Cancer is one of the most devastating diseases
afflicting humans, not only because it negatively affects the
health and quality of life of patients, but also because it places
a great burden on society as a whole (1). A significant amount of progress in
cancer treatment has been made over the last two decades; however,
For all cancers combined, the 5-year relative survival rate of
patients at advance stages with cancer remains less than 20% during
the most recent time period (2007–2013) in United States (2). Surgery, chemotherapy and radiotherapy
are currently the main forms of cancer treatment; however, novel
therapeutic approaches, including precision treatment (3), personalized therapy (4), molecular target therapy (5), complex immunologic therapy (6) and endocrine therapy (7,8), have
also emerged. In addition, targeting cancer stem cells (CSCs) is an
approach that has attracted an increasing amount of attention in
cancer research, both in the lab and in clinical settings (9,10).
As one of the most prevalent and deadly malignancies
in China, esophageal cancer with respective modality and mortality
rates of 478 and 375 per 10,000, has become the focus of an
increasing number of studies (1).
Among different types of esophageal cancer, squamous cell cancer is
the most common malignancy among the Chinese population (22). The present review will focus on the
role of CSCs in esophageal squamous cell cancer (ESCC), including
their incidence and 5-year survive rate, isolation, identification,
signaling pathways, association with viral infection, and their
involvement in diagnosis and treatment.
ESCC is a common malignant tumor of the digestive
tract, and relapse and metastasis to surrounding lymphatic nodes
are major traits of ESCC (23).
Since this type of cancer usually exhibits few or no obvious
symptoms at the early stages of disease, >50% of patients with
ESCC present with metastases and unresectable tumor at the time of
diagnosis (24) and the overall
5-year survive rate remains at 15–25% (25). In addition, due to the anatomical
location, ESCC is difficult to completely resect, and ESCC is less
sensitive to chemotherapy and radiotherapy (26). Given the important roles of CSCs in
cancer initiation, progression, recurrence and metastasis (27), targeting ESCC-specific CSCs has
become an attractive alternative. One of the major obstacles in CSC
research and application is obtaining CSCs. There are currently
four frequently used, well-established methods in CSC isolation.
Although the gold standard for defining a CSC or tissue stem cell
would be serial transplantation, this is a timely and costly
experiment (28).
A common method used to isolate CSCs is to utilize
cell surface markers, including cluster of differentiation (CD) 90
and CD44, for isolation by flow cytometry (10). This was also the earliest reported
method. Using flow cytometry Bonnet and Dick (12) identified that CD34++/CD38- leukemic
cells were acute myeloid leukemia stem cells in 1997. Since then,
it appears that CSCs can be isolated from almost every type of
cancer using this method, i.e. utilizing special cell surface
markers, primarily CD molecules. Although several studies (29–32) have
demonstrated an association between CDs, including CD133, and
self-renewal and multidirectional differentiation abilities, it is
not apparent as to why CDs can be regarded as surface markers for
CSCs. Following the silencing of CD133, the stemness of CSCs was
decreased (33). ESCC was not an
exception to this phenomenon. In a previous study, CD90+ ESCC cells
were isolated using flow cytometry, and further characterization of
this population by mRNA profiling suggested that CD90+ is involved
in tumor growth and metastasis, via the dysregulation of the
Ets-1/matrix metalloproteinase (MMP) signaling pathway and a change
from an epithelial phenotype to a mesenchymal phenotype. The
isolated CD90+ ESCC cells possessed a higher self-renewal
capability and were able to initiate tumor growth, differentiation,
metastasis and chemotherapeutic resistance (34). CD44 is a common stem cell marker that
has been identified in several types of human cancer, including
breast (31,35), gastric (36), prostate (37) and colorectal cancer (38), as well as glioma (39). In addition, CD44 is considered to be
a stem cell marker of ESCC (40).
The second method for isolating CSCs from ESCC is to
use a serum-free medium suspension culture. In the mid-1960s,
serum-free culture was first used to enable the L cell line to
propagate from its low density of inoculum by adding specific
nutrients to substitute the serum (41). Serum-free culture was subsequently
applied in a cytokine supported stroma-free suspension culture
protocols (42,43). To achieve a specific aim, including
collecting cytokines or high molecular weight proteins released
from cultured cells. Serum-free suspension culture was first
applied to isolate human primitive hematopoietic progenitor cells
(44) and is widely regarded as an
effective method for isolating stem cells or CSCs. However,
serum-free suspension culture exhibits limitations, due to the
impaired differentiation and increased proliferation abilities of
CSCs. It is speculated that the cells in the center of spheres are
prone to be senescent. In a sphere, the CSCs are not homogeneous,
but hierarchical. The cells closer to the core of the sphere are
more differentiated, which is possibly explained by the fact that
fewer nutrients reach the core. The cells in the core are prone to
differentiation due to a lack of growth factor (45). It is largely accepted that the number
of CSCs in the sphere continuously increases, as the cells undergo
further passage (46). However, the
risk of gene variation increases with each passage. Theoretically,
only CSCs can form microspheres; however, not every sphere cell is
a CSC. The sphere formation ability can, to a certain extent,
reflect the self-renewal ability of CSCs (46). In numerous types of cancer,
microsphere cells from the serum-free medium suspension culture
have been demonstrated to exhibit stem cell-like characteristics
(47,48). Wang et al (49) reported that sphere cells isolated
from the ESCC cell lines KYSE 150 and TE1 are more resistant to
radiotherapy, when compared with parental cells.
A third method of CSC isolation is to utilize
specific stem cell phenotype determinants, including side
population (SP) cells. In 1997, Goodell et al (50) utilized dual-wavelength flow cytometry
analysis of murine bone marrow cells stained with the fluorescent
DNA-binding dye Hoechst 33342 to obtain a small fraction of cells
that could efflux the Hoechst dye, defining an extremely small and
homogeneous population of cells termed SP cells. These cells are
primarily CD34- and lineage marker-negative, highly enriched for
long-term culture-initiating cells, which is an indicator of
primitive hematopoietic cells, and exhibit the capacity to
differentiate into T cells. In addition, SP cells have been
demonstrated to have a special type of stem cell phenotype in
several normal tissues, including the hematopoietic system
(51) and breast epithelium tissue
(52). SP cells have also been
identified in numerous types of cancer tissue, including leukemia
(53), multiple myeloma (54) and breast cancer tissues (55). Furthermore, SP cells can
trans-differentiate into other tissue cells. For example, bone
marrow-derived SP cells can promote respiratory damage repair in
mice in vivo (56). Although
SP markers can be viewed as a common stem cell phenotype marker,
some debate still exists. ATP binding cassette subfamily G member 2
(ABCG2) is a membrane receptor, which has a pump function that can
remove toxic substances from the cytoplasm, contributing to the
phenotype of SP cells. ABCG2 can protect cancer and normal stem
cells from X-ray damage. Additionally, ABCG2 has been demonstrated
to be a phenotype determinant of SP cells (57). Patrawala et al (58) reported that ~30% of cultured human
cancer cells and xenograft tumors possess a detectable SP. Purified
SP cells from U373 glioma cells, MCF7 breast cancer cells and a
xenograft prostate tumor were more tumorigenic compared with the
corresponding non-SP cells. These SP cells also possess intrinsic
stem cell properties, as they generate non-SP cells in vivo, can be
further transplanted, and preferentially express specific stemness
genes, including Notch-1 and β-catenin (58). Since the SP phenotype is primarily
mediated by ABCG2, an ATP-binding cassette half-transporter
associated with multidrug resistance, the tumorigenicity of ABCG2+
and ABCG2- cancer cells was investigated. Although SP cells
exhibited an increased ABCG2 mRNA expression compared with the
non-SP cells, all examined cancer cells and xenograft tumors
expressed ABCG2 in a small fraction (0.5–3.0%) of the cells, highly
purified ABCG2+ cancer cells have a very similar tumorigenicity to
the ABCG2- cancer cells (58). A
mechanistic study has indicated that ABCG2 expression is associated
with proliferation and ABCG2+ cancer cells can generate ABCG2-
cells (58). However, ABCG2- cancer
cells can also generate ABCG2+ cells. Furthermore, the ABCG2-
cancer cells form more and larger clones in long-term clonal
analyses, and the ABCG2- population preferentially expresses
several stemness genes (58). These
results suggested that, although the SP is enriched with
tumorigenic cancer stem-like cells and the ABCG2 expression
primarily identifies fast-cycling tumor progenitors, the ABCG2-
population contains primitive cancer stem-like cells. Therefore,
ABCG2+ cancer cells are not equal to SP cells.
