Introduction
Pancreatic ductal adenocarcinoma (PDAC) is a highly
malignant tumor of the digestive system, which exhibits a
high-mortality rate (1). Advances in
surgical and chemotherapeutic techniques have improved the
treatment and overall survival (OS) rates of the majority of
patients with gastrointestinal tumors; however, these advances have
not yet conferred improvement on patients with PDAC, who currently
exhibit a 5-year relative OS rate of 8%, in the USA (2). The early diagnosis of PDAC is rare and,
consequently, numerous patients are in the advanced stages upon
primary diagnosis, which results in fewer surgical options and an
increase in chemotherapeutic resistance (2). Currently used clinical biomarkers, such
as carbohydrate antigen 19-9 and carcinoembryonic antigen, are not
able to accurately diagnose early disease or predict the
chemotherapeutic response/OS times in patients with PDAC (3). Therefore, the identification of novel
biomarkers for PDAC progression and treatment evaluation represents
a shared objective for both clinical and laboratory staff, which
may confer great benefit on patient outcomes.
MicroRNAs (miRNAs) are a form of endogenous
non-coding RNAs that consist of 20–24 nucleotides and serve a
post-transcriptional regulatory role, via binding to the
3′-untranslated region (3′-UTR) of mRNAs (4). It has been reported that >1,000
miRNAs are encoded by the mammalian genome (5), with 30% of the human genome (~5,300
genes) targeted by miRNAs (6).
Abnormal expression of miRNAs has been revealed in numerous types
of cancer and is implicated in various physiological and
pathological processes of tumor cells, such as proliferation,
differentiation, apoptosis, metabolism, metastasis and cell
signaling (7). As a result of
significant advances in bioinformatics and microarray techniques,
human genome sequencing represents a novel technique for the
identification of the genes that influence cancer progression. Gene
expression microarray analysis has been used to discover multiple
molecular signatures associated with PDAC progression (8,9). A
plethora of differentially expressed miRNAs have been discovered by
comparing the gene expression profiles between cancer tissues and
adjacent tissues of European and American populations (10). However, due to differences in the
genetic backgrounds and living environments between the
aforementioned populations and Asian populations, the findings may
not be transferable. Therefore, more experiments need to be
conducted on Asian populations in order to identify miRNAs that are
highly specific and sensitive to PDAC, allowing for the accurate
predictions of diagnosis and prognosis.
The aim of the present study was to identify miRNAs
that are able to predict the prognosis of patients with PDAC. miRNA
expression levels were detected in cancerous tissues from six
patients with different prognoses of PDAC via microarray analysis,
and the top 10 miRNAs with the largest differences in fold change
were screened and verified in an independent group of 68 patients.
It was revealed that a low expression level of miRNA-608 was
associated with poor prognosis. Furthermore, overexpression of
miRNA-608 in pancreatic cancer cell lines (Panc-1 and Bxpc-3)
significantly increased apoptosis and decreased the protein level
of bromodomain-containing protein (BRD)-4 (no difference was
observed in transcription). Finally, luciferase reporter gene and
rescue experiments were conducted and confirmed that miRNA-608
promotes apoptosis in PDAC tissues via the negative regulation of
BRD4. The current results provide a theoretical basis for a novel
technique to predict the prognosis of PDAC patients, which may
result in improved individualized treatment and the identification
of new drug targets.
Materials and methods
Gene expression microarray
analysis
A total of six patients with PDAC who had received a
surgical resection at Tianjin Medical University Cancer Institute
& Hospital (TMUCIH; Tianjin, China) between January 2008 and
December 2015 were recruited for the present study, and gene
expression dataset was collected (clinicopathological data of the
patients are detailed in Table I).
The six samples were divided into two groups, dependent on
prognosis. The long-survival group included three patients with OS
times >30 months, and the short-survival group consisted of
three patients with OS times <12 months. The inclusion criteria
were as follows: i) Patients who underwent surgery from the same
surgeon in Tianjin Medical University Cancer Institute &
Hospital; ii) patients who had a pathological diagnosis of PDAC;
iii) patients who had received an R0 resection; iv) patients who
had not received preoperative chemoradiotherapy; and v) patients
who exhibited no serious postoperative complications. The exclusion
criteria were as follows: i) Patients who died with 7–12 days
following surgery; ii) patients with severe postoperative
complications requiring secondary surgery or ICU treatment; and
iii) patients whose death was not related to tumor.
 | Table I.Clinicopathological characteristics of
the six patients. |
Table I.
Clinicopathological characteristics of
the six patients.
|
| Long survival
group | Short survival
group |
|---|
|
|
|
|
|---|
| Patient no. | 1 | 2 | 3 | 4 | 5 | 6 |
|---|
| Sex | Male | Female | Male | Male | Male | Male |
| Age, years | 48 | 36 | 65 | 72 | 69 | 60 |
| Differentation
state | Medium | Poor | High | High | Medium | Poor |
| TNM | T4N0M0 | T1N1M0 | T2N0M0 | T1bN0M0 | T3N1M0 | T1bN1M0 |
| AJCC Stage | III | IIB | I | IIA | IIB | III |
| OS, months | 30 | 39 | 33 | 9 | 7 | 12 |
| RFS, months | 18 | 20 | 33 | 5 | 1 | 11 |
Total RNA was extracted from cancer and
paracancerous tissues using TRIzol® (Invitrogen; Thermo
Fisher Scientific, Inc.)/chloroform and purified with Agencourt
Ampure magnetic beads (cat, no, APN 000132; Beckman Coulter, Inc.).
Sample preparation for the microarray processing was carried out
according to the protocol detailed in the GeneChip® WT
PLUS Reagent kit (Thermo Fisher Scientific, Inc.). Briefly, a total
of 0.5 µg RNA was subjected to two rounds of cDNA synthesis.
Following fragmentation, the second-cycle single-stranded (ss)cDNA
was labeled with biotin using terminal deoxynucleotidyl transferase
(TdT). Subsequently the sample was hybridized using the Affymetrix
Human Gene 1.0ST Array (Thermo Fisher Scientific, Inc.) for 16–18 h
at 45°C. Following hybridization, the microarrays were washed and
stained with streptavidin phycoerythrin on the Affymetrix Fluidics
Station 450. Microarrays were scanned using the
Affymetrix® GeneChip Command Console (AGCC; Thermo
Fisher Scientific, Inc.), which was installed in
GeneChip® Scanner 3000 7G (https://www.thermofisher.com/order/catalog/product/00-0213).
