Introduction
Lung adenocarcinoma is one of the most common
malignant tumors in China (1). It
has a high incidence rate, progresses rapidly and poses a threat to
human health (2). Lung
adenocarcinoma is prone to metastasis at an early stage and is
likely to develop resistance to standard treatments such as
radiotherapy and chemotherapy, resulting in poor clinical efficacy
and poor prognosis (3). A number of
studies (4–6) have been conducted to explore the
molecular mechanism underlying proliferation and drug sensitivity
of lung adenocarcinoma cells, in order to enhance the understanding
of the disease pathogenesis and for the identification of potential
therapeutic targets.
Among these studies, ERK has been frequently studied
as a molecular target (7,8). ERK has several regulatory roles in
various cellular functions, including cell proliferation and
differentiation (9). Moreover, ERK
is a member of the mitogen-activated protein kinase (MAPK)
signaling cascade, which includes three classes of protein kinases:
MAPK kinase kinase, MAPK kinase, MAPK/ERK kinase (MEK) and MAPK
ERK. The ERK signaling pathway is activated by a combination of
growth factors, extracellular matrix, receptor tyrosine kinases and
integrins (10). Phosphorylated
(p)-ERK, the active form of ERK, is an important indicator of ERK
signaling activity (11–13). p-ERK translocates to the nucleus and
regulates gene expression by directly phosphorylating various
nuclear proteins, including transcription factors, and thereby
regulates cell proliferation (14,15).
However, the upstream regulatory mechanism of the ERK signaling
pathway in tumor progression requires further investigation.
Placenta-specific protein 8 (PLAC8) is a protein
with a molecular weight of 12.5 kDa, and its molecular structure is
highly conserved from amphibians to humans (16). PLAC8 was originally discovered in
mice and was considered to be a placental-specific transcriptional
gene (17). It was eventually found
to play an important role in malignant tumor progression, including
in leukemia, pancreatic cancer, colon cancer and osteosarcoma
(18–21), by regulating various cellular
functions, including apoptosis, differentiation and autophagy
(20–22). Recently, a study by Jin et al
(23) found that PLAC8 expression
levels were significantly higher in lung adenocarcinoma cells that
were tolerant to radiotherapy compared with that in lung
adenocarcinoma cells that were sensitive to radiotherapy. Moreover,
it was demonstrated that overexpression of PLAC8 enhanced the
stemness and tolerance to radiotherapy of lung adenocarcinoma
cells, indicating a role for PLAC8 in the regulation of sensitivity
to radiotherapy (23). A study by
Zhang et al (24) revealed
that expression of PLAC8 was high in osteosarcoma cells with a p53
gene mutation, and this was found to be associated with promotion
of osteosarcoma metastasis through the MAPK signaling pathway.
Considering the diversity of PLAC8 function and its regulatory role
in lung adenocarcinoma, it was hypothesized that PLAC8 may be
involved in lung adenocarcinoma cell proliferation and the
regulation of drug sensitivity to gefitinib, an epidermal growth
factor receptor tyrosine kinase inhibitor (EGFR-TKI), via the ERK
signaling pathway. This study aimed to determine whether PLAC8 may
enhance the effect of gefitinib and represent a potential novel
target in lung adenocarcinoma.
Materials and methods
Cell culture
PC-9 cells were purchased from the American Type
Culture Collection. The cells were maintained in RPMI 1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) containing 10% fetal calf
serum (Gibco; Thermo Fisher Scientific, Inc.) and cultured at 37°C
in 5% carbon dioxide.
Lentiviral transfections. In the present study, two
short hairpin (sh) RNA interference sequences for PLAC8, sh1# and
sh2#, were designed as follows: Sh1,
5′-CCGGCAATGAGGACTCTCTACAGGACTCGAGTCCTGTAGAGAGTCCTCATTGTTTTTTG-3′;
and Sh2,
5′-CCGGCTTTGCCAAATCAAGAGATGTCTGGAGATCTCTCTTGATTTGGCAAAGTTTTTTG-3′
(RuboBio Biotechnology Co., Ltd.). PC-9 cells (50% confluence) were
transfected with 2 µg plasmid for 72 h. The DNA sequence of human
PLAC8 (NM_001130715.1) was obtained from human monocytes by PCR and
inserted into the vector using Cloning Kit (Promega Corporation)
(25). The primers for PLAC8 used
for the PCR were as follows: PLAC8 forward,
5′-GAACTCAGATCTCGAAAAATGCAAGCTCAGGCGC-3′, and reverse
5′-CATGACCGGTGGATCGAAAGTACGCATGGCTCTC-3′. The pLKO.1 puro
lentiviral vector (Chengdu Biomart Biotechnology Co., Ltd.) was
used as the interference vector and a scrambled shRNA sequence
5′-CCGGGAGCAATCGCACGAGCTAATTCTCGAGAATTAGCTCGTGCGATTGCTCTTTTTTG-3′
was used as a knockdown negative control (KDNC). The pLVX–IRES-puro
lentiviral vector (Chengdu Biomart Biotechnology Co., Ltd.) was
used as the overexpression vector and an empty vector was used as
an overexpression negative control (OENC). After 72 h of lentivirus
transfection, positive cells were selected with puromycin. To do
so, cells were incubated with 2 µg/ml puromycin for 5 days at 37°C.
Cells that were resistant to puromycin represented cells that were
efficiently transfected.
Western blot analysis
Protein extraction and western blot analyses were
performed as previously described (26). Briefly, IP lysis buffer (Beyotime
Institute of Biotechnology) and protease inhibitor (complete ULTRA
tablets; Roche Diagnostics GmbH) were used for protein extraction
at 4°C for 30 min. Cell lysate was then centrifuged at 12,000 × g
for 5 minutes and the supernatant was discarded. A BCA protein
assay kit (Beyotime Institute of Biotechnology) was used to detect
protein concentrations. Proteins (30 µg) were separated by 10%
SDS-PAGE gels and transferred onto polyvinylidene difluoride
membranes. Membranes were blocked with 5% skimmed milk for 1 h at
room temperature, and incubated with primary antibodies against
PLAC8 (1:1,000; cat. no. 13885), ERK1/2 (1:2,000; cat. no. 4695),
p-ERK1/2 (1:2,000; cat. no. 9101), EGFR (1:3,000; cat. no. 4267),
p-EGFR (1:800; cat. no. 3777), cleaved caspase 3 (1:1,500; cat. no.
9661), cleaved poly (ADP-ribose) polymerase 1 (PARP) (1:3,000; cat.
no. 9532) and GAPDH (1:5,000; cat. no. 5174) overnight at 4°C. All
primary antibodies were purchased from Cell Signaling Technology.
