Introduction
The incidence and mortality of lung cancer rank the
highest among all types of cancer worldwide. In 2018, lung cancer
was the most commonly diagnosed cancer (11.6% of all cancer cases)
and the leading cause of cancer-associated mortality (18.4% of all
cancer-associated mortality cases) across 20 world regions
(1). Malignant epithelial tumors are
the most frequently observed in lung cancer, and can be grouped
into non-small cell lung carcinoma (NSCLC) and small cell lung
carcinoma (2). NSCLC accounts for
85–90% of lung cancer cases, and lung adenocarcinoma (LUAD) is a
common type of NSCLC (3). Although
positive outcomes have been achieved following early diagnosis, the
recurrence rate remains unacceptably high, and the 5-year overall
survival rate of patients with LUAD remains low (4). Without sufficient early detection
methods and effective therapeutic strategies during the early tumor
stages, the mortality rate of patients with LUAD has not markedly
decreased in recent years (5).
Therefore, further insight into the mechanisms responsible for the
development and progression of LUAD is urgently required (6).
Due to the development of high-throughput microarray
technology, an increasing number of genes have been identified to
serve an important role in tumor occurrence and in the progression
of LUAD (7). Gene expression
profiles were used to identify important genes associated with
tumor progression (8). However, the
majority of studies have focused on differentially expressed genes
(DEGs) and not on the interconnection between genes (9–11). In
order to obtain further information on the association between gene
expression levels and important clinical features, scale-free gene
co-expression networks were constructed using co-expression
analysis. Previous studies have applied weighted gene co-expression
network analysis (WGCNA) to analyze gene expression datasets and
screen hub genes (12,13). Tumor stage is crucial to the clinical
prognosis of patients with LUAD, and the survival status of
patients at different tumor stages differs significantly (14). Therefore, tumor stage was selected as
a main clinical feature. Subsequently, co-expression networks of
the association between genes were constructed, and network-centric
genes associated with the clinical features were identified.
Finally, GSE40791 and UALCAN were applied to investigate the value
of the candidate hub genes.
Materials and methods
Data sources and processing
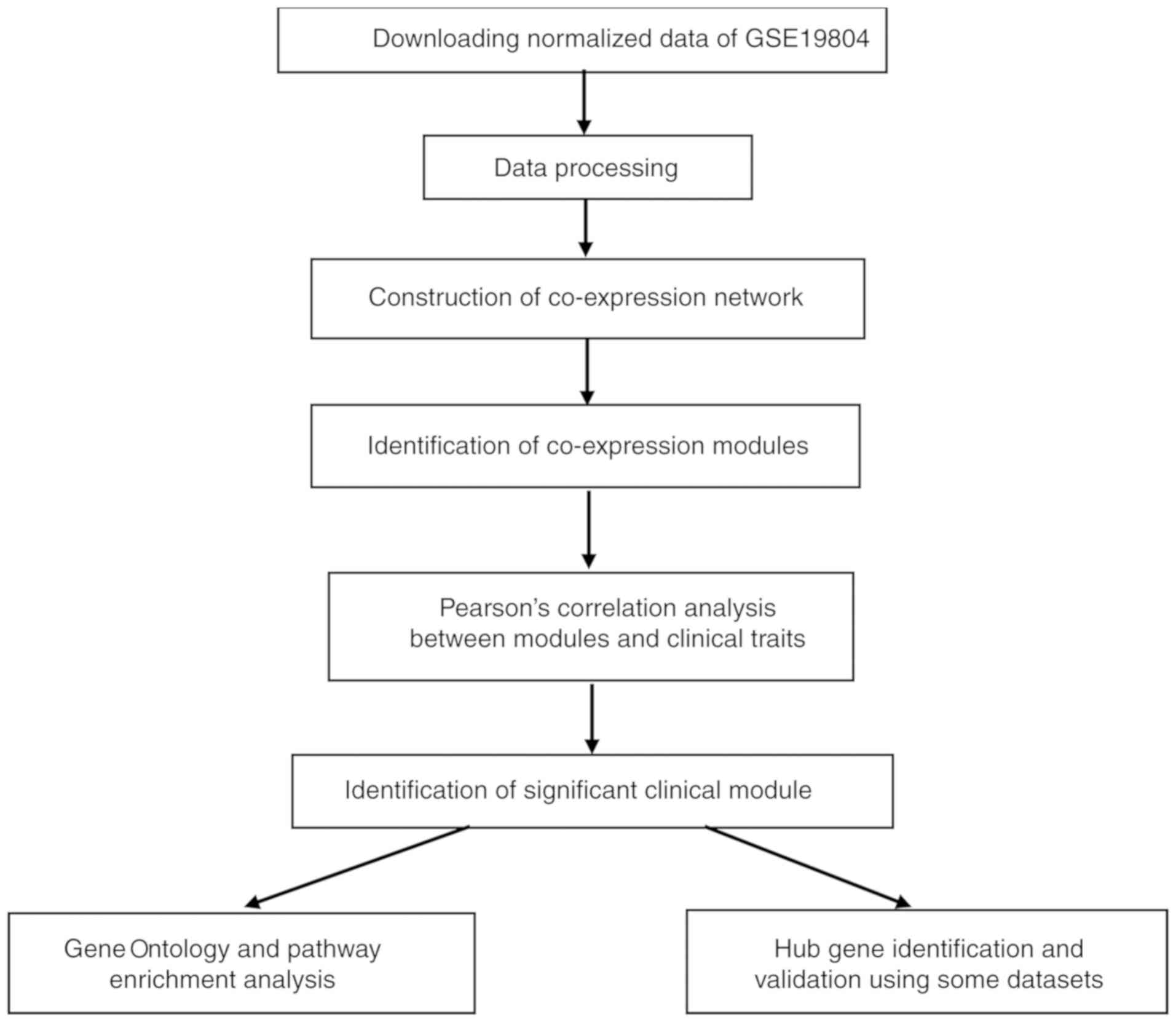
The brief study flow is presented in Fig. 1. The gene expression profile GSE19804
dataset associated with LUAD was downloaded from the Gene
Expression Omnibus database (http://www.ncbi.nlm.nih.gov/geo/). GSE19804, which was
based on the GPL96 platform (Affymetrix Human Genome U133A Array),
contains 120 samples (60 normal and 60 LUAD samples) and 54,675
genes (15). The dataset was
normalized with quantile normalization by the R package ‘affy’
(16). The top 25% most variant
genes (13,669 genes) were then selected by analysis of variance for
further study in R 3.5.1.