The data were analyzed using the Robust Multichip Analysis (RMA)
algorithm in the Partek® Genomics Suite 6.6(http://cgs.hku.hk/portal/index.php/software-for-microarray-analysis/partek-genome-suite-with-partek-pathway),
the default analysis settings were used, and global scaling was
used as the normalization method. The values presented are the log2
RMA signal intensity.
Clinicopathological characteristics
and follow-up of study population
A total of 68 patients with pancreatic cancer, who
had received a surgical resection at Tianjin Medical University
Cancer Institute & Hospital (TMUCIH; Tianjin, China) between
January 2008 and December 2015, were selected for use in the
present study. All patients had received a confirmation of
pancreatic cancer diagnosis from a pathologist in the Department of
Pathology, TMUCIH. A total of 37 males and 31 females (median age,
61 years; range, 36–80 years) were included in the present study.
Of the included patients, five were diagnosed with
well-differentiated tumors (5/68; 7.4%), 55 with
moderately-differentiated tumors (55/68; 80.9%) and eight with
poorly-differentiated tumors (8/68; 11.8%), and a total of 17
patients (17/68; 25.0%) presented with lymphatic invasion.
Furthermore, 21/68 (30.9%), 30/68 (44.1%), 15/68 (22.1%) and 2/68
patients (2.9%) were diagnosed at stages I, IIA, IIB and III,
respectively, according to the 7th American Joint Committee on
Cancer Tumor-Node-Metastasis classification system (11). The median OS time was 18.4 months and
the 2-year OS rate was 35.2%. The clinicopathological
characteristics of the 68 patients with PDAC are detailed in
Table II. The use of all human
tissues in the present study was reviewed and approved by the
Ethics Committee of TMUCIH, and written informed consent was
provided by all participants.
| Table II.Clinicopathological characteristics
and OS times of 68 patients with pancreatic cancer. |
Table II.
Clinicopathological characteristics
and OS times of 68 patients with pancreatic cancer.
| Characteristic | n | % |
|---|
| Sex |
|
|
| Male | 37 | 54.4 |
|
Female | 31 | 45.6 |
| Age |
|
|
| ≤60
years | 31 | 45.6 |
| >60
years | 37 | 54.4 |
|
Differentiation |
|
|
|
Low | 8 | 11.8 |
|
Medium | 55 | 80.9 |
|
High | 5 | 7.3 |
| Tumor size |
|
|
| ≤4
cm | 40 | 58.8 |
| >4
cm | 28 | 41.2 |
| Vascular
invasion |
|
|
|
Present | 13 | 19.1 |
|
Absent | 55 | 80.9 |
| T stage |
|
|
|
T1+T2 | 28 | 41.2 |
| T3 | 40 | 58.8 |
| N stage |
|
|
| N0 | 51 | 75.0 |
| N1 | 17 | 25.0 |
| AJCC stage |
|
|
| I | 21 | 30.9 |
|
IIA | 30 | 44.1 |
|
IIB | 15 | 22.1 |
|
III | 2 | 2.9 |
| OS |
|
|
| ≤12
months | 23 | 33.8 |
| >12
months | 45 | 66.2 |
RNA samples and quantitative (q)
PCR
Genomic RNA samples from 68 patients were collected
from the TMUCIH tumor tissue bank. cDNA was synthesized from 0.5 µg
of total RNA using the miRcute Plus miRNA First-Strand cDNA Kit
(Tiangen Biotech Co., Ltd.) according to the manufacturer's
instructions. Candidate genes and U6 were amplified using qPCR in a
fluorescence reader QuantStudio 5 Real-Time PCR system (Applied
Biosystems). Each sample was run in a miRcute Plus miRNA qPCR kit
(Tiangen Biotech Co., Ltd.). The thermocycling conditions were as
follows: Initial enzyme activation at 95°C for 15 min, followed by
40 cycles of denaturing at 94°C for 20 sec, and then
annealing/extension at 60°C for 34 sec. mRNA levels were quantified
using the 2−ΔΔCq method and normalized to U6 (12). The sequences of the primers used in
the present study are presented in Table III.
| Table III.Sequences designed for the validation
of candidate miR levels and U6 using qPCR. |
Table III.
Sequences designed for the validation
of candidate miR levels and U6 using qPCR.
| Primer | Sequence
(5′-3′) |
|---|
| miR-221 |
CGAGCTACATTGTCTGCTGGGTTTC |
| miR-492 |
AGGACCTGCGGGACAAGATTCTT |
| miR-192 |
GCGCTGACCTATGAATTGACAGCC |
| miR-573 |
GCGCTGAAGTGATGTGTAACTGATCAG |
| miR-222 |
GCGTGTCAGTTTGTCAAATACCCCA |
| miR-106b |
GCGTAAAGTGCTGACAGTGCAGAT |
| miR-409 |
AGGTTACCCGAGCAACTTTGCAT |
| miR-21 |
GGCGCTAGCTTATCAGACTGATGTTG |
| miR-186 |
GCCCAAAGAATTCTCCTTTTGGGCT |
| miR-608 |
AGGGGTGGTGTTGGGACAGC |
| U6-Forward |
GCGCGTCGTGAAGCGTTC |
| U6-Reverse |
GTGCAGGGTCCGAGGT |
Cell transfection
The pancreatic cancer cell lines Panc-1 and Bxpc-3
(American Type Culture Collection; ATCC) were transfected with
miR-608 mimics (50 nM) and their respective negative controls (NC;
non-targeting sequence; 50 nM), which were acquired from Guangzhou
RiboBio Co., Ltd. The sequence of miR-608 mimic was as follow:
5′-AGGGGUGGUGUUGGGACAGCUCCGU-3′. Opti-MEM® I and
Lipofectamine 3000 reagent (both from Thermo Fisher Scientific,
Inc.) were used for transfection. The cells were collected 24 h
after transfection.
Flow cytometry
Flow cytometry was performed using a FITC Annexin V
Apoptosis Detection kit I (Becton, Dickinson and Company). Panc-1
and Bxpc-3 cells (with or without miR-608 mimic transfection) were
seeded in 6-well plates at a density of ~4×105
cells/well. After overnight incubation at 37°C and 5%
CO2, the cells were collected, washed twice with cold
PBS, resuspended in 500 µl 1× binding buffer at a concentration of
1×106 cells/ml, and incubated with 5 µl V-fluorescein
isothiocyanate (FITC) and 5 µl propidium iodide (Thermo Fisher
Scientific, Inc.). The samples were collected via centrifugation at
800 rpm/min and 25°C and incubated for 15 min at 25°C in the dark.
