Epigenetic mechanisms are implicated in
tumorigenesis and cancer progression, and are defined as heritable
changes that do not affect the DNA sequence. Examples include DNA
methylation, histone modification and microRNA (miR) interference
(1). Histone modification serves an
important role in transcriptional regulation, DNA repair and
replication, and chromosomal condensation. Several studies have
indicated that histone modifications typically occur at the
N-terminus, primarily in the form of acetylation, methylation,
phosphorylation or ubiquitination (2,3).
Ubiquitination describes a post-translational
modification of proteins under the conditions of normal homeostasis
or disease, which involves the addition of the evolutionarily
conserved small protein ubiquitin (4) or UBLs (ubiquitin-like proteins)
(5) that tag the protein for
proteasomal degradation or non-degradative processes (6). Ubiquitin is a 76-amino acid polypeptide
that can covalently conjugate with protein substrates via a
mechanism involving three enzymes: Ubiquitin-activating (E1),
ubiquitin-conjugating (E2) and ubiquitin ligase (E3). The
ubiquitination of a specific protein substrate requires the
selective recruitment of E1, E2 and E3 (7–9). In
eukaryotic cells, the structure of ubiquitin is highly conserved
and the protein responds to certain chemical signals (such as
phosphorylation, oxidation, misfolding and damage to the
ubiquitinated protein) to induce the ubiquitin-proteasome
degradation pathway (10). Notably,
the activity of deubiquitinating enzymes (DUBs) directly influences
the turnover rate, activity, regeneration and localization of
various proteins in cells. In addition, DUBs serve an important
role in homeostasis (11), the
stabilization and degradation of proteins, and signal transduction
pathways (11). Changes in protein
structure, abnormal spatial and temporal expression, and
uncontrolled activity can result in the development of certain
conditions, including arthritis, neurodegenerative and
cardiovascular diseases, and tumors. In humans, DUBs can serve a
role in the genesis of tumors as either oncogenes or tumor
suppressor genes.
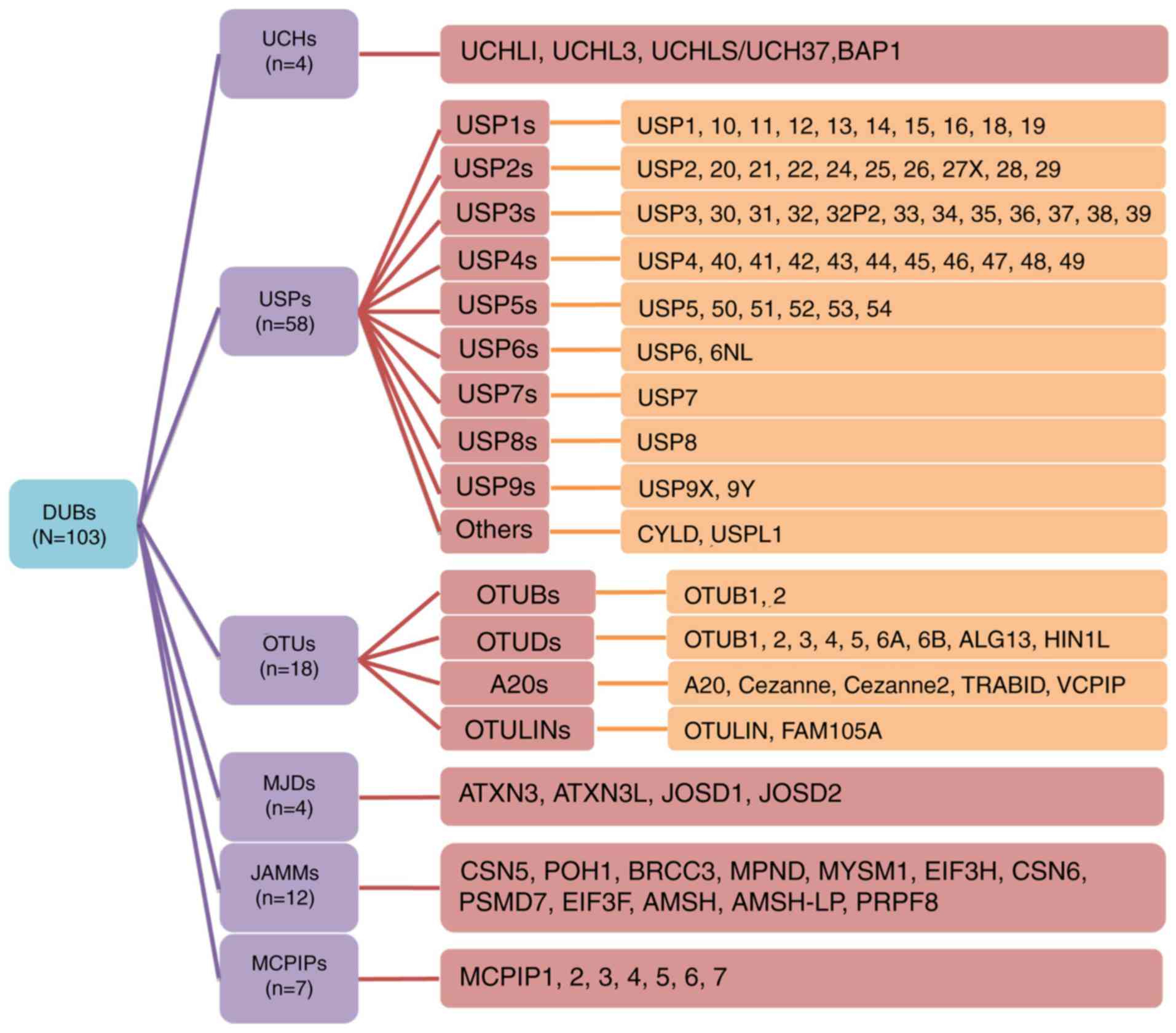
The DUB protein family reportedly comprises 103
members, the majority of which are cysteine proteases. As a result
of similarities in their amino acid sequences and molecular
structures, these proteins can be divided into the following six
families: Ubiquitin C-terminal hydrolases (UCHs) (12), ubiquitin-specific proteases (USPs),
ovarian tumor-related proteases (OTUs), Machado-Joseph disease
protein domain proteases (MJDs), Jab1/MPN domain-associated
metalloisopeptidase (JAMM) domain proteins and monocyte chemotactic
protein-induced proteins (MCPIPs) (13). To further summarize and stratify the
aforementioned proteins, subfamilies are also detailed in Fig. 1. USP16 (14), USP6NL (15), ubiquitin thioesterase otulin (OTULIN)
(16) and family with sequence
similarity 105 member A (FAM105) (17) were also recently discovered. The
majority of these DUBs are associated with tumor progression and
several studies have revealed the association between DUBs and
gastric cancer (GC) (18,19). Of note, GC is the second leading
cause of cancer-associated mortality and the fourth most common
cancer type worldwide (20,21).
The lack of a comprehensive understanding of the
molecular mechanism underpinning GC metastasis and recurrence
suggests that further investigation is required. Thus, DUBs and
their potential association with the progression of GC were the
primary focus of the present review. Within the present study,
subsequent data analyses were performed using the Gene Expression
Profiling Interactive Analysis (GEPIA) website (http://gepia.cancer-pku.cn), which primary collates
data from The Cancer Genome Atlas and the Genotype-Tissue
Expression project databases. The name of a each target gene was
input into the GEPIA website and the corresponding data was
extracted (22).
The enzymes of the UCH protein family contain a
conserved catalytic domain known as the UCH domain, which comprises
~230 amino acids (23). This protein
family includes four members, including UCHL1/protein gene product
9.5, UCHL3, UCHL5/UCH37 and BRCA1 associated protein-1 (BAP1)
(24–27). The activities of these proteins have
been associated with the occurrence and development of cancer, and
several studies have identified that UCHL1, UCHL5 and BAP1 are
specifically involved in the formation of GC (24–26).