Several studies have isolated CSCs from ESCC using
SP cells. For example, in 2008, Zhang et al (59) used flow cytometry to serially sort
stem-like SP cells, demonstrating that radioresistant cell lines
included more SP cells than parent cell lines. Another Chinese
research group reported that the number of SP cells was increased
in tumor spheres, when compared with adherent cells (60). A previous study by Yue et al
(61) demonstrated that stem
cell-like SPs in ESCC were a cause of chemotherapy resistance and
metastasis. The SP subpopulation was detected using Hoechst 33342
staining in five ESCC cell lines, OE19, OE21, OE33, PT1590 and
LN1590. Chemotherapy-resistant cell lines were developed following
long-term exposure to 5-fluorouracil (5-FU) and cisplatin, and were
validated by an analysis of resistance markers, thymidylate
synthase and excision repair cross-complement gene 1. While the
LN1590 and PT1590 cell lines did not exhibit detectable SP cells,
OE19, OE21 and OE33 cells were observed to contain varying levels
of SP cells. With the increasing duration of 5-FU or cisplatin
therapy, the SP subpopulation substantially emerged in the PT1590
and LN1590 cell lines. SP OE19 cells displayed a significantly
higher tumorigenicity compared with non-SP OE19 cells, following
the subcutaneous injection of tumor cells in vivo. SP cells
isolated from OE19 and OE19 5-FU-resistant cells were subsequently
analyzed by an epithelial-to-mesenchymal transition (EMT)
polymerase chain reaction array. Notably, the SP fraction of 5-FU
resistant OE19 cells led to a marked upregulation of EMT-associated
genes, when compared with the SP fraction of OE19 cells (61). These results provided the following
evidence: i) The proportion of SP cells is different in ESCC; ii)
SP cells exhibit stem cell-like properties and are associated with
chemotherapy resistance; and iii) long-term exposure to cytotoxic
drug selects for SP cells with an upregulated EMT gene profile,
which may be the source of systemic disease relapse. Further
investigations are necessary to target these EMT-associated SP
cells in ESCC.
The fourth method for isolating CSCs is the
utilization of chemotherapy resistance markers, including ALDH1.
ALDH1 has been demonstrated to be a candidate CSC biomarker
(62) and has been associated with
CSC-like characteristics in numerous types of human cancer
(63). Early in 2005, ALDH1 was
reported to be expressed at a higher level in CD34+ cells, when
compared with differentiated cells, highlighting the important role
of ALDH1 in normal hemopoietic stem cell biology (64). Seigel et al (65) reported that <1% of retinoblastoma
cells exhibit immunoreactivity against the stem cell markers ABCG2
and ALDH1, suggesting that CSCs account for a small fraction of the
cells in human cancer. In the following decade, ALDH1 was
identified to be overexpressed in various other CSCs, including
those of breast (35), lung
(66) and pancreatic cancer
(67), and can therefore be regarded
as a common CSC marker. In ESCC, the expression of ALDH1 protein in
the nucleus of ESCCs is associated with lymph node metastasis and a
poor survival, suggesting that ALDH1 may be involved in the
aggressive behavior of ESCC (68).
Although there is no direct evidence to validate that ALDH1+ cells
in ESCC are cancer stem-like cells, several studies hypothesize
this to be the case. For example, Song et al (69) demonstrated that the short hairpin RNA
(shRNA)-mediated knockdown of yes-associated protein 1 (YAP1) or
SRY-box (SOX) 9 in transformed cells attenuates CSC phenotypes
in vitro and tumorigenicity in vivo in ESCC. The
small-molecule inhibitor of YAP1, verteporfin, blocks ESCC CSC
properties in cells with high YAP1 and a high proportion of ALDH1+
cells (69). Furthermore, Chen et
al (70) recently revealed that
ALDH1 staining was positively linked to a higher clinical stage,
higher loco-regional failure rate, and shorter survival time in
ESCC.
Following the isolation of a CSC, its identification
and verification is imperative. Due to the abundance of isolation
methods, different isolation methods have been reported to identify
the same CSC with a different cellular phenotype (71). A common method to identify CSCs is to
examine the stemness of isolated cells based on three major
characteristics: i) Unlimited proliferation ability; ii)
self-renewal ability; and iii) strong tumorigenesis (71). Since the concept of CSCs was first
introduced, scientists have attempted to isolate CSCs from human
ESCC cell lines and tissues.
The three aforementioned CSC characteristics have
been applied to investigate the stemness of isolated ESCC cells.
Yang et al (72) identified
leucine zipper and EF-hand containing transmembrane protein 1
(LETM1) as a marker of cancer stem-like cells and predicted a poor
prognosis in ESCC based on the protein expression of LETM1, which
was positively correlated with CSC markers in ESCC cell lines. Liu
et al (73) reported that
Cripto-1 could act as a functional marker of cancer stem-like cells
and predict prognosis in patients with ESCC. The suppression of the
Cripto-1 expression by shRNA markedly reduced the expression of
stemness- and EMT-associated genes, in addition to their
self-renewal capabilities in vitro, and tumorigenicity and
metastasis in vivo in ESCC cells. Aldefluor, a fluorescent
substrate of aldehyde dehydrogenase, has been used to isolate CSCs;
however, it was not as easy to observe the interaction between CSCs
and non-CSCs (74). Almanaa et
al (75) introduced the
attached-cell Aldefluor method to detect CSCs in ESCC cell lines.
Almanaa et al (75) also
demonstrated a novel method for generating and growing tumor
spheres without the growth factor supplements normally used in the
medium for their formation. ALDH-1 as a stem cell marker in
patients with resectable ESCC, ALDH-1 expression can predict the
response or resistance to preoperative chemoradiation (76).
From the signaling pathways dysregulated in ESCC,
certain stem-like characteristics have been identified to be
associated with CSCs in ESCC. Zhang et al (60) reported that R cells in ESCC exhibited
CSC-like traits and highly expressed β-catenin. In addition, a
cyclooxygenase-2 inhibitor NS398 had a radiosensitization effect on
R cells via the downregulation of β-catenin (89). SB525334, a transforming growth
factor-β 1 (TGF-β1) inhibitor, has been identified to markedly
inhibit the migration and invasion of sphere-forming stem-like
cells, yet had no effect on their sphere-forming ability, this
suggested that TGF-β1 may be a biomarker for metastasis in ESCC
stem cells; however, was not required for self-renewal (90). TGF-β1 can be regarded as a molecular
target for the eradication of ESCC stem cell metastasis (90). In addition to its implication in ESCC
stem cells, TGF-β1 has been reported to regulate the stemness of
other CSCs, including those of liver cancer (91,92).
TGF-β1 can also be induced by a bispecific EpCAM+ CD3 antibody to
be released from pre-activated lymphocytes and target EpCAM+
retinoblastoma stem cells (93).
Although immunotherapy applied to ESCC is not yet effective, we
anticipate that in the near future, CSCs may become a highly
efficient therapeutic target cell type for immunotherapy in ESCC.
The hedgehog (Hh) pathway is involved in CSC maintenance in various
types of tumor, and Glioma-associated oncogene homolog 1 (Gli1) is
a key mediator of the Hh pathway (94,95).
Yang et al (96) reported
Gli1 to be expressed in 28.3% of ESCC and to be an indicator of
ESCC stem cells, and its expression was correlated with the
expression of the stemness genes SOX9 and CD44. Gli1, CD44 and SOX9
were highly expressed in poorly differentiated ESCC cell lines,
including TE8 and TE1 cells. Notably, the Gli1 expression was
positively associated with distant metastasis, increased
microvessel density and the expression of cell cycle regulators,
including p21, cyclin D1, cyclin E1 and NF-κB. Fujiwara et
al (97) used a
NanoCulture® Plate for 3-dimensional cell culture to
prepare spheroids from ESCC cells and demonstrated that these
spheroids possessed stem cell-like characteristics. Li et al
(unpublished data) also utilized serum-free medium supplemented
with growth factors to culture ESCC microspheres and observed that
these sphere cells displayed a number of CSC-like characteristics.