Membranes were then incubated with horseradish
peroxidase-conjugated goat anti-rabbit and anti-mouse secondary
antibodies (cat. nos. ZB-2301 and ZB-2305, respectively; 1:10,000;
OriGene Technologies, Inc.). Bands were detected using enhanced
chemiluminescence substrate (Bio-Rad Laboratories, Inc.)
Detection of cell viability and
proliferation by Cell Counting Kit-8 (CCK-8) assay
Cell viability was measured using the CCK-8 assay
kit (Beyotime Institute of Biotechnology). Following 72 h of cell
transfection with lentivirus, cells were seeded in 96-well plates
at 5,000 cells/well. At the time of detection, 10 µl CCK-8 solution
was added to each well, and the absorbance was measured at 450 nm
using a microplate reader (Thermo Fisher Scientific, Inc.) after 90
min. To measure the relative proliferative rate of cells, PC-9 OENC
and PC-9 PLAC8 cells were seeded in 96-well plates, and each group
was administered DMSO or 0.001, 0.01, 0.05, 0.1, 0.5, 1 and 5 µM
gefitinib (Selleck Chemicals; cat. no. S1025). For U0216
(MedChemExpress; cat. no. HY-12031) administration, only PC-9 OENC
and PC-9 PLAC8 groups treated with DMSO were added as controls.
After 72 h, medium was changed and cells were incubated with 100 µl
medium containing 10% CCK-8 for 90 min. Absorbance was then
measured at 450 nm using a microplate reader, and the relative
proliferative rate of the cells was calculated as the ratio of the
optical density (OD) value of the treatment group at 72 h to the OD
value of the control group at 72 h. Eight replicate wells were
tested for each set of samples, and the average absorbance was
calculated. GraphPad Prism software v7.0 (GraphPad Software, Inc.)
was used to plot the proliferation curves and to calculate the
IC50 values.
Statistical analysis
Each experiment was repeated 3 times and
representative results were selected. Data analysis was performed
using SPSS version 18.0 statistical software (IBM Corp.). The data
are expressed as the mean ± SD. The CCK-8 assay was analyzed using
one-way ANOVA, followed by the least significant difference test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Stable infection of the PC-9 cell line
with PLAC8-silencing and PLAC8-overexpressing lentiviruses
Lung adenocarcinoma PC-9 cells were infected with
lentiviruses carrying PLAC8-targeting shRNAs (sh1#, sh2#), a
knockdown control (KDNC), PLAC8 or an empty vector control (OENC).
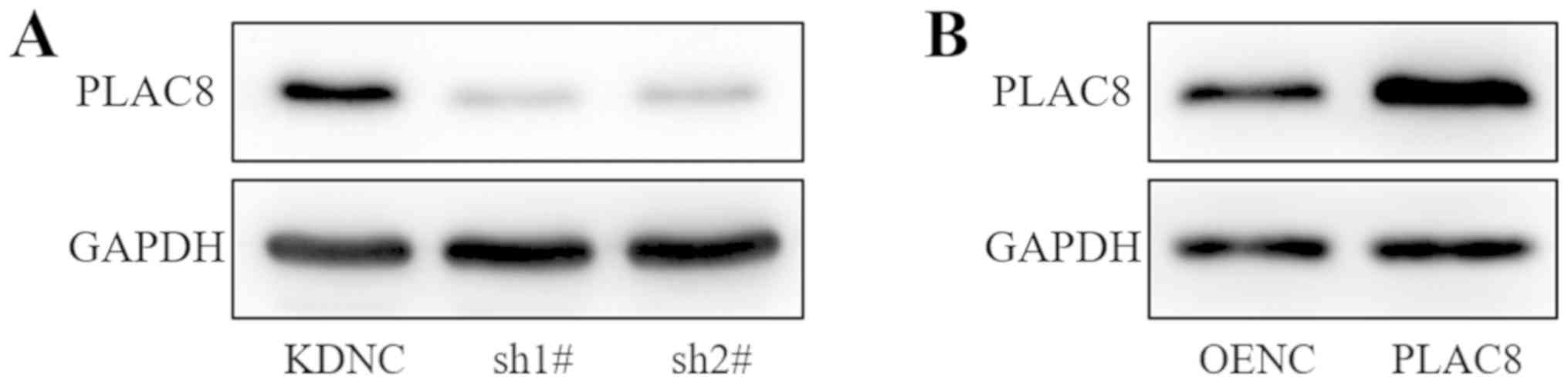
Western blot analysis demonstrated reduced protein expression of
PLAC8 in the sh1# and sh2# groups compared with that in the KDNC
group (Fig. 1A). PLAC8 protein
expression was increased in the PLAC8 group compared with that in
the OENC group (Fig. 1B). Thus, the
western blot analysis confirmed the successful transfection of
PLAC8 shRNAs and overexpression vector.
PLAC8 enhances PC-9 cell
proliferation
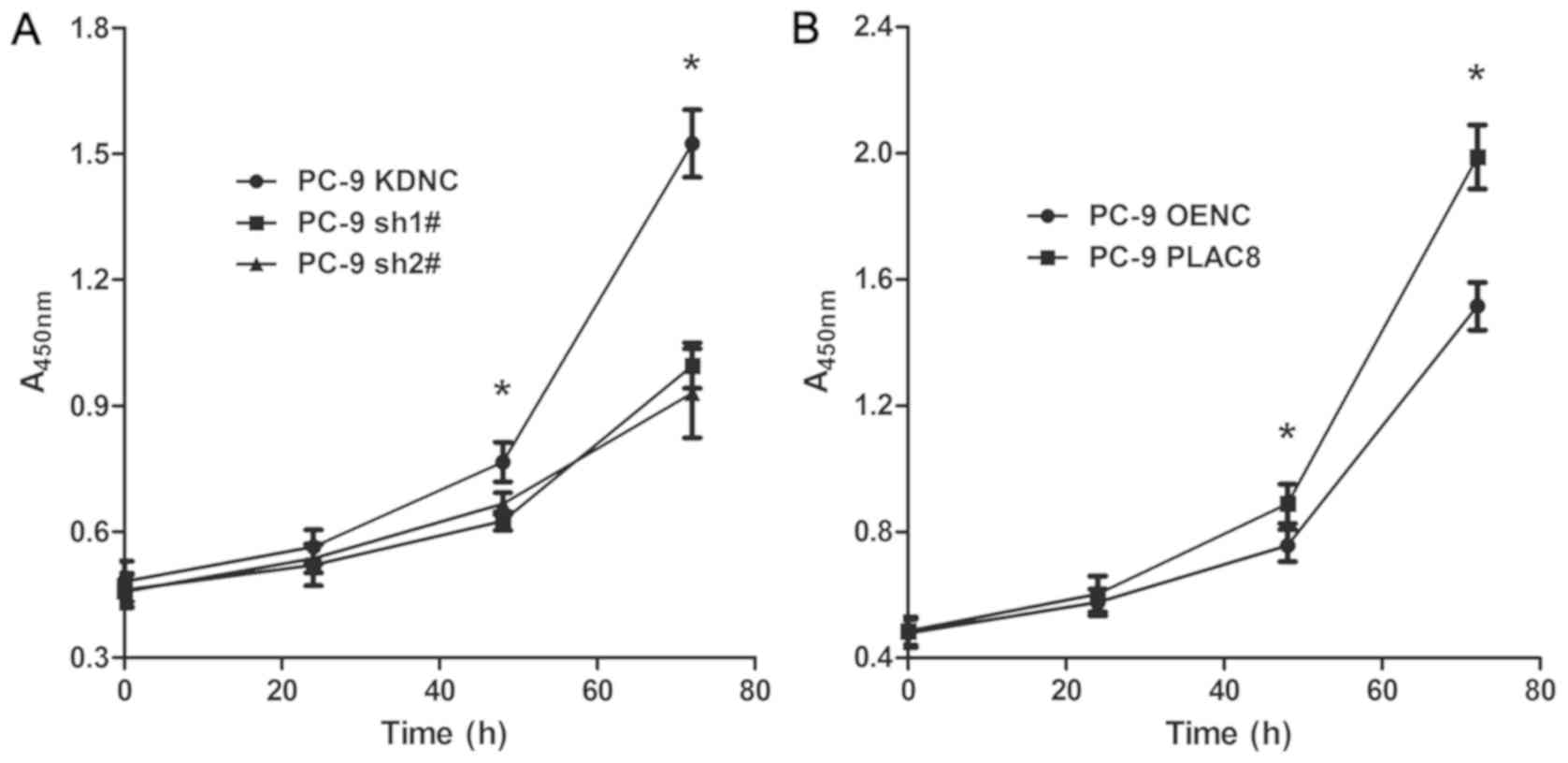
To study the effect of PLAC8 on the proliferation of
lung adenocarcinoma PC-9 cells, the CCK-8 assay was used.