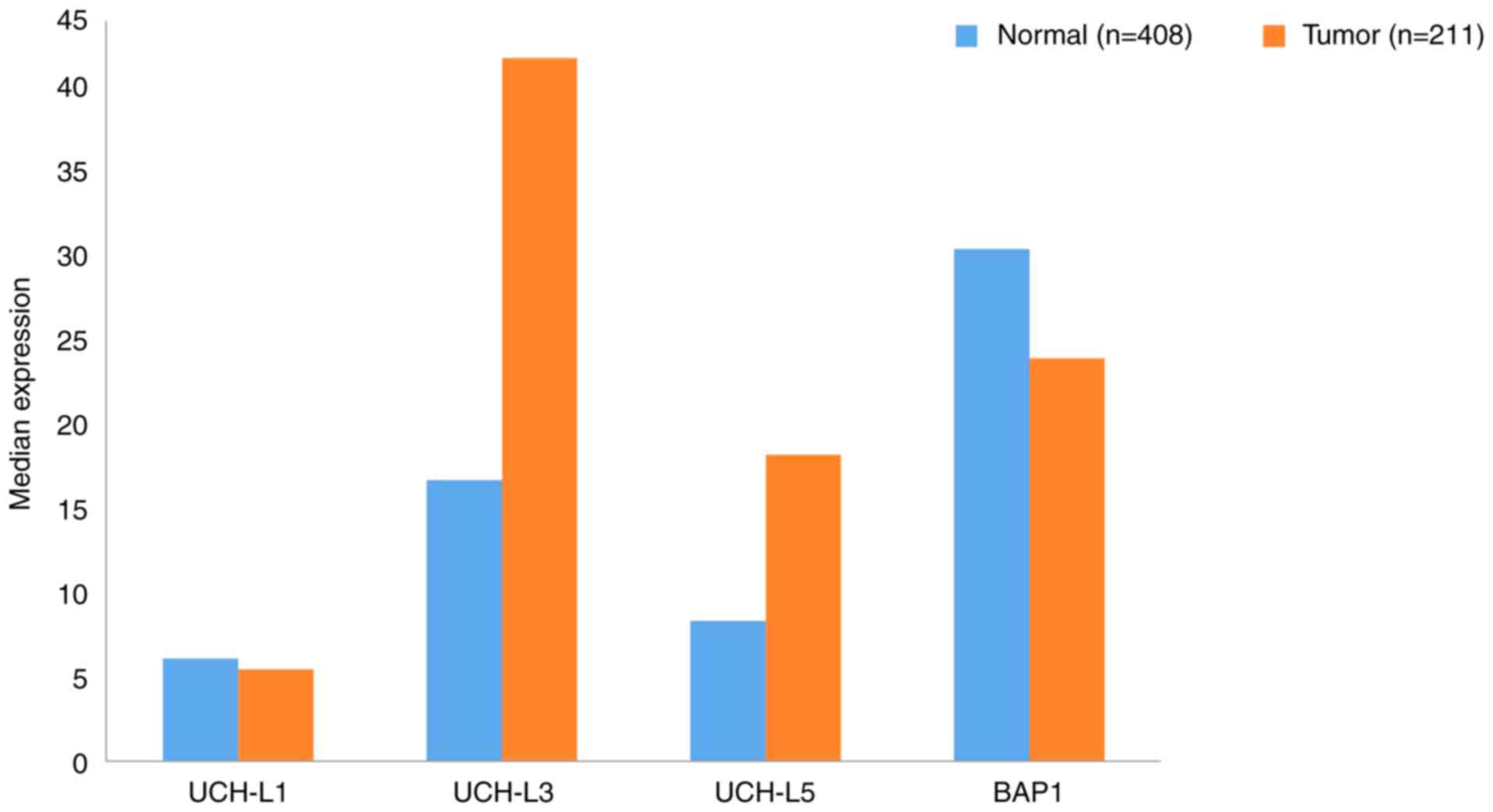
Using data extracted from GEPIA, the gene expression
profiles of UCHs between GC samples and the paired normal tissues
are presented in Fig. 2. The gene
expression levels of UCHL3 and UCHL5 in tumor tissues were
upregulated >2-fold compared with normal tissues. To the best of
our knowledge, no studies have reported the link between UCHL3 and
GC; however, UCHL5 has been identified as a potential biomarker of
GC with novel prognostic value. For elderly patients with
dysregulated protein homeostasis, high levels of UCHL5 inhibited
proteasome activity, and were determined to promote the apoptosis
of cancer cells (28). Regarding
UCHL1, research has shown that it could be a candidate biomarker
and therapeutic target for GC metastasis, as UCHL1 promoted this
process via the Akt and Erk1/2 pathways (29). BAP1 expression is downregulated in
gastric carcinoma and its decreased expression was associated with
a malignant phenotype (histological grade) and a more advanced TNM
stage (30). Furthermore, low BAP1
expression was revealed to be associated with poor prognosis in
patients with gastric adenocarcinoma and GC (30).
Associations between UCHs and certain
clinicopathological features, and the 5-years survival rates of
patients with GC are presented in Table
I. High expression levels of UCHs in patients with GC were
predominantly associated with tumor size and TNM stage, but not sex
or age. Analysis of these expression levels indicated that the
higher the degree of positive BAP1 and UCHL5 expression in GC, the
higher the 5-year survival rate of patients. Conversely, increased
expression of UCHL1 was revealed to reduce the survival rate of
patients.
USPs are the most diverse family of DUBs and the USP
subclass represents the majority of DUBs encoded by the human
genome. Consequently, numerous studies have investigated their
function, substrates and mechanisms of action in various diseases.
The discovery of gene mutations and the upregulation of USPs in
various types of cancer, and their potential for targeted small
molecule-mediated inhibition, indicates USPs as a promising
therapeutic target. There is also increasing interest in the
development of USP-specific inhibitors as antiviral and anticancer
agents (31). In the present study,
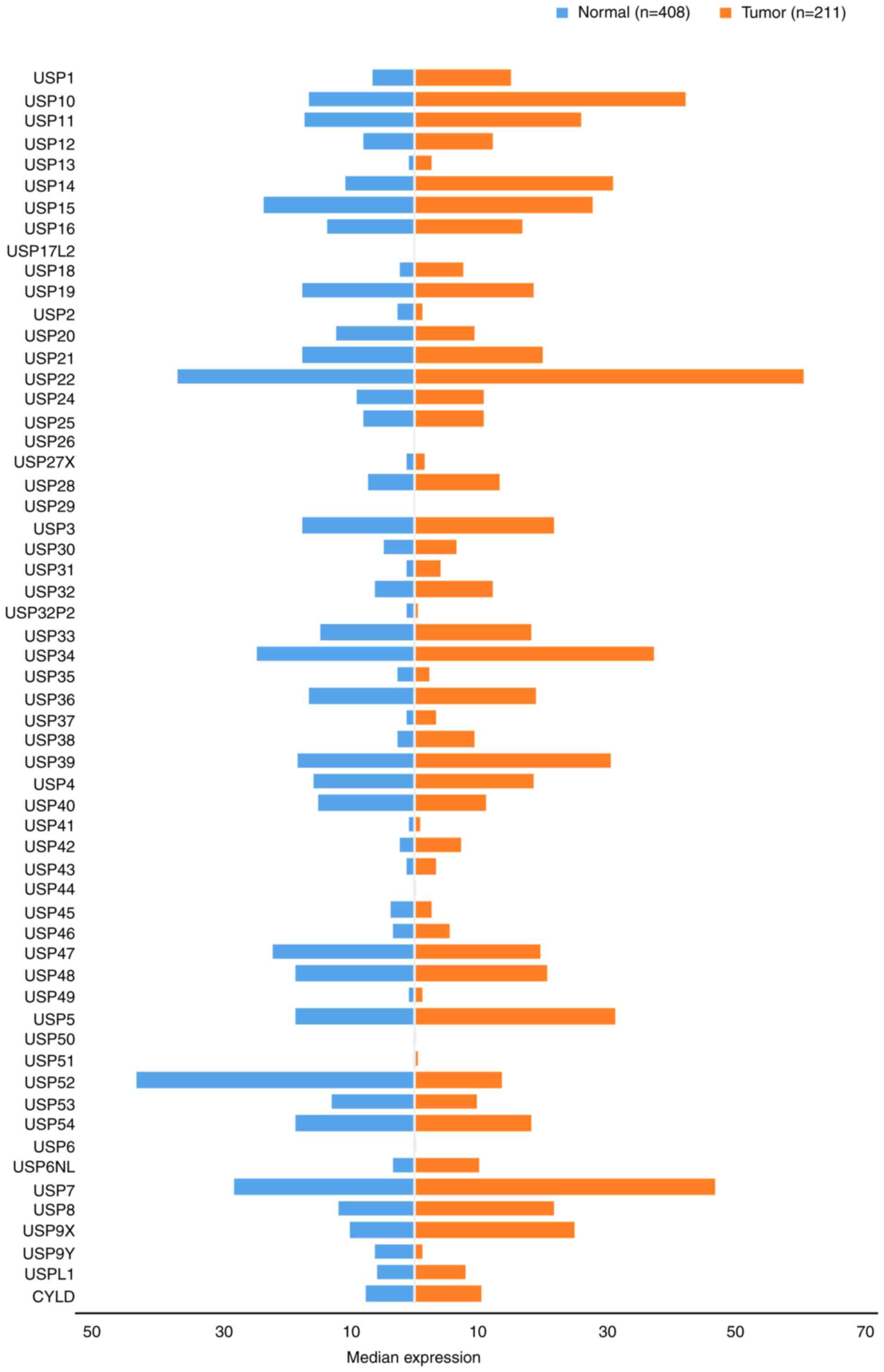
the USP family was stratified into 10 subgroups comprising USPs
1–10. The gene expression profile of USPs was compared between GC
and paired normal tissues (Fig.
3).
The expression of the majority of USP1s has been
associated with tumor growth, though studies into GC have
predominantly investigated USP10, USP14 and USP15 (43). USP10 and USP14 are independent
predictors of prognosis for patients with GC, and the increased
expression of USP10 in GC has been associated with the 5-year
survival rate of patients. A previous study demonstrated that the
downregulation of USP10, as well as the absence of USP14 expression
in GC cells had significant effects on gastric wall invasion and
lymph node metastasis, increased malignant biological behavior and
reduced survival rate, as determined from a large number of
clinical samples (44,45). Conversely, vimentin expression was
upregulated in human GC tissues and cell lines as a result of
deubiquitination, following interactions with USP14 and miR-320a,
which may contribute to the aggressiveness of GC cells (46). It was also reported that the direct
targeting of USP14 and vimentin by miR-320a inhibited GC cell
proliferation, migration and invasion. miR-320a not only directly
suppresses vimentin expression, but also binds to USP14, indirectly
downregulating vimentin in GC tissues. Therefore, a high positive
expression rate of USP14 correlates with a high recurrence rate in
patients with GC (44,45).
USP20, USP22 and USP28 have been previously
determined to be associated with GC. Compared with normal tissues,
high expression levels of USP28 were detected in GC tissues, and
were also associated with the distant metastasis of tumors.
Conversely, USP28 downregulation may significantly inhibit the
proliferation and migration of GC cells; however, the effects of
USP28 expression on the proliferation and migration of gastric
epithelial mucosal cell lines were not significant (46). The aforementioned findings provide a
novel insight for the development of therapeutic strategies to
treat GC via the regulation of USP28 (56). USP20 also serves an important role in
gastric tumorigenesis and progression. A negative association
between USP20 expression and tumor size, tumor invasion and TNM
stage has previously been reported (Table I). It was revealed that USP20
expression negatively correlated with patient prognosis and its
anti-tumor activity. The mechanism underlying the effects of USP20
included the positive regulation of claspin stabilization in GC,
thus, USP20 represents a promising molecular target for the
development of novel therapeutic drugs (57). USP22-mediated protein stabilization
of B cell-specific Moloney murine leukemia virus integration site 1
promotes the stemness of GC stem cells as well as GC progression,
and its expression may also serve an important role in gastric
carcinoma (58–60). Yang et al discussed that USP22
expresion is correlate with cancer progression. Where they found
that around 57% of gastric cancer tisues showed high expression of
USP22 comparing with normal connective tissue. This overexpression
of USP22 consequentially effecct on tumor size, inavsion and
metastasis (60). Additionally, both
USP20 and USP22 expression are positively correlated with the
5-year survival rate of patients with GC (57,60).
