Neill BP: Glioblastoma
survival in the United States before and during the temozolomide
era. J Neurooncol. 107:359–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: Effects of radiotherapy with concomitant and
adjuvant temozolomide versus radiotherapy alone on survival in
glioblastoma in a randomised phase III study: 5-year analysis of
the EORTC-NCIC trial. Lancet Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Friedmann-Morvinski D, Narasimamurthy R,
Xia Y, Myskiw C, Soda Y and Verma IM: Targeting NF-κB in
glioblastoma: A therapeutic approach. Sci Adv. 2:e15012922016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Campos B, Wan F, Farhadi M, Ernst A,
Zeppernick F, Tagscherer KE, Ahmadi R, Lohr J, Dictus C, Gdynia G,
et al: Differentiation therapy exerts antitumor effects on
stem-like glioma cells. Clin Cancer Res. 16:2715–2728. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cancer Genome Atlas Research Network, .
Comprehensive genomic characterization defines human glioblastoma
genes and core pathways. Nature. 455:1061–1068. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sundar SJ, Hsieh JK, Manjila S, Lathia JD
and Sloan A: The role of cancer stem cells in glioblastoma.
Neurosurg Focus. 37:E62014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Auffinger B, Tobias AL, Han Y, Lee G, Guo
D, Dey M, Lesniak MS and Ahmed AU: Conversion of differentiated
cancer cells into cancer stem-like cells in a glioblastoma model
after primary chemotherapy. Cell Death Differ. 21:1119–1131. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Olmez I, Shen W, McDonald H and Ozpolat B:
Dedifferentiation of patient-derived glioblastoma multiforme cell
lines results in a cancer stem cell-like state with
mitogen-independent growth. J Cell Mol Med. 19:1262–1272. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lathia JD, Hitomi M, Gallagher J, Gadani
SP, Adkins J, Vasanji A, Liu L, Eyler CE, Heddleston JM, Wu Q, et
al: Distribution of CD133 reveals glioma stem cells self-renew
through symmetric and asymmetric cell divisions. Cell Death Dis.
2:e2002011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
O'Brien CA, Kreso A, Ryan P, Hermans KG,
Gibson L, Wang Y, Tsatsanis A Gallinger S and Dick JE: ID1 and ID3
regulate the self-renewal capacity of human colon cancer-initiating
cells through p21. Cancer Cell. 21:777–792. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pece S, Tosoni D, Confalonieri S, Mazzarol
G, Vecchi M, Ronzoni S, Bernard L, Viale G, Pelicci PG and Di Fiore
PP: Biological and molecular heterogeneity of breast cancers
correlates with their cancer stem cell content. Cell. 140:62–73.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sugiarto S, Persson AI, Munoz EG,
Waldhuber M, Lamagna C, Andor N, Hanecker P, Ayers-Ringler J,
Phillips J, Siu J, et al: Asymmetry-defective oligodendrocyte
progenitors are glioma precursors. Cancer Cell. 20:328–340. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang M, Song T, Yang L, Chen R, Wu L,
Yang Z and Fang J: Nestin and CD133: Valuable stem cell-specific
markers for determining clinical outcome of glioma patients. J Exp
Clin Cancer Res. 27:852008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Karsy M, Albert L, Tobias ME, Murali R and
Jhanwar-Uniyal M: All-trans retinoic acid modulates cancer stem
cells of glioblastoma multiforme in an MAPK-dependent manner.
Anticancer Res. 30:4915–4920. 2010.PubMed/NCBI
|
|
16
|
Yu SC, Ping YF, Yi L, Zhou ZH, Chen JH,
Yao XH, Gao L, Wang JM and Bian XW: Isolation and characterization
of cancer stem cells from a human glioblastoma cell line U87.
Cancer Lett. 265:124–134. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun Y, Goderie SK and Temple S: Asymmetric
distribution of EGFR receptor during mitosis generates diverse CNS
progenitor cells. Neuron. 45:873–886. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Singh SK, Clarke ID, Terasaki M, Bonn VE,
Hawkins C, Squire J and Dirks PB: Identification of a cancer stem
cell in human brain tumors. Cancer Res. 63:5821–5828.
2003.PubMed/NCBI
|
|
19
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Howard CM, Valluri J, Alberico A, Julien
T, Mazagri R, Marsh R, Alastair H, Cortese A, Griswold M, Wang W,
et al: Analysis of Chemopredictive assay for targeting cancer stem
cells in glioblastoma patients. Transl Oncol. 10:241–254. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zheng X, Shen G, Yang X and Liu W: Most C6
cells are cancer stem cells: Evidence from clonal and population
analyses. Cancer Res. 67:3691–3697. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen J, Li Y, Yu TS, McKay RM, Burns DK,
Kernie SG and Parada LF: A restricted cell population propagates
glioblastoma growth after chemotherapy. Nature. 488:522–526. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu Y, Richard JP, Wang SD, Rath P, Laterra
J and Xia S: Regulation of glioblastoma multiforme stem-like cells
by inhibitor of DNA binding proteins and oligodendroglial
lineage-associated transcription factors. Cancer Sci.