Flow cytometry was performed within 1 h, using a FACScan flow
cytometer (version V10.0; BD Biosciences).
The prediction of miR-608 target by
bioinformatics analyses
The potential target genes of miR-608 were predicted
by using TargetScan (http://www.targetscan.org/), miRDB (http://mirdb.org/) and miRanda (http://miranda.org.uk/). Venn diagram was drawn usinkg
R Project (X64; version 3.2.5) to obtain genes that appeared
simultaneously on the three websites. Subsequently RNAhybrid
(https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid/)
was used to evaluate the binding capacity of miR-608 to its
target.
Western blotting
Total protein was extracted from Bxpc-3 and Panc-1
cell lines using RIPA buffer supplemented with protease and
phosphatase inhibitors (Roche Diagnostics; cat. no. 04906837001).
Total protein was quantified using a BCA protein assay kit.
Proteins (20 µg) were separated via SDS-PAGE (10–15% gel). The
separated proteins were transferred to nitrocellulose membranes,
which were then blocked with 5% non-fat milk in Tris-buffered
saline-Tween-20 (TBST) overnight at 4°C. The membranes were
incubated with primary antibodies against BRD4 (1:500; Abcam; cat.
no. ab244221) and GAPDH (1:1,000; Cell Signaling Technology, Inc.;
cat. no. 8884). The proteins were then incubated with secondary
antibodies conjugated with horseradish peroxidase (OriGene
Technologies, Inc.; cat. no. DS-0002) for 1 h at room temperature.
The immunoreactive protein bands were visualized using enhanced
chemiluminescence (ECL kit; Santa Cruz Biotechnology) and scanned
using Amersham Imager 600 (GE Healthcare). Western blotting
experiments were performed three times.
Dual-luciferase assay
A dual-luciferase reporter assay was performed in
order to determine the effect of miR-608 on the BRD4 3′
untranslated region (3′UTR) and the functional binding sites in the
BRD4 3′UTR. BRD4-3′UTR-Wild type (Wt) and BRD4-3′UTR-Mutant (Mut;
all from Shanghai GeneChem Co., Ltd.)] were produced by sub-cloning
the full-length 3′UTR fragments of the BRD4 gene and its mutant
into the miR-608 binding sites using a pmirGlo Dual-Luciferase
miRNA target expression vector (Promega Corporation). 293T (ATCC),
Panc-1 and Bxpc-3 cells were seeded into 96-well plates
(5×103 cells/well). Following incubation for 24 h at
37°C, the cells were co-transfected with either BRD4-3′UTR-Wt,
BRD4-3′UTR-Mut, miR-608 or miR-608 NC. The cells were divided into
three groups as follows: miR-608 negative control (NC) +
BRD4-3′UTR-Wt; miR-608 + BRD4-3′UTR-Wt; and miR-608 +
BRD4-3′UTR-Mut. Following transfection for 24 h, luciferase assays
were performed using the Dual-Luciferase Reporter Assay system
(Promega Corporation), and the firefly luciferase activity was
normalized to Renilla luciferase activity.
BRD4 rescue experiment
The BRD4 plasmid was purchased from Shanghai
Genechem Co., Ltd. and short hairpin (sh) BRD4 was purchased from
Guangzhou RiboBio Co., Ltd. The experiment included four groups:
miR-608 mimics + NC plasmid; miR-608 mimics + BRD4 plasmid; NC
mimics + siBRD4 plasmid; and NC mimics + NC plasmid. Each group of
mimics and BRD4 were co-transfected into Panc-1 and Bxpc-3 cells.
The cells were harvested 24 h after transfection and were analyzed
via flow cytometry. The sequence of shBRD4 was as follows:
5′-GATCCGCCTGGAGATGACATAGTCTTATTCAAGAGATAAGACTATGTCATCTCCAGGTTTTTTC-3′
Statistical analysis
Statistical analysis was performed using SPSS