The USP3 subfamily represents the largest family of
USPs and consists of the following 12 members: USP3 (61), USP 30 (62), USP31 (63,64),
USP32, USP32P2 (22), USP33
(64), USP34 (65), USP35 (66), USP36 (67), USP37 (68), USP38 (69) and USP39 (70). As shown in Fig. 3, excluding USP32P2 and USP35, the
expression of each member was upregulated in GC tissues compared
with normal gastric tissues. The highest expression levels were
exhibited by USP34 (37.21), while the lowest were reported for
USP32P2 (0.33). The expression levels of USP30, USP31, USP32P2,
USP35, USP37 and USP38 were <10 TPM.
USP3 s may also serve as useful biomarkers to
predict the prognosis of patients with GC. Studies investigating
USP3s revealed their ability to influence cell proliferation, cell
cycle regulation and transfer-related protein expression (61). In vivo experiments revealed
that USP3s promoted the growth and metastasis of GC. Additionally,
the high expression levels of these proteins imparted a lower
survival rate in patients (71).
Studies have also discovered that tumor location, tumor
infiltration depth and TNM stage are all associated with USP33
upregulation, and affect the overall survival rate and prognosis of
patients with GC. USP33 may also be linked with the prognosis of GC
(62), and its high expression
levels indicated longer survival times in patients (72).
It was also determined that short hairpin
RNA-mediated downregulation of USP39, another member of the USP3
subfamily, inhibited GC cell proliferation and colony formation.
USP39 inhibition also induced G2/M phase arrest and
increased poly (ADP-ribose) polymerase cleavage (Asp214) suggesting
that USP39 is critical for GC cell proliferation. As USP39 is
upregulated in certain types of cancer, and hyperproliferation is a
hallmark of cancer, USP39 may represent a potential therapeutic
target for the treatment of several cancer types (73). By contrast, miR-133a expression was
inversely correlated with USP39, which it directly targets by
binding at the 3′-untranslated region; the high expression rate of
USP39 indicated a longer survival time for patients (74).
Studies into GC have investigated USP42, USP44 and
USP47. It has been reported that USP47 may represent a drug
resistant target for GC. Additionally, it was determined that
miR-204-5p was downregulated in GC, and may inhibit the
proliferation of GC cells by targeting USP47 and RAB22A, thus
serving a role in suppressing cancer development. Therefore, the
recovery of miR-204-5p expression may be a potential therapeutic
strategy for the treatment of GC (83,84).
In vitro analyses also demonstrated that USP42 silencing
suppressed cell proliferation by inducing
G0/G1 arrest, and inhibited cellular invasion
via matrix metalloprotease and epithelial-mesenchymal transition
regulation. The increased expression of USP42 may be important in
tumor progression and the metastasis of GC, and may serve as a
prognostic marker (85). The
combination of USP44 expression and DNA ploidy status may also
serve as an independent prognostic marker in GC. Notably, the
expression rate of USP44 in GC is negatively correlated with the
5-year survival rate of patients (86).
USP7 is currently the only member of the USP7
subfamily. Its expression levels in GC tissues are higher than