103:1028–1037. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Joo KM, Kim SY, Jin X, Song SY, Kong DS,
Lee JI, Jeon JW, Kim MH, Kang BG, Jung Y, et al: Clinical and
biological implications of CD133-positive and CD133-negative cells
in glioblastomas. Lab Invest. 88:808–815. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clement V, Dutoit V, Marino D, Dietrich PY
and Radovanovic I: Limits of CD133 as a marker of glioma
self-renewing cells. Int J Cancer. 125:244–248. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang J, Sakariassen PØ, Tsinkalovsky O,
Immervoll H, Bøe SO, Svendsen A, Prestegarden L, Røsland G, Thorsen
F, Stuhr L, et al: CD133 negative glioma cells form tumors in nude
rats and give rise to CD133 positive cells. Int J Cancer.
122:761–768. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Barrantes-Freer A, Renovanz M, Eich M,
Braukmann A, Sprang B, Spirin P, Pardo LA, Giese A and Kim EL:
CD133 expression is not synonymous to immunoreactivity for AC133
and fluctuates throughout the cell cycle in glioma stem-like cells.
PLoS One. 10:e01305192015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang X, Xing H, Kim TM, Jung Y, Huang W,
Yang HW, Song S, Park PJ, Carroll RS and Johnson MD: Numb regulates
glioma stem cell fate and growth by altering epidermal growth
factor receptor and Skp1-Cullin-F-box ubiquitin ligase activity.
Stem Cells. 30:1313–1326. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Di Marcotullio L, Greco A, Mazzá D,
Canettieri G, Pietrosanti L, Infante P, Coni S, Moretti M, De
Smaele E, Ferretti E, et al: Numb activates the E3 ligase Itch to
control Gli1 function through a novel degradation signal. Oncogene.
30:65–76. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Beres BJ, George R, Lougher EJ, Barton M,
Verrelli BC, McGlade CJ, Rawls JA and Wilson-Rawls J: Numb
regulates Notch1, but not Notch3, during myogenesis. Mech Dev.
128:247–257. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shen Q, Zhong W, Jan YN and Temple S:
Asymmetric Numb distribution is critical for asymmetric cell
division of mouse cerebral cortical stem cells and neuroblasts.
Development. 129:4843–4853. 2002.PubMed/NCBI
|
|
32
|
Bajaj J, Zimdahl B and Reya T: Fearful
symmetry: Subversion of asymmetric division in cancer development
and progression. Cancer Res. 75:792–797. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Euskirchen P, Skaftnesmo KO, Huszthy PC,
Brekkå N, Bjerkvig R, Jacobs AH and Miletic H: NUMB does not impair
growth and differentiation status of experimental gliomas. Exp Cell
Res. 317:2864–2873. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Beier CP and Beier D: CD133 negative
cancer stem cells in glioblastoma. Front Biosci (Elite Ed).
3:701–710. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schenk T, Stengel S and Zelent A:
Unlocking the potential of retinoic acid in anticancer therapy. Br
J Cancer. 111:2039–2045. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ling GQ, Liu YJ, Ke YQ, Chen L, Jiang XD,
Jiang CL and Ye W: All-trans retinoic acid impairs the vasculogenic
mimicry formation ability of U87 stem-like cells through promoting
differentiation. Mol Med Rep. 12:165–172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Friedman MD, Jeevan DS, Tobias M, Murali R
and Jhanwar-Uniyal M: Targeting cancer stem cells in glioblastoma
multiforme using mTOR inhibitors and the differentiating agent
all-trans retinoic acid. Oncol Rep. 30:1645–1650. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bu P, Chen KY, Lipkin SM and Shen X:
Asymmetric division: A marker for cancer stem cells in early stage
tumors? Oncotarget. 4:950–951. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Armstrong JL, Taylor GA, Thomas HD, Boddy
AV, Redfern CP and Veal GJ: Molecular targeting of retinoic acid
metabolism in neuroblastoma: The role of the CYP26 inhibitor
R116010 in vitro and in vivo. Br J Cancer. 96:1675–1683. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chakravarti N, Lotan R, Diwan AH, Warneke
CL, Johnson MM and Prieto VG: Decreased expression of retinoid
receptors in melanoma: Entailment in tumorigenesis and prognosis.
Clin Cancer Res. 13:4817–4824. 2007. View Article : Google Scholar : PubMed/NCBI
|