Co-expression network
construction
The R (R 3.5.1; http://www.r-project.org/) package ‘WGCNA’ (7) was used to construct gene co-expression
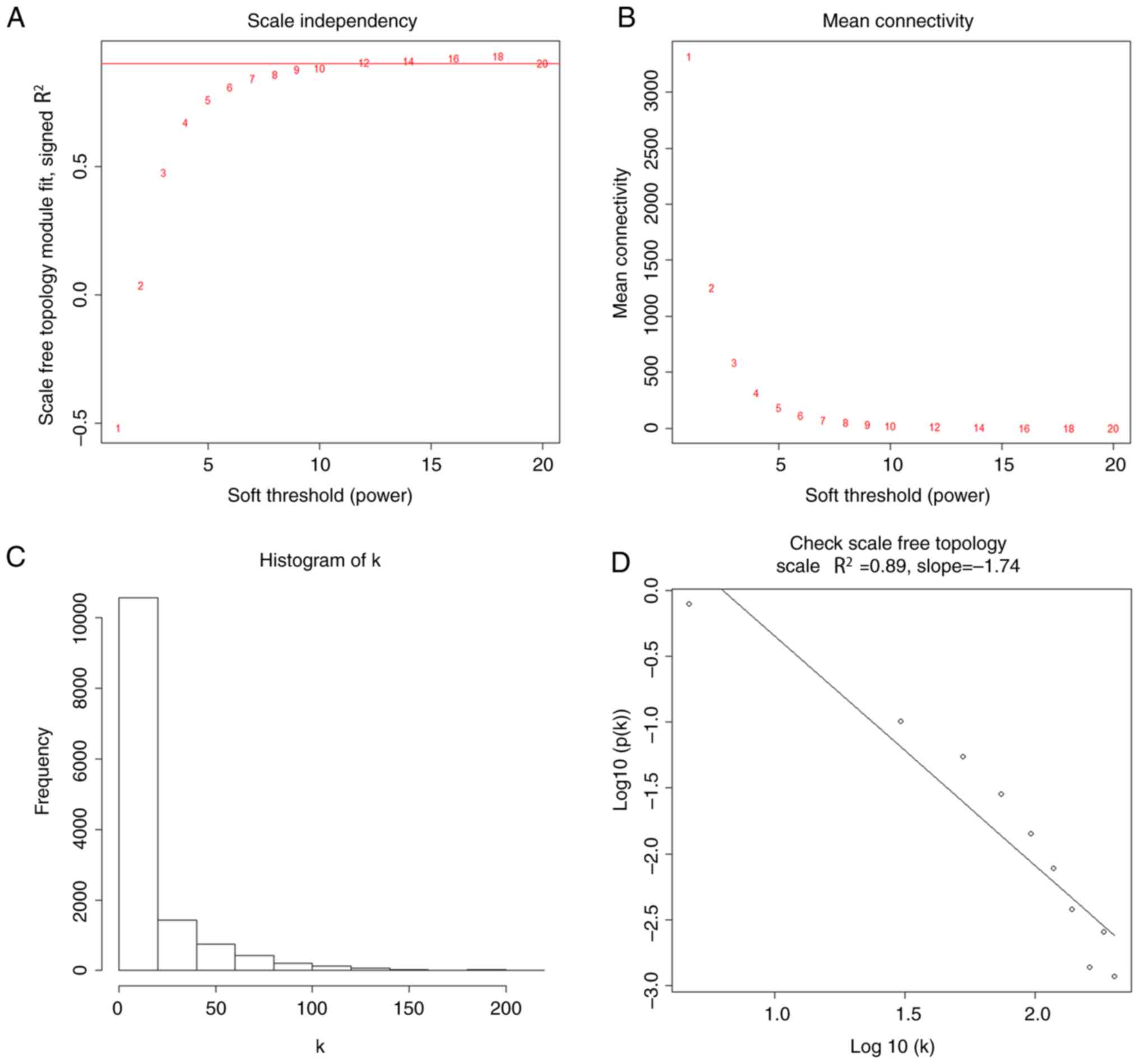
networks for the filtered gene expression matrix. To construct a
scale-free network, the power of β=12 (scale-free
R2=0.89) was selected as the soft-thresholding
parameter. After transforming the adjacency into a topological
overlap measurement (TOM), the corresponding dissimilarity (1-TOM)
was calculated and the dissimilarity of module eigengenes (MEs) was
estimated. Using the DynamicTreeCut algorithm (17), the genes, which had similar
expression profiles, were categorized into the same module.
Identification of clinically
significant modules
The clinical trait of focus was the T stage of LUAD.
The association between the clinical phenotype and MEs was
determined to identify clinically significant modules. MEs were
deemed to represent the expression levels of all genes in the
associated module. In addition, the mediated P-value of each gene
was calculated and the gene significance (GS=lg P) was identified.
Finally, the most clinically significant module was selected
according to module significance, which was the average GS of genes
involved in the associated module.
Functional and pathway enrichment
analysis
The Database for Annotation, Visualization and
Integrated Discovery 5 (https://david-d.ncifcrf.gov/) (DAVID) is a database
for several types of functional annotation. With the aid of DAVID,
the biological relevance of the genes in a given module was
identified according to false discovery rate (FDR) <0.05. Gene
Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG)
enrichment analyses of the genes in the hub module were performed
by DAVID.
Identification and validation of hub
genes
The connectivity of the module can be measured by
the absolute value of the Pearson's correlation. Additionally, the
association between clinical traits and genes can be measured by
the absolute value of the Pearson's correlation. The genes that had
a high connectivity with the module and selected phenotype were
regarded to be hub genes in hub module (cor.geneModuleMembership
>0.8 and cor.geneTraitSignificance >0.2) (18). Survival analysis was performed to
explore the association between the expression level of hub genes
and overall survival rate in lung adenocarcinoma (based on The
Cancer Genome Atlas data in Gene Expression Profiling Interactive
Analysis, http://gepia.cancer-pku.cn/).
Furthermore, other data of LUAD from GSE40791 (19) and UALCAN (http://ualcan.path.uab.edu/) were used for validation.
GSE40791 was used to identify DEGs between normal tissues and LUAD
tissues by using the ‘limma’ package in R (20). There are 100 normal samples and 94
tumor samples in the GSE40791 dataset. To overlap the genes in the
turquoise module and DEGs, a Venn diagram was constructed using the
online tool jvenn (http://jvenn.toulouse.inra.fr/app/example.html).
UALCAN is a useful online tool for analyzing cancer transcriptome
data, which is based on public cancer transcriptome data from The
Cancer Genome Atlas (https://portal.gdc.cancer.gov) (TCGA) and MET500
(21) transcriptome sequencing
(22). Independent-sample t-test was
used to validate the hub genes in UALCAN. Validation of the genes
that were selected from protein levels using The Human Protein
Atlas (http://www.proteinatlas.org) was also
performed.
Results
Weighted co-expression network
construction and key module identification
Using the method of average linkage hierarchical
clustering, 120 samples from the GSE19804 dataset were clustered
(Fig. 2). Using the ‘WGCNA’ package,
genes with similar expression levels were divided into modules to
construct co-expression networks. The power of β=12 (scale free
R2=0.89) was selected as the soft-thresholding parameter
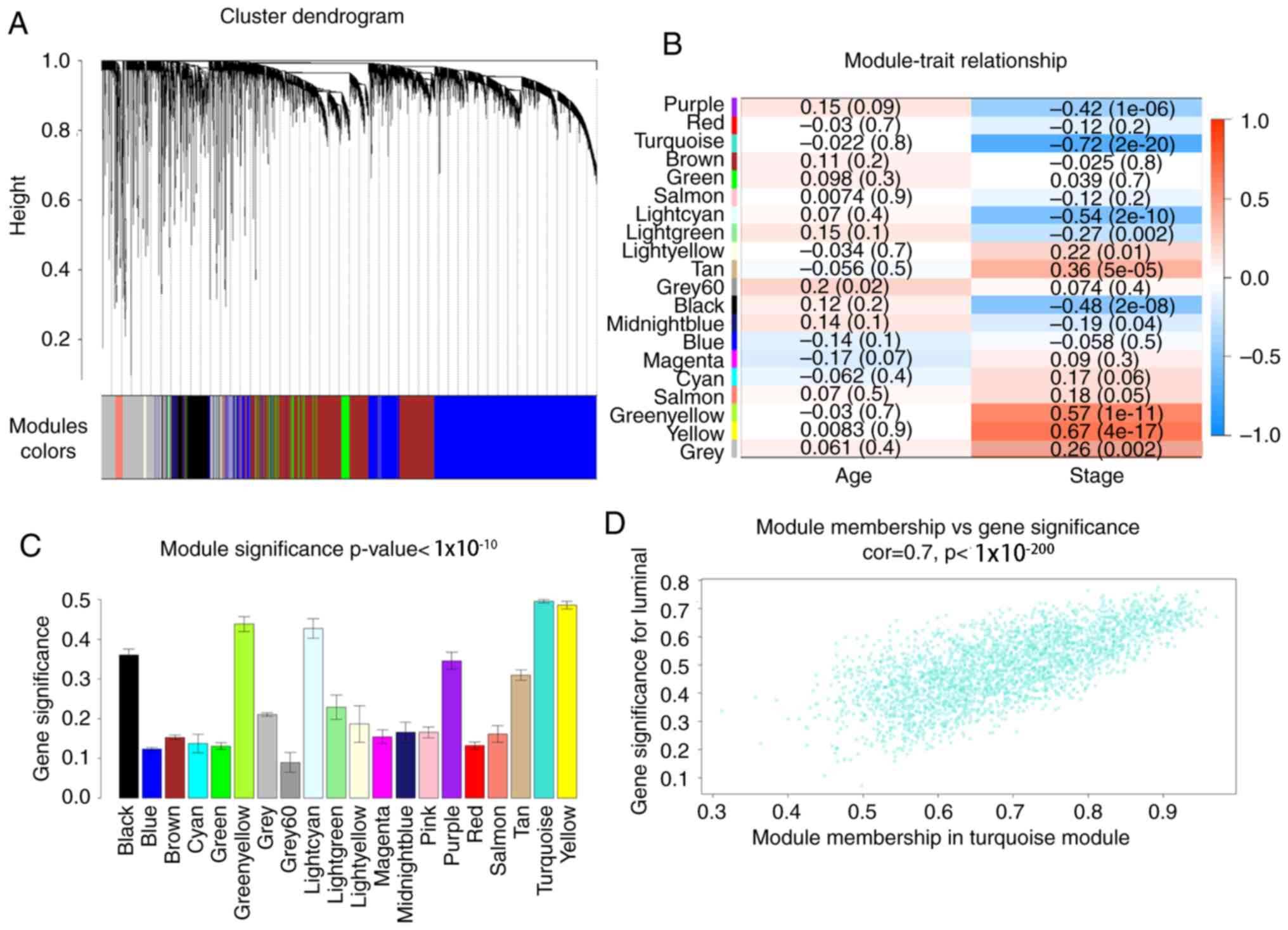
(Fig. 3). In total, 20 modules were
identified and the turquoise module exhibited the highest
association with the T stage of LUAD (Fig. 4). Therefore, the turquoise module was
selected for further analysis as the clinically significant
module.