These spheroid cells highly expressed the ALDH1 enzyme, and the
mRNA expression of the stem cell-associated genes SOX2, NANOG,
OCT3/4 and LIN28 were also elevated (97). Notably, several common stem cell
surface markers, including CD44, CD133, CD338 (ABCG2), CD318
(CDCP1) and CD326 (EpCAM), were not observed be elevated. However,
Kanamoto et al (98) reported
that a single dose of irradiation can induce EMT and CD44
expression, conferring ESCC to acquire cancer stem-like cell
properties. miRNAs were also identified to be involved in the
signaling pathways of ESCC cancer stem cells. Downregulation of
miRNA-644a was suggested to promote ESCC aggressiveness and stem
cell-like phenotype via dysregulation of paired like homeodomain 2
(99).
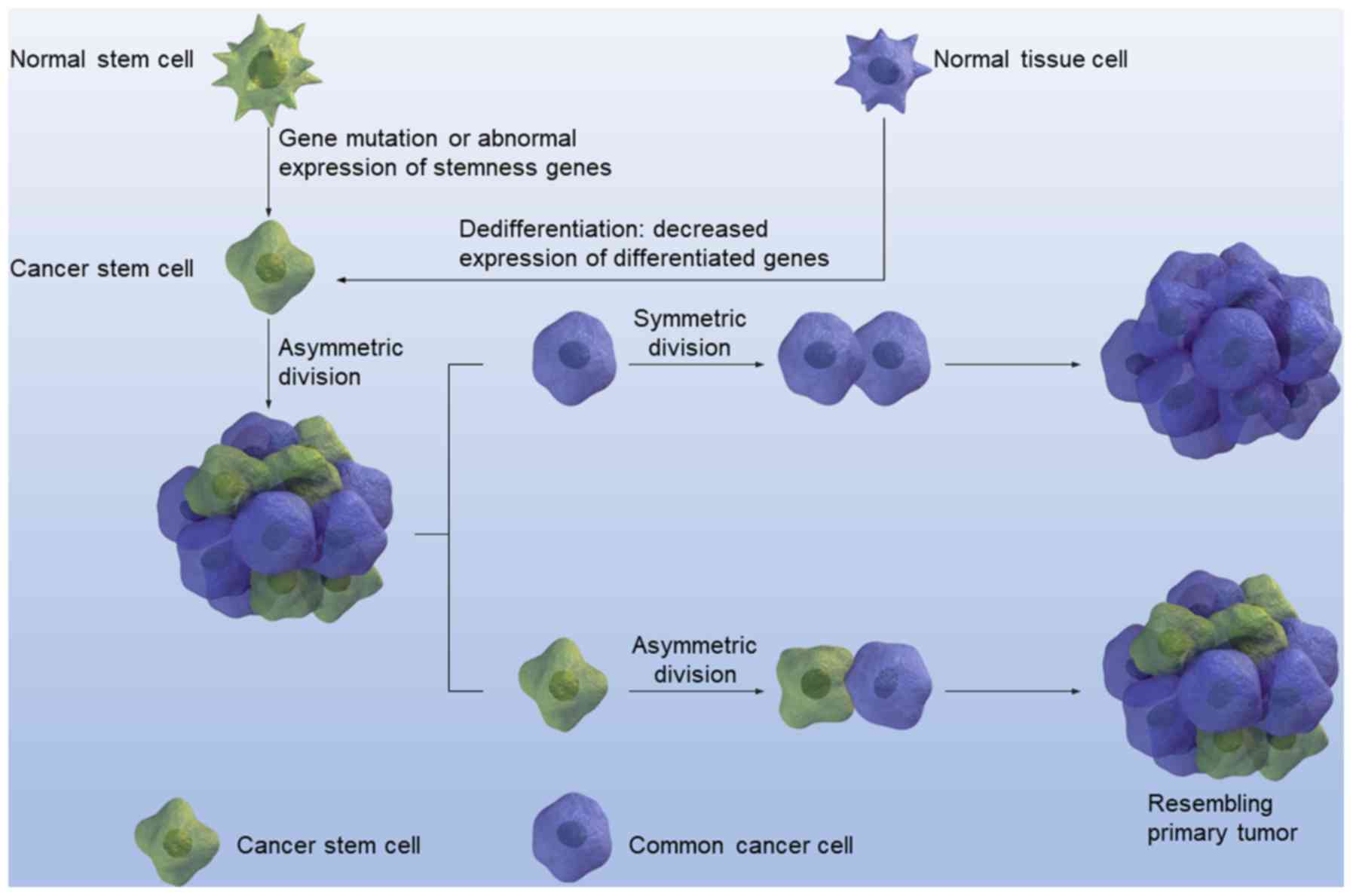
Until now, stem cells could be classified into
several categories, including embryonic or totipotent stem cells,
pluripotent stem cells, and tissue-specific stem or precursor
cells, thus constituting a hierarchy of stem cells (100). CSCs can also be organized in a
hierarchical fashion (101). This
theory was initially proposed by Bonnet and Dick (12) in a pioneering study that isolated and
identified CSCs in human acute myeloid leukemia. Cancer research
then began to focus on stem cells. To a certain extent, a large
portion of the CSC research that followed was derived from this
initial study. As with normal stem cells, different signaling
pathways such as Wnt and Hedgehog signaling participate in the
maintenance and regulation of CSCs (102,103).
Due to the fact that CSCs are aberrant stem cells, stemness genes
are abnormally upregulated or downregulated in CSCs. In addition,
CSCs are cancer cells; thus these signaling molecules should also
be specific to cancer and dysregulated in cancer cells. Therefore,
stemness genes or signaling pathways dysregulated in CSCs should
possess two properties: i) The ability to signal the activation of
downstream targets that contribute to the maintenance of the three
major characteristics of stem cells; and ii) the ability to
maintain malignant characteristics (104).