Proliferation was detected at 0, 24, 48 and 72 h (Fig. 2). As presented in Fig. 2A, the proliferative capacity of PC-9
cells was significantly decreased following PLAC8 knockdown. At 24
h, there was no significant difference between the three groups
(P>0.05). At 48 h, the proliferation was significantly reduced
in sh1# and sh2# groups compared with that in the KDNC group
(P<0.05). At 72 h, the difference between the KDNC group and the
sh1# and sh2# groups was greater (P<0.05). As presented in
Fig. 2B, the proliferation of PC-9
cell was significantly increased following PLAC8 overexpression.
There was no significant difference between the PLAC8 and OENC
groups at 0 and 24 h. However, by 48 h, the proliferation was
higher in the PLAC8 overexpression group compared with the OENC
group and at 72 h, the difference between the two groups was
further increased (P<0.05). These results indicated that PLAC8
influenced PC-9 cell proliferation, as it was inhibited following
knockdown and enhanced following overexpression of PLAC8.
PLAC8 enhances ERK signaling pathway
activity
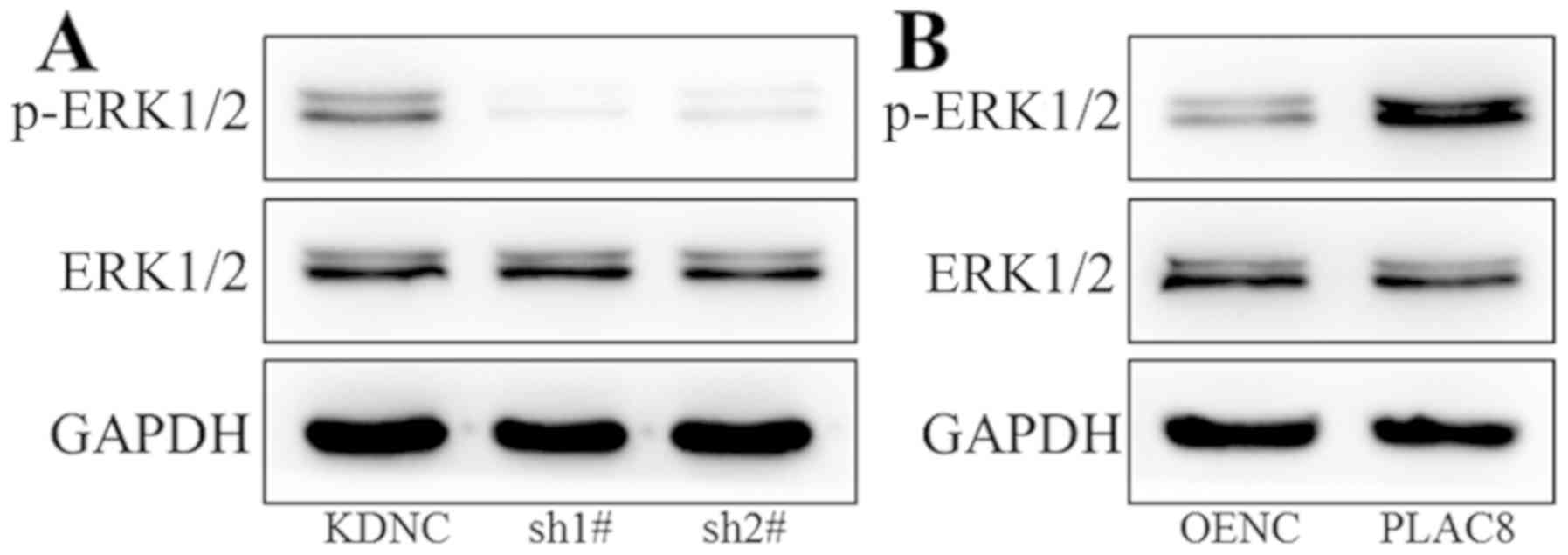
Western blot analysis was performed to study the
mechanism underlying the impact of PLAC8 on proliferation of PC-9
cells. The ERK signaling pathway was investigated as it plays an
important regulatory role in tumor cell viability (27,28). As
presented in Fig. 3A, there was no
change in ERK1/2 total protein expression after knockdown of PLAC8,
however p-ERK1/2 expression was decreased, indicating decreased ERK
signaling activity. As shown in Fig.
3B, protein expression of ERK1/2 did not change after PLAC8
overexpression, however p-ERK1/2 protein levels were increased in
cells with overexpression of PLAC8. These results indicate a
regulatory role for PLAC8 in the activity of the ERK signaling
pathway.