GO and pathway enrichment
analysis
To obtain further information on the function of
candidate genes, the genes from the turquoise module were
categorized into biological process (BP), cellular component (CC)
and molecular function (MF) terms. The outcome of GO enrichment
analysis is presented in Table I.
The BP terms were generally enriched in the ‘positive regulation of
transcription from RNA polymerase II promoter’, ‘signal
transduction’, ‘negative regulation of transcription from RNA
polymerase II promoter’, ‘cell adhesion’ and ‘positive regulation
of GTPase activity’; the CC terms were mainly focused on
‘cytoplasm’, ‘plasma membrane’, ‘extracellular exosome’,
‘extracellular region’ and ‘integral component of plasma membrane’;
and the MF terms were focused on ‘protein binding’, ‘calcium ion
binding’, ‘actin binding’, ‘transcriptional activator activity, RNA
polymerase II core promoter proximal region sequence-specific
binding’ and ‘heparin binding’. In addition, KEGG analysis was
performed to obtain the pathways in the turquoise module. The
results presented in Table II
indicated that these genes were included in ‘focal adhesion’,
‘cGMP-PKG signaling pathway’ and ‘tight junction’. Overall, the
genes in the turquoise module were primarily associated with signal
transduction.
 | Table I.GO enrichment analysis of the
turquoise module. |
Table I.
GO enrichment analysis of the
turquoise module.
| Category | Term | Count, n | % | FDR |
|---|
| BP | GO:0045944-positive
regulation of transcription from RNA polymerase II promoter | 157 | 8.26751 |
3.54×10−7 |
| BP | GO:0007165-signal
transduction | 157 | 8.26751 |
2.14×10−2 |
| BP | GO:0000122-negative
regulation of transcription from RNA polymerase II promoter | 114 | 6.00316 |
2.66×10−4 |
| BP | GO:0007155-cell
adhesion | 98 | 5.16061 |
1.54×10−10 |
| BP | GO:0043547-positive
regulation of GTPase activity | 96 | 5.05529 |
1.08×10−4 |
| CC |
GO:0005737-cytoplasm | 577 | 30.38441 |
1.68×10−2 |
| CC | GO:0005886-plasma
membrane | 531 | 27.96209 |
1.33×10−12 |
| CC |
GO:0070062-extracellular exosome | 367 | 19.32596 |
5.00×10−8 |
| CC |
GO:0005576-extracellular region | 213 | 11.21643 |
6.62×10−4 |
| CC | GO:0005887-ntegral
component of plasma membrane | 206 | 10.84781 |
5.04×10−7 |
| MF | GO:0005515-protein
binding | 953 | 50.18431 |
8.86×10−7 |
| MF | GO:0005509-calcium
ion binding | 112 | 5.89784 |
2.45×10−4 |
| MF | GO:0003779-actin
binding | 54 | 2.84360 |
1.10×10−3 |
| MF |
GO:0001077-transcriptional activator
activity, RNA polymerase II core promoter proximal region
sequence-specific binding | 45 | 2.36967 |
1.72×10−2 |
| MF | GO:0008201-heparin
binding | 36 | 1.895735 |
3.57×10−3 |
| Table II.KEGG pathway enrichment analysis of
the turquoise module. |
Table II.
KEGG pathway enrichment analysis of
the turquoise module.
| Category | Term | Count, n | Percentage | FDR |
|---|
| KEGG | hsa04510: focal
adhesion | 47 | 2.474987 | 0.001208 |
| KEGG | hsa04022: cGMP-PKG
signaling pathway | 37 | 1.948394 | 0.036805 |
| KEGG | hsa04530: tight
junction | 34 | 1.790416 | 0.007826 |
Identification and validation of hub
genes
Given the threshold of |module membership
(MM)|>0.8 and |GS|>0.2, a total of 415 genes in the turquoise
module were recognized as hub genes. Additionally, carbonic
anhydrase 4 (CA4), platelet and endothelial cell adhesion molecule
1 (PECAM1), DnaJ heat shock protein family (Hsp40) member B4
(DNAJB4), advanced glycosylation end-product specific receptor
(AGER), GTPase, IMAP family member 6 (GIMAP6), chromosome 10 open
reading frame 54 (C10orf54), dedicator of cytokinesis 4 (DOCK4),
Golgi membrane protein 1 (GOLM1) and platelet activating factor
acetylhydrolase 1b catalytic subunit 3 (PAFAH1B3) were associated
with overall survival and relapse-free survival (Figs. 5 and 6). After clarifying the hub genes, some
methods were used for validation. Firstly, the ‘limma’ package in R
was used to identify DEGs between normal tissues and LUAD tissues
in the GSE40791 dataset. Defined by the threshold of
|log2 fold change|≥2 and FDR ≤0.05, 3,295 DEGs were
obtained. To overlap the genes in the turquoise module and DEGs, a
Venn diagram was constructed using the online tool jvenn (Fig. 7). Secondly, the expression levels of
these nine genes differed between the normal and LUAD samples in
UALCAN (Fig. 8). CA4, PECAM1,
DNAJB4, AGER, GIMAP6, C10orf54 and DOCK4 were expressed at lower
levels in the tumor samples, whereas GOLM1 and PAFAH1B3 were highly
expressed in the tumor samples. Finally, in the Human Protein Atlas
database, the protein expression levels of six genes (PECAM1,
DNAJB4, AGER, GIMAP6, GOLM1 and PAFAH1B3) in the LUAD samples were
distinct from the normal samples (Fig.
9). There were no associated IHC samples of CA4, C10orf54 and
DOCK4 in the database.
