Introduction
Breast cancer is a malignant tumor with high
incidence and heterogeneity in females worldwide (1). In 2000, Perou et al (2) divided breast cancer into luminal-like,
basal-like, human epidermal growth factor receptor 2, and normal
breast-like, according to the clinical detection of molecular
markers. In addition, breast cancer can be classified into
triple-negative breast cancer (TNBC), based on immunohistochemical
techniques and hormone receptor status, which is negative for the
estrogen receptor, progesterone receptor and human epidermal growth
factor receptor 2 (HER2), and non-negative breast cancer (non-TNBC)
(3). TNBC is a specific type of
breast cancer that accounts for 15–20% of total breast cancer
cases, having unique biological characteristics and clinical
pathological features, such as not expressing Estrogen receptor
(ER), progesterone receptor (PR) and human epiderma lgrowth factor
receptor 2 (her-2) (4). TNBC is
characterized by high recurrence, early metastasis and a poor
prognosis (5,6). Considering that there is no
corresponding hormone receptor or HER2 expression, traditional
therapies, such as endocrine therapy, anti-HER2 targeted therapy
and chemotherapy, are not effective for the treatment of TNBC
(7). Therefore, there is a need to
examine the etiopathogenesis and molecular mechanism of TNBC.
Over the past 20 years, our research has focused
exclusively on surgery and chemotherapy to understand the molecular
subtypes that cause major changes in clinical practice (8). Genome-wide analysis has been extensively
used to identify important cancer genes in humans. For example,
gene expression profiles have been constructed in order to explore
a variety of changes in different types of cancer, including
colorectal, gastric and breast cancer (9–11).
Furthermore, according to reports, genetically modified core
pathways and regulatory systems become obvious in cases in which
the coding area of the genome is analyzed, as the cause of
tumorigenesis can be explained by the dysregulation of core
pathways and processes (12,13). Therefore, different signaling pathways
serve important roles in the pathogenesis of cancer. It was
observed that EGFR was closely associated with the proliferation,
invasion and vascular formation of cancer, and it was markedly
overexpressed in basal-like TNBC, which was significantly
negatively correlated with clinical prognosis (14,15). MAPK
signaling pathways regulate tumor proliferation and survival
studies have demonstrated that MAPK was abnormally activated in the
occurrence and development of TNBC, which may provide TNBC cells
with the ability to proliferate and resist apoptosis (16–18). The
PI3K/AKT/mTOR signaling pathway controls the proliferation,
metabolism, survival and movement of cancer and has frequent
communication with the MAPK signaling pathway (19–22). Other
signaling pathways, such as DNA repair, are also reportedly
dysregulated in TNBC (23).
Breast cancer susceptibility gene 1 (BRCA1) is a
well-established tumor suppressor gene, which is linked to
hereditary breast cancer (24). A
person with a BRCA1 mutation may have a 60–80% chance of developing
breast cancer, which is called BRCA1-related breast cancer
(25). According to
immunohistochemical results, the majority of BRCA1-associated
breast cancer cases are TNBC (26).
It has been suggested that these two types of tumors may have the
same or similar carcinogenic pathways (27), thus providing an important basis to
identify potential therapeutic targets. Transcription factors (TFs)
regulate DNA expression by binding to target gene promoter regions
(28). Previous studies have
recognized that there are numerous cancer-associated TFs, such as
transmembrane serine protease 2 (TMPRSS2) and ETS in prostate
cancer, and KLF4 and KLF5 in esophageal cancer; these TFs influence
the proliferation, apoptosis and invasion of cancer cells (29,30). The
transcriptome section of human TNBC has described previously
(31). However, the precise molecular
mechanisms have not been completely elucidated. It is possible to
examine the BRCA1-associated network, which may represent an
essential mechanism in facilitating TNBC metabolism, using existing
data. In the present study, differentially expressed genes (DEGs)
between TNBC and non-TNBC were ascertained, and functional
specifications including, gene ontology and signalling pathways
analysis, forecast of transcription factors and protein-protein
interaction (PPI) networks were subsequently conducted. The TFs
were later determined, and a BRCA1-connected protein-protein
interaction (PPI) network was constructed. Using this
bioinformatics knowledge, the influences of both BRCA1 and
associated signaling pathways were analyzed with respect to the
etiopathogenesis of TNBC.
Materials and methods
Affymetrix microarray data
The Affymetrix microarray data were obtained from
the National Center for Biotechnology Information Gene Expression
Omnibus database (http://www.ncbi.nlm.nih.gov/geo/) using the series
accession number, GSE27447 (32). In
total, 14 pre-treated non-triple-negative breast tumors and 5
triple-negative breast tumors were collected based on the GPL6244
(HuGene-1_0-st) Affymetrix Human Gene 1.0 ST Array.
Gene ontology and signaling pathways
impact the analysis of DEGs
The DEGs were ascertained by means of Significant
Analysis of Microarray (SAM) (33).
GO analysis determined that there were 132 upregulated and 198
downregulated DEGs (https://www.ncbi.nlm.nih.gov/geo/), applying the
Database for Annotation, Visualization, and Integrated Discovery
(DAVID; http://david.ncifcrf.gov/) (34). Using the KEGG pathway map tool, the
signaling pathways that the DEGs would likely impact were
predicted.
Forecast of transcription factors
The transcription factor target genes associated
with DEGs were evaluated by using a bioinformatics tool named TFatS
(www.tfacts.org) (35). The 132 upregulated and 198
downregulated genes were imported to TfactS to determine the
TFs.
Protein-protein interaction (PPI)
network for BRCA1
BRCA1 and DEGs were mapped to the CHARACTER STRING
database to discover the possible interacting proteins. CHARACTER
STRING (https://string-db.org/) is a repository
of forecasted protein interrelationships (36). The PPI network was subsequently
visualized by Cytoscape v3. 5.1 software.
Patient samples
Patients with breast cancer were chosen from the
pathological diagnosis of Changzheng hospital, including three TNBC
patient samples and three non-TNBC patient samples from ACKERMAN
PATHOLOGY & DIAGNOSTICS center, and total RNA was extracted
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
protocol. The present study was approved by the Ethics Committee of
Changzheng Hospital, Second Military Medical University (Shanghai,
China). Patients provided written informed consent.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
According to the instructions included in the
PrimeScript™ RT reagent Kit (Perfect Real Time) (TaKaRa Bio, Inc.,
Otsu, Japan) kit, the RT reaction was performed for the mRNA (37°C
for 15 min or 85°C for 5 sec) and miRNA (42°C for 60 min or 70°C
for 10 min). Then, they were subjected to an RT reaction using the
ABI 7900HT Fast Real-Time PCR System (Thermo Fisher Scientific,
Inc.). The miRNA primers were designed by Guangzhou RiboBio Co.,
Ltd (Guangzhou, China). The fold-changes for miRNA and mRNA
expression were calculated by the 2−ΔΔCq method. The
amplification primers of CR2, PRKCB, CARD11, PLCG2, CD79A, CD27,
STARD3, SRD5A3, CACNA1H, UGT2B4, SDR16C5 and MED1 were then
designed (Table I).
 | Table I.Primers used in RT-qPCR. |
Table I.