software (version 16.0; SPSS, Inc.). Receiver operating
characteristic (ROC) curves were constructed and the area under the
curve (AUC) was calculated in order to estimate the OS times for
the selected candidate genes. The difference between two groups was
assessed using the Student's t-test and one-way ANOVA followed by
Bonferroni post hos test was used for comparisons between multiple
groups. Kaplan-Meier analysis was utilized to generate survival
curves and these were compared using the log-rank test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Screening for differentially expressed
miRNAs (DEMs) between patients with pancreatic cancer exhibiting
different prognoses using a gene expression microarray
In order to identify the potential prognostic
markers correlated with OS time, a microarray analysis was
conducted on samples from six patients with PDAC. Compared with the
short-survival group (OS, <12 months), 390 miRNAs (65.9%) were
upregulated and 201 (34.1%) were downregulated (|fold change|
>1) in the long-survival group (OS, >30 months; data not
shown). Combined with our preliminary study (13) and associated literature retrieved
from PubMed, 10 miRNAs (miRNA-21, −106b, −186, −192, −221, −222,
−409, −492, −573 and −608) were selected to investigate their
association with OS in a population of 68 patients with PDAC. The
expression levels of these 10 miRNAs were detected using qPCR, and
a Kaplan-Meier plot was constructed in order to analyze the
association between the miRNA expression levels and OS. It was
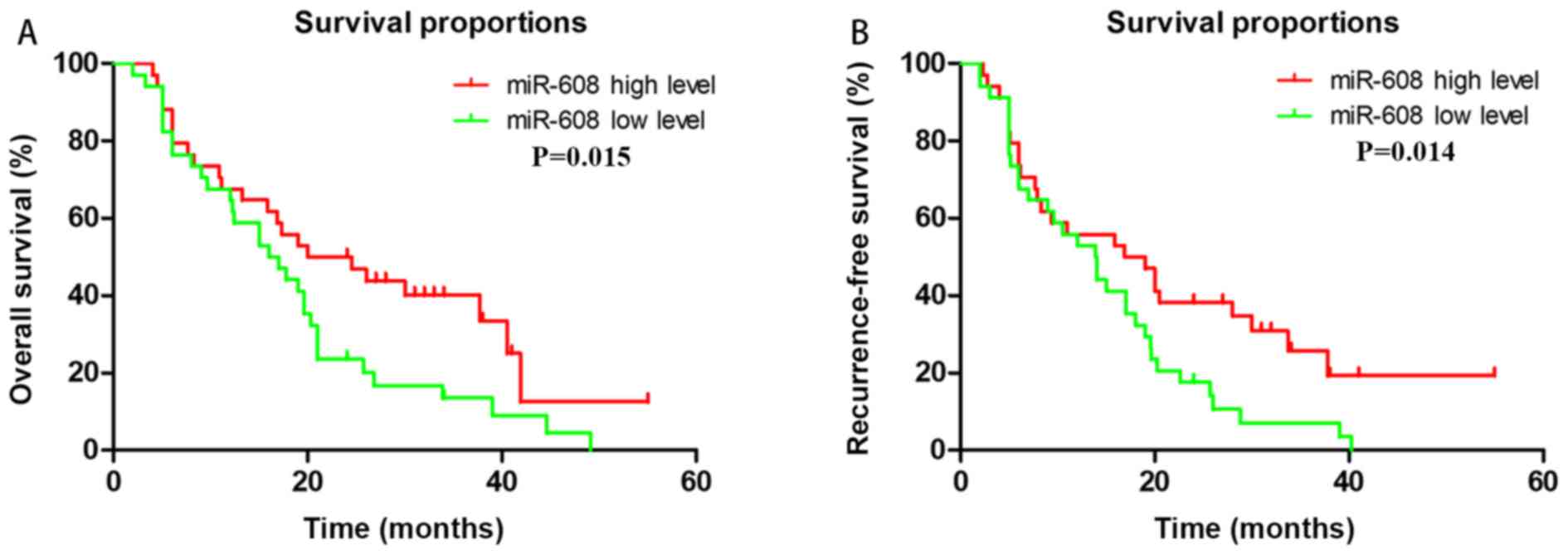
revealed that only miRNA-608 expression was able to accurately
predict patient outcomes. The results demonstrated that high
expression of miRNA-608 was associated with improved OS (P=0.015)
and RFS (P=0.014; Fig. 1).
miRNA-608 induces apoptosis in
pancreatic cancer cell lines
miRNA-608 was associated with the prognosis of
patients with PDAC. However, the mechanism behind the association
between miR-608 and pancreatic cancer was unclear. miRNAs
post-transcriptionally regulate their target genes, thus, 3 online
bioinformatics tools were utilized to estimate potential target
genes; TargetScan, miRDB and miRanda websites yielded 5,267, 916
and 2,840 possible miR-608 targets, respectively. R Project was
then used to obtain 638 genes from all three bioinformatics tools.
Subsequently, the online bioinformatics tool RNAhybrid indicated
that there were high scoring binding sites from miRNA-608 in the
3′-UTR of BRD4 mRNA. It was previously reported that BRD4 serves a
role in the modulation of the transcription of certain essential
genes associated with apoptosis, including c-Myc and BCL2 (14). These data suggested that miRNA-608
was primarily associated with ‘apoptosis’ and ‘proliferation’ via
the targeting of BRD4.
In order to verify the results predicted using
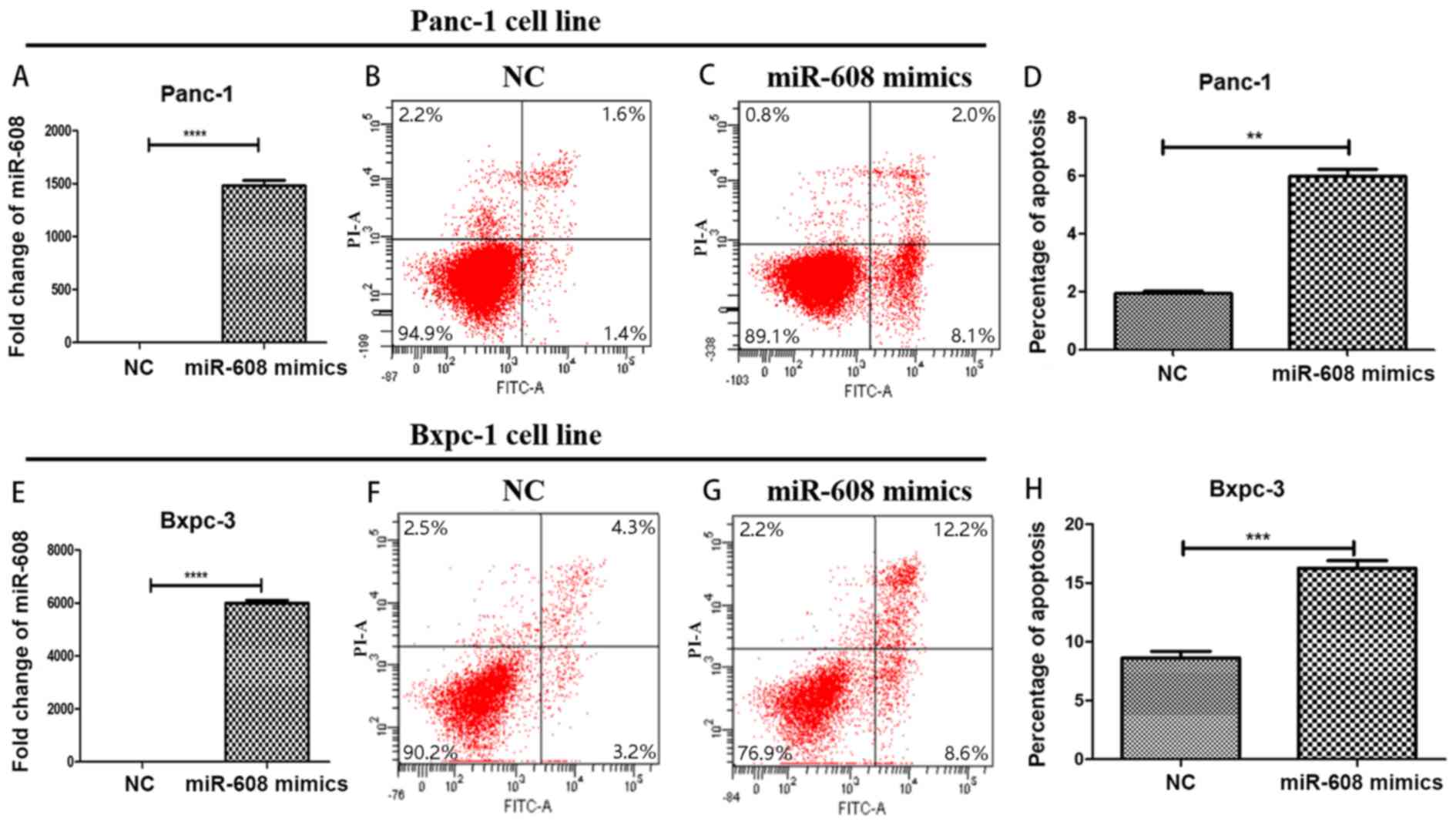
bioinformatics, miRNA-608 was overexpressed in two pancreatic
cancer cell lines: Panc-1 and Bxpc-3, using mimics. qPCR was used
to confirm the transfection success and was compared with the NC
group. The expression level of miRNA-608 in the mimic group was
significantly higher. The fold changes were increased by ~1,500 and
~6,000 times in Panc-1 and Bxpc-3, respectively (P<0.0001). Flow
cytometry was then performed in order to detect the function of
miRNA-608, and it was revealed that a high level of miRNA-608
expression promoted apoptosis in both cell lines used (Fig. 2).