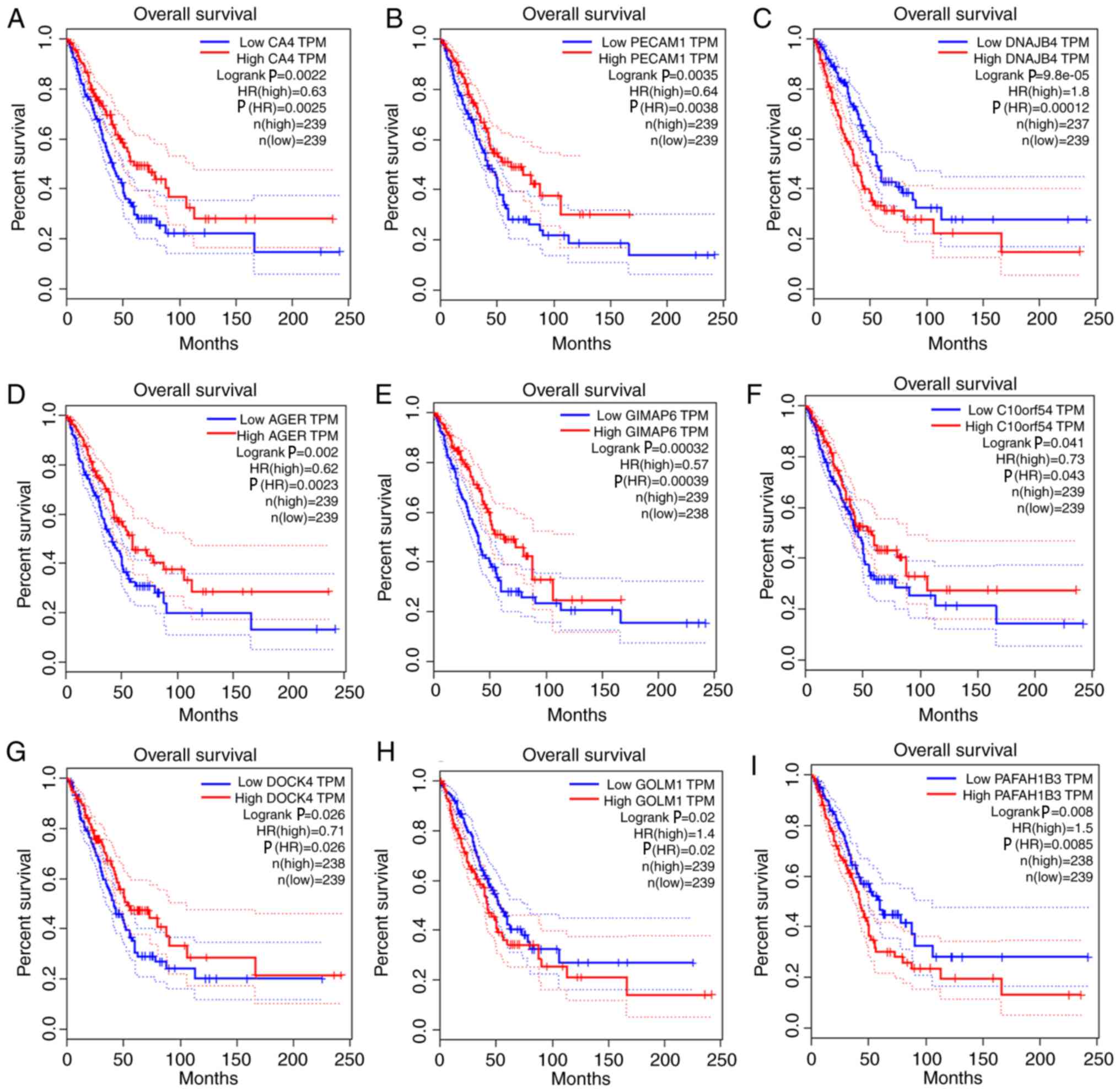 | Figure 5.Survival analysis of the association
between the expression levels of hub genes and overall survival
rate in lung adenocarcinoma (based on The Cancer Genome Atlas data
in Gene Expression Profiling Interactive Analysis). (A) CA4. (B)
PECAM1. (C) DNAJB4. (D) AGER. (E) GIMAP6. (F) C10orf54. (G) DOCK4.
(H) GOLM1. (I) PAFAH1B3. The red line indicates the samples with a
high gene expression, and the blue line indicates the samples with
low gene expression. P≤0.05 was regarded as statistically
significant. CA4, carbonic anhydrase 4; PECAM1, platelet and
endothelial cell adhesion molecule 1; DNAJB4, DnaJ member B4; AGER,
advanced glycosylation end-product specific receptor; GIMAP6,
GTPase, IMAP family member 6; C10orf54, chromosome 10 open reading
frame 54; DOCK4, dedicator of cytokinesis 4; GOLM1, Golgi membrane
protein 1; PAFAH1B3, platelet activating factor acetylhydrolase 1b
catalytic subunit 3; HR, hazard ratio; TPM, transcripts per
million. |
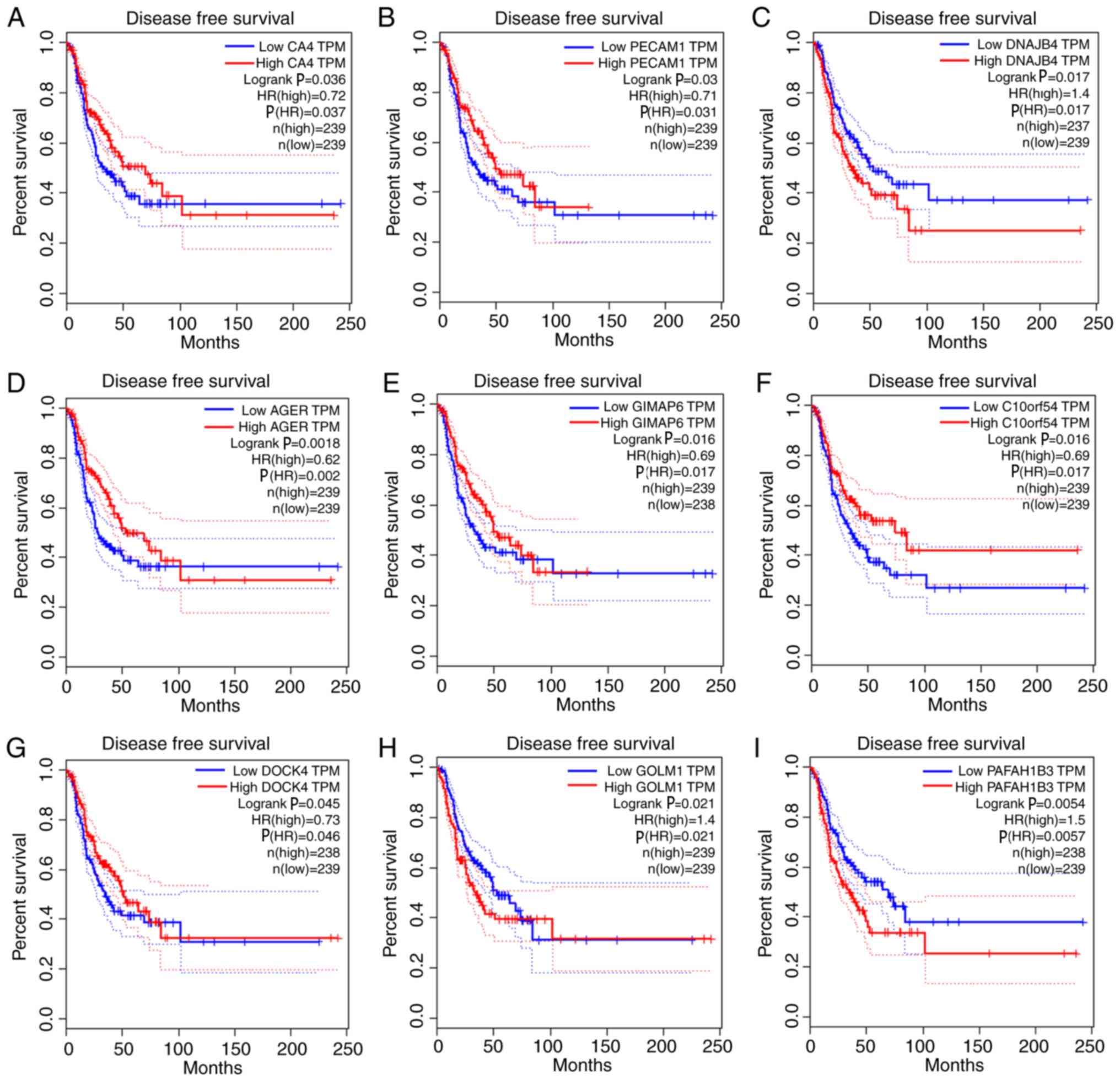 | Figure 6.Survival analysis of the association
between the expression levels of hub genes and disease-free
survival time in lung adenocarcinoma (based on The Cancer Genome
Atlas data in Gene Expression Profiling Interactive Analysis). (A)
CA4. (B) PECAM1. (C) DNAJB4. (D) AGER. (E) GIMAP6. (F) C10orf54.