Primers used in RT-qPCR.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| CR2 |
GGCTACCTTATGGCTGGAGAG |
AGAGTCACAGTAGTCCCAAACC |
| PRKCB |
GCCTACCCCAAGGTCCATGT |
CTTGGTCATGAGCCCTTTG |
| CARD11 |
CAGGGTGCCTGCCTCATAG |
TATAGGGAGAAGCAAGGCAGGG |
| PLCG2 |
CTGGCAACCGACTCAAAGGA |
GCTGATGCTGTTTCTTCGGG |
| CD79A |
CTACGGCTTCTCCAGCTGAAT |
CAGCTGAATGTCTTCCTCACA |
| CD27 |
ATGGAAAGGGAAGCACGTC |
TTGGCCAACTCCTCTCCTAA |
| STARD3 |
GACCTGGTTCCTTGACTTCAA |
CGGCAAGACGTTTATCCTGAA |
| SRD5A3 |
TTTAATCAGGCCCTGTCTGC |
GGGGTATAGAAATGGAATGGAGA |
| CACNA1H |
GGATCCCAAGCTTGGTACCG |
CTTCATGGCCCTCTAGAGGATCC |
| UGT2B4 |
TGGACTCATCACCTGACTCATGTAA |
GTCAAAGAGACTGCAGGAACATGA |
| SDR16C5 |
TGGAAACTCTTAAAGTTTTGCTCCTTACAATC |
GAAAGATATTCATGGTGAATTCGAATC |
| MED1 |
GAGACTCCGCCCACTTACCTG |
GGACACACTTCAAACTGGAGG |
| GAPDH |
ACGGCAAGTTCAACGGCACAG |
CCACGACATACTCAGCACCAGC |
Western blot analysis
Patient sample was homogenized in a lysis buffer
(P0013; Beyotime Institute of Biotechnology, Haimen, China)
supplemented with 1% protease inhibitor cocktail (Pierce; Thermo
Fisher Scientific, Inc.). Antibodies against the following proteins
were used for Western blot analysis: MYC (1:1,000 dilution; no.
#2272; Cell Signalling Technology, Inc., Danvers, MA, USA), SP1
(1:1,000 dilution; no. #5931; Cell Signalling Technology, Inc.,),
β-actin (1:1,000 dilution; no. #4967; Cell Signalling Technology,
Inc.,) and CTNNB1 (1:1,000 dilution; Phospho-β-Catenin (Ser33/37)
Antibody no. #2009; Cell Signalling Technology, Inc.). Following
centrifugation at 12,000 × g for 30 min at 4°C, the supernatant was
collected, and the protein concentration was determined using an
enhanced bicinchoninic acid protein assay kit according to the
manufacturer's protocol (Nanjing Jiancheng Bioengineering
Institute, Nanjing, China). The membrane proteins were transferred
to a polyvinylidene difluoride (PVDF) membrane following
electrophoresis with 10% SDS-PAGE. The membranes were blocked with
5% non-fat dry milk in Tris-buffered saline prior to overnight
incubation at 4°C, incubated with the primary antibodies and were
then incubated with the secondary antibodies (Abmart, Shanghai
China) for 60 min at 37°C. Following washing with TBST 3 times for
10 min each, the membranes were developed with an enhanced
chemiluminescent ECL assay kit (Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA).
Statistical analysis
Data are presented as the mean ± standard error of
the mean. The primary data were formatted into expression
measurements and normalized by the robust multi-array average (RMA)
algorithm (23). The criteria for
screening DEGs were |logFC|>1.5 and the false discovery rate,
(FDR) <0.05. An unpaired student's t-test was used to analyze
data between two groups. P<0.05 was considered to indicate a
statistically significant difference. All analyses were performed
using SPSS v.22.0 (IBM Corp., Armonk, NY, USA).
Results
Discernment and GO analysis of
DEGs
On the basis of the SAM analysis, a total of 132
upregulated and 198 downregulated DEGs were identified. The GO
analysis was subsequently conducted (Table II). The results demonstrated that the
upregulated DEGs, which included CR2, IGHM, PRKCB, CARD11, PLCG2,
CD79A, IGKC and CD27, were relative to the immune response, such as
lymphocyte activation (P=1.49×10−11), leukocyte
activation (P=4.68×10−11) and B-cell activation (P=6.02×
10−8) (Table IIA). The
downregulated DEGs, which included STARD3, ALDH8A1, SRD5A3,
CACNA1H, UGT2B4, SDR16C5 and MED1, were primarily enriched in the
hormone metabolic process (P=5.45×10−5) (Table IIB).
| Table II.GO terms of DEGs. |
Table II.
GO terms of DEGs.
| A, The top 10 GO
terms of the 132 upregulated DEGs |
|---|
|
|---|
| Category | Term | Count | P-value | Genes |
|---|
| GO:0046649 | Lymphocyte
activation | 24 |
1.49×10−11 | GAPT, TCF7, CR2,
IGHM |
| GO:0045321 | Leukocyte
activation | 25 |
4.68×10−11 | GAPT, TCF7, CR2,
IGHM |
| GO:0002684 | Positive regulation
of immune system process | 27 |
2.66×10−10 | BLK, CR2, IGHM,
PRKCB |
| GO:0050778 | Positive regulation
of immune response | 23 |
3.37×10−10 | CR2, IGHM, PRKCB,
CARD11 |
| GO:0006955 | Immune
response | 33 |
1.46×10−9 | IL16, BLK, PAX5,
GPRC5B |
| GO:0002376 | Immune system
process | 42 |
2.11×10−9 | IL16, BLK, PAX5,
GPRC5B |
| GO:0001775 | Cell
activation | 25 |
2.52×10−9 | GAPT, TCF7, CR2,
IGHM |
| GO:0002682 | Regulation of
immune system process | 30 |
6.97×10−9 | BLK, CR2, IGHM,
PRKCB |
| GO:0050776 | Regulation of
immune response | 24 |
1.34×10−8 | CR2, IGHM, PRKCB,
CARD11 |
| GO:0042113 | B cell
activation | 13 |
6.02×10−8 | GAPT, CR2, IGHM,
PRKCB |
|
| B, Top 10 GO
terms of the 198 downregulated DEGs |
|
|
Category | Term | Count | P-value | Genes |
|
| GO:0042445 | Hormone metabolic
process | 10 |
5.45×10−5 | STARD3, TG,
ALDH8A1, FOXA1 |
| GO:0044699 | Single-organism
process | 143 |
4.38×10−4 | PDP1, ALDH8A1,
ADCY1, TSPAN1 |
| GO:0034754 | Cellular hormone
metabolic process | 7 |
5.22×10−4 | STARD3, ALDH8A1,
SRD5A3, CACNA1H |
| GO:0010817 | Regulation of
hormone levels | 14 |
5.62×10−4 | ALDH8A1, TG, FOXA1,
IYD |
| GO:0009952 | Anterior/posterior
pattern specification | 9 |
6.17×10−4 | HOXC10, MSX2,
PCGF2, HOXC11 |
KEGG pathways analysis
On the basis of the SPIA analysis, a total of 13
KEGG signaling pathways were examined to determine if they were
dysregulated in TNBC (Table III).