PLAC8 promotes PC-9 cell proliferation
through the ERK signaling pathway
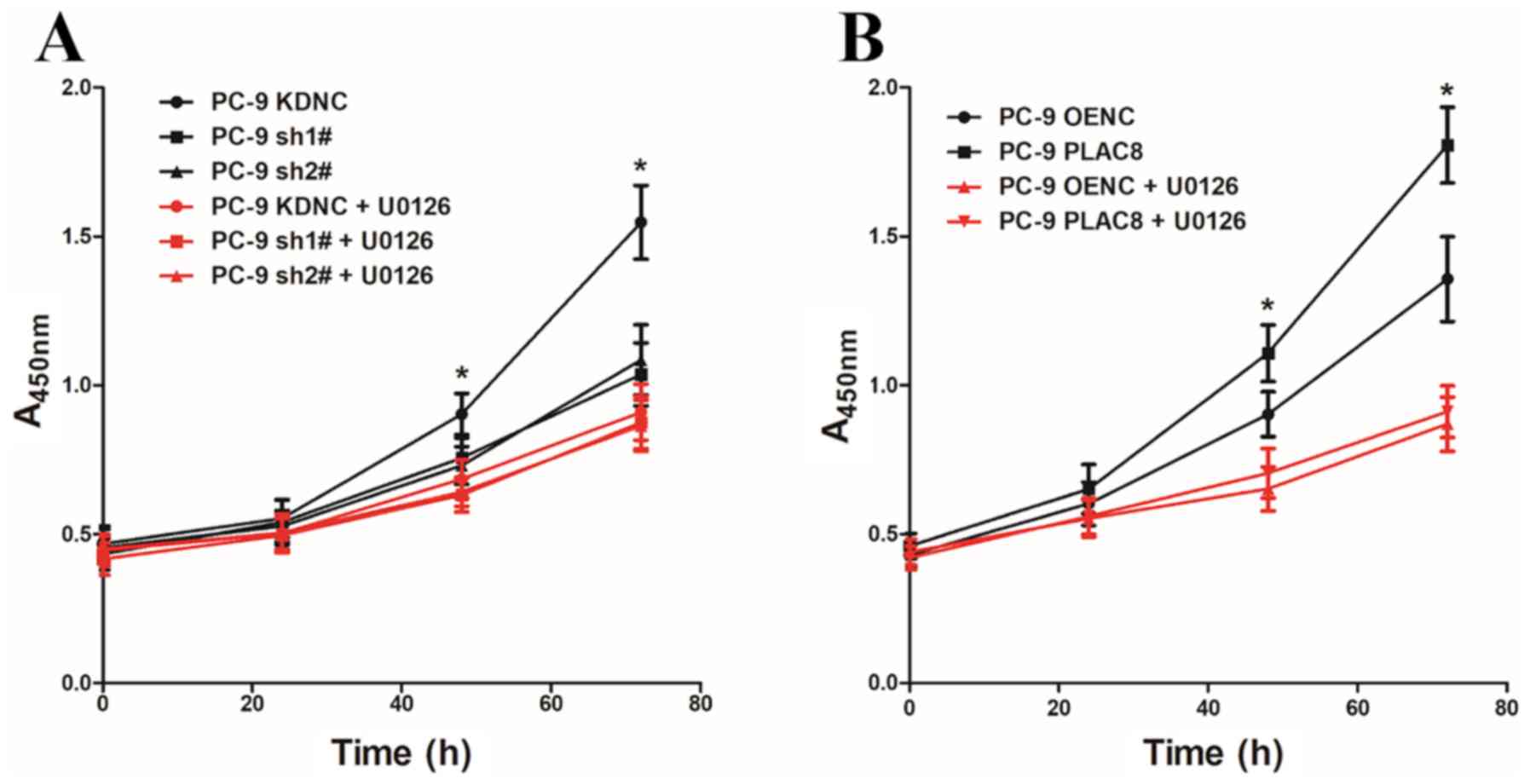
To investigate whether PLAC8 regulates the
proliferation of PC-9 cells via the ERK signaling pathway, PC-9
cells with PLAC8 overexpression and knockdown were treated with the
ERK signaling pathway inhibitor U0126 (5 µM). The CCK-8 assay was
used to detect cell proliferation with and without the addition of
the ERK signaling pathway inhibitor U0126 at 0, 24, 48 and 72 h
(Fig. 4). As presented in Fig. 4A, when U0126 was not added, cell
proliferation was significantly lower in cells transfected with two
PLAC8-targeting shRNAs compared to control KDNC cells. However,
there were no significant differences in the proliferation rate
between KDNC cells and those with PLAC8 knocked down that had been
treated with the ERK signaling pathway inhibitor U0126. Similarly,
the proliferation of PC-9 PLAC8 cells was significantly higher
compared with that of the PC-9 OENC group in untreated cells, but
there was no significant difference in the proliferation ability of
the two groups after the addition of U0126 (Fig. 4B). These results demonstrated that
altering the expression of PLAC8 while inhibiting the ERK signaling
activity did not affect the proliferation of PC-9 cells.
PLAC8 decreases the sensitivity of
PC-9 cells to gefitinib
PC-9 cells carry EGFR-sensitive mutations and can be
treated with gefitinib, an EGFR-TKI. OENC and PLAC8-overexpressing
cells were treated with 0.001, 0.01, 0.05, 0.1, 0.5, 1 and 5 µM
gefitinib. Cells treated with DMSO alone were used as controls.
Following 72 h, the effect of gefitinib was detected by CCK-8 assay
and the cell viability of OENC and PLAC8-overexpressing cells was
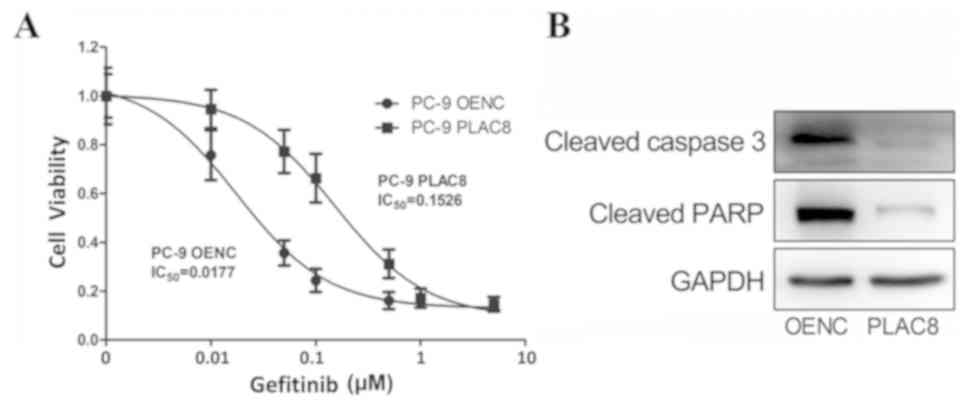
determined. Fig. 5A shows that the
PC-9 cells with overexpression of PLAC8 were more tolerant to
gefitinib than PC-9 OENC cells; the PLAC8 group had an
IC50 value of 0.1526 µM, which was higher than that of
the PC-9 OENC group (0.0177 µM). Subsequently, OENC and PLAC8
groups were treated with 0.5 µM gefitinib for 24 h to detect the
expression of apoptosis-associated proteins. As shown in Fig. 5B, apoptosis-associated protein levels
were lower in PC-9 PLAC8 cells compared with those in PLAC8 OENC
cells, further indicating that the sensitivity of PC-9 cells to
gefitinib was decreased following PLAC8 overexpression.