(G) DOCK4. (H) GOLM1. (I) PAFAH1B3. Red line indicates the samples
with highly expressed genes, and blue line shows the samples with
lowly expressed gene. P≤0.05 was regarded as statistically
significant. HR, hazard ratio; CA4, carbonic anhydrase 4; PECAM1,
platelet and endothelial cell adhesion molecule 1; DNAJB4, DnaJ
member B4; AGER, advanced glycosylation end-product specific
receptor; GIMAP6, GTPase, IMAP family member 6; C10orf54,
chromosome 10 open reading frame 54; DOCK4, dedicator of
cytokinesis 4; GOLM1, Golgi membrane protein 1; PAFAH1B3, platelet
activating factor acetylhydrolase 1b catalytic subunit 3; TPM,
transcripts per million. |
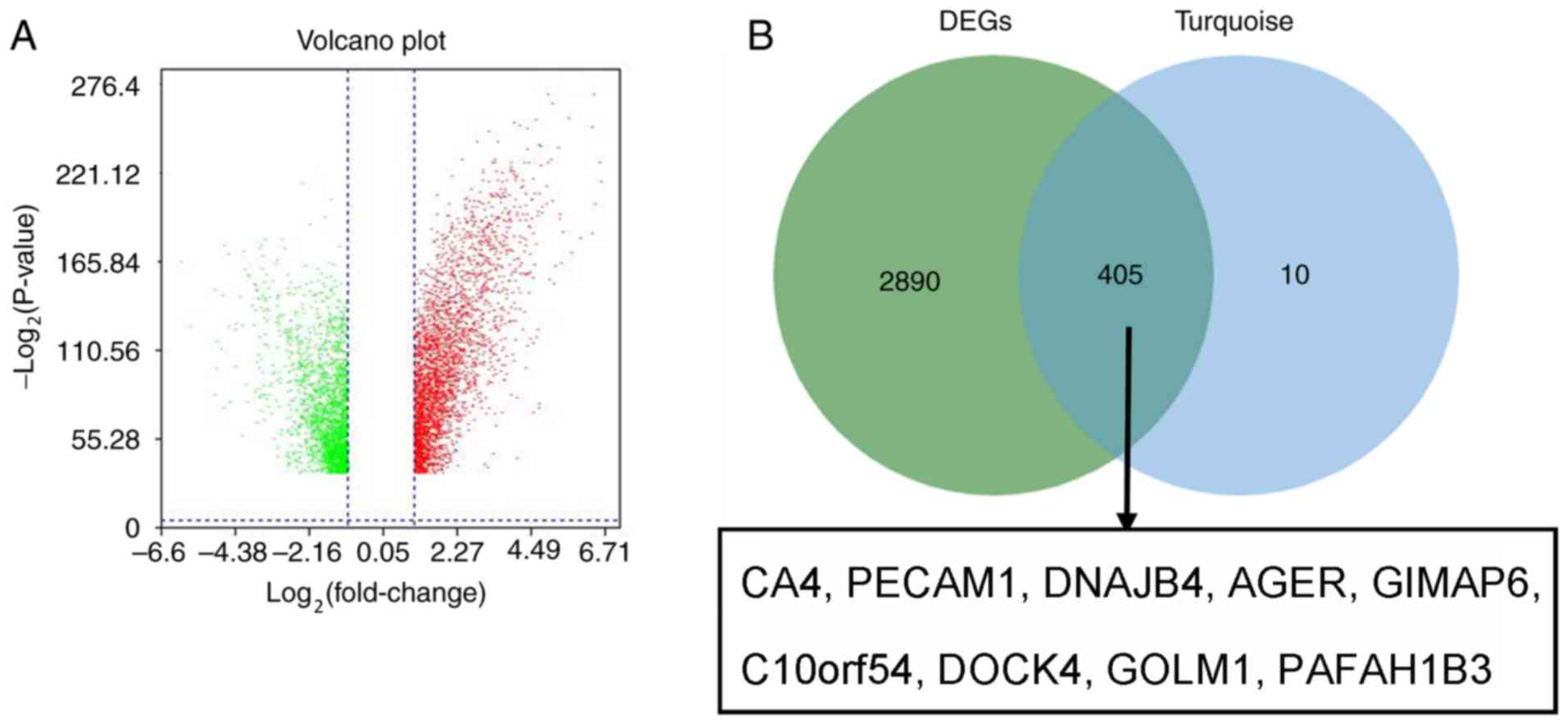 | Figure 7.Validation of hub genes in GSE40791.
(A) Volcano plot visualizing DEGs in GSE40791 (100 normal samples
and 94 lung adenocarcinoma samples). The vertical lines demarcate
the fold change values. The right vertical line corresponds to
≥2-fold change (upregulation) and the left vertical line to ≥2-fold
change (downregulation), whereas the horizontal line marks a
-log10 adjusted P-value of 0.01. (B) Identification of
common genes between DEGs and the turquoise module by overlapping
them. The nine hub genes in the turquoise module were also DEGs in
the GSE40791 dataset. DEGs, differentially expressed genes; CA4,
carbonic anhydrase 4; PECAM1, platelet and endothelial cell
adhesion molecule 1; DNAJB4, DnaJ member B4; AGER, advanced
glycosylation end-product specific receptor; GIMAP6, GTPase, IMAP
family member 6; C10orf54, chromosome 10 open reading frame 54;
DOCK4, dedicator of cytokinesis 4; GOLM1, Golgi membrane protein 1;
PAFAH1B3, platelet activating factor acetylhydrolase 1b catalytic
subunit 3. |
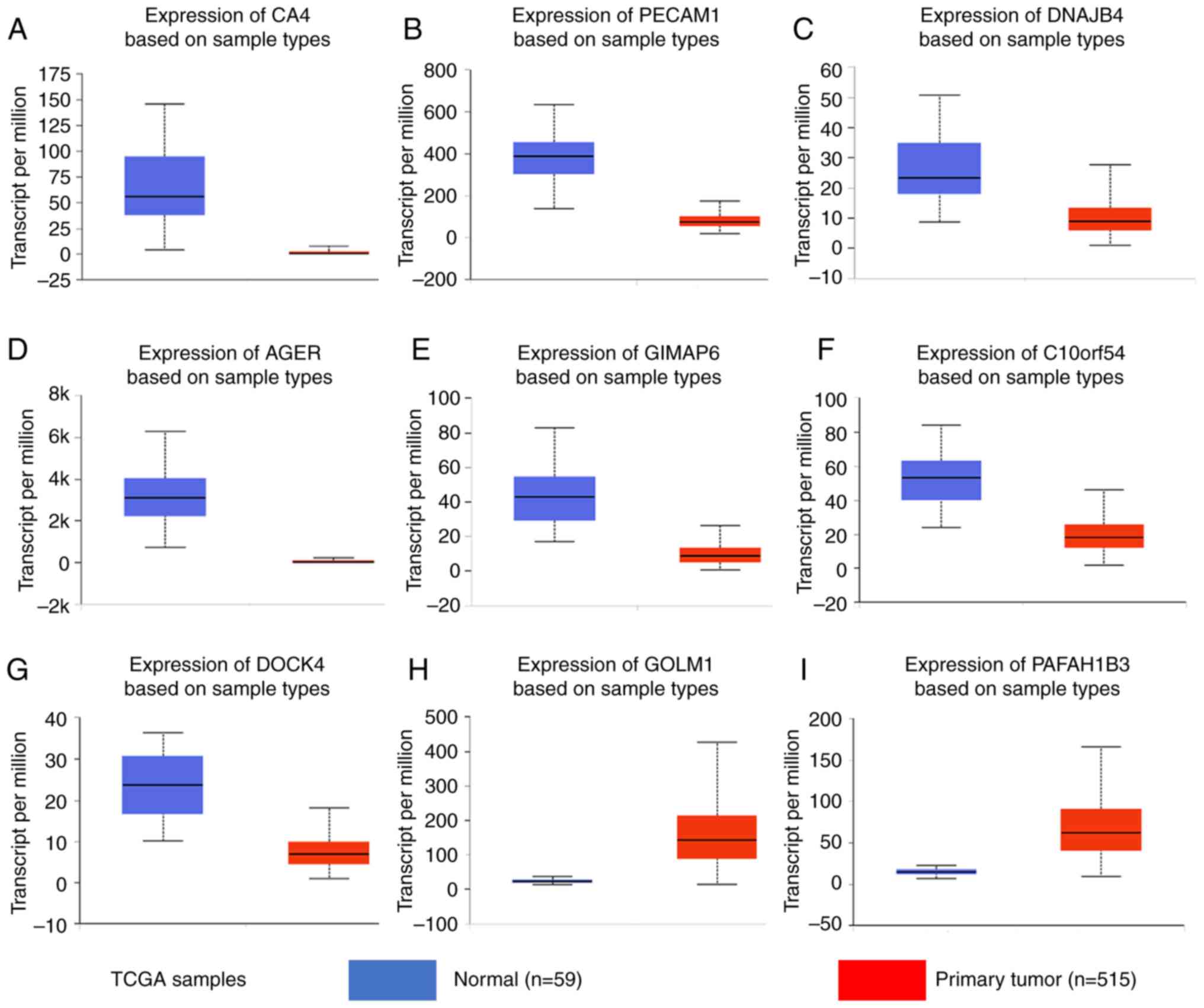 | Figure 8.Gene expression levels in normal lung
and tumor samples (based on The Cancer Genome Atlas data in
UALCAN). mRNA levels of (A) CA4, (B) PECAM1, (C) DNAJB4, (D) AGER,
(E) GIMAP6, (F) C10orf54, (G) DOCK4, (H) GOLM1 and (I) PAFAH1B3.