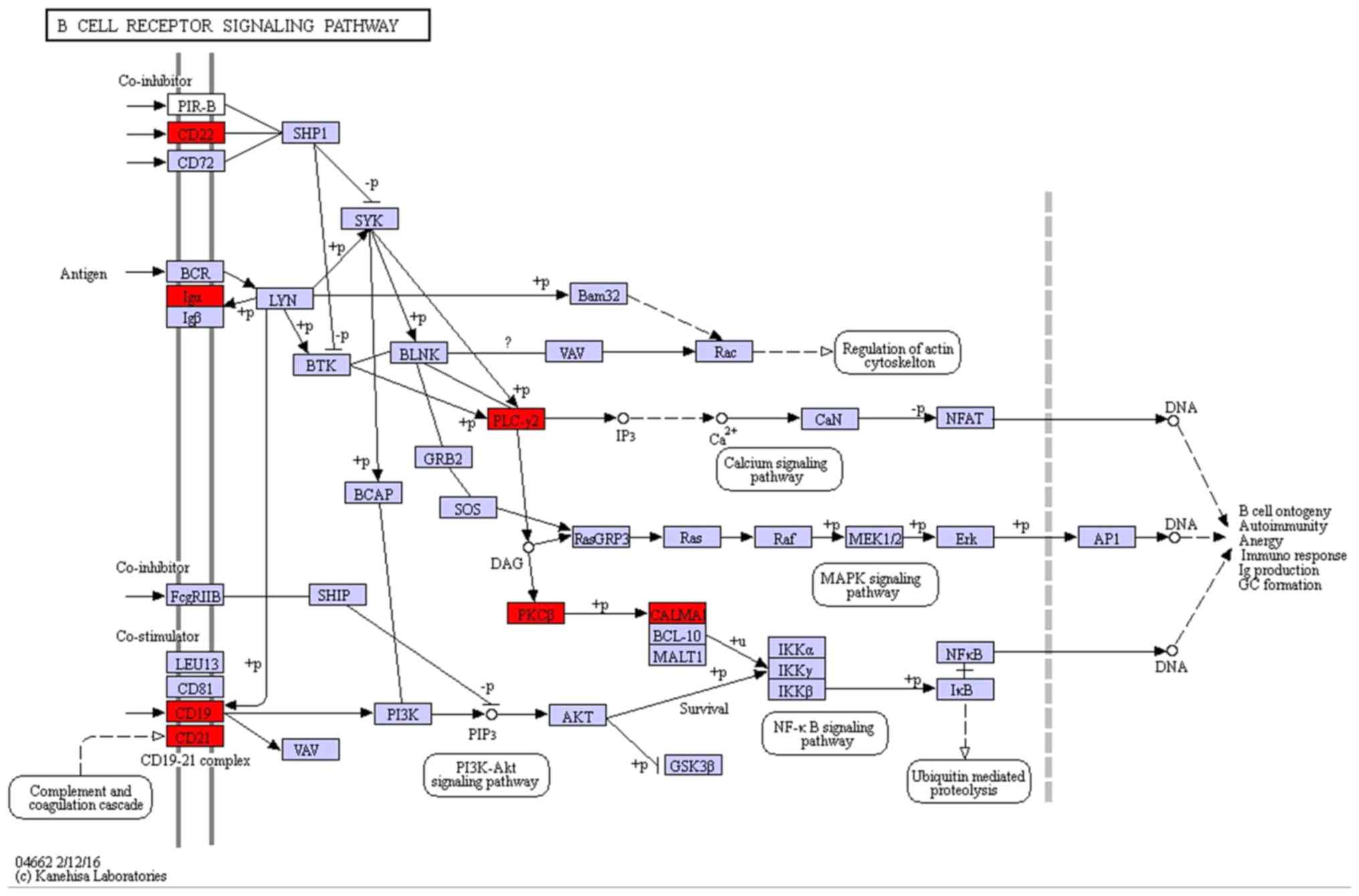
The B cell receptor signaling pathway (P=3.99×10−3)
(Fig. 1), hormone synthesis signaling
pathway (P=2.21×10−2) and oxytocin signaling pathway
(P=3.41×10−2) were each identified to exhibit
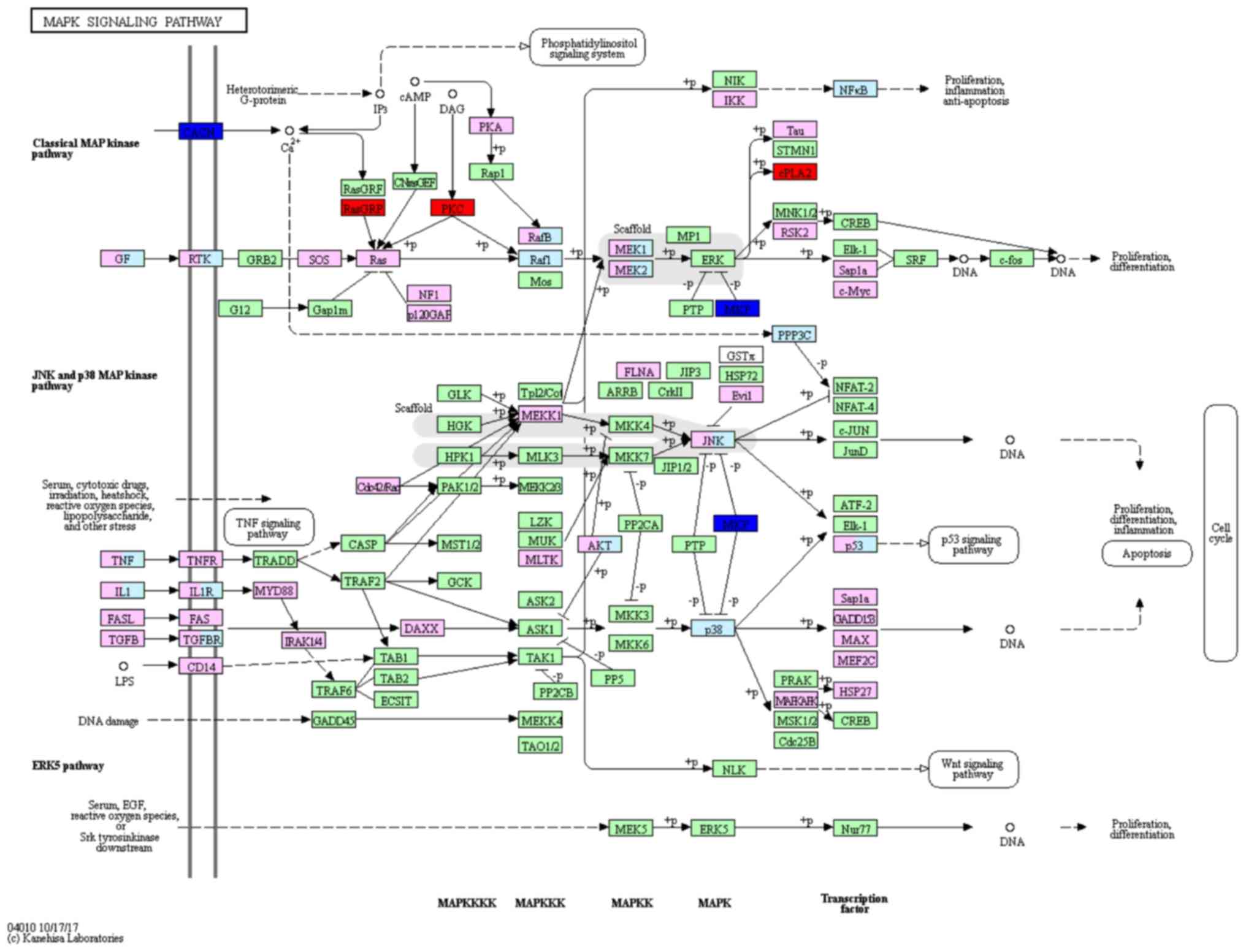
significant differences. MAPK/RAS and mTOR signaling pathways that
correlated with cancer were selected for subsequent investigation.