PLAC8 decreases the sensitivity of
PC-9 cells to gefitinib by activating the ERK pathway
The role of ERK in the PLAC8-mediated decrease in
sensitivity to gefitinib was investigated. OENC and
PLAC8-overexpressing cells were treated with 0.05 µM gefitinib
alone or in combination with 5 µM U0126. Cells treated with DMSO
were considered as the control groups. The expression of ERK
proteins and the relative cell viability was detected by western
blot and CCK-8 assays, respectively.
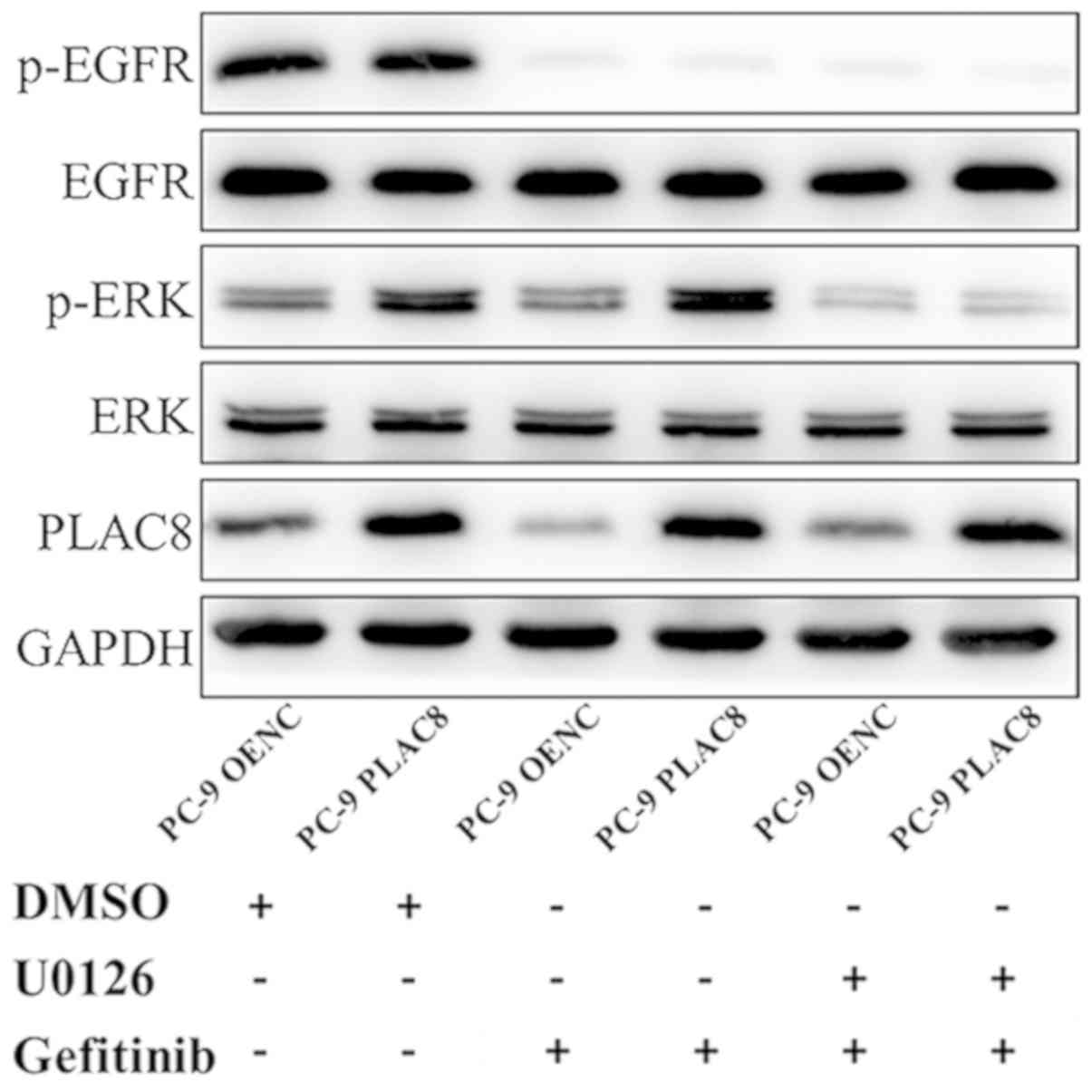
As presented in Fig.
6, western blot analysis revealed a decrease in the protein
expression of p-EGFR in PC-9 OENC and PC-9 PLAC8 cells following 72
h of gefitinib treatment, but EGFR expression level was not
changed. The anticancer effect exerted by gefitinib is mediated by
the inhibition of EGFR phosphorylation (29,30).
Following PLAC8 overexpression, ERK was phosphorylated in PC-9
cells, but ERK protein expression was not significantly changed.
Following treatment with U0126 and gefitinib, ERK phosphorylation
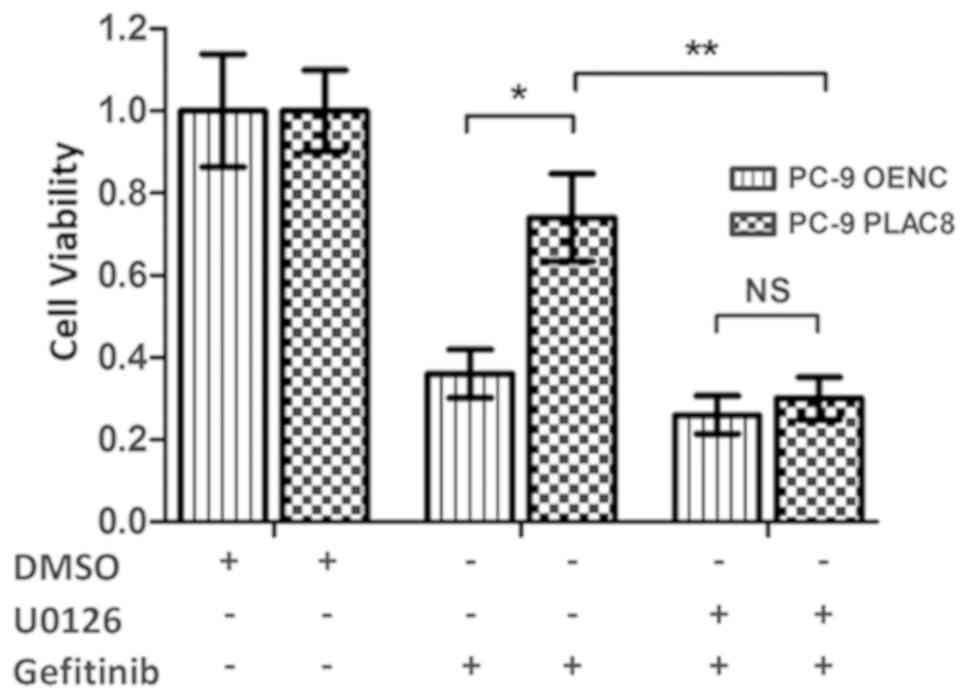
was inhibited in PLAC8 overexpressing cells. The CCK-8 results
presented in Fig. 7 revealed that
the relative survival rates of OENC cells and PLAC8-overexpressing
cells were significantly different following 72 h of gefitinib
treatment. The survival rate of PLAC8-overexpressiong cells was
significantly higher compared with that of OENC cells (P<0.05).