(A-I) P<0.0001. TCGA, The Cancer Genome Atlas; CA4, carbonic
anhydrase 4; PECAM1, platelet and endothelial cell adhesion
molecule 1; DNAJB4, DnaJ member B4; AGER, advanced glycosylation
end-product specific receptor; GIMAP6, GTPase, IMAP family member
6; C10orf54, chromosome 10 open reading frame 54; DOCK4, dedicator
of cytokinesis 4; GOLM1, Golgi membrane protein 1; PAFAH1B3,
platelet activating factor acetylhydrolase 1b catalytic subunit
3. |
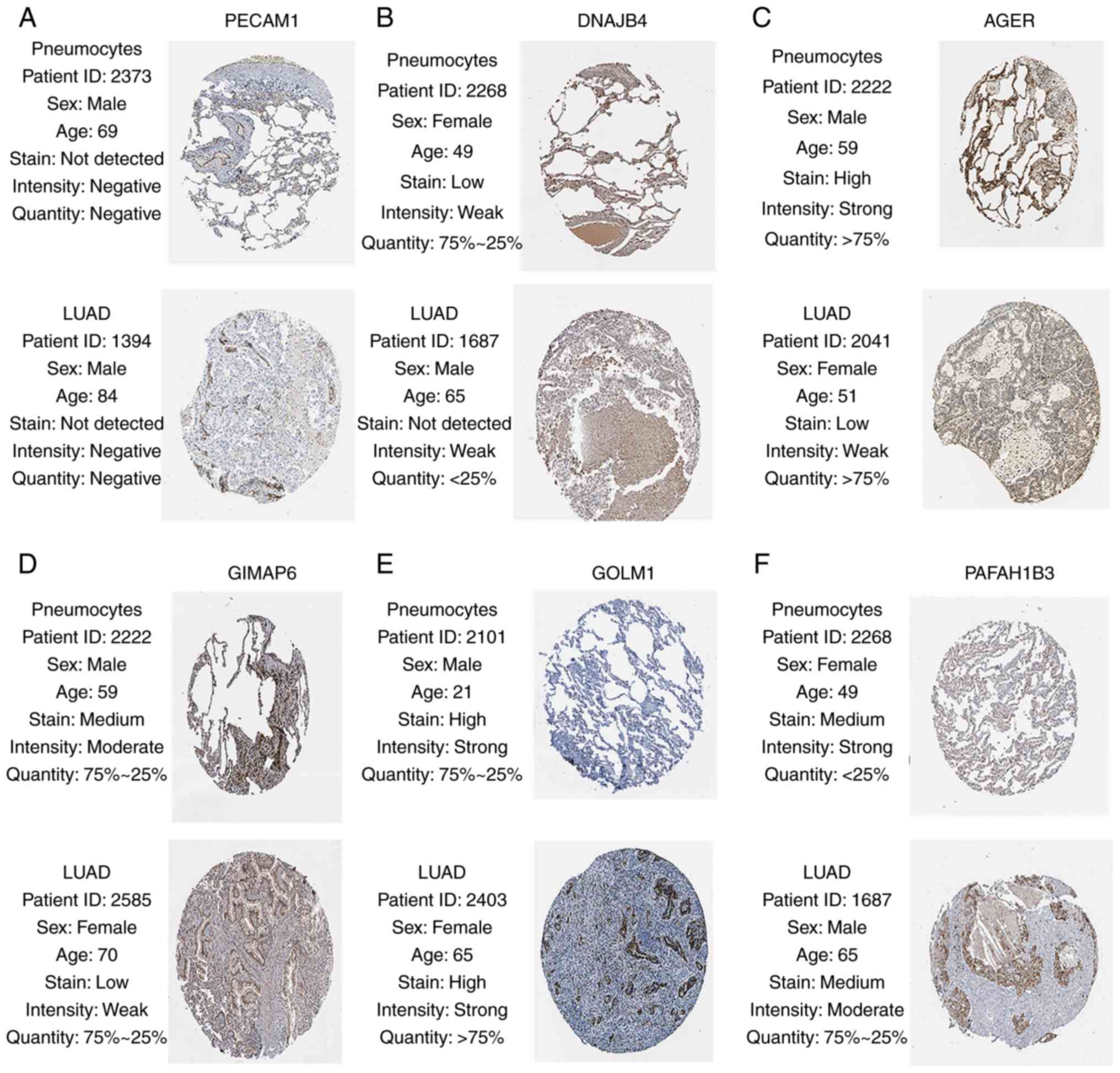 | Figure 9.Validation of six hub genes in the
turquoise module using The Human Protein Atlas database. There were
no associated immunohistochemistry samples of carbonic anhydrase 4,
chromosome 10 open reading frame 54 and dedicator of cytokinesis 4
in the database. Expression of (A) PECAM1, (B) DNAJB4, (C) AGER,
(D) GIMAP6, (E) GOLM1, (F) PAFAH1B3. (A-F) Translational expression
levels of six hub genes were positively associated with the disease
status, as they were upregulated in the LUAD samples. PECAM1,
platelet and endothelial cell adhesion molecule 1; DNAJB4, DnaJ
member B4; AGER, advanced glycosylation end-product specific
receptor; GIMAP6, GTPase, IMAP family member 6; GOLM1, Golgi
membrane protein 1; PAFAH1B3, platelet activating factor
acetylhydrolase 1b catalytic subunit 3; LUAD, lung
adenocarcinoma. |
Discussion
The early diagnosis and recurrence prediction of
LUAD are crucial for effective prevention and treatment. Therefore,
a deeper understanding of the molecular mechanisms associated with
the development of these tumors is of utmost importance. In the
present study, free-scale gene co-expression networks were
constructed to identify genes with a high connectivity with the T
stage of LUAD. WGCNA is a method of analyzing the association
between the expression levels of genes and important clinical
features (12). Although
co-expression does not mean causality, the module and the genes
that are closely associated with a certain clinical phenotype in
co-expression networks could be identified. Additionally, survival
analysis was performed to screen hub genes, which were associated
with overall survival and relapse-free survival rate. The hub genes
(CA4, PECAM1, DNAJB4, AGER, GIMAP6, C10orf54, DOCK4, GOLM1 and
PAFAH1B3) were differentially expressed between the normal and LUAD
samples at the transcriptional and protein level, and were
significantly associated with prognosis. Therefore, the hub genes
serving an important role in tumor progression have the potential
to be prognostic biomarkers for LUAD.
In the present study, the tumor stage of the
patients with LUAD was of prime concern. The tumor samples of the
GSE19804 dataset included different stages, and the findings of
WGCNA for GSE19804 would be more convincing and more significant
compared with other datasets in Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo). Although the
GSE19804 dataset refers to lung cancer in female non-smokers, the
result of WGCNA, in which the stage of the tumor was selected as a
main clinical feature, could not be significantly influenced.
Furthermore, other data of LUAD from the GSE40791 dataset and TCGA
(in UALCAN), which exhibited no significant differences in sex and
smoking status, were used for the validation of the hub genes.
Following adjustment for other predictors, the results demonstrated
the important role of the hub genes in the progression and
prognosis of LUAD. Validation of the hub genes based on protein
levels in the Human Protein Atlas was also performed.
There were 100 normal samples and 94 tumor samples
in the GSE40791 dataset. For validation, the GSE40791 dataset was
used to identify DEGs between normal and LUAD tissues, and the
dataset had a sufficient sample number. Additionally, the tumor
tissues in the GSE40791 dataset included 69, 12 and 13 stage I, II
and III LUAD frozen tissues, respectively. The tumor samples with
various stages of LUAD provided more reliable results.