Certain protein kinase genes, such as GRP, PKC and CPLA2, were
upregulated, and MKP and CACN were downregulated, in the MAPK
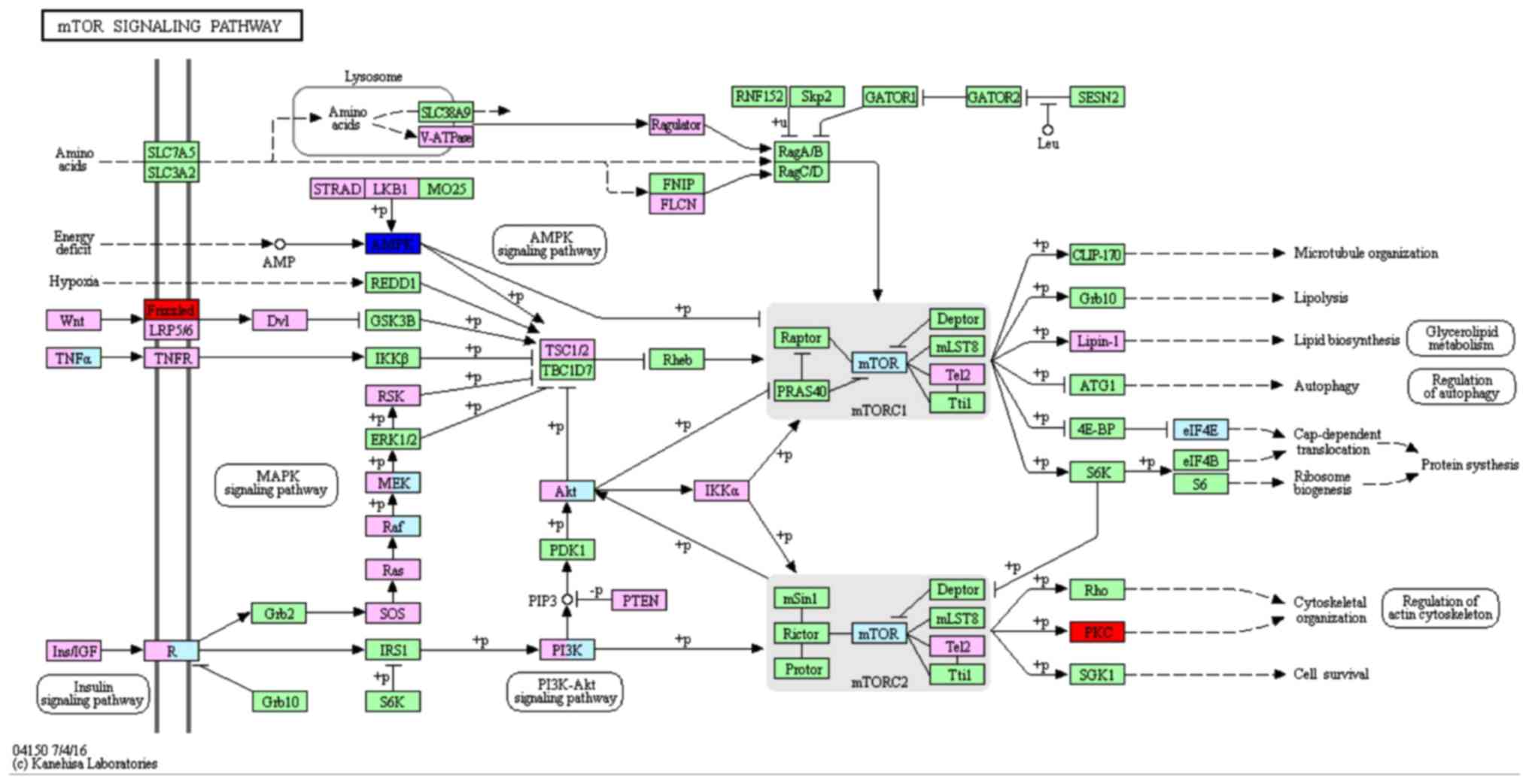
signaling pathway (Fig. 2). In the
mTOR signaling pathway, PKC was upregulated and AMPK was
downregulated (Fig. 3).
| Table III.Fourteen pathways identified based on
KEGG. |
Table III.
Fourteen pathways identified based on
KEGG.
| Pathway | Count | P-value | Genes |
|---|
| B cell receptor
signaling pathway | 6 | 0.003988 | CARD11, CD19, CR2,
PLCG2, CD22, CD79A |
| Hormone
synthesis | 5 | 0.022133 | TG, ADCY1, CREB3L4,
PRKCB, IYD |
| Oxytocin signaling
pathway | 7 | 0.034094 | ADCY1, PLA2G4A,
RGS2, RYR3, CACNG4, PRKAA2, PRKCB |
| Hematopoietic cell
lineage | 5 | 0.041129 | CD19, CR2, CD3E,
MS4A1, CD22 |
| Platelet
activation | 6 | 0.049048 | ADCY1, PLA2G4A,
FGA, RASGRP1, PLCG2, RASGRP2 |
| Non-small cell lung
cancer | 4 | 0.054172 | ERBB2, PLCG2, RARB,
PRKCB |
| Calcium signaling
pathway | 7 | 0.056584 | ADCY1, CHRM3,
ERBB2, RYR3, PLCG2, CACNA1H, PRKCB |
| Wnt signaling
pathway | 6 | 0.060444 | TCF7, DKK1, SFRP1,
WIF1, FZD7, PRKCB |
| Inflammatory
mediator regulation of TRP channels | 5 | 0.063253 | ADCY1, PLA2G4A,
P2RY2, PLCG2, PRKCB |
| Melanogenesis | 5 | 0.067119 | ADCY1, TCF7,
CREB3L4, FZD7, PRKCB |
| Circadian
rhythm | 3 | 0.081308 | RORC, PRKAA2,
BHLHE41 |
| Primary
immunodeficiency | 3 | 0.095274 | CD19, CD3E,
CD79A |
| MAPK signaling
pathway | 8 | 0.096136 | DUSP4, PLA2G4A,
RASGRP1, RASGRP2, DUSP10, CACNG4, CACNA1H, PRKCB |
Transcription factor-regulated
DEGs
TFactS analysis was performed to ascertain
transformation in the degree of transcription factor activity in
upregulated and downregulated genes in TNBC (Table IV). The results indicated that MYC,
SP1 and CTNNB1 were stimulated in TNBC. In total, 14 target genes
of CTNNB1 were identified, including 5 upregulated and 9
downregulated genes, while for SP1, 3 target genes were upregulated
and 6 genes were downregulated. Additionally, 4 target genes of MYC
were upregulated and 2 genes were downregulated.
| Table IV.Results of the TfactS analysis. |
Table IV.
Results of the TfactS analysis.