those in normal tissue (46.63 TPM and 28.04 TPM, respectively;
Fig. 3). Studies investigating USP7
and its relation to GC are yet to be performed; however, H.
pylori was reported to affect the expression of the USP family
via alternative H. pylori-specific mechanisms distinct from
the conserved signaling pathways, during the activation of the
innate immune response (18).
The roles of USP8 and its substrate (epidermal
growth factor) have been evaluated in cancer therapy, and their
possible targeting for the treatment of Cushing's disease has been
investigated (96); USP8 is the only
member of the USP8 subfamily. As shown in Fig. 3, USP8 expression in GC tissues was
increased ~2-fold compared with that in normal tissues.
Additional USP family members include CYLD lysine 63
deubiquitinase (CYLD) and USPL1. Their expression in GC tissues was
notably increased compared with normal tissues (Fig. 3). The CYLD signaling pathway serves a
biological function similar to that of the oncogenes in
gastrointestinal tumors, and has been associated with the
occurrence and development of GC (99,100).
Moreover, genetic variations affecting USPL1 expression have been
linked to breast cancer (101).
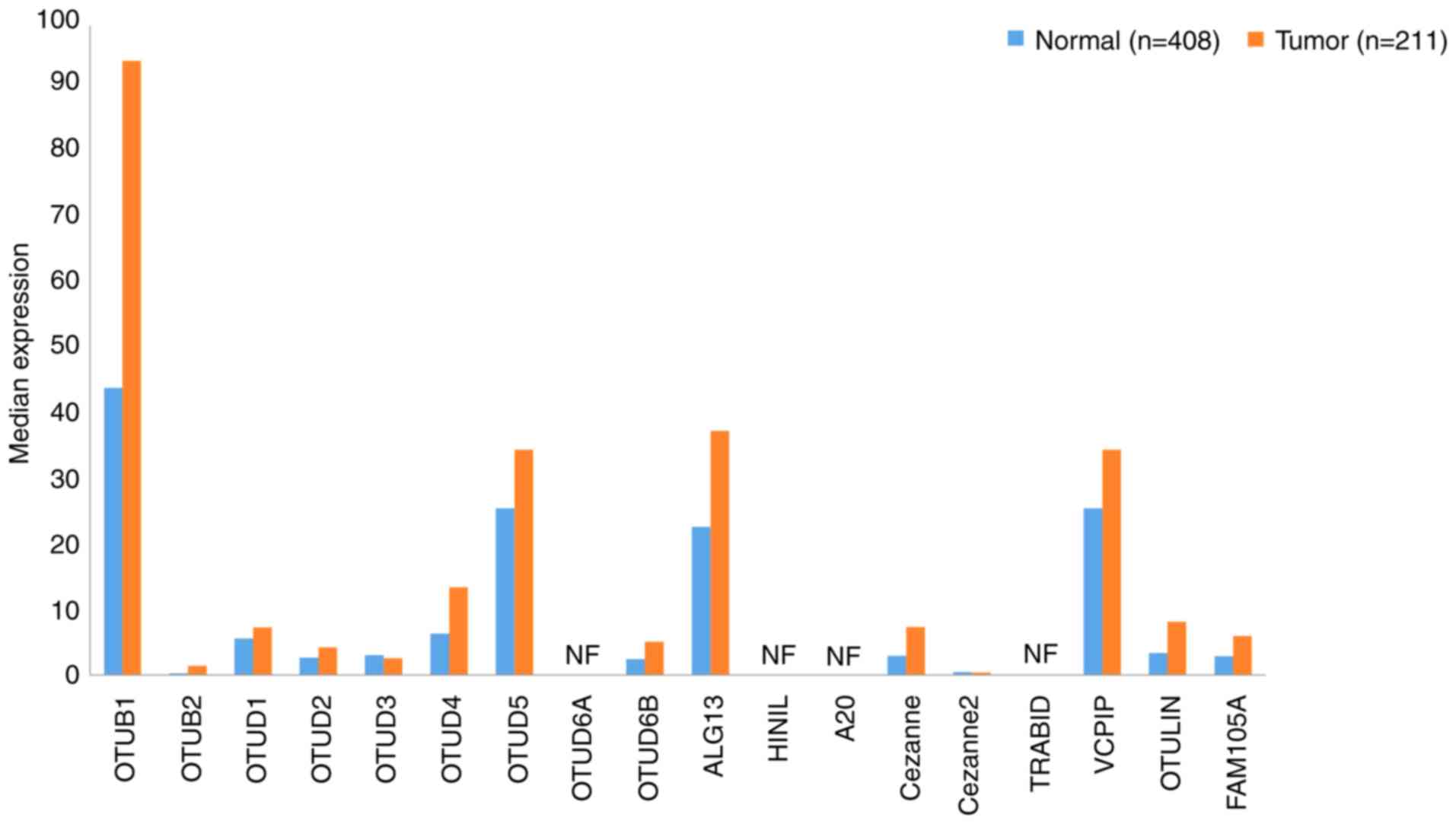
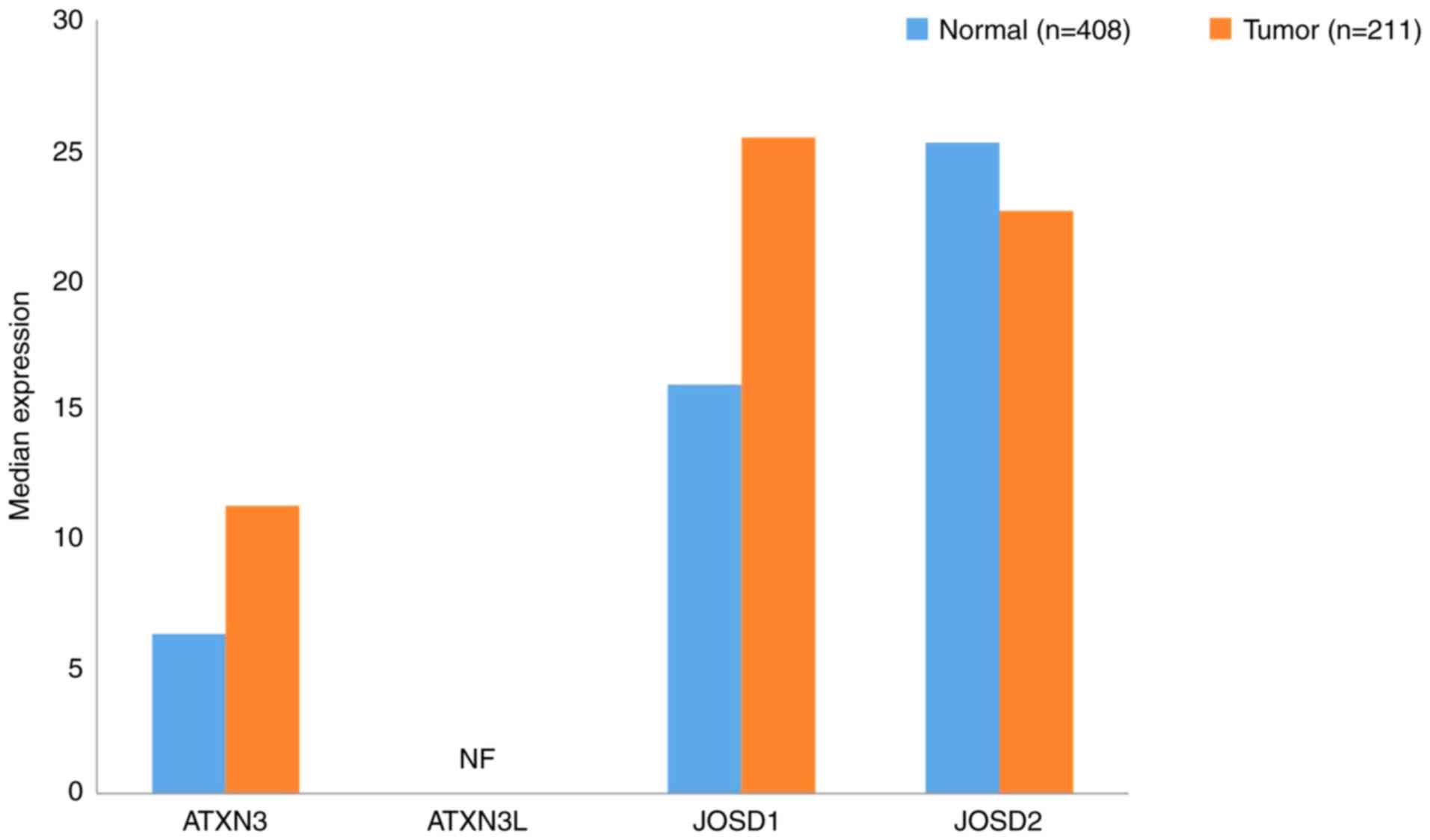
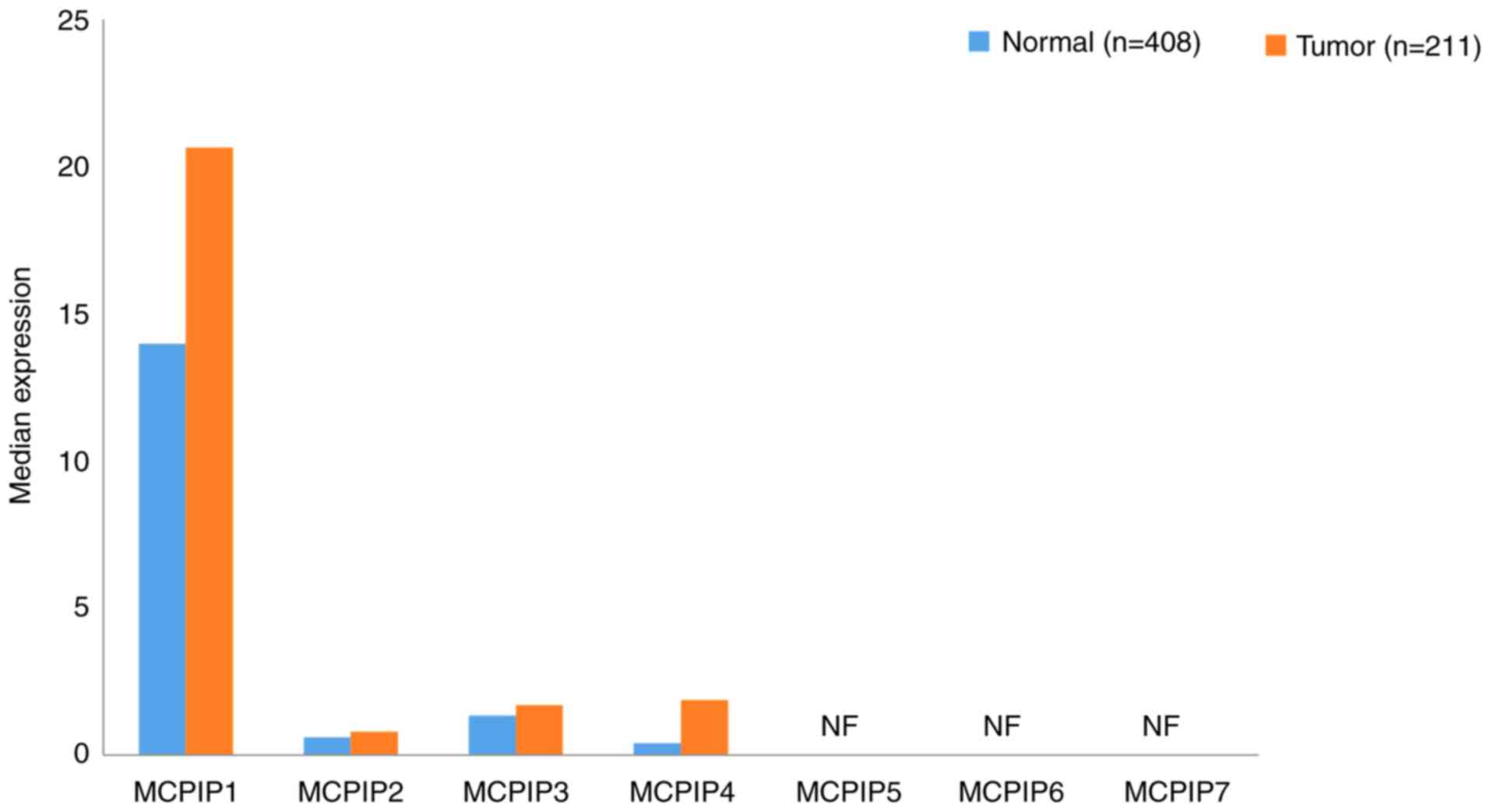
A total of 18 OTU family DUBs exist in humans, the
majority of which have been associated with the prognosis of
patients with tumors. OTUs can be divided into four categories:
OTUBs, OTUDs, A20s and OTULINs (102). The gene expression profiles of OTUs
in GC samples and paired normal tissues ware presented in Fig. 4.
OTUDs are the largest class of DUBs, which comprises
the following nine members: OTUD1-5 (106–110),
OTUD6A (111) and B (112), UDP-N-acetylglucosamine transferase
subunit ALG13 homolog (ALG13) and hematological and neurological
expressed 1 protein (HIN1 L) (102). Using data extracted from GEPIA, the
expression levels of both of the OTUD subfamily members in GC and
normal tissues were determined to be relatively low. OTUD6A and
HIN1L were undetectable, although the expression levels of OTUD4
and OTUD5 were >10 in GC tissues. Further investigation is
required to determine the pathophysiological role of OTUDs in GC
progression.
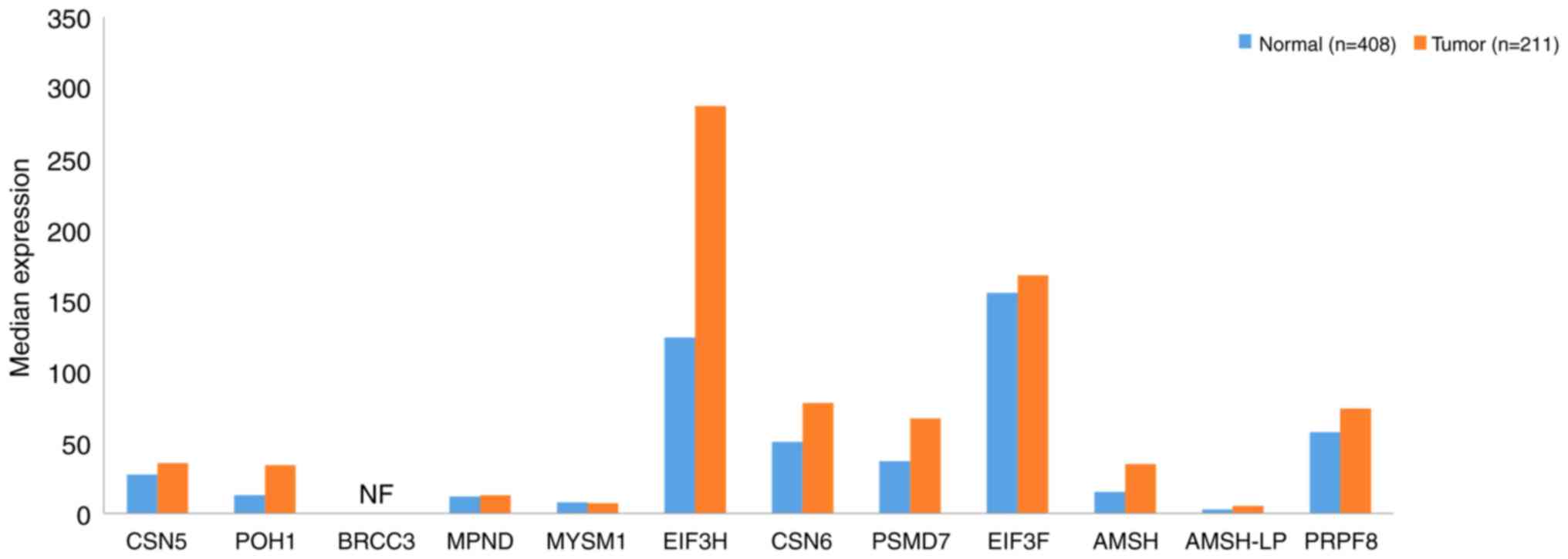
The JAMM subfamily comprises 12 members, including
COP9 signalsome subunit (CSN)5, 26S proteasome non-ATPase
regulatory subunit 14 (POH1) (122), BRCA1/BRCA2-containing complex
subunit 3 (BRCC3) (123), MPN
domain containing (MPND) (124),
myb-like SWIRM and MPN domains 1 (MYSM1) (125), eukaryotic translation initiation
factor 3 subunit (EIF3)H, CSN6 (126), 26S proteasome non-ATPase regulatory
subunit 7 (PSMD7) (127), EIF3F,
anti-Müllerian hormone (AMSH) (128), AMSH-LP (129) and pre-mRNA-processing-splicing
factor 8 (PRPF8) (130). The data
presented in Fig. 6 demonstrate that
the expression levels of JAMMs in GC tissues were upregulated
compared with those in normal tissues, particularly EIF3H and
EIF3F, in which the expression levels were >120. The expression
of BRCC3 was not detected. These findings suggest that the
inhibition of CSN5 may result in a significant increase in p53
levels, indicating that CSN5 may be a crucial regulator of p53 and
its associated intracellular signaling pathway, via CSN5-mediated
cell activity.