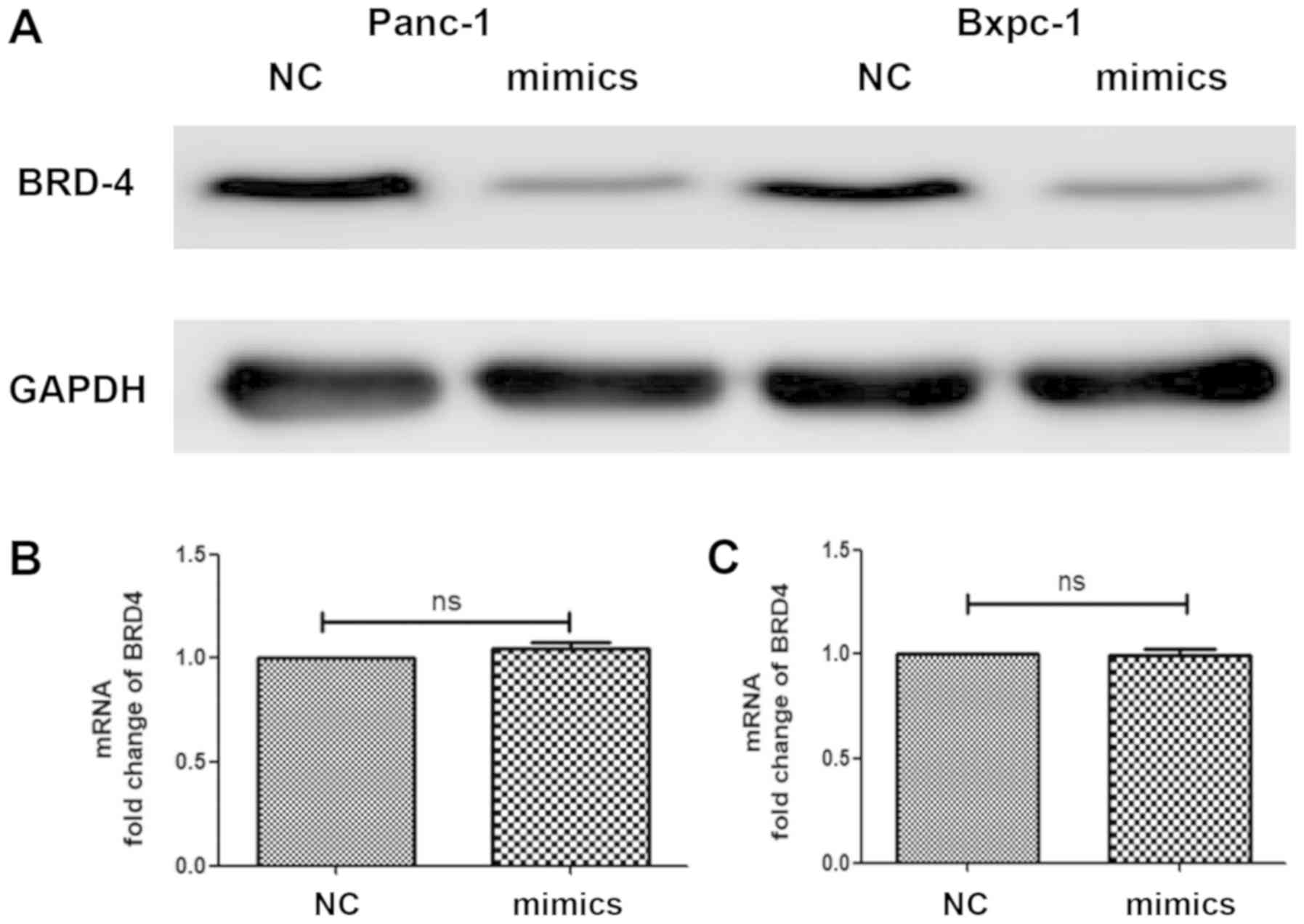
Furthermore, mimics were used to increase miRNA-608
expression and cells were collected in order to detect the
expression level of BRD4 in both transcription and translation
processes, using qPCR and western blot analyses, respectively. It
was revealed that the protein levels of BRD4 in Panc-1 and Bxpc-3
were significantly decreased following overexpression of miRNA-608,
but there was no change in the BRD4 mRNA level (Fig. 3).
In order to investigate miRNA-608 regulation of BRD4
after transcription, the fragment of the BRD4 3′-UTR containing the
predicted wild-type miRNA-608 binding site was incorporated into
the downstream region of firefly luciferase, in the Dual-Luciferase
miRNA Target Expression Vector, and a vector with the corresponding
mutated binding sites was simultaneously constructed. The relative
luciferase activity in 293T, Panc-1 and Bxpc-3 cells co-transfected
with miRNA-608 mimic and a Wt vector was inhibited, while in cells
transfected with the miR-608 mimic mutant vector, luciferase
activity was unaffected (Fig. 4).
The results of the present study supports the hypothesis that
miRNA-608 may directly inhibit the expression of BRD4, via binding
to the 3′-UTR of its mRNA.