To obtain further information on the role of the hub
module (turquoise module) in tumor progression, GO and KEGG pathway
enrichment analyses were performed based on DAVID. The genes were
generally enriched for ‘positive regulation of transcription from
RNA polymerase II promoter’, ‘signal transduction’, ‘negative
regulation of transcription from RNA polymerase II promoter’, ‘cell
adhesion’ and ‘positive regulation of GTPase activity’. Currently,
inhibitors of RNA polymerase II have the potential to be effective
anticancer drugs (23). The hub
genes, which are associated with regulating RNA polymerase II, may
serve as therapeutic targets for drug design. Signaling by small
GTPase, Ras-related protein 1/Rac1, is one of the major signaling
pathways controlling cancer cell migration and tumor metastasis
(24). The results revealed that the
hub genes were involved in tumor progression by regulating GTPase
activity. Additionally, a literature review of the hub genes was
conducted, and identified that the hub genes led to the induction
of apoptotic programs by regulating certain enzymes (25). For example, the molecular mechanisms
by which CA4 carries out its anti-invasive functions have been
identified to be mediated by the enhancement of E-cadherin
expression and the inhibition of N-cadherin and vimentin expression
(25). E-cadherin functions as a
suppressor of invasion, whereas N-cadherin and vimentin promote
cell motility and invasion in cancer (25–27).
The protein encoded by CA4 is one of 12 active human
isozymes. It is also one of four existing on the extracellular
surfaces of certain epithelial and endothelial cells (28). By interacting with Wilms' tumor
1-associating protein (WTAP), CA4 influences WTAP protein
degradation through polyubiquitination (25). Furthermore, it has been demonstrated
that a low expression of carbonic anhydrase IV can promote the
proliferation of cancer cells (29).
PECAM1 can code for CD31, which belongs to the adhesion molecule in
the immunoglobulin superfamily (30). Via the wingless-related integration
site signaling pathway, PECAM1 can maintain and restore vascular
integrity (30). This indicates that
PECAM1 is involved in the tumorigenesis of LUAD by regulating the
expression of vascular endothelial growth factor. Additionally,
CD31 is a member of I transmembrane glycoprotein that is enriched
in platelets, monocytes, endothelial cells and discrete circulating
lymphocytes (31). DNAJB4 belongs to
the heat shock protein 40 family (Hsp40/DnaJ), and serves an
important role in suppressing cancer metastasis (32). AGER is a member of the immunoglobulin
superfamily of cell surface molecules. AGER engagement activates
multiple intracellular signaling mechanisms to fuel chronic
inflammatory conditions, which can lead to malignant transformation
(33,34). Previous studies have demonstrated
that long non-coding RNA AGER can influence the development of lung
cancer by regulating the expression of AGER (34,35).
GIMAP6 belongs to the GIMAP gene family, which includes one
pseudogene and seven functional members. By regulating the
activation of T cells and cell death, GIMAP6 can modulate immune
function accurately (36). Although
it has been indicated that the association between
autophagy-related protein 8 and GIMAP6 is of importance to
autophagy (37), a deeper
understanding of the functions of GIMAP6 is required. C10orf54
encodes for V-set immunoregulatory receptor (VISTA), which belongs
to I transmembrane proteins. There is an ~30-amino acid stalk, a
single N-terminal immunoglobulin V domain, a 95 amino acid
cytoplasmic tail and a transmembrane domain in VISTA (38,39). A
recent study found that the expression levels of C10orf54 were
closely associated with tumor immune evasion (40). The protein encoded by DOCK4 is a
member of a large family of proteins (CED5/DOCK180/MYOBLAST CITY
class), and a previous study suggested that it could suppress tumor
growth in several types of cancer, including ovarian cancer, breast
cancer and glioblastoma (41–43).
GOLM1 is a resident cis-Golgi membrane protein. On the surface of
the Golgi apparatus, there is an extensive C-terminal, coiled-coil
domain and a single N-terminal transmembrane domain (44). It has been confirmed that the Golgi
apparatus serves an active role in cell migration, which can be
activated by post-translational modification and prominent
alterations (45). The
overexpression of microRNA-200a, which has been reported to be
involved in blocking the increase in cell proliferation, can
repress LUAD cell proliferation induced by the overexpression of
GOLM1 (46). PAFAH1B3 serves an
important role in tumorigenic features and aggressiveness. PAFAH1B3
encodes a catalytic subunit of platelet-activating factor (PAF)
acetyl hydrolase 1b (Pafah1b) (47).
By hydrolyzing PAF, Pafah1b can regulate the intracellular PAF
levels and may lead to the evasion of apoptosis caused by high
intracellular PAF concentrations (47,48). In
addition, by regulating an optimal landscape of signaling lipids,
PAFAH1B3 can weaken the aggressiveness of cancer and regulate
cancer cell pathogenicity (49).
In conclusion, the present study identified the
turquoise module and nine hub genes (CA4, PECAM1, DNAJB4, AGER,
GIMAP6, C10orf54, DOCK4, GOLM1 and PAFAH1B3), which are of
importance to the development of LUAD. Through the turquoise
module, further information on the mechanisms of tumorigenesis in
LUAD was obtained. In the future, these nine hub genes that serve a
vital role in LUAD tumorigenesis may also contribute to early
diagnosis and treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by Key Projects of
Hubei Provincial Health and Family Planning Commission (grant no.
WJ2017Z006), the Zhongnan Hospital of Wuhan University Science,
Technology and Innovation Cultivating Fund (grant no. cxpy2017041),
and the 351 Talent Project of Wuhan University (Luojia Young
Scholars: SL).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
WH and SL conceived and designed the study. DY, JH
and XL performed the analysis procedures. DY, JH, XR, CC and XL
analyzed the results. XR, XL, WH and SL contributed to the analysis
tools. DY and JH contributed to the writing of the manuscript. All
authors reviewed the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Knight SB, Crosbie PA, Balata H, Chudziak
J, Hussell T and Dive C: Progress and prospects of early detection
in lung cancer. Open Biol. 7(pii): 1700702017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chang JT, Lee YM and Huang RS: The impact
of the cancer genome atlas on lung cancer. Transl Res. 166:568–585.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Martin P and Leighl NB: Review of the use
of pretest probability for molecular testing in non-small cell lung
cancer and overview of new mutations that may affect clinical
practice. Ther Adv Med Oncol. 9:405–413. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao X, Li X, Zhou L, Ni J, Yan W, Ma R,
Wu J, Feng J and Chen P: LncRNA HOXA11-AS drives cisplatin
resistance of human LUAD cells via modulating miR-454-3p/Stat3.
Cancer Sci. 109:3068–3079. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gan TQ, Chen WJ, Qin H, Huang SN, Yang LH,
Fang YY, Pan LJ, Li ZY and Chen G: Clinical value and prospective
pathway signaling of MicroRNA-375 in lung adenocarcinoma: A study
based on the cancer genome atlas (TCGA), gene expression omnibus
(GEO) and bioinformatics analysis. Med Sci Monit. 23:2453–2464.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shi YX, Zhu T, Zou T, Zhuo W, Chen YX,
Huang MS, Zheng W, Wang CJ, Li X, Mao XY, et al: Prognostic and
predictive values of CDK1 and MAD2L1 in lung adenocarcinoma.
Oncotarget. 7:85235–85243. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shang J, Song Q, Yang Z, Li D, Chen W, Luo
L, Wang Y, Yang J and Li S: Identification of lung adenocarcinoma
specific dysregulated genes with diagnostic and prognostic value
across 27 TCGA cancer types. Oncotarget. 8:87292–87306. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kober KM, Olshen A, Conley YP, Schumacher
M, Topp K, Smoot B, Mazor M, Chesney M, Hammer M, Paul SM, et al:
Expression of mitochondrial dysfunction-related genes and pathways
in paclitaxel-induced peripheral neuropathy in breast cancer
survivors. Mol Pain. 14:17448069188164622018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
El-Aarag SA, Mahmoud A, Hashem MH, Abd
Elkader H, Hemeida AE and ElHefnawi M: In silico identification of
potential key regulatory factors in smoking-induced lung cancer.
BMC Med Genomics. 10:402017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fan L, Yu X, Huang Z, Zheng S, Zhou Y, Lv
H, Zeng Y, Xu JF, Zhu X and Yi X: Analysis of Microarray-identified
genes and microRNAs associated with idiopathic pulmonary fibrosis.