Moreover, upregulation of CSN5 has been
significantly associated with the progression of GC; therefore,
CSN5 may represent a novel target for the treatment of this disease
(131). EIF3H was also reported to
influence the progression of GC (132), and therefore, may serve as a
potential therapeutic target. In particular, the strategy of
inhibiting EIF3H expression may suppress the progression of GC and
improve patient prognosis (131).
Furthermore, EIF3F was determined to serve an important role in the
recurrence of GC; increased expression rates of EIF3F in GC were
associated with higher 5-year survival rates of patients (133).
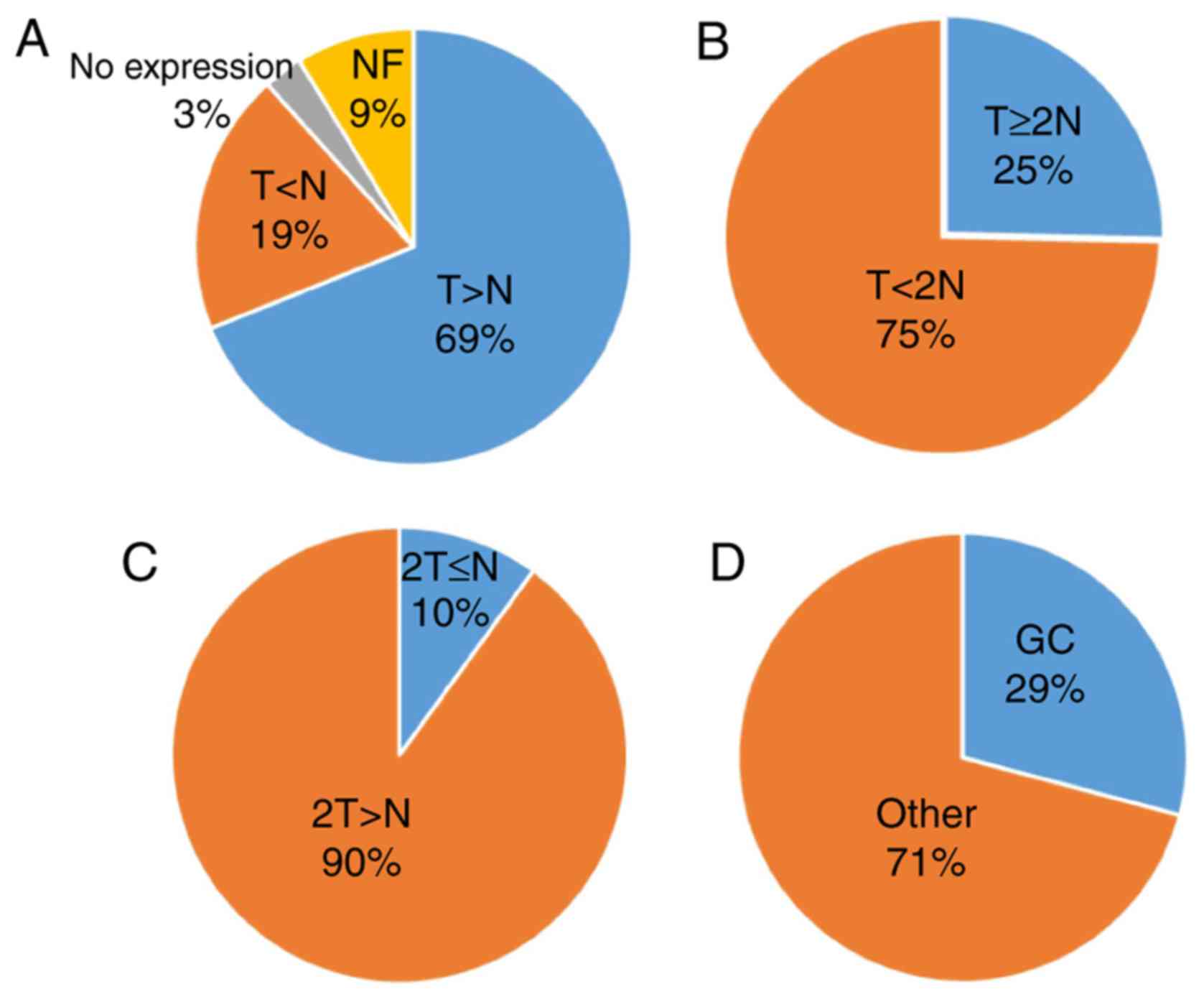
On the contrary, the number of DUBs associated with
GC was determined to be 29%. Following analysis of data from
previously published studies (Table
I), the expression of DUBs in GC and normal tissues was not
determined to be associated with either sex or age; however, an
association between DUBs and tumor size, classification and staging
was observed. In addition, the expression level of DUBs was
significantly associated with the 5-year survival rate of patients
with GC. Among the upregulated genes in GC, six DUBs were linked to
a high 5-year survival rate, though the difference between the two
was not significant. Thus, DUBs may serve a dual role in the
prognosis of GC. However, further investigation is required.
Providing that DUBs can be divided into two categories according to
the prognosis of GC, the common features associated with this
disease and DUBs may be identified, in which DUBs may be considered
in the development of treatments for GC.
Not applicable.
No funding was received.
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
JS, XS and YG conducted literature searching and
wrote this review. JS conducted the data analysis. The language of
the review was edited by MAAM.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Pidsley R, Lawrence MG, Zotenko E,
Niranjan B, Statham A, Song J, Chabanon RM, Qu W, Wang H, Richards
M, et al: Enduring epigenetic landmarks define the cancer
microenvironment. Genome Res. 28:625–638. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zentner GE and Henikoff S: High-resolution
digital profiling of the epigenome. Nat Rev Genet. 15:814–827.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Onder O, Sidoli S, Carroll M and Garcia
BA: Progress in epigenetic histone modification analysis by mass
spectrometry for clinical investigations. Expert Rev Proteomics.
12:499–517. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Swatek KN and Komander D: Ubiquitin
modifications. Cell Res. 26:399–422. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rogov V, Dotsch V, Johansen T and Kirkin
V: Interactions between autophagy receptors and ubiquitin-like
proteins form the molecular basis for selective autophagy. Mol
Cell. 53:167–178. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hershko A: Ubiquitin: Roles in protein
modification and breakdown. Cell. 34:11–12. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pickart CM: Mechanisms underlying
ubiquitination. Annu Rev Biochem. 70:503–533. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hershko A and Ciechanover A: The ubiquitin
system. Annu Rev Biochem. 67:425–479. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Finley D, Ciechanover A and Varshavsky A:
Ubiquitin as a central cellular regulator. Cell. 116 (Suppl
2):S29–S32. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhou MJ, Chen FZ and Chen HC:
Ubiquitination involved enzymes and cancer. Med Oncol. 31:932014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Johnston SC, Riddle SM, Cohen RE and Hill
CP: Structural basis for the specificity of ubiquitin C-terminal
hydrolases. EMBO J. 18:3877–3887. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fang Y and Shen X: Ubiquitin
carboxyl-terminal hydrolases: Involvement in cancer progression and
clinical implications. Cancer Metastasis Rev. 36:669–682. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
McDonough M, Sangan P and Gonda DK:
Characterization of novel yeast RAD6 (UBC2) ubiquitin-conjugating
enzyme mutants constructed by charge-to-alanine scanning
mutagenesis. J Bacteriol. 177:580–585. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu JC, Dawson VL and Dawson TM: Usp16: Key
controller of stem cells in Down syndrome. EMBO J. 32:2788–2789.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Avanzato D, Pupo E, Ducano N, Isella C,
Bertalot G, Luise C, Pece S, Bruna A, Rueda OM, Caldas C, et al:
High USP6NL levels in breast cancer sustain chronic AKT
phosphorylation and GLUT1 stability fueling aerobic glycolysis.
Cancer Res. 78:3432–3444. 2018.PubMed/NCBI
|
|
16
|
Weber A, Elliott PR, Pinto-Fernandez A,
Bonham S, Kessler BM, Komander D, El Oualid F and Krappmann D: A
linear diubiquitin-based probe for efficient and selective
detection of the deubiquitinating enzyme OTULIN. Cell Chem Biol.
24:1299–1313.e7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Taneera J, Fadista J, Ahlqvist E, Atac D,
Ottosson-Laakso E, Wollheim CB and Groop L: Identification of novel
genes for glucose metabolism based upon expression pattern in human
islets and effect on insulin secretion and glycemia. Hum Mol Genet.
24:1945–1955. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Coombs N, Sompallae R, Olbermann P,
Gastaldello S, Goppel D, Masucci MG and Josenhans C: Helicobacter
pylori affects the cellular deubiquitinase USP7 and
ubiquitin-regulated components TRAF6 and the tumour suppressor p53.
Int J Med Microbiol. 301:213–224. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saldana M, VanderVorst K, Berg AL, Lee H
and Carraway KL: Otubain 1: A non-canonical deubiquitinase with an
emerging role in cancer. Endocr Relat Cancer. 26:R1–R14. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rahman R, Asombang AW and Ibdah JA:
Characteristics of gastric cancer in Asia. World J Gastroenterol.
20:4483–5890. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Todi SV and Paulson HL: Balancing act:
Deubiquitinating enzymes in the nervous system. Trends Neurosci.
34:370–382. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fang Y, Fu D and Shen XZ: The potential
role of ubiquitin c-terminal hydrolases in oncogenesis. Biochim
Biophys Acta. 1806:1–6. 2010.PubMed/NCBI
|
|
25
|
Kim HJ, Kim YM, Lim S, Nam YK, Jeong J,
Kim HJ and Lee KJ: Ubiquitin C-terminal hydrolase-L1 is a key
regulator of tumor cell invasion and metastasis. Oncogene.
28:117–127. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dang LC, Melandri FD and Stein RL: Kinetic
and mechanistic studies on the hydrolysis of ubiquitin C-terminal
7-amido-4-methylcoumarin by deubiquitinating enzymes. Biochemistry.