By contrast, when U0126 treatment was used to inhibit the ERK
signaling pathway, the relative cell survival rate was not
significantly different between OENC and PLAC8 cells following 72 h
of gefitinib treatment, indicating that PLAC8 could not induce
resistance in PC-9 cells to gefitinib following inhibition of the
ERK signaling pathway. These results suggested that the ERK
signaling pathway was involved in the regulation of PC-9 cell
sensitivity to gefitinib by PLAC8.
PLAC8 decreases the expression of
apoptosis-associated proteins via the ERK pathway in PC-9 cells
treated with gefitinib
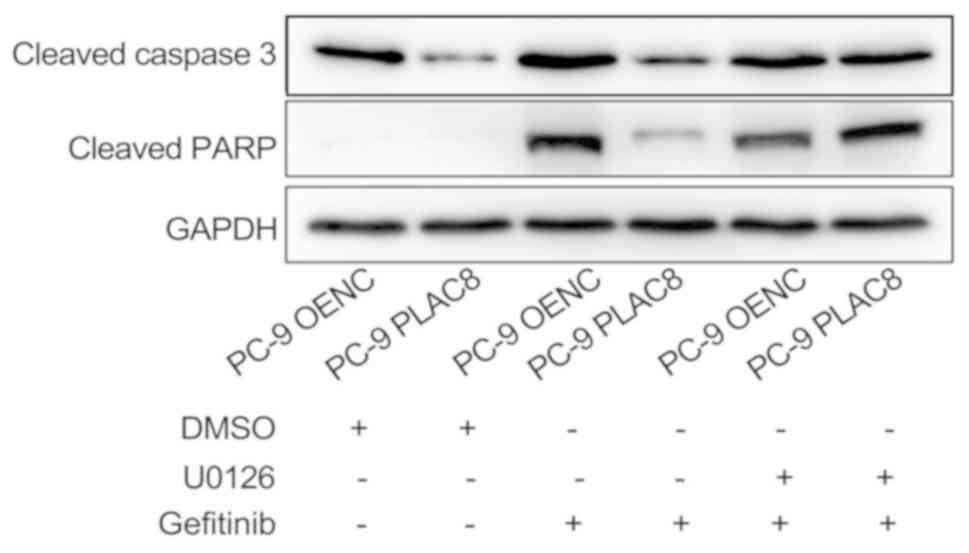
OENC and PLAC8-overexpressing cells were treated
with 0.5 µM gefitinib alone or in combination with 5 µM U0126 and
the expression levels of apoptosis-associated proteins, cleaved
caspase 3 and cleaved PARP, were detected by western blot analysis
after 24 h. As presented in Fig. 8,
the expression of cleaved caspase 3 and cleaved PARP was reduced in
PLAC8-overexpressing cells compared with that in OENC cells
following 24 h of gefitinib treatment. This suggested that PLAC8
decreased the sensitivity of PC-9 cells to gefitinib-induced
apoptosis. However, inhibition of the ERK signaling pathway by
treatment with U0126 reversed the effect of PLAC8 overexpression on
the expression of cleaved caspase 3 and cleaved PARP in cells
treated with gefitinib. This suggested that PLAC8-mediated
resistance to gefitinib-induced apoptosis may be dependent on the
ERK signaling pathway.
Discussion
In recent years, the role of PLAC8 in tumor
progression has begun to be uncovered (18,31).
However, to the best of our knowledge, the regulatory role and
mechanism of PLAC8 in lung adenocarcinoma have not yet been
reported. Recently, Jin et al (23) reported that PLAC8 is involved in the
resistance of lung adenocarcinoma to radiotherapy. No studies have
been performed on the regulation of lung adenocarcinoma cell
proliferation by PLAC8.
Firstly, the present study demonstrated a role for
PLAC8 in regulating the proliferation of lung adenocarcinoma PC-9
cells. Cell proliferation was decreased following the knockdown of
PLAC8 and significantly increased with overexpression of PLAC8 in
PC-9 cells. These results indicate that PLAC8 can enhance the
proliferation ability of PC-9 cells. Subsequently, it was revealed
that PLAC8 plays a regulatory role in the ERK signaling pathway, as
the inhibition of PLAC8 resulted in decreased p-ERK expression.
Finally, it was found that PLAC8 was unable to regulate PC-9 cell
proliferation following the inhibition of the ERK signaling
pathway. This indicates that the regulation of PC-9 cell
proliferation by PLAC8 is mediated via the ERK signaling
pathway.
U0126 is an effective compound that inhibits ERK1/2
activation by specifically inhibiting the activity of the ERK1/2
upstream kinase MEK1/2 (32). In the
present study, cells were treated with U0126, which can induce
apoptosis rapidly via inhibiting the ERK signaling pathway
activity, and the impact on PC-9 cell proliferation was
investigated using the CCK-8 assay. This revealed that the
inhibition of ERK activity had no effect on cell proliferation. The
proliferation of cells at 72 h after U0126 administration was the
latest time point detected; therefore, a low concentration of U0126
(5 µM) was used to prevent the drug from directly causing apoptosis
(33). The results indicated that
inhibition of ERK signaling pathway activity resulted in the loss
of PLAC8-mediated regulation of PC-9 cell proliferation. These
findings demonstrate that PLAC8 regulates PC-9 cell proliferation
in a manner dependent on ERK signaling pathway activation. In
addition, these results confirm that the ERK signaling pathway
activation may be critical for promoting lung adenocarcinoma cell
proliferation, and this observation is consistent with previous
studies showing that ERK signaling pathway activation promotes
tumor proliferation (34–36).
Certain subtypes of lung cancer have EGFR mutations.