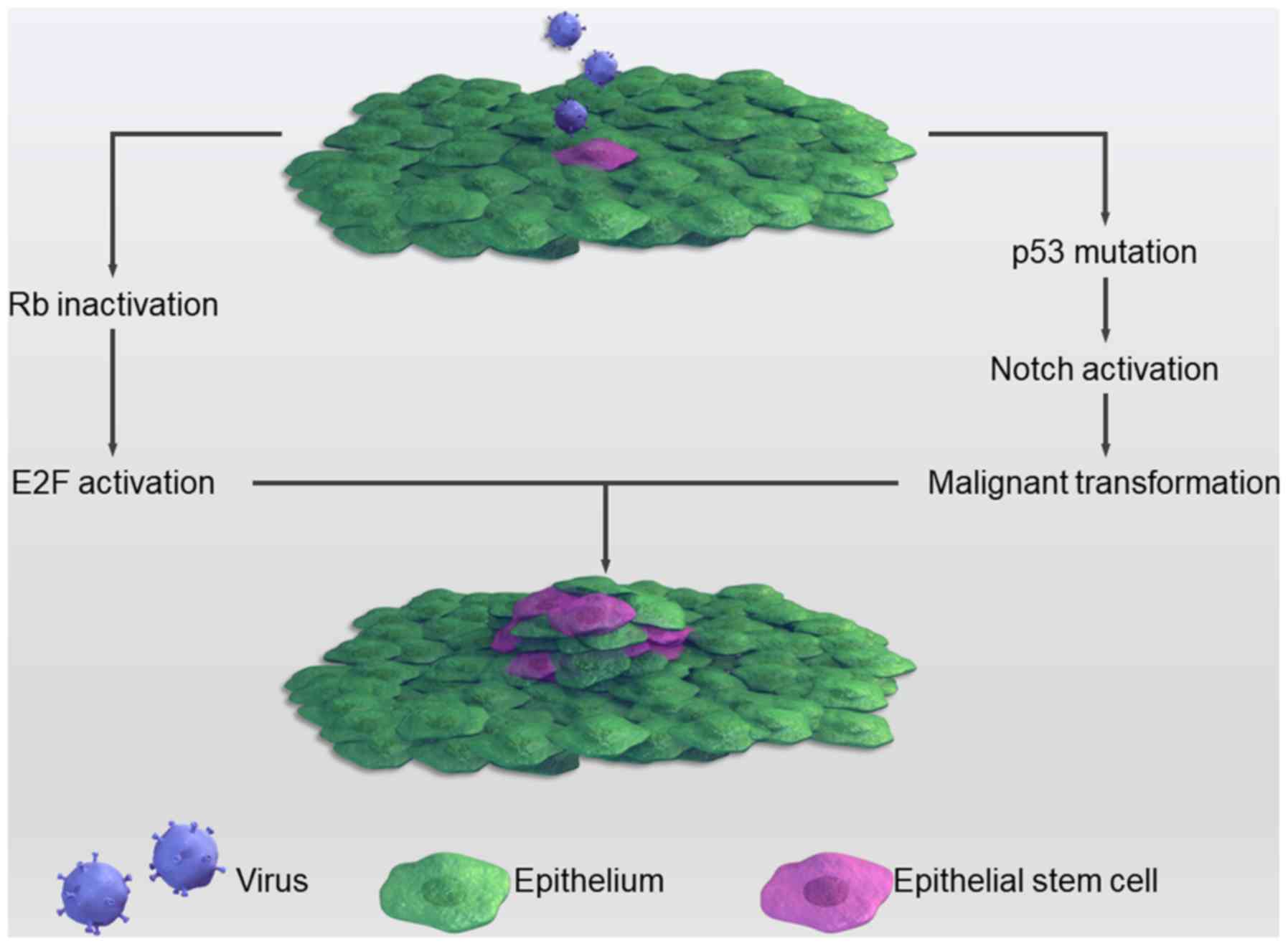
Although the mechanism through which viruses
contribute to the stemness of cancers remains to be completely
elucidated, several studies have attempted to investigate it. For
example, the HPV E6 protein can dysregulate p53, which has been
observed to be closely associated with CSCs (120). p53 is ‘the guardian of the genome’,
since genome mutation or instability leads to tumor occurrence
(121). Recently, several studies
have implied that p53-deficiency confers a CSC-like phenotype
(122,123). In cases of cell damage, p53 can
lead to cell growth arrest and apoptosis with expression level
changes of Nanog and Notch via binding to the promoter region of
these two stemness genes. Under normal circumstances, p53 inhibits
the Notch pathway; however, when infected with HPV this inhibition
is relieved, causing a more proliferative state through the
activation of Notch (124). Another
common tumor suppressor, the retinoblastoma (Rb) gene, is often
mutated in several types of tumors and is associated with CSCs. HPV
can inactivate Rb, thereby causing cancer. Inactivated Rb
predisposes cells to a highly proliferative state through binding
to and activating the E2F protein. The E2F protein is a
transcriptional factor that promotes cell division (125). The HPV E7 protein forms complexes
with Rb, which frees Rb from E2F and promotes unregulated cell
proliferation (126). This process
is summarized in Fig. 2.
Numerous viruses, including HPV, herpes simplex
virus, cytomegalovirus and Epstein-Barr virus, can infect the
esophageal epithelium (127–129).
As a result, infection with these viruses may contribute to the
pathogenesis of ESCCs. HPV was the first virus to be associated
with the pathogenesis of ESCC. In 1990, a study performed in an
area of China with a high ESCC-associated morbidity rate revealed
that HPV was associated with the carcinogenesis of ESCC, as
demonstrated by histological analysis and in situ DNA
hybridization (130). Subsequently,
a number of studies (131–133) have reinforced this association,
although to the best of our knowledge, evidence for HPV as an
initiating factor has yet to be established. Furthermore, to the
best of our knowledge, the molecular mechanism of HPV
infection-associated carcinogenesis in esophagus mucosa and ESCC
tissue is unclear. In 2016, Xi et al (134) utilized a lentiviral vector to
stably overexpress the high-risk HPV 16, E6 and E7 proteins, and
conferred stem-like characteristics on an ESCC cell line, thus
providing direct evidence to support HPV participation in the
formation of CSCs. However, in ESCC low-morbidity areas, such as
Italy, Syrjänen (135) reported
that HPV cannot be detected in the ESCC tissue. This result was
consistent with those of other studies (136–138),
suggesting that, in ESCC low-morbidity areas, HPV is not crucial
for esophageal epithelium carcinogenesis. A case-controlled study
in Zambia implied that HPV infection was not a risk factor;
however, HIV infection and domestic smoke exposure were (139). Consequently, a conclusion can be
drawn that HPV alone is insufficient for the occurrence of ESCC,
neither is it essential. HPV infection may be a promoting factor in
the pathogenesis of ESCC, but not an initiating one.
In conclusion, CSCs are regarded as target cells
that when effectively eradicated have the potential to treat human
cancer. However, there are currently no therapeutics or treatments
aimed at eradicating CSCs. Such is the case in ESCC, although CSCs
have been identified in ESCC. Due to a lack of early detection
methods in ESCC, the disease is frequently diagnosed at the mid or
late stages of progression, accompanied by lymph-node metastasis.
Systemic therapy, such as targeting CSCs, may ultimately prove
crucial for the eradication of this malignant disease. CSCs have
been isolated and identified in ESCC for over a decade; however,
effective therapies targeting these cells are lacking. Further
studies along these lines of investigation are urgently required in
order to identify novel and effective therapeutics for ESCC.
Not applicable.
The present study was funded by grants from the
Natural Science Funding of China (grant no. 71473073) and the
Department of Science and Technology of Hubei province (grant nos.
2015CFB470 and 2015CFC773).
All data generated or analyzed during the present
study are included in this published article.
QW, ZW, CB and WL wrote the manuscript. HH, YS and
ZC contributed to the review of the literature. HZ and ZN
contributed to the concept of the manuscript. All authors read and
approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China
2015. Ca Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang Y, Qu S, Zhu G, Wang F, Liu R, Shen
X, Viola D, Elisei R, Puxeddu E, Fugazzola L, et al: BRAF V600E
mutation-assisted risk stratification of solitary intrathyroidal
papillary thyroid cancer for precision treatment. J Natl Cancer
Inst. 110:362–370. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Leonard KL and Wazer DE: Genomic assays
and individualized treatment of ductal carcinoma in situ in
the era of value-based cancer care. J Clin Oncol. 34:3953–2955.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dos Santos M, Brachet PE, Chevreau C and
Joly F: Impact of targeted therapies in metastatic renal cell
carcinoma on patient-reported outcomes: Methodology of clinical
trials and clinical benefit. Cancer Treat Rev. 53:53–60. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen DS and Mellman I: Elements of cancer
immunity and the cancer-immune set point. Nature. 541:321–330.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Başaran GA, Twelves C, Diéras V, Cortés J
and Awada A: Ongoing unmet needs in treating estrogen
receptor-positive/HER2-negative metastatic breast cancer. Cancer
Treat Rev. 63:144–155. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bourke L, Kirkbride P, Hooper R, Rosario
AJ, Chico TJ and Rosario DJ: Endocrine therapy in prostate cancer:
Time for reappraisal of risks, benefits and cost-effectiveness? Br
J Cancer. 108:9–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Han SH, Kim JW, Kim M, Kim JH, Lee KW, Kim
BH, Oh HK, Kim DW, Kang SB, Kim H and Shin E: Prognostic
implication of ABC transporters and cancer stem cell markers in
patients with stage III colon cancer receiving adjuvant FOLFOX-4
chemotherapy. Oncol Lett. 17:5572–5580. 2019.PubMed/NCBI
|
|
10
|
Xu PP, Fu D, Li JY, Hu JD, Wang X, Zhou
JF, Yu H, Zhao X, Huang YH, Jiang L, et al: Anthracycline dose
optimisation in patients with diffuse large B-cell lymphoma: A
multicentre, phase 3, randomised, controlled trial. Lancet
Haematol. 6:e328–e337. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rous P: The relations of embryonic tissue
and tumor in mixed grafts. J Exp Med. 13:239–247. 1911. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
ZD: CarcinogenesisMedicine. Zaridze D.G.:
pp. 1–567. 2004
|
|
14
|
Dick JE and Tsvee L: Biology of normal and
acute myeloid leukemia stem cells. Int J Hematol. 82:389–396. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wright MH, Calcagno AM, Salcido CD,
Carlson MD, Ambudkar SV and Lyuba V: Brca1 breast tumors contain
distinct CD44+/CD24- and CD133+ cells with cancer stem cell
characteristics. Breast Cancer Res. 10:R102008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Collins AT, Berry PA, Hyde C, Stower MJ
and Maitland NJ: Prospective identification of tumorigenic prostate
cancer stem cells. Cancer Res. 65:10946–10951. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Odoux C, Fohrer H, Hoppo T, Guzik L, Stolz
DB, Lewis DW, Gollin SM, Gamblin TC, Geller DA and Lagasse E: A
stochastic model for cancer stem cell origin in metastatic colon
cancer. Cancer Research. 68:6932–6941. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vermeulen L, Todaro M, de Sousa Mello F,
Sprick MR, Kemper K, Perez Alea M, Richel DJ, Stassi G and Medema
JP: Single-cell cloning of colon cancer stem cells reveals a
multi-lineage differentiation capacity. Proc Natl Acad Sci USA.