| Gene name | TF | Regulation |
|---|
| WIF1 | CTNNB1 | Up |
| FZD7 | CTNNB1 | Up |
| QPCT | CTNNB1 | Up |
| TCF7 | CTNNB1 | Up |
| SOX10 | CTNNB1 | Up |
| INHBB | CTNNB1 | Down |
| SEMA3C | CTNNB1 | Down |
| DKK1 | CTNNB1 | Down |
| MSX2 | CTNNB1 | Down |
| AR | CTNNB1 | Down |
| CEACAM6 | CTNNB1 | Down |
| MUC6 | CTNNB1 | Down |
| SCGB2A2 | CTNNB1 | Down |
| CEACAM5 | CTNNB1 | Down |
| CD19 | SP1 | Up |
| UGT8 | SP1 | Up |
| PLA2G4A | SP1 | Up |
| PAPSS2 | SP1 | Down |
| KRT19 | SP1 | Down |
| AR | SP1 | Down |
| CEACAM5 | SP1 | Down |
| FOXA1 | SP1 | Down |
| SLC9A2 | SP1 | Down |
| RARB | MYC | Up |
| SPIB | MYC | Up |
| PLA2G4A | MYC | Up |
| RGS2 | MYC | Up |
| RPL23 | MYC | Down |
| CEACAM5 | MYC | Down |
Verifying the expression of DEGs and
transcription factors
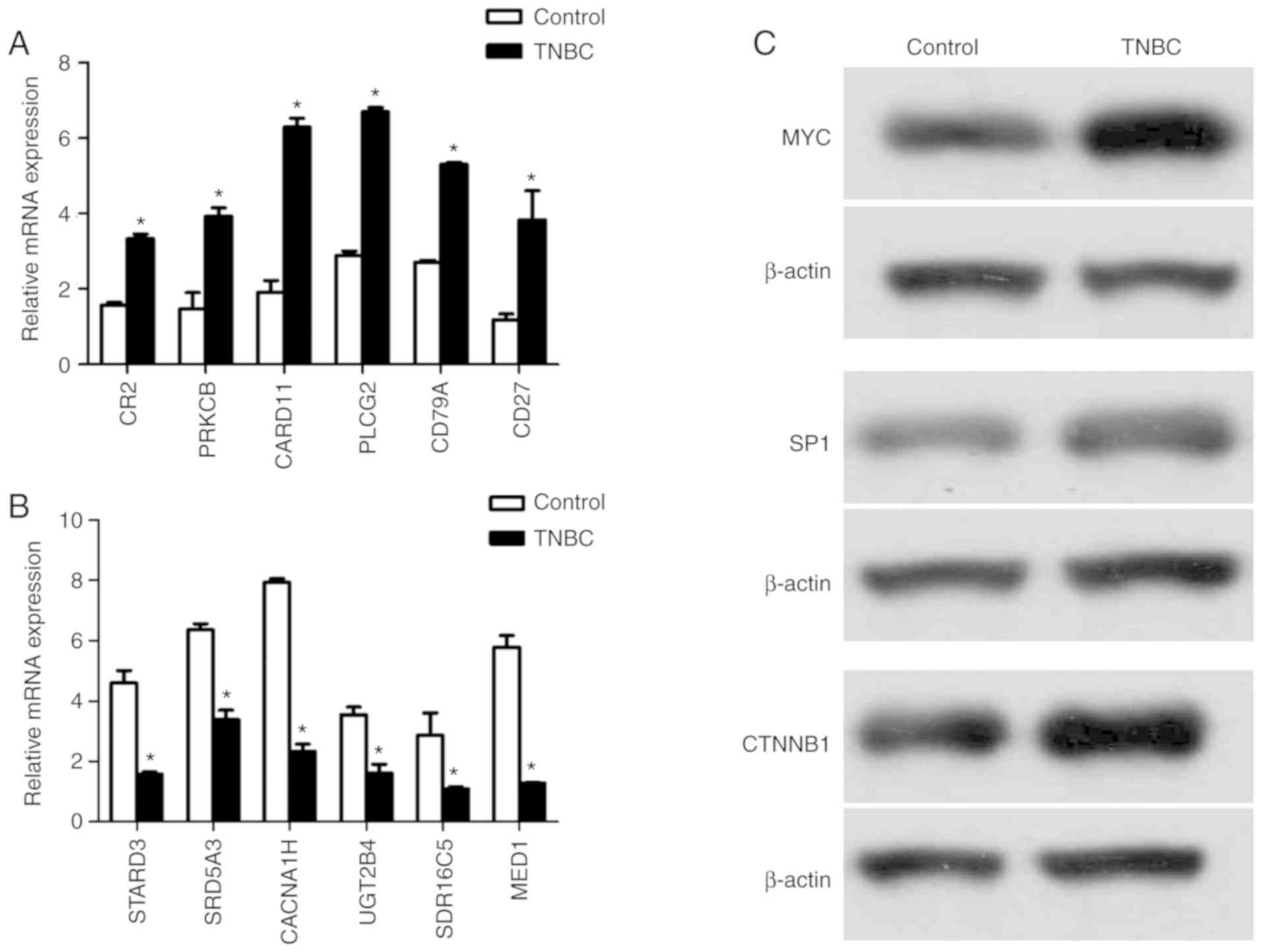
The RT-qPCR results demonstrated that the expression
of CR2, PRKCB, CARD11, PLCG2, CD79A and CD27 was increased, and the
expression of STARD3, SRD5A3, CACNA1H, UGT2B4, SDR16C5 and MED1 was
decreased in TNBC-patient samples (Fig.
4A and B). The western blotting results revealed that the
expression of MYC, SP1 and CTNNB1 was increased in TNBC-patient
samples (Fig. 4C).
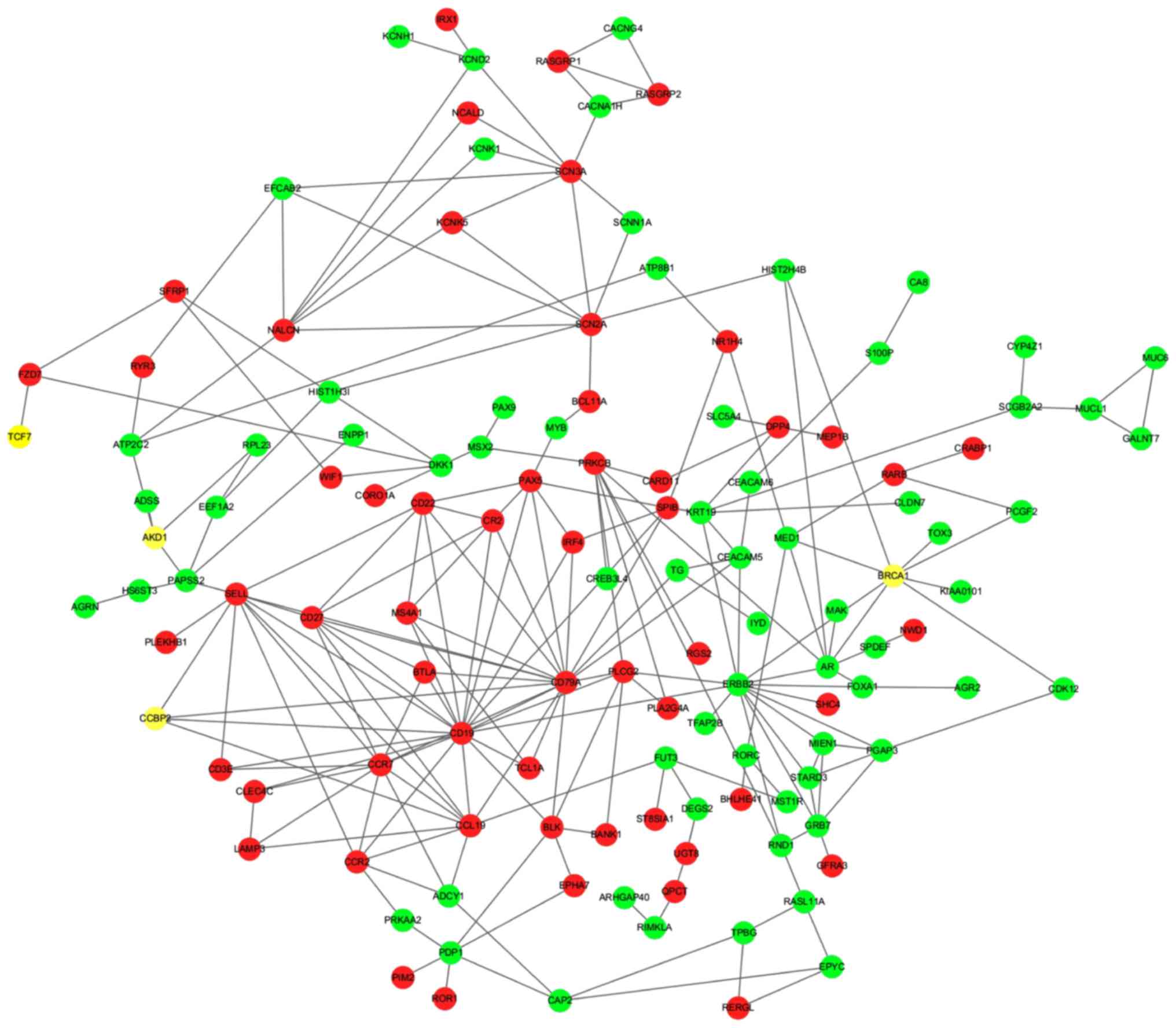
BRCA1 associated with PPI network
Metastasis is the main cause of cancer-related death
(27). Mutations in BRCA1, one of
nucleoprotein species, have been discovered to improve
tumorigenesis (23,24). To examine the role of BRCA1 in TNBC, a
BRCA1-related PPI network was constructed (Fig. 5). The results demonstrated that 8
proteins, AR, CDK12, KIAA0101, PCGF2, TOX3, HIST2H4B, MED1 and
ERBB2, which were encoded by downregulated genes, interacted with
BRCA1, while no proteins encoded by upregulated genes interacted
with BRCA1. Additionally, an important interaction with BRCA1 in
the expression levels of the androgen receptor (AR) was
observed.