Approximately 10–50% of patients with non-small cell lung cancer
have EGFR-activating mutations (37,38).
These patients are sensitive to EGFR-TKIs such as gefitinib and
erlotinib, and benefit from better therapeutic effects. However,
such patients will eventually develop EGFR-TKI resistance (39). Although the number of studies on the
mechanism of EGFR-TKI resistance and resistance reversal has
increased, current, alternative treatments are limited, and there
is still no effective way to reverse EGFR-TKI resistance. The
present study found that PC-9 cells are sensitive to gefitinib. A
low concentration of gefitinib significantly inhibited cell
proliferation, and overexpression of PLAC8 significantly enhanced
the resistance of PC-9 cells to gefitinib, demonstrating that PLAC8
may play an important role in mediating resistance to EGFR-TKIs in
lung adenocarcinoma. Furthermore, it was found that PLAC8 activated
the ERK signaling pathway and that ERK signaling was required for
the PLAC8-mediated resistance of PC-9 cells to gefitinib. In
addition, overexpression of PLAC8 decreased expression of
gefitinib-induced apoptosis-associated proteins in PC-9 cells, and
inhibition of ERK signaling pathway blocked this effect. These
results indicated that PLAC8 induced resistance of PC-9 cells to
gefitinib via the ERK signaling pathway. The present study revealed
the mechanism underlying the resistance of lung adenocarcinoma cell
to EGFR-TKIs, thus providing a novel approach for reversing
EGFR-TKI resistance.
One of the limitations of the present study was that
the mechanistic studies were not comprehensive. The mechanism by
which PLAC8 activates the ERK signaling pathway remains to be
determined. Furthermore, in addition to its regulatory role in lung
adenocarcinoma cells, the role of PLAC8 in other diseases remains
to be further explored.
In conclusion, the present study demonstrated that
PLAC8 enhanced proliferation and resistance to EGFR-TKIs in lung
adenocarcinoma PC-9 cells by activating the ERK signaling pathway.
These findings demonstrate the functional diversity of the role of
PLAC8 in lung adenocarcinoma and provide a novel therapeutic target
for future studies.
Acknowledgements
Not applicable.
Funding
This study was supported by the Scientific Research
Topics of Sichuan Education Department (grant no. 18ZA0165).
Availability of data and material
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XZ and QL analyzed the data and wrote the paper. YY
and SL performed the CCK-8 and western blotting assays. DH, WJ and
RM designed the experiment and revised the paper. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cao M and Chen W: Epidemiology of lung
cancer in China. Thorac Cancer. 10:3–7. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Imielinski M, Berger AH, Hammerman PS,
Hernandez B, Pugh TJ, Hodis E, Cho J, Suh J, Capelletti M,
Sivachenko A, et al: Mapping the hallmarks of lung adenocarcinoma
with massively parallel sequencing. Cell. 150:1107–1120. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen J, Liu X, Xu Y, Zhang K, Huang J, Pan
B, Chen D, Cui S, Song H, Wang R, et al: TFAP2C-activated MALAT1
modulates the chemoresistance of docetaxel-resistant lung
adenocarcinoma cells. Mol Ther Nucleic Acids. 14:567–582. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dai B, Kong DL, Tian J, Liu TW, Zhou H and
Wang ZF: microRNA-1205 promotes cell growth by targeting APC2 in
lung adenocarcinoma. Eur Rev Med Pharmacol Sci. 23:1125–1133.
2019.PubMed/NCBI
|
|
6
|
Song J, Wang W, Wang Y, Qin Y, Wang Y,
Zhou J, Wang X, Zhang Y and Wang Q: Epithelial-mesenchymal
transition markers screened in a cell-based model and validated in
lung adenocarcinoma. BMC Cancer. 19:6802019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cong Z, Diao Y, Xu Y, Li X, Jiang Z, Shao
C, Ji S, Shen Y, De W and Qiang Y: Long non-coding RNA linc00665
promotes lung adenocarcinoma progression and functions as ceRNA to
regulate AKR1B10-ERK signaling by sponging miR-98. Cell Death Dis.
10:842019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu C, Li H, Jia J, Ruan X, Liu Y and
Zhang X: High metastasis-associated lung adenocarcinoma transcript
1 (MALAT1) expression promotes proliferation, migration, and
invasion of non-small cell lung cancer via ERK/mitogen-activated
protein kinase (MAPK) signaling pathway. Med Sci Monit.
25:5143–5149. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Samatar AA and Poulikakos PI: Targeting
RAS-ERK signalling in cancer: Promises and challenges. Nat Rev Drug
Discov. 13:928–942. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tanimura S and Takeda K: ERK signalling as
a regulator of cell motility. J Biochem. 162:145–154. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Corcoran RB, Ebi H, Turke AB, Coffee EM,
Nishino M, Cogdill AP, Brown RD, Della Pelle P, Dias-Santagata D,
Hung KE, et al: EGFR-mediated re-activation of MAPK signaling
contributes to insensitivity of BRAF mutant colorectal cancers to
RAF inhibition with vemurafenib. Cancer Discov. 2:227–235. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang LF, Li X, Gao YB, Wang SM, Zhao L,
Dong J, Yao BW, Xu XP, Chang GM, Zhou HM, et al: Activation of
VEGF/Flk-1-ERK pathway induced blood-brain barrier injury after
microwave exposure. Mol Neurobiol. 52:478–491. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Garces S, Yin CC, Patel KP, Khoury JD,
Manning JT Jr, Li S, Xu J, Pina-Oviedo S, Johnson MR, González S,
et al: Focal Rosai-Dorfman disease coexisting with lymphoma in the
same anatomic site: A localized histiocytic proliferation
associated with MAPK/ERK pathway activation. Mod Pathol. 32:16–26.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen RH, Sarnecki C and Blenis J: Nuclear
localization and regulation of erk- and rsk-encoded protein
kinases. Mol Cell Biol. 12:915–927. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lenormand P, Sardet C, Pages G, L'Allemain
G, Brunet A and Pouyssegur J: Growth factors induce nuclear
translocation of MAP kinases (p42mapk and p44mapk) but not of their
activator MAP kinase kinase (p45mapkk) in fibroblasts. J Cell Biol.