105:13427–13432. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang H, Hao C, Wang H, Shang H and Li Z:
Carboxypeptidase A4 promotes proliferation and stem cell
characteristics of hepatocellular carcinoma. Int J Exp Pathol.
100:133–138. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li X, Zhang Y, Ding J, Wang M, Li N, Yang
H, Wang K, Wang D, Lin PP, Li M, et al: Clinical significance of
detecting CSF-derived tumor cells in breast cancer patients with
leptomeningeal metastasis. Oncotarget. 9:2705–2714. 2017.PubMed/NCBI
|
|
22
|
Lin Y, Totsuka Y, He Y, Kikuchi S, Qiao Y,
Ueda J, Wei W, Inoue M and Tanaka H: Epidemiology of esophageal
cancer in Japan and China. J Epidemiol. 23:233–242. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lao-Sirieix P and Fitzgerald RC: Screening
for oesophageal cancer. Nat Rev Clin Oncol. 9:278–287. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pennathur A, Gibson MK, Jobe BA and
Luketich JD: Oesophageal carcinoma. Lancet. 381:400–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen M, Liu P, Chen Y, Chen Z, Shen M, Liu
X, Li X, Lin Y, Yang R, Ni W, et al: Primary tumor regression
patterns in esophageal squamous cell cancer treated with definitive
chemoradiotherapy and implications for surveillance schemes. Cancer
Manag Res. 11:3361–3369. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vira D, Basak SK, Veena MS, Wang MB, Batra
RK and Srivatsan ES: Cancer stem cells, microRNAs, and therapeutic
strategies including natural products. Cancer Metastasis Rev.
31:733–751. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fu W, Lei C, Yu Y, Liu S, Li T, Lin F, Fan
X, Shen Y, Ding M, Tang Y, et al: EGFR/Notch antagonists enhance
the response to inhibitors of the PI3K-Akt pathway by decreasing
tumour-initiating cell frequency. Clin Cancer Res. 25:2835–2847.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jia ZF, Wu YH, Cao DH, Cao XY, Jiang J and
Zhou BS: Polymorphisms of cancer stem cell marker gene CD133 are
associated with susceptibility and prognosis of gastric cancer.
Future Oncol. 13:979–989. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kalantari E, Asgari M, Nikpanah S,
Salarieh N, Lari MH and Madjd Z: Co-expression of putative cancer
stem cell markers CD44 and CD133 in prostate carcinomas. Pathol
Oncol Res. 23:793–802. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yan Y, Zuo X and Wei D: Concise review:
Emerging role of CD44 in cancer stem cells: A promising biomarker
and therapeutic target. Stem Cells Transl Med. 4:1033–1043. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liou GY: CD133 as a regulator of cancer
metastasis through the cancer stem cells. Int J Biochem Cell Biol.
106:1–7. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tang KH, Dai YD, Tong M, Chan YP, Kwan PS,
Fu L, Qin YR, Tsao SW, Lung HL, Lung ML, et al: A CD90(+)
tumor-initiating cell population with an aggressive signature and
metastatic capacity in esophageal cancer. Cancer Res. 73:2322–2332.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Moreira MP, da Conceição Braga L and Silva
LM: STAT3 as a promising chemoresistance biomarker associated with
the CD44+/high/CD24-/low/ALDH+ BCSCs-like subset of the
triple-negative breast cancer (TNBC) cell line. Exp Cell Res.
363:283–290. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nguyen PH, Giraud J, Staedel C,
Chambonnier L, Dubus P, Chevret E, Bœuf H, Gauthereau X, Rousseau
B, Fevre M, et al: All-trans retinoic acid targets gastric cancer
stem cells and inhibits patient-derived gastric carcinoma tumor
growth. Oncogene. 35:5619–5628. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Erb HHH, Guggenberger F, Santer FR and
Culig Z: Interleukin-4 induces a CD44high/CD49bhigh PC3
subpopulation with tumor-initiating characteristics. J Cell
Biochem. 119:4103–4112. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ogawa T, Hirohashi Y, Murai A, Nishidate
T, Okita K, Wang L, Ikehara Y, Satoyoshi T, Usui A, Kubo T, et al:
ST6GALNAC1 plays important roles in enhancing cancer stem
phenotypes of colorectal cancer via the Akt pathway. Oncotarget.
8:112550–112564. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang HH, Liao CC, Chow NH, Huang LL,
Chuang JI, Wei KC and Shin JW: Whether CD44 is an applicable marker
for glioma stem cells. Am J Transl Res. 9:4785–4806.
2017.PubMed/NCBI
|
|
40
|
Zhao JS, Li WJ, Ge D, Zhang PJ, Li JJ, Lu
CL, Ji XD, Guan DX, Gao H, Xu LY, et al: Tumor initiating cells in
esophageal squamous cell carcinomas express high levels of CD44.
PLoS One. 6:e214192011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Matsuya Y: A serum-free culture medium for
the minor inoculum of L line cells. Tohoku J Exp Med. 86:1–8. 1965.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Haylock DN, To LB, Dowse TL, Juttner CA
and Simmons PJ: Ex vivo expansion and maturation of peripheral
blood CD34+ cells into the myeloid lineage. Blood. 80:1405–1412.
1992.PubMed/NCBI
|
|
43
|
Petzer AL, Zandstra PW, Piret JM and Eaves
CJ: Differential cytokine effects on primitive (CD34+CD38-) human
hematopoietic cells: Novel responses to Flt3-ligand and
thrombopoietin. J Exp Med. 183:2551–2558. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Möbest D, Goan SR, Junghahn I, Winkler J,
Fichtner I, Hermann M, Becker M, de Lima-Hahn E and Henschler R:
Differential kinetics of primitive hematopoietic cells assayed
in vitro and in vivo during serum-free suspension
culture of CD34+ blood progenitor cells. Stem Cells. 17:152–161.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jimenez-Pascual A, Hale JS, Kordowski A,
Pugh J, Silver DJ, Bayik D, Roversi G, Alban TJ, Rao S, Chen R, et
al: ADAMDEC1 maintains a growth factor signaling loop in cancer
stem cells. Cancer Discov. (pii): CD-18-1308. 2019.PubMed/NCBI
|
|
46
|
Abbaszadegan MR, Bagheri V, Razavi MS,
Momtazi AA, Sahebkar A and Gholamin M: Isolation, identification
and characterization of cancer stem cells: A review. J Cell
Physiol. 232:2008–2018. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xiao G, Li X, Li G, Zhang B, Xu C, Qin S,
Du N, Wang J, Tang SC, Zhang J, et al: MiR-129 blocks estrogen
induction of NOTCH signaling activity in breast cancer stem-like
cells. Oncotarget. 8:103261–103273. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Trisciuoglio D, Tupone MG, Desideri M, Di
Martile M, Gabellini C, Buglioni S, Pallocca M, Alessandrini G,
D'Aguanno S and Del Bufalo D: BCL-XL overexpression promotes tumor
progression-associated properties. Cell Death Dis. 8:32162017.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang JL, Yu JP, Sun ZQ and Sun SP:
Radiobiological characteristics of cancer stem cells from
esophageal cancer cell lines. World J Gastroenterol.