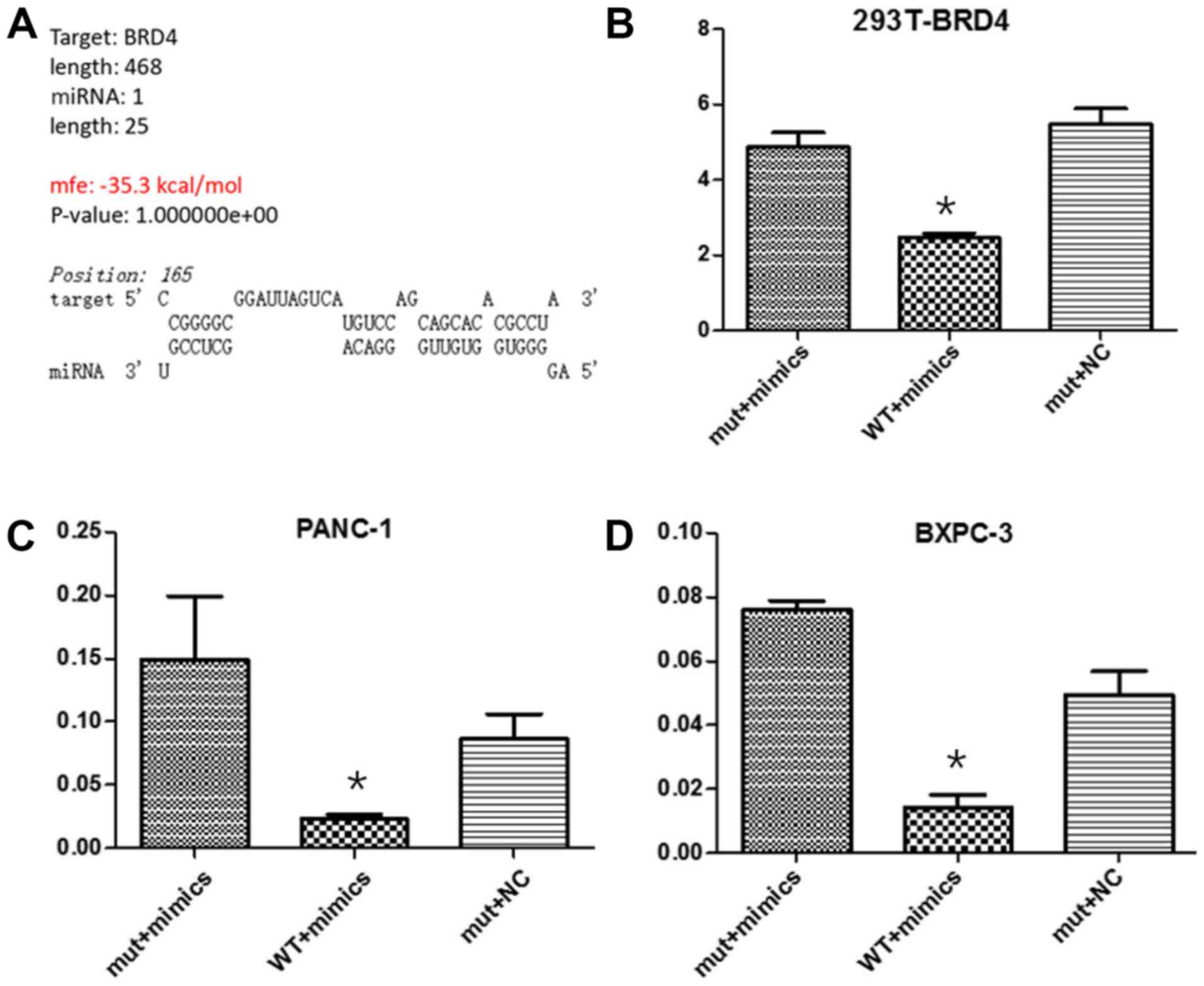 | Figure 4.miR-608 decreases the BRD4 protein
level by directly targeting the 3′-UTR. A dual-luciferase reporting
assay was performed to detect that miRNA-608 may directly inhibit
the expression of BRD4 by binding to the 3′-UTR of its mRNA in
Panc-1 and Bxpc-3 cell lines. (A) Online biological information
predicted the binding site and capacity between miRNA-608 and BRD4.
The relative luciferase activity in (B) 293T cells, (C) Panc-1
cells and (D) Bxpc-3 cells, co-transfected with miR-608 mimics and
the WT vector, were significantly inhibited, while in cells
simultaneously transfected with the Mut type vector and miR-608
mimics or NC mimics, luciferase activity was unaffected.
*P<0.05. miRNA, microRNA; NC, negative control; WT, wild type;
Mut, mutant; BRD4, bromodomain-containing protein 4; 3′-UTR,
3′-untranslated region. |
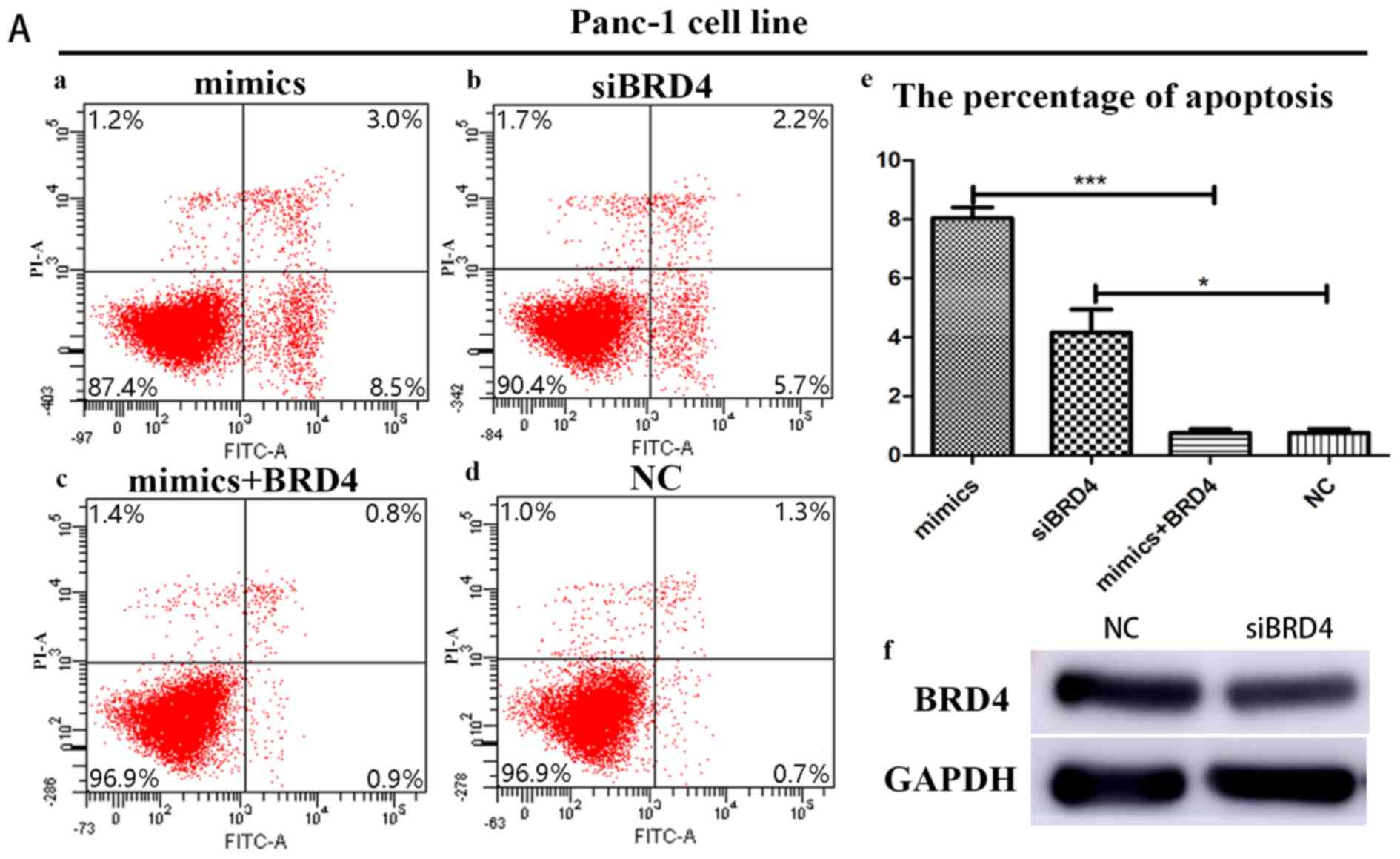
Suppression of BRD4 serves a crucial
role in miRNA-608-induced apoptosis
In order to further confirm the influence of BRD4 on
miRNA-608-induced apoptosis, Panc-1 and Bxpc-3 cells were
transfected with siBRD4 in order to specifically knock-down the
expression of BRD4. Transfection with BRD4 increased apoptosis in
both cell lines, a result consistent with the effects of miRNA-608
overexpression, using mimics. Finally, Panc-1 and Bxpc-3 cells were
transfected with both the miRNA-608 and BRD4 plasmid, and
upregulation of BRD4 decreased the miRNA-608-mediated induction of
apoptosis (Fig. 5).
Discussion
The present study demonstrated that miRNA-608 is
associated with the prognosis of patients with PDAC; a high level
of miRNA-608 expression was associated with a longer OS time. In
addition, miRNA-608 exhibited a low expression level in cancerous
tissues compared with adjacent paracancerous tissues, and the
overexpression of miRNA-608 promoted the apoptosis of pancreatic
cancer cells. Furthermore, the results of the dual-luciferase
reporter assay and bioinformatics database analysis revealed that
miRNA-608 targets the BRD4 gene, which results in the
downregulation of BRD4 expression. Finally, using a rescue assay,
it was determined that miRNA-608 serves a tumor-suppressive role
via the targeting of BRD4.
Pancreatic cancer is highly malignant and has a low
5-year OS rate, which often results in patient mortality only
several months post-diagnosis (1).
However, in TMUCIH, it was observed that in patients at the same
pathological stage, certain patients exhibited significantly longer
OS times compared with others. The present study aimed to elucidate
the causal factor behind this difference, improving the
understanding of disease progression and potentially conferring