Discussion
Breast cancer is the most common type of cancer in
females worldwide, and is characterized by a high mortality rate
(37,38). TNBC is the most dangerous type of
breast cancer and is characterised by high recurrence, early
metastasis and a poor prognosis (5,6). Although
it has been the focus of numerous studies, the potential mechanisms
of TNBC have not been completely elucidated. In the present study,
the DEGs of TNBC and non-TNBC samples were identified, and the
functions were predicted through GO analysis and signaling
pathways. A PPI network associated with BRCA1 was also identified.
Taken together the BRAC1-associated interactions offer novel
insights into the pathogenesis of TNBC.
GO analysis revealed that the upregulated genes were
primarily concentrated in the immune response, including lymphocyte
activation, cell cycle progression, leukocyte activation and B cell
activation. Complement receptor type 2 (CR2; CD21), which binds
fragments of C3, may ligate CD19 or CD23 to present antigens to B
cells (39). This outcome can
facilitate the interaction of B cells with other cells essential
for cellular activation (39). CR2
also serves an important role in enhancing humoral immunity to
T-dependent and T-independent foreign antigens, and in regulating
T-cell immunity to self and non-self-antigens. CR2 is an important
receptor that amplifies B lymphocyte activation by bridging the
innate and adaptive immune systems (40). Variations or deletions of the CR2 gene
in humans are associated with a variety of autoimmune and
inflammatory conditions, including but not limited to rheumatoid
arthritis, systemic lupus erythmatosus, psoriatic arthritis,
reactive arthritis, Crohn's disease, ulcerative colitis and
scleroderma (40). In aggressive
adult T-cell leukemia/lymphoma, a multivariate analysis
incorporating clinical factors and genetic alterations demonstrated
that being at high-risk, including patient age, the presence of
PRKCB mutations and PD-L1 amplifications, were all independent poor
prognosticators listed in the Japan Clinical Oncology Group
prognostic index (41). Through
pathway analysis, these upregulated genes closely associated with
immune related pathways, such as the B cell receptor-signalling
pathway. Conversely, a possible target for further clinical
verification is conducive to TNBC immunotherapy.
The results of the present study revealed that
downregulated genes, including ALDH8A1 and SRD5A3, in TNBC were
significantly concentrated in the hormone metabolic biological
process. Retinoic acid (RA) is required for cellular
differentiation and is known to arrest tumor development (42). RA is synthesized from retinaldehyde by
retinaldehyde dehydrogenases, specifically ALDH1A1, ALDH1A2,
ALDH1A3 and ALDH8A1 (42). ALDH8A1 is
considered to be a candidate marker to identify and isolate normal
and cancer stem cells (42). In
addition, SRD5A3 is associated with the production of 5
α-dihydrotestosterone and the activation of the androgen-androgen
receptor-pathway is a promising molecular target for prostate
cancer therapy (43).
Signaling pathways serve an important role in the
investigation of cancer pathogenesis (44). According to the latest research, CD19,
CD22 and CD79A were concentrated in the B cell receptor-signaling
pathway (44–46). These genes encode a cell surface
molecule that assembles with the antigen receptor of B-lymphocytes
in order to decrease the threshold for antigen receptor-dependent
stimulation (44–46). In there present study, these DEGs were
closly associated with potential therapeutic targets for TNBC.
Previous study (47) has demonstrated
that the most prominent signaling pathway that is dysregulated in
TNBC is the PI3K/AKT/mTOR signaling pathway. PKC (protein kinase C)
is an intracellular effector of the C kinase pathway. PKC is
associated with the mTOR signaling pathway, which is involved in
the regulation of metabolism and gene expression, including
serine/threonine residue phosphorylation, cell proliferation and
nuclear type changes (48). It has
been previously demonstrated that the attenuation of the mTOR
signaling pathway presents an obvious curative possible for the
diagnosis of cancer. The present study provides detailed insights
into the novel cytotoxic mechanism of an anti-tumor compound
originated from the herbal plant, which may be useful in promoting
autophagy mediated- cell death in cancer cell that is resistant to
apoptosis. Mechanistically, tetrandrine induces autophagy that
depends on mTOR inactivation (49).
The results of the present study indicated that the B-cell receptor
signaling pathway by enriched genes was differentially significant,
when compared with the PI3K/AKT/mTOR signaling pathway in TNBC.
Transcription factors are targets for certain
anticancer drugs; however, a limited list of transcription factors
are overactive in most human cancer cells, which makes them targets
for the development of anticancer drugs. That they are the most
direct and hopeful targets for treating cancer is proposed, and
this is supported by the fact that there are many more human
oncogenes in signalling pathways than there are oncogenic
transcription factors (50). NF-κB,
is associated with control of cell cycle, apoptosis, migration and
cell differentiation, and is one of the most appropriate
transcription factors as a target for the development of cancer
immunotherapeutics (51,52). HIF-1α and STAT have also been reported
to be involved in the development of cancer therapeutics (53,54).
Specific transcription factors including, MYC, SP1 and CTNNB1, have
been demonstrated to function as regulators of certain DEGs
(55). In the Wnt/APC/β-catenin
signaling pathway in colorectal cancer, the activation and
overexpression of MYC, marked as a downstream step, is crucial for
tumour metastasis (55,56). Sp1, a stress-responsive factor,
inhibits the sensitivity of cancer cells to chemotherapy by
regulating stemness gene expression in glioblastoma (57). CTNNB1 is the key regulatory factor of
the Wnt/β-catenin signaling pathway, and the overexpression pattern
promotes the growth of hepatocellular carcinoma cells by targeting
CD44 and c-Myc protein (58). These
findings may help to develop drugs for tumor therapy.
The present study has helped to elucidate the
molecular mechanisms of tumor development and metastasis. The BRCA1
mutation is associated with neoplastic transformation (59), and the results of the present study
suggest that BRCA1 may interact with DEGs to participate in the
occurrence of TNBC. Among the mass of genes associated with BRCA1,
the interaction with the androgen receptor (AR) is significant.
Several lines of evidence suggest that androgen may have direct
effects on breast cancer cells, by binding to their selective
androgen receptors (ARs) (60).
Therefore, in the case of oestrogen receptor negative, evidence has
revealed that TNBC is associated with the hormone pathway. The
network of BRCA1 was predicated in order to help elucidate the
mechanism of tumor cell metastasis, and provide therapeutic targets
for the development of cancer therapeutics. However, there
limitations to the present research, as all differentially
expressed genes and transcription factors did not undergo RT-qPCR
and western blotting for further verification.
The present study demonstrated that the B cell
receptor-signaling pathway and hormone synthesis-signaling pathway
are of vital importance in the development of TNBC. In addition,
numerous genes that may function as potential targets for TNBC have
been identified. Nevertheless, to reveal TNBC's potential molecular
mechanisms, further research should be performed to investigate
other signaling pathways and key cancer-associated proteins.
Acknowledgements
The authors would like to thank Ackerman Pathology
and Diagnostics Center (Shanghai China) for providing specimen
tissues.
Funding
The present study was funded by Chinese National
Science Foundation (grant no. 81502260, 2016).