122:1079–1088. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chang WL, Liu YW, Dang YL, Jiang XX, Xu H,
Huang X, Wang YL, Wang H, Zhu C, Xue LQ, et al: PLAC8, a new marker
for human interstitial extravillous trophoblast cells, promotes
their invasion and migration. 145(pii): dev148932. 2018.
|
|
17
|
Galaviz-Hernandez C, Stagg C, de Ridder G,
Tanaka TS, Ko MS, Schlessinger D and Nagaraja R: Plac8 and Plac9,
novel placental-enriched genes identified through microarray
analysis. Gene. 309:81–89. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kaistha BP, Lorenz H, Schmidt H, Sipos B,
Pawlak M, Gierke B, Kreider R, Lankat-Buttgereit B, Sauer M,
Fiedler L, et al: PLAC8 localizes to the inner plasma membrane of
pancreatic cancer cells and regulates cell growth and disease
progression through critical cell-cycle regulatory pathways. Cancer
Res. 76:96–107. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li C, Ma H, Wang Y, Cao Z, Graves-Deal R,
Powell AE, Starchenko A, Ayers GD, Washington MK, Kamath V, et al:
Excess PLAC8 promotes an unconventional ERK2-dependent EMT in colon
cancer. J Clin Invest. 124:2172–2187. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu SF, Huang Y, Hou JK, Yuan TT, Zhou CX,
Zhang J and Chen GQ: The downregulation of onzin expression by
PKCepsilon-ERK2 signaling and its potential role in AML cell
differentiation. Leukemia. 24:544–551. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kinsey C, Balakrishnan V, O'Dell MR, Huang
JL, Newman L, Whitney-Miller CL, Hezel AF and Land H: Plac8 links
oncogenic mutations to regulation of autophagy and is critical to
pancreatic cancer progression. Cell Rep. 7:1143–1155. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mourtada-Maarabouni M, Watson D, Munir M,
Farzaneh F and Williams GT: Apoptosis suppression by candidate
oncogene PLAC8 is reversed in other cell types. Curr Cancer Drug
Targets. 13:80–91. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jin Z, Guan L, Xiang GM and Gao BA:
Radiation resistance of the lung adenocarcinoma is related to the
AKT-Onzin-POU5F1 axis. Biochem Biophys Res Commun. 499:538–543.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Hu Q, Li G, Li L, Liang S, Zhang
Y, Liu J, Fan Z, Li L, Zhou B, et al: ONZIN upregulation by mutant
p53 contributes to osteosarcoma metastasis through the CXCL5-MAPK
signaling pathway. Cell Physiol Biochem. 48:1099–1111. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Segawa S, Kondo Y, Nakai Y, Iizuka A,
Kaneko S, Yokosawa M, Furuyama K, Tsuboi H, Goto D, Matsumoto I and
Sumida T: Placenta specific 8 suppresses IL-18 production through
regulation of autophagy and is associated with adult still disease.
J Immunol. 201:3534–3545. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao L, Liu S, Xu J, Li W, Duan G, Wang H,
Yang H, Yang Z and Zhou R: A new molecular mechanism underlying the
EGCG-mediated autophagic modulation of AFP in HepG2 cells. Cell
Death Dis. 8:e31602017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gong C, Fang J, Li G, Liu HH and Liu ZS:
Effects of microRNA-126 on cell proliferation, apoptosis and tumor
angiogenesis via the down-regulating ERK signaling pathway by
targeting EGFL7 in hepatocellular carcinoma. Oncotarget.
8:52527–52542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang G, Sun J, Liu G, Fu Y and Zhang X:
Bradykinin promotes cell proliferation, migration, invasion, and
tumor growth of gastric cancer through ERK signaling pathway. J
Cell Biochem. 118:4444–4453. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Verma N, Muller AK, Kothari C,
Panayotopoulou E, Kedan A, Selitrennik M, Mills GB, Nguyen LK, Shin
S, Karn T, et al: Targeting of PYK2 Synergizes with EGFR
Antagonists in Basal-like TNBC and Circumvents HER3-Associated
Resistance via the NEDD4-NDRG1 Axis. Cancer Res. 77:86–99. 2010.
View Article : Google Scholar
|
|
30
|
Tanimoto A, Takeuchi S, Arai S, Fukuda K,
Yamada T, Roca X, Ong ST and Yano S: Histone deacetylase 3
inhibition overcomes BIM deletion polymorphism-mediated osimertinib
resistance in EGFR-mutant lung cancer. Clin Cancer Res.
23:3139–3149. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jia Y, Ying X, Zhou J, Chen Y, Luo X, Xie
S, Wang QC, Hu W and Wang L: The novel KLF4/PLAC8 signaling pathway
regulates lung cancer growth. Cell Death Dis. 9:6032018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zou ZQ, Zhang LN, Wang F, Bellenger J,
Shen YZ and Zhang XH: The novel dual PI3K/mTOR inhibitor GDC-0941
synergizes with the MEK inhibitor U0126 in non-small cell lung
cancer cells. Mol Med Rep. 5:503–508. 2012.PubMed/NCBI
|
|
33
|
Ochi N, Takigawa N, Harada D, Yasugi M,
Ichihara E, Hotta K, Tabata M, Tanimoto M and Kiura K: Src mediates
ERK reactivation in gefitinib resistance in non-small cell lung
cancer. Exp Cell Res. 322:168–177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ito Y, Sasaki Y, Horimoto M, Wada S,
Tanaka Y, Kasahara A, Ueki T, Hirano T, Yamamoto H, Fujimoto J, et
al: Activation of mitogen-activated protein kinases/extracellular
signal-regulated kinases in human hepatocellular carcinoma.
Hepatology. 27:951–958. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Marks JL, Gong Y, Chitale D, Golas B,
McLellan MD, Kasai Y, Ding L, Mardis ER, Wilson RK, Solit D, et al:
Novel MEK1 mutation identified by mutational analysis of epidermal
growth factor receptor signaling pathway genes in lung
adenocarcinoma. Cancer Res. 68:5524–5528. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gao X, Chen G, Gao C, Zhang DH, Kuan SF,
Stabile LP, Liu G and Hu J: MAP4K4 is a novel MAPK/ERK pathway
regulator required for lung adenocarcinoma maintenance. Mol Oncol.
11:628–639. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shigematsu H, Lin L, Takahashi T, Nomura
M, Suzuki M, Wistuba II, Fong KM, Lee H, Toyooka S, Shimizu N, et
al: Clinical and biological features associated with epidermal
growth factor receptor gene mutations in lung cancers. J Natl
Cancer Inst. 97:339–346. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shu Y, Wu X, Tong X, Wang X, Chang Z, Mao
Y, Chen X, Sun J, Wang Z, Hong Z, et al: Circulating tumor DNA
mutation profiling by targeted next generation sequencing provides
guidance for personalized treatments in multiple cancer types. Sci
Rep. 7:5832017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kato Y, Hosomi Y, Watanabe K, Yomota M,
Kawai S, Okuma Y, Kubota K, Seike M, Gemma A and Okamura T: Impact
of clinical features on the efficacy of osimertinib therapy in
patients with T790M-positive non-small cell lung cancer and
acquired resistance to epidermal growth factor receptor tyrosine
kinase inhibitors. J Thorac Dis. 11:2350–2360. 2019. View Article : Google Scholar : PubMed/NCBI
|