improvements on the outcome of patients with PDAC.
Numerous studies have revealed the capacity of
miRNAs to serve as promising therapeutic targets, and to be
significant diagnostic or prognostic biomarkers in the treatment of
malignant tumors (15–17). In addition, miRNA-608 has been widely
reported to be a tumor suppressor gene and is downregulated in
multiple types of malignant tumor, including glioma, bladder,
colon, liver and lung cancer (18–22).
However, the function of miRNA-608 in pancreatic cancer progression
is yet to be determined.
In the present study, tissue samples were collected
from six patients with similar clinicopathological characteristics,
but exhibiting significantly different prognostic outcomes. The
samples were analyzed using a whole-genome expression microarray
and a total of 10 miRNAs, with significant fold-changes between
patient groups, were selected for further analyses. A limitation of
the present study was that the microarray was conducted on a small
sample population size (n=6), and that the groups did not have an
even sex distribution. Thus, future studies should be conducted on
a larger population size, with an even sex distribution.
The 10 pre-identified DEMs were verified using
another population comprising 68 patients with PDAC. qPCR analysis
revealed that miRNA-608 was associated with OS time. In addition,
miRNA-608 expression level was significantly decreased in cancerous
tissues compared with paracancerous tissues (68 vs. 11 tissues;
P<0.05; data not shown). However, the mechanism behind the
effect of miRNA-608 on pancreatic cancer is yet to be determined.
Therefore, overexpression of miRNA-608 was induced in Panc-1 and
Bxpc-3 cells in order to investigate the biological function of
miRNA-608, and it was revealed that the rate of apoptosis was
increased in the miRNA-608-overexpression group. Furthermore, it
was revealed that miR-608 also inhibited the proliferation of
pancreatic cells, but the association was not significant.
Furthermore, there was no significant association between cell
cycle stage and expression level of miR-608. Consequently, the
results concerning proliferation and cell cycle are not shown.
Subsequent experiments determined that miRNA-608 may promote
apoptosis in pancreatic cancer cells via the targeting of the
bromodomain and extra-terminal (BET) family protein, BRD4. BRD4
contains two tandem bromodomains that recognize acetylated-lysine
residues in nucleosome histones, facilitating the recruitment of
transcriptional proteins to chromatin (23). Furthermore, BRD4 may downregulate MYC
Proto-Oncogene, BHLH transcription factor (24), which is a well-characterized
apoptotic gene.