37:1868–1879. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Case A and Stein RL: Mechanistic studies
of ubiquitin C-terminal hydrolase L1. Biochemistry. 45:2443–2452.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Arpalahti L, Laitinen A, Hagström J,
Mustonen H, Kokkola A, Böckelman C, Haglund C and Holmberg CI:
Positive cytoplasmic UCHL5 tumor expression in gastric cancer is
linked to improved prognosis. PLoS One. 13:e01931252018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gu YY, Yang M, Zhao M, Luo Q, Yang L, Peng
H, Wang J, Huang SK, Zheng ZX, Yuan XH, et al: The de-ubiquitinase
UCHL1 promotes gastric cancer metastasis via the Akt and Erk1/2
pathways. Tumour Biol. 36:8379–8387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yan S, He F, Luo R, Wu H, Huang M, Huang
C, Li Y and Zhou Z: Decreased expression of BRCA1-associated
protein 1 predicts unfavorable survival in gastric adenocarcinoma.
Tumour Biol. 37:6125–6133. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nijman SM, Luna-Vargas MP, Velds A,
Brummelkamp TR, Dirac AM, Sixma TK and Bernards R: A genomic and
functional inventory of deubiquitinating enzymes. Cell.
123:773–786. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Das DS, Das A, Ray A, Song Y, Samur MK,
Munshi NC, Chauhan D and Anderson KC: Blockade of deubiquitylating
enzyme USP1 inhibits DNA repair and triggers apoptosis in multiple
myeloma cells. Clin Cancer Res. 23:4280–4289. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kedersha N, Panas MD, Achorn CA, Lyons S,
Tisdale S, Hickman T, Thomas M, Lieberman J, McInerney GM, Ivanov P
and Anderson P: G3BP-Caprin1-USP10 complexes mediate stress granule
condensation and associate with 40S subunits. J Cell Biol.
212:845–860. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kapadia B, Nanaji NM, Bhalla K, Bhandary
B, Lapidus R, Beheshti A, Evens AM and Gartenhaus RB: Fatty Acid
Synthase induced S6Kinase facilitates USP11-eIF4B complex formation
for sustained oncogenic translation in DLBCL. Nat Commun.
9:8292018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Aron R, Pellegrini P, Green EW, Maddison
DC, Opoku-Nsiah K, Wong JS, Daub AC, Giorgini F and Finkbeiner S:
Publisher correction: Deubiquitinase Usp12 functions
noncatalytically to induce autophagy and confer neuroprotection in
models of Huntington's disease. Nat Commun. 9:43332018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang S, Zhang M, Jing Y, Yin X, Ma P,
Zhang Z, Wang X, Di W and Zhuang G: Deubiquitinase USP13 dictates
MCL1 stability and sensitivity to BH3 mimetic inhibitors. Nat
Commun. 9:2152018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee BH, Lee MJ, Park S, Oh DC, Elsasser S,
Chen PC, Gartner C, Dimova N, Hanna J, Gygi SP, et al: Enhancement
of proteasome activity by a small-molecule inhibitor of USP14.
Nature. 467:179–184. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Eichhorn PJ, Rodon L, Gonzalez-Junca A,
Dirac A, Gili M, Martinez-Saez E, Aura C, Barba I, Peg V, Prat A,
et al: USP15 stabilizes TGF-β receptor I and promotes oncogenesis
through the activation of TGF-β signaling in glioblastoma. Nat Med.
18:429–435. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Adorno M, Sikandar S, Mitra SS, Kuo A,
Nicolis Di Robilant B, Haro-Acosta V, Ouadah Y, Quarta M, Rodriguez
J, Qian D, et al: Usp16 contributes to somatic stem-cell defects in
Down's syndrome. Nature. 501:380–384. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shaw JA, Page K, Blighe K, Hava N, Guttery
D, Ward B, Brown J, Ruangpratheep C, Stebbing J, Payne R, et al:
Genomic analysis of circulating cell-free DNA infers breast cancer
dormancy. Genome Res. 22:220–231. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Malakhov MP, Malakhova OA, Kim KI, Ritchie
KJ and Zhang DE: UBP43 (USP18) specifically removes ISG15 from
conjugated proteins. J Biol Chem. 277:9976–9981. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Combaret L, Adegoke OA, Bedard N, Baracos
V, Attaix D and Wing SS: USP19 is a ubiquitin-specific protease
regulated in rat skeletal muscle during catabolic states. Am J
Physiol Endocrinol Metab. 288:E693–E700. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xie L, Wei J, Qian X, Chen G, Yu L, Ding Y
and Liu B: CXCR4, a potential predictive marker for docetaxel
sensitivity in gastric cancer. Anticancer Res. 30:2209–2216.
2010.PubMed/NCBI
|
|
44
|
Fu Y, Ma G, Liu G, Li B, Li H, Hao X and
Liu L: USP14 as a novel prognostic marker promotes cisplatin
resistance via Akt/ERK signaling pathways in gastric cancer. Cancer
Med. 7:5577–5588. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zeng Z, Wu HX, Zhan N, Huang YB, Wang ZS,
Yang GF, Wang P and Fu GH: Prognostic significance of USP10 as a
tumor-associated marker in gastric carcinoma. Tumour Biol.
35:3845–3853. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhu Y, Zhang Y, Sui Z, Zhang Y, Liu M and
Tang H: USP14 de-ubiquitinates vimentin and miR-320a modulates
USP14 and vimentin to contribute to malignancy in gastric cancer
cells. Oncotarget. 8:48725–48736. 2017.PubMed/NCBI
|
|
47
|
Renatus M, Parrado SG, D'Arcy A, Eidhoff
U, Gerhartz B, Hassiepen U, Pierrat B, Riedl R, Vinzenz D,
Worpenberg S and Kroemer M: Structural basis of ubiquitin
recognition by the deubiquitinating protease USP2. Structure.
14:1293–1302. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Berthouze M, Venkataramanan V, Li Y and
Shenoy SK: The deubiquitinases USP33 and USP20 coordinate beta2
adrenergic receptor recycling and resensitization. EMBO J.
28:1684–1796. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ye Y, Akutsu M, Reyes-Turcu F, Enchev RI,
Wilkinson KD and Komander D: Polyubiquitin binding and
cross-reactivity in the USP domain deubiquitinase USP21. EMBO Rep.
12:350–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang XY, Varthi M, Sykes SM, Phillips C,
Warzecha C, Zhu W, Wyce A, Thorne AW, Berger SL and McMahon SB: The
putative cancer stem cell marker USP22 is a subunit of the human
SAGA complex required for activated transcription and cell-cycle
progression. Mol Cell. 29:102–111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhang L, Lubin A, Chen H, Sun Z and Gong
F: The deubiquitinating protein USP24 interacts with DDB2 and
regulates DDB2 stability. Cell Cycle. 11:4378–4384. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Stouffs K, Lissens W, Tournaye H, Van
Steirteghem A and Liebaers I: Possible role of USP26 in patients
with severely impaired spermatogenesis. Eur J Hum Genet.
13:336–340. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Weber A, Heinlein M, Dengjel J, Alber C,
Singh PK and Häcker G: The deubiquitinase Usp27× stabilizes the
BH3-only protein Bim and enhances apoptosis. EMBO Rep. 17:724–738.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Popov N, Wanzel M, Madiredjo M, Zhang D,
Beijersbergen R, Bernards R, Moll R, Elledge SJ and Eilers M: The
ubiquitin-specific protease USP28 is required for MYC stability.
Nat Cell Biol. 9:765–774. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Liu J, Chung HJ, Vogt M, Jin Y, Malide D,
He L, Dundr M and Levens D: JTV1 co-activates FBP to induce USP29
transcription and stabilize p53 in response to oxidative stress.
EMBO J. 30:846–858. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhao LJ, Zhang T, Feng XJ, Chang J, Suo
FZ, Ma JL, Liu YJ, Liu Y, Zheng YC and Liu HM: USP28 contributes to
the proliferation and metastasis of gastric cancer. J Cell Biochem.
Nov 28–2018.(Epub ahead of print). doi: 10.1002/jcb.28040.
|
|
57
|
Wang C, Yang C, Ji J, Jiang J, Shi M, Cai
Q, Yu Y, Zhu Z and Zhang J: Deubiquitinating enzyme USP20 is a
positive regulator of Claspin and suppresses the malignant
characteristics of gastric cancer cells. Int J Oncol. Mar
8–2017.(Epub ahead of print). doi: 10.3892/ijo.2017.3904.
|
|
58
|
Ma Y, Fu HL, Wang Z, Huang H, Ni J, Song
J, Xia Y, Jin WL and Cui DX: USP22 maintains gastric cancer stem
cell stemness and promotes gastric cancer progression by
stabilizing BMI1 protein. Oncotarget. 8:33329–33342.
2017.PubMed/NCBI
|
|
59
|
He Y, Jin YJ, Zhang YH, Meng HX, Zhao BS,
Jiang Y, Zhu JW, Liang GY, Kong D and Jin XM: Ubiquitin-specific
peptidase 22 overexpression may promote cancer progression and poor
prognosis in human gastric carcinoma. Transl Res. 16:407–416. 2015.
View Article : Google Scholar
|
|
60
|
Yang DD, Cui BB, Sun LY, Zheng HQ, Huang
Q, Tong JX and Zhang QF: The co-expression of USP22 and BMI-1 may
promote cancer progression and predict therapy failure in gastric
carcinoma. Cell Biochem Biophys. 61:703–710. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Nicassio F, Corrado N, Vissers JH, Areces
LB, Bergink S, Marteijn JA, Geverts B, Houtsmuller AB, Vermeulen W,
Di Fiore PP and Citterio E: Human USP3 is a chromatin modifier
required for S phase progression and genome stability. Curr Biol.
17:1972–1977. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Bingol B, Tea JS, Phu L, Reichelt M,
Bakalarski CE, Song Q, Foreman O, Kirkpatrick DS and Sheng M: The
mitochondrial deubiquitinase USP30 opposes parkin-mediated
mitophagy. Nature. 510:370–375. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Tzimas C, Michailidou G, Arsenakis M,
Kieff E, Mosialos G and Hatzivassiliou EG: Human ubiquitin specific
protease 31 is a deubiquitinating enzyme implicated in activation
of nuclear factor-kappaB. Cell Signal. 18:83–92. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Akhavantabasi S, Akman HB, Sapmaz A,
Keller J, Petty EM and Erson AE: USP32 is an active, membrane-bound
ubiquitin protease overexpressed in breast cancers. Mamm Genome.