Availability of data and materials
The Affymetrix microarray data were obtained from
the National Center for Biotechnology Information Gene Expression
Omnibus database (http://www.ncbi.nlm.nih.gov/geo/) using the series
accession number, GSE27447. Patient samples and three non-TNBC
patient samples from ACKERMAN PATHOLOGY & DIAGNOSTICS
Centre.
Authors' contributions
FQ and WXQ performed the majority of experiments;
FQ, WXQ and YSZ provided vital reagents and analytical tools and
were also involved in editing the manuscript; FQ and WXQ designed
the study and wrote the manuscript. YSZ analyzed and interpreted
the data, and agreed to be accountable for all aspects of the work
in ensuring that question related to the accuracy or integrity of
any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Changzheng Hospital, Second Military Medical
University (Shanghai, China). Patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chu J, Bae H, Seo Y, Cho SY, Kim SH and
Cho EY: The prognostic impact of synchronous ipsilateral multiple
breast cancer: Survival Outcomes according to the Eighth American
Joint Committee on cancer staging and molecular subtype. J Pathol
Transl Med. 52:396–403. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Perou CM, Sørlie T, Eisen MB, van de Rijn
M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA,
et al: Molecular portraits of human breast tumours. Nature.
406:747–752. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zubeda S, Kaipa PR, Shaik NA, Mohiuddin
MK, Vaidya S, Pavani B, Srinivasulu M, Latha MM and Hasan Q:
Her-2/neu status: A neglected marker of prognostication and
management of patients with breast cancer in India. Asian Pac J
Cancer Prev. 14:2231–2235. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bauer KR, Brown M, Cress RD, Parise CA and
Caggiano V: Descriptive analysis of estrogen receptor
(ER)-negative, progesterone receptor (PR)-negative, and
HER2-negative invasive breast cancer, the so-called triple-negative
phenotype: A population-based study from the California cancer
Registry. Cancer. 109:1721–1728. 2010. View Article : Google Scholar
|
|
5
|
Haque R, Ahmed SA, Inzhakova G, Shi J,
Avila C, Polikoff J, Bernstein L, Enger SM and Press MF: Impact of
breast cancer subtypes and treatment on survival: An analysis
spanning two decades. Cancer Epidemiol Biomarkers Prev.
21:1848–1855. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Carey LA, Perou CM, Livasy CA, Dressler
LG, Cowan D, Conway K, Karaca G, Troester MA, Tse CK, Edmiston S,
et al: Race, breast cancer subtypes, and survival in the Carolina
Breast Cancer Study. JAMA. 295:2492–2502. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bosch A, Eroles P, Zaragoza R, Viña JR and
Lluch A: Triple-negative breast cancer: Molecular features,
pathogenesis, treatment and current lines of research. Cancer Treat
Rev. 36:206–215. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stratton MR, Campbell PJ and Futreal PA:
The cancer genome. Nature. 458:719–724. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lanza G, Ferracin M, Gafà R, Veronese A,
Spizzo R, Pichiorri F, Liu CG, Calin GA, Croce CM and Negrini M:
mRNA/microRNA gene expression profile in microsatellite unstable
colorectal cancer. Mol Cancer. 6:542007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jones S, Zhang X, Parsons DW, Lin JC,
Leary RJ, Angenendt P, Mankoo P, Carter H, Kamiyama H, Jimeno A, et
al: Core signaling pathways in human pancreatic cancers revealed by
global genomic analyses. Science. 321:1801–1806. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lustig B and Behrens J: The Wnt signaling
pathway and its role in tumor development. J Cancer Res Clin Oncol.
129:199–221. 2003.PubMed/NCBI
|
|
12
|
Cancer Genome Atlas Network: Comprehensive
molecular portraits of human breast tumours. Nature. 490:61–70.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang X and Sommer RJ: Antagonism of
LIN-17/Frizzled and LIN-18/Ryk in nematode vulva induction reveals
evolutionary alterations in core developmental pathways. PLoS Biol.
9:e10011102011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Williams CB, Soloff AC, Ethier S and Yeh
ES: Perspectives on epidermal growth factor receptor regulation in
triple-negative breast cancer: Ligand-mediated mechanisms of
receptor regulation and potential for clinical targetin. Adv Cancer
Res. 127:253–281. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tao JJ, Castel P, Radosevic-Robin N,
Elkabets M, Auricchio N, Aceto N, Weitsman G, Barber P, Vojnovic B,
Ellis H, et al: Antagonism of EGFR and HER3 enhances the response
to inhibitors of the PI3K-Akt pathway in triple-negative breast
cancer. Sci Signal. 7:ra292014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Duncan JS, Whittle MC, Nakamura K, Abell
AN, Midland AA, Zawistowski JS, Johnson NL, Granger DA, Jordan NV,
Darr DB, et al: Dynamic reprogramming of the kinome in response to
targeted MEK inhibition in triple-negative breast cancer. Cell.
149:307–321. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee J, Galloway R, Grandjean G, Jacob J,
Humphries J, Bartholomeusz C, Goodstal S, Lim B, Bartholomeusz G,
Ueno NT and Rao A: Comprehensive two- and three-dimensional RNAi
screening identifies PI3K inhibition as a complement to MEK
inhibitor AS703026 for combination treatment of triple-negative
breast cancer. J Cancer. 6:1306–1319. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim S, Lee J, Jeon M, Lee JE and Nam SJ:
MEK-dependent IL-8 induction regulates the invasiveness of
triple-negative breast cancer cells. Tumour Biol. 37:4991–4999.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Aksamitiene E, Kiyatkin A and Kholodenko
BN: Cross-talk between mitogenicRas/MAPK and survival PI3K/Akt
pathways: A fine balance. Biochem Soc Trans. 40:139–146. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fruman DA and Rommel C: PI3K and cancer:
Lessons, challenges and opportunities. Nat Rev Drug Discov.
13:140–156. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ganesan P, Moulder S, Lee JJ, Janku F,
Valero V, Zinner RG, Naing A, Fu S, Tsimberidou AM, Hong D, et al:
Triple-negative patients with breast cancer treated at MD Anderson
cancer center in phase I trials: Improved outcomes with combination
chemotherapy and targeted agents. Mol Cancer Ther. 13:3175–3184.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Juvekar A, Burga LN, Hu H, Lunsford EP,
Ibrahim YH, Balmañà J, Rajendran A, Papa A, Spencer K, Lyssiotis
CA, et al: Combining a PI3K inhibitor with a PARP inhibitor
provides an effective therapy for BRCA1-related breast cancer.
Cancer Discov. 2:1048–1063. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Murai J, Huang SN, Das BB, Renaud A, Zhang
Y, Doroshow JH, Ji J, Takeda S and Pommier Y: Trapping of PARP1 and
PARP2 by clinical PARP inhibitors. Cancer Res. 72:5588–5599. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Paul A and Paul S: The breast cancer
susceptibility genes (BRCA) in breast and ovarian cancers. Front
Biosci (Landmark Ed). 19:605–618. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tischkowitz MD and Foulkes WD: The basal
phenotype of BRCA1-related breast cancer: Past, present and future.
Cell Cycle. 5:963–967. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He BS, Pan YQ, Lin K, Ying HQ, Wang F,
Deng QW, Sun HL, Gao TY and Wang SK: Evaluation the susceptibility
of five polymorphisms in microRNA-binding sites to female breast
cancer risk in Chinese population. Gene. 573:160–165. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang L, Fang C, Xu X, Li A, Cai Q and
Long X: Androgen receptor, EGFR, and BRCA1 as biomarkers in
triple-negative breast cancer: A meta-analysis. Biomed Res Int.