Certain studies have suggested that miRNAs regulate
gene expression by serving as molecular sponges, resulting in mRNA
degradation. This indicates that miRNA-mRNA interactions with high
degrees of complementarity result in mRNA degradation (25). Despite this, it has been hypothesized
that miRNA-mediated inhibition of expression may occur following
the initiation of translation, but not transcription. miRNA has
been revealed to decrease the rate of translation, and ribosomes
are able to prematurely terminate the translation of miRNA-targeted
mRNAs. Following miRNA-mediated mRNA translation, miRNA is often
rapidly hydrolyzed; however, the full extent of the influence of
miRNAs on protein expression levels, after the initiation of
translation, is yet to be elucidated (26). Notably, in the present study,
overexpression of miRNA-608 (via transfection with mimics in
pancreatic cancer cell lines) caused the protein level of BRD4 to
decrease significantly; however, no change in mRNA level was
observed following qPCR analysis.
In conclusion, the present study revealed that
miRNA-608 is associated with OS times in patients with pancreatic
cancer, and that a high-expression level of miRNA-608 is associated
with longer OS times. It was also discovered that miRNA-608 may
promote apoptosis in pancreatic cancer cells via the targeting of
the BET family protein, BRD4. The results indicate that miRNA-608
may represent a potential target for the treatment of human
pancreatic cancer.
Acknowledgements
Not applicable.
Funding
This study was supported by the Key Projects of
Tianjin Science and Technology Support Program (grant no.
13ZCZCSY20300).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ML performed the experiment and analyzed data. TL
interpreted the data. WM collected the samples and patients' data.
XW interpreted the clinical and survival analysis. GZ interpreted
the data, wrote and revised the manuscript. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Tianjin Medical University Cancer Institute and Hospital. All
patients or their family members were fully informed and consented
to this study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Berezikov E, van Tetering G, Verheul M,
van de Belt J, van Laake L, Vos J, Verloop R, van de Wetering M,
Guryev V, Takada S, et al: Many novel mammalian microRNA candidates
identified by extensive cloning and RAKE analysis. Genome Res.
16:1289–1298. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hata A and Lieberman J: Dysregulation of
microRNA biogenesis and gene silencing in cancer. Sci Signal.
8:re32015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stratford JK, Bentrem DJ, Anderson JM, Fan
C, Volmar KA, Marron JS, Routh ED, Caskey LS, Samuel JC, Der CJ, et
al: A six-gene signature predicts survival of patients with
localized pancreatic ductal adenocarcinoma. PLoS Med.
7:e10003072010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Collisson EA, Sadanandam A, Olson P, Gibb
WJ, Truitt M, Gu S, Cooc J, Weinkle J, Kim GE, Jakkula L, et al:
Subtypes of pancreatic ductal adenocarcinoma and their differing
responses to therapy. Nat Med. 17:500–503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Donahue TR, Tran LM, Hill R, Li Y,
Kovochich A, Calvopina JH, Patel SG, Wu N, Hindoyan A, Farrell JJ,
et al: Integrative survival-based molecular profiling of human
pancreatic cancer. Clin Cancer Res. 18:1352–1363. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
AJCC 7th Edition Cancer Staging Manual.
https://cancerstaging.org/references-tools/deskreferences/Documents/AJCC%207th%20Ed%20Cancer%20Staging%20Manual.pdfApril
10–2019
|
|
12
|
Li D, Liu H, Li Y, Yang M, Qu C, Zhang Y,
Liu Y and Zhang X: Identification of suitable endogenous control
genes for quantitative RT-PCR analysis of miRNA in bovine solid
tissues. Mol Biol Rep. 41:6475–6480. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma W, Li T, Wu S, Li J, Wang X and Li H:
LOX and ACSL5 as potential relapse markers for pancreatic cancer
patients. Cancer Biol Ther. 20:787–798. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jung M, Gelato KA, Fernández-Montalván A,
Siegel S and Haendler B: Targeting BET bromodomains for cancer
treatment. Epigenomics. 7:487–501. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sahraei M, Chaube B, Liu Y, Sun J, Kaplan
A, Price NL, Ding W, Oyaghire S, García-Milian R, Mehta S, et al:
Suppressing miR-21 activity in tumor-associated macrophages
promotes an antitumor immune response. J Clin Invest.
129:5518–5536. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Juang V, Chang CH, Wang CS, Wang HE and Lo
YL: pH-Responsive PEG-shedding and targeting peptide-modified
nanoparticles for dual-delivery of irinotecan and microRNA to
enhance tumor-specific therapy. Small. 15:e19032962019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Basu A, Jiang X, Negrini M and Haldar S:
MicroRNA-mediated regulation of pancreatic cancer cell
proliferation. Oncol Lett. 1:565–568. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liang Z, Wang X, Xu X, et al: MicroRNA-608
inhibits proliferation of bladder cancer via AKT/FOXO3a signaling
pathway. Mol Cancer. 16:962017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang H, Li Q, Niu J, Li B, Jiang D, Wan Z,
Yang Q, Jiang F, Wei P and Bai S: microRNA-342-5p and miR-608
inhibit colon cancer tumorigenesis by targeting NAA10. Oncotarget.
7:2709–2720. 2016.PubMed/NCBI
|
|
20
|
Wang Z, Xue Y, Wang P, Zhu J and Ma J:
MiR-608 inhibits the migration and invasion of glioma stem cells by
targeting macrophage migration inhibitory factor. Oncol Rep.
35:2733–2742. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang K, Liang Q, Wei L, Zhang W and Zhu P:
MicroRNA-608 acts as a prognostic marker and inhibits the cell
proliferation in hepatocellular carcinoma by macrophage migration
inhibitory factor. Tumour Biol. 37:3823–3830. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Othman N and Nagoor NH: miR-608 regulates
apoptosis in human lung adenocarcinoma via regulation of AKT2. Int
J Oncol. 51:1757–1764. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Filippakopoulos P, Qi J, Picaud S, Shen Y,
Smith WB, Fedorov O, Morse EM, Keates T, Hickman TT, Felletar I, et
al: Selective inhibition of BET bromodomains. Nature.
468:1067–1073. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Delmore JE, Issa GC, Lemieux ME, et al:
BET bromodomain inhibition as a therapeutic strategy to target
c-Myc. Cell. 146:904–917. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Seggerson K, Tang L and Moss EG: Two
genetic circuits repress the Caenorhabditis elegans heterochronic
gene lin-28 after translation initiation. Dev Biol. 243:215–225.
2002. View Article : Google Scholar : PubMed/NCBI
|