21:388–397. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sy SM, Jiang J, O WS, Deng Y and Huen MS:
The ubiquitin specific protease USP34 promotes ubiquitin signaling
at DNA double-strand breaks. Nucleic Acids Res. 41:8572–8580. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang Y, Serricchio M, Jauregui M, Shanbhag
R, Stoltz T, Di Paolo CT, Kim PK and McQuibban GA: Deubiquitinating
enzymes regulate PARK2-mediated mitophagy. Autophagy. 11:595–606.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Endo A, Matsumoto M, Inada T, Yamamoto A,
Nakayama KI, Kitamura N and Komada M: Nucleolar structure and
function are regulated by the deubiquitylating enzyme USP36. J Cell
Sci. 122:678–686. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Huang X, Summers MK, Pham V, Lill JR, Liu
J, Lee G, Kirkpatrick DS, Jackson PK, Fang G and Dixit VM:
Deubiquitinase USP37 is activated by CDK2 to antagonize APC(CDH1)
and promote S phase entry. Mol Cell. 42:511–523. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Lin M, Zhao Z, Yang Z, Meng Q, Tan P, Xie
W, Qin Y, Wang RF and Cui J: USP38 Inhibits type I interferon
signaling by editing TBK1 Ubiquitination through NLRP4 Signalosome.
Mol Cell. 64:267–281. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
van Leuken RJ, Luna-Vargas MP, Sixma TK,
Wolthuis RM and Medema RH: Usp39 is essential for mitotic spindle
checkpoint integrity and controls mRNA-levels of aurora B. Cell
Cycle. 7:2710–2719. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Fang CL, Lin CC, Chen HK, Hseu YC, Hung
ST, Sun DP, Uen YH and Lin KY: Ubiquitin-specific protease 3
overexpression promotes gastric carcinogenesis and is predictive of
poor patient prognosis. Cancer Sci. 109:3438–3449. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Chen Y, Pang X, Ji L, Sun Y and Ji Y:
Reduced expression of deubiquitinase USP33 is associated with tumor
progression and poor prognosis of gastric adenocarcinoma. Med Sci
Monit. 24:3496–505. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang X, Yu Q, Huang L and Yu P:
Lentivirus-mediated inhibition of USP39 suppresses the growth of
gastric cancer cells via PARP activation. Mol Med Rep. 14:301–306.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Dong X, Su H, Jiang F, Li H, Shi G and Fan
L: miR-133a, directly targeted USP39, suppresses cell proliferation
and predicts prognosis of gastric cancer. Oncol Lett. 15:8311–3818.
2018.PubMed/NCBI
|
|
75
|
Zhang L, Zhou F, Drabsch Y, Gao R,
Snaar-Jagalska BE, Mickanin C, Huang H, Sheppard KA, Porter JA, Lu
CX and ten Dijke P: USP4 is regulated by AKT phosphorylation and
directly deubiquitylates TGF-β type I receptor. Nat Cell Biol.
14:717–726. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Li Y, Schrodi S, Rowland C, Tacey K,
Catanese J and Grupe A: Genetic evidence for ubiquitin-specific
proteases USP24 and USP40 as candidate genes for late-onset
Parkinson disease. Hum Mutat. 27:1017–1023. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Pinilla-Vera M, Xiong Z, Zhao Y, Zhao J,
Donahoe MP, Barge S, Horne WT, Kolls JK, McVerry BJ, Birukova A, et
al: Full spectrum of LPS activation in alveolar macrophages of
healthy volunteers by whole transcriptomic profiling. PLoS One.
11:e01593292016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Hock AK, Vigneron AM, Carter S, Ludwig RL
and Vousden KH: Regulation of p53 stability and function by the
deubiquitinating enzyme USP42. EMBO J. 30:4921–4930. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
He L, Liu X, Yang J, Li W, Liu S, Liu X,
Yang Z, Ren J, Wang Y, Shan L, et al: Imbalance of the reciprocally
inhibitory loop between the ubiquitin-specific protease USP43 and
EGFR/PI3K/AKT drives breast carcinogenesis. Cell Res. 28:934–951.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Borrero J, Jimenez JJ, Gutiez L, Herranz
C, Cintas LM and Hernandez PE: Use of the usp45 lactococcal
secretion signal sequence to drive the secretion and functional
expression of enterococcal bacteriocins in Lactococcus lactis. Appl
Microbiol Biotechnol. 89:131–143. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Schweitzer K and Naumann M: CSN-associated
USP48 confers stability to nuclear NF-kappaB/RelA by trimming
K48-linked Ub-chains. Biochim Biophys Acta. 1853:453–469. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Weinstock J, Wu J, Cao P, Kingsbury WD,
McDermott JL, Kodrasov MP, McKelvey DM, Suresh Kumar KG, Goldenberg
SJ, Mattern MR and Nicholson B: Selective dual inhibitors of the
cancer-related deubiquitylating proteases USP7 and USP47. ACS Med
Chem Lett. 3:789–792. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zhang B, Yin Y, Hu Y, Zhang J, Bian Z,
Song M, Hua D and Huang Z: MicroRNA-204-5p inhibits gastric cancer
cell proliferation by downregulating USP47 and RAB22A. Med Oncol.
32:3312015. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Naghavi L, Schwalbe M, Ghanem A and
Naumann M: Deubiquitinylase USP47 promotes RelA phosphorylation and
survival in gastric cancer cells. Biomedicines. 6(pii): E622018.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Hou K, Zhu Z, Wang Y, Zhang C, Yu S, Zhu Q
and Yan B: Overexpression and biological function of
ubiquitin-specific protease 42 in gastric cancer. PLoS One.
11:e01529972016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Nishimura S, Oki E, Ando K, Iimori M,
Nakaji Y, Nakashima Y, Saeki H, Oda Y and Maehara Y: High
ubiquitin-specific protease 44 expression induces DNA aneuploidy
and provides independent prognostic information in gastric cancer.
Cancer Med. 6:1453–1464. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Dayal S, Sparks A, Jacob J, Allende-Vega
N, Lane DP and Saville MK: Suppression of the deubiquitinating
enzyme USP5 causes the accumulation of unanchored polyubiquitin and
the activation of p53. J Biol Chem. 284:5030–5041. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Aressy B, Jullien D, Cazales M, Marcellin
M, Bugler B, Burlet-Schiltz O and Ducommun B: A screen for
deubiquitinating enzymes involved in the G2/M checkpoint
identifies USP50 as a regulator of HSP90-dependent Wee1 stability.
Cell Cycle. 9:3815–3822. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Wang Z, Zhang H, Liu J, Cheruiyot A, Lee
JH, Ordog T, Lou Z, You Z and Zhang Z: USP51 deubiquitylates
H2AK13,15ub and regulates DNA damage response. Genes Dev.
30:946–959. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yang S, Liu L, Cao C, Song N, Wang Y, Ma
S, Zhang Q, Yu N, Ding X, Yang F, et al: USP52 acts as a
deubiquitinase and promotes histone chaperone ASF1A stabilization.
Nat Commun. 9:12852018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Kazmierczak M, Harris SL, Kazmierczak P,
Shah P, Starovoytov V, Ohlemiller KK and Schwander M: Progressive
hearing loss in mice carrying a mutation in Usp53. J Neurosci.
35:15582–15598. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Fraile JM, Campos-Iglesias D, Rodriguez F,
Espanol Y and Freije JM: The deubiquitinase USP54 is overexpressed
in colorectal cancer stem cells and promotes intestinal
tumorigenesis. Oncotarget. 7:74427–74434. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Liu Y, Wang WM, Zou LY, Li L, Feng L, Pan
MZ, Lv MY, Cao Y, Wang H, Kung HF, et al: Ubiquitin specific
peptidase 5 mediates Histidine-rich protein Hpn induced cell
apoptosis in hepatocellular carcinoma through P14-P53 signaling.
Proteomics. 172017.doi: 10.1002/pmic.201600350. PubMed/NCBI
|
|
94
|
Oliveira AM, Perez-Atayde AR, Inwards CY,
Medeiros F, Derr V, Hsi BL, Gebhardt MC, Rosenberg AE and Fletcher
JA: USP6 and CDH11 oncogenes identify the neoplastic cell in
primary aneurysmal bone cysts and are absent in so-called secondary
aneurysmal bone cysts. Am J Pathol. 165:1773–1780. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Kang A, Kumar JB, Thomas A and Bourke AG:
A spontaneously resolving breast lesion: Imaging and cytological
findings of nodular fasciitis of the breast with FISH showing USP6
gene rearrangement. BMJ Case Rep. 2015(pii): bcr20152130762015.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Jian F, Cao Y, Bian L and Sun Q: USP8: A
novel therapeutic target for Cushing's disease. Endocrine.
50:292–296. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Fu X, Xie W, Song X, Wu K, Xiao L, Liu Y
and Zhang L: Aberrant expression of deubiquitylating enzyme USP9X
predicts poor prognosis in gastric cancer. Clin Res Hepatol
Gastroenterol. 41:687–692. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Deng S, Zhou H, Xiong R, Lu Y, Yan D, Xing
T, Dong L, Tang E and Yang H: Over-expression of genes and proteins
of ubiquitin specific peptidases (USPs) and proteasome subunits
(PSs) in breast cancer tissue observed by the methods of RFDD-PCR
and proteomics. Breast Cancer Res Treat. 104:21–30. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Xia JT, Chen LZ, Jian WH, Wang KB, Yang
YZ, He WL, Chen D and Li W: MicroRNA-362 induces cell proliferation
and apoptosis resistance in gastric cancer by activation of NF-B
signaling. J Transl Med. 12:332014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Sun B, Li L, Ma W, Wang S and Huang C:
MiR-130b inhibits proliferation and induces apoptosis of gastric
cancer cells via CYLD. Tumour Biol. 37:7981–9787. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Bermejo JL, Kabisch M, Dunnebier T,
Schnaidt S, Melchior F, Fischer HP, Harth V, Rabstein S, Pesch B,
Brüning T, et al: Exploring the association between genetic
variation in the SUMO isopeptidase gene USPL1 and breast cancer
through integration of data from the population-based GENICA study
and external genetic databases. Int J Cancer. 133:362–372. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Mevissen TE, Hospenthal MK, Geurink PP,
Elliott PR, Akutsu M, Arnaudo N, Ekkebus R, Kulathu Y, Wauer T, El
Oualid F, et al: OTU deubiquitinases reveal mechanisms of linkage
specificity and enable ubiquitin chain restriction analysis. Cell.