Mediators Inflamm. 2017:18042402017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li S, Liu X, Liu T, Meng X, Yin X, Fang C,
Huang D, Cao Y, Weng H, Zeng X and Wang X: Identification of
biomarkers correlated with the TNM staging and overall survival of
patients with bladder cancer. Front Physiol. 8:9472017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kadara H, Choi M, Zhang J, Parra ER,
Rodriguez-Canales J, Gaffney SG, Zhao Z, Behrens C, Fujimoto J,
Chow C, et al: Whole-exome sequencing and immune profiling of
early-stage lung adenocarcinoma with fully annotated clinical
follow-up. Ann Oncol. 29:1072. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu TP, Lee JM, Hsu CP, et al: Genome-wide
screening of genomic alterations and transcriptional modulation in
non-smoking female lung cancer in Taiwan. Cancer Res. 69:2009.
|
|
16
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: Affy - analysis of affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Langfelder P, Zhang B and Horvath S:
Defining clusters from a hierarchical cluster tree: The Dynamic
Tree Cut package for R. Bioinformatics. 24:719–720. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang J, Kong D, Cui Q, Wang K, Zhang D,
Gong Y and Wu G: Prognostic genes of breast cancer identified by
gene co-expression network analysis. Fron Oncol. 8:4742018.
|
|
19
|
Zhang Y, Foreman O, Wigle DA, Kosari F,
Vasmatzis G, Salisbury JL, van Deursen J and Galardy PJ: USP44
regulates centrosome positioning to prevent aneuploidy and suppress
tumorigenesis. J Clin Invest. 122:4362–4374. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Robinson DR, Wu YM, Lonigro RJ, Vats P,
Cobain E, Everett J, Cao X, Rabban E, Kumar-Sinha C, Raymond V, et
al: Integrative clinical genomics of metastatic cancer. Nature.
548:297–303. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oronsky B, Reid TR, Oronsky A and Carter
CA: What's new in SCLC? A review. Neoplasia. 19:842–847. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee JW, Ryu YK, Ji YH, Kang JH and Moon
EY: Hypoxia/reoxygenation-experienced cancer cell migration and
metastasis are regulated by Rap1-and Rac1-GTPase activation via the
expression of thymosin beta-4. Oncotarget. 6:9820–9833.
2015.PubMed/NCBI
|
|
25
|
Zhang J, Tsoi H, Li X, Wang H, Gao J, Wang
K, Go MY, Ng SC, Chan FK, Sung JJ and Yu J: Carbonic anhydrase IV
inhibits colon cancer development by inhibiting the Wnt signalling
pathway through targeting the WTAP-WT1-TBL1 axis. Gut.
65:1482–1493. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Canel M, Serrels A, Frame MC and Brunton
VG: E-cadherin-integrin crosstalk in cancer invasion and
metastasis. J Cell Sci. 126:393–401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pan TL, Wang PW, Huang CC, Yeh CT, Hu TH
and Yu JS: Network analysis and proteomic identification of
vimentin as a key regulator associated with invasion and metastasis
in human hepatocellular carcinoma cells. J Proteomics.
75:4676–4692. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Waheed A and Sly WS: Membrane associated
carbonic anhydrase IV (CA IV): A personal and historical
perspective. Subcell Biochem. 75:157–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen J, Hu L, Zhang F, Wang J, Chen J and
Wang Y: Downregulation of carbonic anhydrase IV contributes to
promotion of cell proliferation and is associated with poor
prognosis in non-small cell lung cancer. Oncol Lett. 14:5046–5050.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Villar J, Zhang H and Slutsky AS: Lung
repair and regeneration in acute respiratory distress syndrome:
Role of PECAM1 and Wnt signaling. Chest. 155:587–594. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ren Q, Ren L, Ren C, Liu X, Dong C and
Zhang X: Platelet endothelial cell adhesion molecule-1 (PECAM1)
plays a critical role in the maintenance of human vascular
endothelial barrier function. Cell Biochem Funct. 33:560–565. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen CH, Chang WH, Su KY, Ku WH, Chang GC,
Hong QS, Hsiao YJ, Chen HC, Chen HY, Wu R, et al: HLJ1 is an
endogenous Src inhibitor suppressing cancer progression through
dual mechanisms. Oncogene. 35:5674–5685. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Riehl A, Nemeth J, Angel P and Hess J: The
receptor RAGE: Bridging inflammation and cancer. Cell Commun
Signal. 7:122009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sparvero LJ, Asafu-Adjei D, Kang R, Tang
D, Amin N, Im J, Rutledge R, Lin B, Amoscato AA, Zeh HJ, et al:
RAGE (Receptor for Advanced Glycation Endproducts), RAGE Ligands,
and their role in cancer and inflammation. J Transl Med. 7:172009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pan Z, Liu L, Nie W, Miggin S, Qiu F, Cao
Y, Chen J, Yang B, Zhou Y, Lu J and Yang L: Long non-coding RNA
AGER-1 functionally upregulates the innate immunity gene AGER and
approximates its anti-tumor effect in lung cancer. Mol Carcinog.
57:305–318. 2018. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ho CH and Tsai SF: Functional and
biochemical characterization of a T cell-associated anti-apoptotic
protein, GIMAP6. J Biol Chem. 292:9305–9319. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pascall JC, Rotondo S, Mukadam AS, Oxley
D, Webster J, Walker SA, Piron J, Carter C, Ktistakis NT and
Butcher GW: The immune system GTPase GIMAP6 interacts with the Atg8
homologue GABARAPL2 and is recruited to autophagosomes. PLoS One.
8:e777822013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang L, Rubinstein R, Lines JL, Wasiuk A,
Ahonen C, Guo Y, Lu LF, Gondek D, Wang Y, Fava RA, et al: VISTA, a
novel mouse Ig superfamily ligand that negatively regulates T cell
responses. J Exp Med. 208:577–592. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Flies DB, Wang S, Xu H and Chen L: Cutting
edge: A monoclonal antibody specific for the programmed death-1
homolog prevents graft-versus-host disease in mouse models. J
Immunol. 187:1537–1541. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xie S, Huang J, Qiao Q, Zang W, Hong S,
Tan H, Dong C, Yang Z and Ni L: Expression of the inhibitory B7
family molecule VISTA in human colorectal carcinoma tumors. Cancer
Immunol Immunother. 67:1685–1694. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu J-R, Tai Y, Jin Y, Hammell MC,
Wilkinson JE, Roe J-S, Vakoc CR and Van Aelst L: TGF-β/Smad
signaling through DOCK4 facilitates lung adenocarcinoma metastasis.
Genes Dev. 29:250–261. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Westbrook JA, Wood SL, Cairns DA, McMahon
K, Gahlaut R, Thygesen H, Shires M, Roberts S, Marshall H, Oliva
MR, et al: Identification and validation of DOCK4 as a potential
biomarker for risk of bone metastasis development in patients with
early breast cancer. J Pathol. 247:381–391. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Debruyne DN, Turchi L, Burel-Vandenbos F,
Fareh M, Almairac F, Virolle V, Figarella-Branger D, Baeza-Kallee
N, Lagadec P, Kubiniek V, et al: DOCK4 promotes loss of
proliferation in glioblastoma progenitor cells through nuclear
beta-catenin accumulation and subsequent miR-302-367 cluster
expression. Oncogene. 37:241–254. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Donizy P, Kaczorowski M, Biecek P, Halon
A, Szkudlarek T and Matkowski R: Golgi-related proteins GOLPH2
(GP73/GOLM1) and GOLPH3 (GOPP1/MIDAS) in cutaneous melanoma:
Patterns of expression and prognostic significance. Int J Mol Sci.
17(pii): E16192016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hu L, Li L, Xie H, Gu Y and Peng T: The
Golgi localization of GOLPH2 (GP73/GOLM1) is determined by the
transmembrane and cytoplamic sequences. PLoS One. 6:e282072011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Arun a and Li LM: Overexpression of golgi
membrane protein 1 promotes non-small-cell carcinoma aggressiveness
by regulating the matrix metallopeptidase 13. Am J Cancer Res.
8:551–565. 2018.PubMed/NCBI
|
|
47
|
McIntyre TM, Prescott SM and Stafforini
DM: The emerging roles of PAF acetylhydrolase. J Lipid Res.
(Suppl):50:S255–S259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bonin F, Ryan SD, Migahed L, Mo F, Lallier
J, Franks DJ, Arai H and Bennett SAL: Anti-apoptotic actions of the
platelet-activating factor acetylhydrolase I alpha2 catalytic
subunit. J Biol Chem. 279:52425–52436. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mulvihill MM, Benjamin DI, Ji X, Le Scolan
E, Louie SM, Shieh A, Green M, Narasimhalu T, Morris PJ, Luo K and
Nomura DK: Metabolic profiling reveals PAFAH1B3 as a critical
driver of breast cancer pathogenicity. Chem Biol. 21:831–840. 2014.
View Article : Google Scholar : PubMed/NCBI
|