2015:3574852015.PubMed/NCBI
|
|
28
|
Sankpal UT, Goodison S, Abdelrahim M and
Basha R: Targeting Sp1 transcription factors in prostate cancer
therapy. Med Chem. 7:518–525. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tomlins SA, Rhodes DR, Perner S,
Dhanasekaran SM, Mehra R, Sun XW, Varambally S, Cao X, Tchinda J,
Kuefer R, et al: Recurrent fusion of TMPRSS2 and ETS transcription
factor genes in prostate cancer. Science. 310:644–648. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang Y, Goldstein BG, Chao HH and Katz JP:
KLF4 and KLF5 regulate proliferation, Apoptosis and invasion in
esophageal cancer cells. Cancer Biol Ther. 4:1216–1221. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang L, Wu X, Wang Y, Zhang K, Wu J, Yuan
YC, Deng X, Chen L, Kim CC, Lau S, et al: FZD7 has a critical role
in cell proliferation in triple negative breast cancer. Oncogene.
30:4437–4446. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tusher VG, Tibshirani R and Chu G:
Significance analysis of microarrays applied to the ionizing
radiation response. Proc Natl Acad Sci USA. 98:5116–5121. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2008. View Article : Google Scholar
|
|
34
|
Essaghir A, Toffalini F, Knoops L, Kallin
A, van Helden J and Demoulin JB: Transcription factor regulation
can be accurately predicted from the presence of target gene
signatures in microarray gene expression data. Nucleic Acids Res.
38:e1202010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
von Mering C, Huynen M, Jaeggi D, Schmidt
S, Bork P and Snel B: STRING: A database of predicted functional
associations between proteins. Nucleic Acids Res. 31:258–261. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fan L, Strasser-Weippl K, Li JJ, St Louis
J, Finkelstein DM, Yu KD, Chen WQ, Shao ZM and Goss PE: Breast
cancer in China. Lancet Oncol. 15:e279–e289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fearon DT and Carter RH: The
CD19/CR2/TAPA-1 complex of B lymphocytes: Linking natural to
acquired immunity. Annu Rev Immunol. 13:127–149. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Donius LR and Weis JH: Quantification of
complement receptor 2 calcium signaling enhancement using flow
cytometry. Methods Mol Biol. 1100:311–317. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kataoka K, Iwanaga M, Yasunaga JI, Nagata
Y, Kitanaka A, Kameda T, Yoshimitsu M, Shiraishi Y, Sato-Otsubo A,
Sanada M, et al: Prognostic relevance of integrated genetic
profiling in adult T-cell leukemia/lymphoma. Blood. 131:215–225.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Singh S, Arcaroli J, Thompson DC,
Messersmith W and Vasiliou V: Acetaldehyde and
retinaldehyde-metabolizing enzymes in colon and pancreatic cancers.
Adv Exp Med Biol. 815:281–294. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Uemura M, Tamura K, Chung S, Honma S,
Okuyama A, Nakamura Y and Nakagawa H: Novel 5 alpha-steroid
reductase (SRD5A3, type-3) is overexpressed in
hormone-refractoryprostate cancer. Cancer Sci. 99:81–86.
2008.PubMed/NCBI
|
|
43
|
Vogelstein B and Kinzler KW: Cancer genes
and the pathways they control. Nat Med. 10:789–799. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chung HJ, Chi HS, Cho YU, Lee EH, Jang S,
Park CJ and Seo EJ: Prognostic effect of cytoplasmic CD79a
expression in acute myeloid leukemia with t(8;21). Korean J Lab
Med. 27:388–393. 2007.(In Korean). View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Berahovich R, Xu S, Zhou H, Harto H, Xu Q,
Garcia A, Liu F, Golubovskaya VM and Wu L: FLAG-tagged
CD19-specific CAR-T cells eliminate CD19-bearing solid tumor cells
in vitro and in vivo. Front Biosci (Landmark Ed). 22:1644–1654.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bednar KJ, Shanina E, Ballet R, Connors
EP, Duan S, Juan J, Arlian BM, Kulis MD, Butcher EC, Fung-Leung WP,
et al: Human CD22 inhibits murine B cell receptor activation in a
human CD22 transgenic mouse model. J Immunol. 199:3116–3128. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang Y, Gao M, Lin Z, Chen L, Jin Y, Zhu
G, Wang Y and Jin T: DEK promoted EMT and angiogenesis through
regulating PI3K/AKT/mTOR pathway in triple-negative breast cancer.
Oncotarget. 8:98708–98722. 2017.PubMed/NCBI
|
|
48
|
Szewczyk B, Pochwat B, Rafało A,
Palucha-Poniewiera A, Domin H and Nowak G: Activation of mTOR
dependent signaling pathway is a necessary mechanism of
antidepressant-like activity of zinc. Neuropharmacology.
99:517–526. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wong VKW, Zeng W, Chen J, Yao XJ, Leung
ELH, Wang QQ, Chiu P, Ko BCB and Law BYK: Tetrandrine, an activator
of autophagy, induces autophagic cell death via PKC-α inhibition
and mTOR-dependent mechanisms. Front Pharmacol. 8:3512017.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Darnell JE Jr: Transcription factors as
targets for cancer therapy. Nat Rev Cancer. 2:740–749. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
O'neill LA and Kaltschmidt C: NF-κB: A
crucial transcription factor for glial and neuronal cell function.
Trends Neurosci. 20:252–258. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Barnes PJ: Nuclear factor-kappa B. Int J
Biochem Cell Biol. 29:867–870. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhu X, Mao Z, Na Y, Guo Y, Wang X and Xin
D: Significance of pituitary tumor transforming gene 1 (PTTG1) in
prostate cancer. Anticancer Res. 26:1253–1259. 2006.PubMed/NCBI
|
|
54
|
Gorynia S, Bandeiras TM, Pinho FG, McVey
CE, Vonrhein C, Round A, Svergun DI, Donner P, Matias PM and
Carrondo MA: Structural and functional insights into a dodecameric
molecular machine-the RuvBL1/RuvBL2 complex. J Struct Biol.
176:279–291. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bièche I, Laurendeau I, Tozlu S, Olivi M,
Vidaud D, Lidereau R and Vidaud M: Quantitation of MYC gene
expression in sporadic breast tumors with a real-time reverse
transcription-PCR assay. Cancer Res. 59:2759–2765. 1999.PubMed/NCBI
|
|
56
|
Le Floch N, Rivat C, De Wever O, Bruyneel
E, Mareel M, Dale T and Gespach C: The proinvasive activity of
Wnt-2 is mediated through a noncanonical Wnt pathway coupled to
GSK-3beta and c-Jun/AP-1 signaling. FASEB J. 19:144–146. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chang KY, Huang CT, Hsu TI, Hsu CC, Liu
JJ, Chuang CK, Hung JJ, Chang WC, Tsai KK and Chuang JY: Stress
stimuli induce cancer-stemness gene expression via Sp1 activation
leading to therapeutic resistance in glioblastoma. Biochem Biophys
Res Commun. 493:14–19. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shibata T and Aburatani H: Exploration of
liver cancer genomes. Nat Rev Gastroenterol Hepatol. 11:340–349.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lim E, Vaillant F, Wu D, Forrest NC, Pal
B, Hart AH, Asselin-Labat ML, Gyorki DE, Ward T, Partanen A, et al:
Aberrant luminal progenitors as the candidate target population for
basal tumor development in BRCA1 mutation carriers. Nat Med.
15:907–913. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Azariadis K, Kiagiadaki F, Pelekanou V,
Bempi V, Alexakis K, Kampa M, Tsapis A, Castanas E and Notas G:
Androgen triggers the pro-migratory CXCL12/CXCR4 axis in
AR-positive breast cancer cell lines: Underlying mechanism and
possible implications for the use of aromatase inhibitors in breast
cancer. Cell Physiol Biochem. 44:66–84. 2017. View Article : Google Scholar : PubMed/NCBI
|