154:169–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Wiener R, Zhang X, Wang T and Wolberger C:
The mechanism of OTUB1-mediated inhibition of ubiquitination.
Nature. 483:618–622. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Kato K, Nakajima K, Ui A, Muto-Terao Y,
Ogiwara H and Nakada S: Fine-tuning of DNA damage-dependent
ubiquitination by OTUB2 supports the DNA repair pathway choice. Mol
Cell. 53:617–630. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Wang YQ, Zhang QY, Weng WW, Wu Y, Yang YS,
Shen C, Chen XC, Wang L, Liu KJ, Xu MD and Sheng WQ: Upregulation
of the Non-coding RNA OTUB1-isoform 2 contributes to gastric cancer
cell proliferation and invasion and predicts poor gastric cancer
prognosis. Int J Biol Sci. 12:545–557. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Carneiro AP, Reis CF, Morari EC, Maia YC,
Nascimento R, Bonatto JM, de Souza MA, Goulart LR and Ward LS: A
putative OTU domain-containing protein 1 deubiquitinating enzyme is
differentially expressed in thyroid cancer and identifies
less-aggressive tumours. Br J Cancer. 111:551–558. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Flierman D, van der Heden van Noort GJ,
Ekkebus R, Geurink PP, Mevissen TE, Hospenthal MK, Komander D and
Ovaa H: Non-hydrolyzable diubiquitin probes reveal linkage-specific
reactivity of deubiquitylating enzymes mediated by S2 pockets. Cell
Chem Biol. 23:472–482. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Yuan L, Lv Y, Li H, Gao H, Song S, Zhang
Y, Xing G, Kong X, Wang L, Li Y, et al: Deubiquitylase OTUD3
regulates PTEN stability and suppresses tumorigenesis. Nat Cell
Biol. 17:1169–1181. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zhao Y, Majid MC, Soll JM, Brickner JR,
Dango S and Mosammaparast N: Noncanonical regulation of alkylation
damage resistance by the OTUD4 deubiquitinase. EMBO J.
34:1687–1703. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Luo J, Lu Z, Lu X, Chen L, Cao J, Zhang S,
Ling Y and Zhou X: OTUD5 regulates p53 stability by
deubiquitinating p53. PLoS One. 8:e776822013. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Kim SY, Kwon SK, Lee SY and Baek KH:
Ubiquitin-specific peptidase 5 and ovarian tumor deubiquitinase 6A
are differentially expressed in p53+/+ and
p53−/− HCT116 cells. Int J Oncol. Mar 5–2018.(Epub ahead
of print). doi: 10.3892/ijo.2018.4302. View Article : Google Scholar
|
|
112
|
Santiago-Sim T, Burrage LC, Ebstein F,
Tokita MJ, Miller M, Bi W, Braxton AA, Rosenfeld JA, Shahrour M,
Lehmann A, et al: Biallelic variants in OTUD6B cause an
intellectual disability syndrome associated with seizures and
dysmorphic features. Am J Hum Genet. 100:676–688. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Evans PC, Smith TS, Lai MJ, Williams MG,
Burke DF, Heyninck K, Kreike MM, Beyaert R, Blundell TL and Kilshaw
PJ: A novel type of deubiquitinating enzyme. J Biol Chem.
278:23180–23186. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Xu Z, Pei L, Wang L, Zhang F, Hu X and Gui
Y: Snail1-dependent transcriptional repression of Cezanne2 in
hepatocellular carcinoma. Oncogene. 33:2836–2845. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Virdee S, Ye Y, Nguyen DP, Komander D and
Chin JW: Engineered diubiquitin synthesis reveals Lys29-isopeptide
specificity of an OTU deubiquitinase. Nat Chem Biol. 6:750–757.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Guo T, Zhang Y, Qu X, Che X, Li C, Fan Y,
Wan X, Ma R, Hou K, Zhou H, et al: miR-200a enhances TRAIL-induced
apoptosis in gastric cancer cells by targeting A20. Cell Biol Int.
42:506–514. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Lork M, Verhelst K and Beyaert R: CYLD,
A20 and OTULIN deubiquitinases in NF-B signaling and cell death: So
similar, yet so different. Cell Death Differ. 24:1172–1183. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Weng W, Zhang Q, Xu M, Wu Y, Zhang M, Shen
C, Chen X, Wang Y and Sheng W: OTUB1 promotes tumor invasion and
predicts a poor prognosis in gastric adenocarcinoma. Am J Transl
Res. 8:2234–2244. 2016.PubMed/NCBI
|
|
119
|
Wang X, Zhang L, Zhang Y, Zhao P, Qian L,
Yuan Y, Liu J, Cheng Q, Xu W, Zuo Y, et al: JOSD1 negatively
regulates type-I interferon antiviral activity by deubiquitinating
and stabilizing SOCS1. Viral Immunol. 30:342–349. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Zhang B, Zheng A, Hydbring P, Ambroise G,
Ouchida AT, Goiny M, Vakifahmetoglu-Norberg H and Norberg E: PHGDH
defines a metabolic subtype in lung adenocarcinomas with poor
prognosis. Cell Rep. 19:2289–2303. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Zhang J, Huang JY, Chen YN, Yuan F, Zhang
H, Yan FH, Wang MJ, Wang G, Su M, Lu G, et al: Whole genome and
transcriptome sequencing of matched primary and peritoneal
metastatic gastric carcinoma. Sci Rep. 5:137502015. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Butler LR, Densham RM, Jia J, Garvin AJ,
Stone HR, Shah V, Weekes D, Festy F, Beesley J and Morris JR: The
proteasomal de-ubiquitinating enzyme POH1 promotes the
double-strand DNA break response. EMBO J. 31:3918–3934. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Py BF, Kim MS, Vakifahmetoglu-Norberg H
and Yuan J: Deubiquitination of NLRP3 by BRCC3 critically regulates
inflammasome activity. Mol Cell. 49:331–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Sun H, Guo D, Su Y, Yu D, Wang Q, Wang T,
Zhou Q, Ran X and Zou Z: Hyperplasia of pericytes is one of the
main characteristics of microvascular architecture in malignant
glioma. PLoS One. 9:e1142462014. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zhou L, Shi L, Guo H and Yao X: MYSM-1
suppresses migration and invasion in renal carcinoma through
inhibiting epithelial-mesenchymal transition. Tumour Biol. Sep
27–2015.(Epub ahead of print).
|
|
126
|
Xiao D, Yang S, Huang L, He H, Pan H and
He J: COP9 signalosome subunit CSN5, but not CSN6, is upregulated
in lung adenocarcinoma and predicts poor prognosis. J Thorac Dis.
10:1596–1606. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Niu Z, Lei R, Shi J, Wang D, Shou W, Wang
Z, Wang Y, Wang Z and Huang W: A polymorphism rs17336700 in the
PSMD7 gene is associated with ankylosing spondylitis in Chinese
subjects. Ann Rheum Dis. 70:706–907. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
McCullough J, Clague MJ and Urbe S: AMSH
is an endosome-associated ubiquitin isopeptidase. J Cell Biol.
166:487–492. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Zhu W, Liu Y and Ling B: Quantum mechanics
and molecular mechanics study of the catalytic mechanism of human
AMSH-LP domain deubiquitinating enzymes. Biochemistry.
54:5225–5234. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Wickramasinghe VO, Gonzalez-Porta M,
Perera D, Bartolozzi AR, Sibley CR, Hallegger M, Ule J, Marioni JC
and Venkitaraman AR: Regulation of constitutive and alternative
mRNA splicing across the human transcriptome by PRPF8 is determined
by 5′ splice site strength. Genome Biol. 16:2012015. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Sang MM, Du WQ, Zhang RY, Zheng JN and Pei
DS: Suppression of CSN5 promotes the apoptosis of gastric cancer
cells through regulating p53-related apoptotic pathways. Bioorg Med
Chem Lett. 25:2897–2901. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Wang X, Wang H, Zhao S, Sun P, Wen D, Liu
T, Liu H, Yang Z and Ma Z: Eukaryotic translation initiation factor
EIF3H potentiates gastric carcinoma cell proliferation. Tissue
Cell. 53:23–29. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Cheng Y, Jia C, Li G and Li H: Expression
of eukaryotic initiation factor 3f is associated with prognosis in
gastric carcinomas. Oncol Res Treat. 37:198–202. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Tahara H, Kay MA, Yasui W and Tahara E:
MicroRNAs in cancer: The 22nd hiroshima cancer Seminar/the 4th
Japanese Association for RNA interference joint international
symposium, 30 August 2012, grand prince hotel Hiroshima. Jpn J Clin
Oncol. 43:579–582. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Huang S, Liu S, Fu JJ, Tony Wang T, Yao X,
Kumar A, Liu G and Fu M: Monocyte chemotactic protein-induced
protein 1 and 4 form a complex but act independently in regulation
of interleukin-6 mRNA degradation. J Biol Chem. 290:20782–20792.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Roy A and Kolattukudy PE: Monocyte
chemotactic protein-induced protein (MCPIP) promotes inflammatory
angiogenesis via sequential induction of oxidative stress,
endoplasmic reticulum stress and autophagy. Cell Signal.
24:2123–2131. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Mansour MA: Ubiquitination: Friend and foe
in cancer. Int J Biochem Cell Biol. 101:80–93. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Suk FM, Chang CC, Lin RJ, Lin SY, Chen YT
and Liang YC: MCPIP3 as a potential metastasis suppressor gene in
human colorectal cancer. Int J Mol Sci. 19:E13502018. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|