20:18296–18305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Goodell MA, Rosenzweig M, Kim H, Marks DF,
DeMaria M, Paradis G, Grupp SA, Sieff CA, Mulligan RC and Johnson
RP: Dye efflux studies suggest that hematopoietic stem cells
expressing low or undetectable levels of CD34 antigen exist in
multiple species. Nat Med. 3:1337–1345. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Parmar K, Sauk-Schubert C, Burdick D,
Handley M and Mauch P: Sca+CD34- murine side population cells are
highly enriched for primitive stem cells. Exp Hematol. 31:244–250.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Alvi AJ, Clayton H, Joshi C, Enver T,
Ashworth A, Vivanco Md, Dale TC and Smalley MJ: Functional and
molecular characterisation of mammary side population cells. Breast
Cancer Research. 5:R1–R8. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gross E, L'Faqihiolive FE, Ysebaert L,
Brassac M, Struski S, Kheirallah S, Fournié JJ, Laurent G and
Quillet-Mary A: B-chronic lymphocytic leukemia chemoresistance
involves innate and acquired leukemic side population cells.
Leukemia. 24:1885–1892. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Du J, Liu S, He J, Liu X, Qu Y, Yan W, Fan
J, Li R, Xi H, Fu W, et al: MicroRNA-451 regulates stemness of side
population cells via PI3K/Akt/mTOR signaling pathway in multiple
myeloma. Oncotarget. 6:14993–15007. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Britton KM, Kirby JA, Lennard TW and
Meeson AP: Cancer stem cells and side population cells in breast
cancer and metastasis. Cancers. 3:2106–2130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Macpherson H, Keir P, Webb S, Samuel K,
Boyle S, Bickmore W, Forrester L and Dorin J: Bone marrow-derived
SP cells can contribute to the respiratory tract of mice in
vivo. J Cell Sci. 118:2441–2450. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Shimoda M, Ota M and Okada Y: Isolation of
cancer stem cells by side population method. Methods Mol Biol.
1692:49–59. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Patrawala L, Calhoun T,
Schneider-Broussard R, Zhou J, Claypool K and Tang DG: Side
population is enriched in tumorigenic, stem-like cancer cells,
whereas ABCG2+ and ABCG2-cancer cells are similarly tumorigenic.
Cancer Res. 65:6207–6019. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang X, Komaki R, Wang L, Fang B and
Chang JY: Treatment of radioresistant stem-like esophageal cancer
cells by an apoptotic gene-armed, telomerase-specific oncolytic
adenovirus. Clin Cancer Res. 14:2813–2823. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhang G, Ma L, Xie YK, Miao XB and Jin C:
Esophageal cancer tumorspheres involve cancer stem-like populations
with elevated aldehyde dehydrogenase enzymatic activity. Mol Med
Rep. 6:519–524. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yue Z, Qi B, Bettina S, Zhao L, Mysliwietz
J, Ellwart J, Renner A, Hirner H, Niess H, Camaj P, et al: Stem
cell-like side populations in esophageal cancer: A source of
chemotherapy resistance and metastases. Stem Cells Dev. 23:180–192.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chen J, Xia Q, Jiang B, Chang W, Yuan W,
Ma Z, Liu Z and Shu X: Prognostic value of cancer stem cell marker
ALDH1 expression in colorectal cancer: A systematic review and
meta-analysis. PLoS One. 10:e01451642015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhou Y, Wang Y, Ju X, Lan J, Zou H, Li S,
Qi Y, Jia W, Hu J, Liang W, et al: Clinicopathological significance
of ALDH1A1 in lung, colorectal, and breast cancers: A
meta-analysis. Biomark Med. 9:777–790. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ferrell CM, Dorsam ST, Ohta H, Humphries
RK, Derynck MK, Haqq C, Largman C and Lawrence HJ: Activation of
stem-cell specific genes by HOXA9 and HOXA10 homeodomain proteins
in CD34+ human cord blood cells. Stem Cells. 23:644–655. 2010.
View Article : Google Scholar
|
|
65
|
Seigel GM, Campbell LM, Narayan M and
Gonzalez-Fernandez F: Cancer stem cell characteristics in
retinoblastoma. Mol Vis. 11:729–737. 2005.PubMed/NCBI
|
|
66
|
Macdonagh L, Gallagher MF, Ffrench B,
Gasch C, Breen E, Gray SG, Nicholson S, Leonard N, Ryan R, Young V,
et al: Targeting the cancer stem cell marker, aldehyde
dehydrogenase 1, to circumvent cisplatin resistance in NSCLC.
Oncotarget. 8:72544–72563. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Fu Z, Chen C, Zhou Q, Wang Y, Zhao Y, Zhao
X, Li W, Zheng S, Ye H, Wang L, et al: LncRNA HOTTIP modulates
cancer stem cell properties in human pancreatic cancer by
regulating HOXA9. Cancer Lett. 410:68–81. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ji Y, Li X, Li Y, Zhong Y, Cao J, Xu R,
Wang J, Zhou F, Li X, Yu D, et al: Aldehyde dehydrogenase-1
expression predicts unfavorable outcomes in patients with
esophageal squamous cell carcinoma. Anticancer Res. 36:343–349.
2016.PubMed/NCBI
|
|
69
|
Song S, Ajani JA, Honjo S, Maru DM, Chen
Q, Scott AW, Heallen TR, Xiao L, Hofstetter WL, Weston B, et al:
Hippo coactivator YAP1 upregulates SOX9 and endows esophageal
cancer cells with stem-like properties. Cancer Res. 74:4170–4182.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Chen MF, Chen PT, Lu MS and Chen WC: Role
of ALDH1 in the prognosis of esophageal cancer and its relationship
with tumor microenvironment. Mol Carcinog. 57:78–88. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Akbarzadeh M, Maroufi NF, Tazehkand AP,
Akbarzadeh M, Bastani S, Safdari R, Farzane A, Fattahi A, Nejabati
HR, Nouri M and Samadi N: Current approaches in identification and
isolation of cancer stem cells. J Cell Physiol. Feb 11–2019.doi:
10.1002/jcp.28271 (Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Yang Z, Ni W, Cui C, Qi W, Piao L and Xuan
Y: Identification of LETM1 as a marker of cancer stem-like cells
and predictor of poor prognosis in esophageal squamous cell
carcinoma. Hum Pathol. 81:148–156. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu Q, Cui X, Yu X, Bian BS, Qian F, Hu
XG, Ji CD, Yang L, Ren Y, Cui W, et al: Cripto-1 acts as a
functional marker of cancer stem-like cells and predicts prognosis
of the patients in esophageal squamous cell carcinoma. Mol Cancer.
16:812017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Cabrera MC, Hollingsworth RE and Hurt EM:
Cancer stem cell plasticity and tumor hierarchy. World J Stem
Cells. 7:27–36. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Almanaa TN, Geusz ME and Jamasbi RJ: A new
method for identifying stem-like cells in esophageal cancer cell
lines. J Cancer. 4:536–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Ajani JA, Wang X, Song S, Suzuki A, Taketa
T, Sudo K, Wadhwa R, Hofstetter WL, Komaki R, Maru DM, et al:
ALDH-1 expression levels predict response or resistance to
preoperative chemoradiation in resectable esophageal cancer
patients. Mol Oncol. 8:142–149. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Chang L, Graham P, Hao J, Ni J, Deng J,
Bucci J, Malouf D, Gillatt D and Li Y: Cancer stem cells and
signaling pathways in radioresistance. Oncotarget. 7:11002–11017.
2016.PubMed/NCBI
|
|
78
|
Lynam-Lennon N, Heavey S, Sommerville G,
Bibby BA, Ffrench B, Quinn J, Gasch C, O'Leary JJ, Gallagher MF,
Reynolds JV and Maher SG: MicroRNA-17 is downregulated in
esophageal adenocarcinoma cancer stem-like cells and promotes a
radioresistant phenotype. Oncotarget. 8:11400–11413. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chen KH, Guo Y, Li L, Qu S, Zhao W, Lu QT,
Mo QY, Yu BB, Zhou L, Lin GX, et al: Cancer stem cell-like
characteristics and telomerase activity of the nasopharyngeal
carcinoma radioresistant cell line CNE-2R. Cancer Med. 7:4755–4764.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Chen Y, Jiang T, Mao A and Xu J:
Esophageal cancer stem cells express PLGF to increase cancer
invasion through MMP9 activation. Tumour Biol. 35:12749–12755.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Tsai ST, Wang PJ, Liou NJ, Lin PS, Chen CH
and Chang WC: ICAM1 is a potential cancer stem cell marker of
esophageal squamous cell carcinoma. PLoS One. 10:e01428342015.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Sauzay C, Voutetakis K, Chatziioannou A,
Chevet E and Avril T: CD90/Thy-1, a cancer-associated cell surface
signaling molecule. Front Cell Dev Biol. 7:662019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Ji N, Yu JW, Ni XC, Wu JG, Wang SL and
Jiang BJ: Bone marrow-derived mesenchymal stem cells increase drug
resistance in CD133-expressing gastric cancer cells by regulating
the PI3K/AKT pathway. Tumor Biol. 37:14637–14651. 2016. View Article : Google Scholar
|
|
84
|
Fan H and Lu S: Fusion of human bone
hemopoietic stem cell with esophageal carcinoma cells didn't
generate esophageal cancer stem cell. Neoplasma. 61:540–545. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Mo JS, Park HW and Guan KL: The Hippo
signaling pathway in stem cell biology and cancer. EMBO Rep.
15:642–656. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Sharon N, Vanderhooft J, Straubhaar J,
Mueller J, Chawla R, Zhou Q, Engquist EN, Trapnell C, Gifford DK
and Melton DA: Wnt signaling separates the progenitor and endocrine
compartments during pancreas development. Cell Rep.
27:2281–2291.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ma L, Wang Y, Hui Y, Du Y, Chen Z, Feng H,
Zhang S, Li N, Song J, Fang Y, et al: WNT/NOTCH pathway is
essential for the maintenance and expansion of human MGE
progenitors. Stem Cell Reports. 12:934–949. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Huynh DL, Koh H, Chandimali N, Zhang JJ,
Kim N, Kang TY, Ghosh M, Gera M, Park YH, Kwon T and Jeong DK:
BRM270 inhibits the proliferation of CD44 positive pancreatic
ductal adenocarcinoma cells via downregulation of sonic hedgehog
signaling. Evid Based Complement Alternat Med. 2019:86204692019.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Che SM, Zhang XZ, Liu XL, Chen X and Hou
L: The radiosensitization effect of NS398 on esophageal cancer stem
cell-like radioresistant cells. Dis Esophagus. 24:265–273. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yue D, Zhang Z, Li J, Chen X, Ping Y, Liu
S, Shi X, Li L, Wang L, Huang L, et al: Transforming growth
factor-beta1 promotes the migration and invasion of sphere-forming
stem-like cell subpopulations in esophageal cancer. Exp Cell Res.
336:141–149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Ding W, Mouzaki M, You H, Laird JC, Mato
J, Lu SC and Rountree CB: CD133+ liver cancer stem cells from
methionine adenosyl transferase 1A-deficient mice demonstrate
resistance to transforming growth factor (TGF)-beta-induced
apoptosis. Hepatology. 49:1277–1286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Mima K, Okabe H, Ishimoto T, Hayashi H,
Nakagawa S, Kuroki H, Watanabe M, Beppu T, Tamada M, Nagano O, et
al: CD44s regulates the TGF-β-mediated mesenchymal phenotype and is
associated with poor prognosis in patients with hepatocellular
carcinoma. Cancer Res. 72:3414–3423. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Mitra M, Kandalam M, Harilal A, Verma RS,
Krishnan UM, Swaminathan S and Krishnakumar S: EpCAM is a putative
stem marker in retinoblastoma and an effective target for
T-cell-mediated immunotherapy. Mol Vis. 18:290–308. 2012.PubMed/NCBI
|
|
94
|
Zhang M, Tan S, Yu D, Zhao Z, Zhang B,
Zhang P, Lv C, Zhou Q and Cao Z: Triptonide inhibits lung cancer
cell tumorigenicity by selectively attenuating the Shh-Gli1
signaling pathway. Toxicol Appl Pharmacol. 365:1–8. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Arai MA, Ochi F, Makita Y, Chiba T,
Higashi K, Suganami A, Tamura Y, Toida T, Iwama A, Sadhu SK, et al:
GLI1 inhibitors identified by target protein oriented natural
products isolation (TPO-NAPI) with hedgehog inhibition. ACS Chem
Biol. 13:2551–2559. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yang Z, Cui Y, Ni W, Kim S and Xuan Y:
Gli1, a potential regulator of esophageal cancer stem cell, is
identified as an independent adverse prognostic factor in
esophageal squamous cell carcinoma. J Cancer Res Clin Oncol.
143:243–254. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Fujiwara D, Kato K, Nohara S, Iwanuma Y
and Kajiyama Y: The usefulness of three-dimensional cell culture in
induction of cancer stem cells from esophageal squamous cell
carcinoma cell lines. Biochem Biophys Res Commun. 434:773–778.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Kanamoto A, Ninomiya I, Harada S, Tsukada
T, Okamoto K, Nakanuma S, Sakai S, Makino I, Kinoshita J, Hayashi
H, et al: Valproic acid inhibits irradiation-induced
epithelial-mesenchymal transition and stem cell-like
characteristics in esophageal squamous cell carcinoma. Int J Oncol.
49:1859–1869. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zhang JX, Chen ZH, Xu Y, Chen JW, Weng HW,
Yun M, Zheng ZS, Chen C, Wu BL, Li EM, et al: Downregulation of
MicroRNA-644a promotes esophageal squamous cell carcinoma
aggressiveness and stem-cell-like phenotype via dysregulation of
PITX2. Clin Cancer Res. 23:298–310. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
De Luca M, Aiuti A, Cossu G, Parmar M,
Pellegrini G and Robey PG: Advances in stem cell research and
therapeutic development. Nat Cell Biol. 21:801–811. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
de Sousa EM, Vermeulen L, Richel D and
Medema JP: Targeting Wnt signaling in colon cancer stem cells. Clin
Cancer Res. 17:647–653. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Merchant AA and William M: Targeting
Hedgehog-a cancer stem cell pathway. Clin Cancer Res. 16:3130–3140.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Galoczova M, Coates P and Vojtesek B:
STAT3, stem cells, cancer stem cells and p63. Cell Mol Biol Lett.
23:122018. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Fu J and Wang H: Precision diagnosis and
treatment of liver cancer in China. Cancer Lett. 412:283–288. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Irwin CR, Hitt MM and Evans DH: Targeting
nucleotide biosynthesis: A strategy for improving the oncolytic
potential of DNA viruses. Front Oncol. 7:2292017. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Pandey S and Robertson ES: Oncogenic
Epstein-Barr virus recruits Nm23-H1 to regulate chromatin
modifiers. Lab Invest. 98:258–268. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Lin TA, Garden AS, Elhalawani H, Elgohari
B, Jethanandani A, Ng SP, Mohamed AS, Frank SJ, Glisson BS, Debnam
JM, et al: Radiographic retropharyngeal lymph node involvement in
human papillomavirus-associated oropharyngeal carcinoma: Patterns
of involvement and impact on patient outcomes. Cancer.
125:1536–1546. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Wang D, Plukker JTM and Coppes RP: Cancer
stem cells with increased metastatic potential as a therapeutic
target for esophageal cancer. Semin Cancer Biol. 44:60–66. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Hirai M, Kelsey LS, Vaillancourt M,
Maneval DC, Watanabe T and Talmadge JE: Purging of human breast
cancer cells from stem cell products with an adenovirus containing
p53. Cancer Gene Ther. 7:197–206. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Eriksson M, Guse K, Bauerschmitz G,
Virkkunen P, Tarkkanen M, Tanner M, Hakkarainen T, Kanerva A,
Desmond RA, Pesonen S and Hemminki A: Oncolytic adenoviruses kill
breast cancer initiating CD44+CD24-/low cells. Mol Ther.
15:2088–2093. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Cho RW, Wang X, Diehn M, Shedden K, Chen
GY, Sherlock G, Gurney A, Lewicki J and Clarke MF: Isolation and
molecular characterization of cancer stem cells in MMTV-Wnt-1
murine breast tumors. Stem Cells. 26:364–371. 2010. View Article : Google Scholar
|
|
113
|
Mui UN, Haley CT and Tyring SK: Viral
oncology: Molecular biology and pathogenesis. J Clin Med. 6(pii):
1112017. View Article : Google Scholar
|
|
114
|
Ali SM, Ross JS and Wang K: Reply to
Genomic profiles of nasopharyngeal carcinoma: The importance of
histological subtyping and Epstein-Barr virus in situ
assays. Cancer. 124:435–436. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Satoru K, Naohiro W, Masamichi M, Zen Y,
Endo K, Murono S, Sugimoto H, Yamaoka S, Pagano JS and Yoshizaki T:
Epstein-Barr virus latent membrane protein 1 induces cancer
stem/progenitor-like cells in nasopharyngeal epithelial cell lines.
J Virol. 85:11255–11264. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Chris C, Figueroa JA, Leonardo M, Colombo
M, Summers G, Figueroa A, Aulakh A, Konala V, Verma R, Riaz J, et
al: The role of human papilloma virus (HPV) infection in
non-anogenital cancer and the promise of immunotherapy: A review.
Int Rev Immunol. 33:383–401. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Swanson MS, Kokot N and Sinha UK: The role
of HPV in head and neck cancer stem cell formation and
tumorigenesis. Cancers (Basel). 8(pii): E242016. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Ortiz-Sánchez E, Santiago-López L,
Cruz-Domínguez VB, Toledo-Guzmán ME, Hernández-Cueto D,
Muñiz-Hernández S, Garrido E, Cantú De León D and García-Carrancá
A: Characterization of cervical cancer stem cell-like cells:
Phenotyping, stemness, and human papilloma virus co-receptor
expression. Oncotarget. 7:31943–31954. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Lanfredini S, Olivero C, Borgogna C,
Calati F, Powell K, Davies KJ, De Andrea M, Harries S, Tang HKC,
Pfister H, et al: HPV8 field cancerization in a transgenic mouse
model is due to Lrig1+ keratinocyte stem cell expansion. J Invest
Dermatol. 137:2208–2216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Zhang M, Kumar B, Piao L, Xie X, Schmitt
A, Arradaza N, Cippola M, Old M, Agrawal A, Ozer E, et al: Elevated
intrinsic cancer stem cell population in human
papillomavirus-associated head and neck squamous cell carcinoma.
Cancer. 120:992–1001. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Zhang M, Zhuang G, Sun X, Shen Y, Wang W,
Li Q and Di W: TP53 mutation-mediated genomic instability induces
the evolution of chemoresistance and recurrence in epithelial
ovarian cancer. Diagn Pathol. 12:162017. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Chiche A, Moumen M, Romagnoli M, Petit V,
Lasla H, Jézéquel P, de la Grange P, Jonkers J, Deugnier MA,
Glukhova MA and Faraldo MM: p53 deficiency induces cancer stem cell
pool expansion in a mouse model of triple-negative breast tumors.
Oncogene. 36:2355–2365. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Shetzer Y, Molchadsky A and Rotter V:
Oncogenic mutant p53 gain of function nourishes the vicious cycle
of tumor development and cancer stem-cell formation. Cold Spring
Harb Perspect Med. 6(pii): a0262032016. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Tan MJ, White EA, Sowa ME, Harper JW,
Aster JC and Howley PM: Cutaneous β-human papillomavirus E6
proteins bind Mastermind-like coactivators and repress Notch
signaling. Proc Natl Acad Sci USA. 109:E1473–E1480. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Shamir ER, Devine WP, Pekmezci M, Umetsu
SE, Krings G, Federman S, Cho SJ, Saunders TA, Jen KY, Bergsland E,
et al: Identification of high-risk human papillomavirus and Rb/E2F
pathway genomic alterations in mutually exclusive subsets of
colorectal neuroendocrine carcinoma. Mod Pathol. 32:290–305. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Dyson N, Howley PM, Münger K and Harlow E:
The human papilloma virus-16 E7 oncoprotein is able to bind to the
retinoblastoma gene product. Science. 243:934–937. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Shanmugarajah R, Bin W, Snow ET, Sharma P,
Pavey D, Merrett N, Ball MJ, Brain T, Fernando R and Robertson IK:
Transcriptionally active human papillomavirus is strongly
associated with Barrett's dysplasia and esophageal adenocarcinoma.
Am J Gastroenterol. 108:1082–1093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Anders M, Rösch T, Küster K, Becker I,
Höfler H, Stein HJ, Meining A, Wiedenmann B and Sarbia M:
Expression and function of the coxsackie and adenovirus receptor in
Barrett's esophagus and associated neoplasia. Cancer Gene Ther.
16:508–515. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Chang F, Syrjänen S, Wang L and Syrjänen
K: Infectious agents in the etiology of esophageal cancer.
Gastroenterology. 103:1336–1348. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Chang F, Syrjänen S, Shen Q, Ji HX and
Syrjänen K: Human papillomavirus (HPV) DNA in esophageal precancer
lesions and squamous cell carcinomas from China. Int J Cancer.
45:21–25. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
He Z, Xu Z, Hang D, Guo F, Abliz A, Weiss
NS, Xi L, Liu F, Ning T, Pan Y, et al: Anti-HPV-E7 seropositivity
and risk of esophageal squamous cell carcinoma in a high-risk
population in China. Carcinogenesis. 35:816–821. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Wang L, Li J, Hou J, Li M, Cui X, Li S, Yu
X, Zhang Z, Liang W, Jiang J, et al: p53 expression but not
p16(INK4A) correlates with human papillomavirus-associated
esophageal squamous cell carcinoma in Kazakh population. Infect
Agent Cancer. 11:192016. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Ludmir EB, Stephens SJ, Palta M, Willett
CG and Czito BG: Human papillomavirus tumor infection in esophageal
squamous cell carcinoma. J Gastrointest Oncol. 6:287–295.
2015.PubMed/NCBI
|
|
134
|
Xi R, Pan S, Chen X, Hui B, Zhang L, Fu S,
Li X, Zhang X, Gong T, Guo J, et al: HPV16 E6-E7 induces cancer
stem-like cells phenotypes in esophageal squamous cell carcinoma
through the activation of PI3K/Akt signaling pathway in
vitro and in vivo. Oncotarget. 7:57050–57065. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Syrjänen KJ: HPV infections and
oesophageal cancer. J Clin Pathol. 55:721–728. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Halec G, Schmitt M, Egger S, Abnet CC,
Babb C, Dawsey SM, Flechtenmacher C, Gheit T, Hale M, Holzinger D,
et al: Mucosal alpha-papillomaviruses are not associated with
esophageal squamous cell carcinomas: Lack of mechanistic evidence
from South Africa, China and Iran and from a world-wide
meta-analysis. Int J Cancer. 139:85–98. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Yang L, Ji Y, Chen L, Li M, Wu F, Hu J,
Jiang J, Cui X, Chen Y, Pang L, et al: Genetic variability in LMP2
and LMP7 is associated with the risk of esophageal squamous cell
carcinoma in the Kazakh population but is not associated with HPV
infection. PLoS One. 12:e01863192017. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
da Costa AM, Fregnani JHTG, Pastrez PRA,
Mariano VS, Silva EM, Neto CS, Guimarães DP, Villa LL, Sichero L,
Syrjanen KJ and Longatto-Filho A: HPV infection and p53 and p16
expression in esophageal cancer: are they prognostic factors?
Infect Agent Cancer. 12:542017. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Kayamba V, Bateman AC, Asombang AW,
Shibemba A, Zyambo K, Banda T, Soko R and Kelly P: S HIV infection
and domestic smoke exposure, but not human papilloma virus, are
risk factors for oesophageal squamous cell carcinoma in Zambia: A
case-control study. Cancer Med. 4:588–595. 2015. View Article : Google Scholar : PubMed/NCBI
|