Introduction
Despite the advances made over the past decades in
the research area, melanoma remains the most aggressive type of
skin cancer. It shows an alarming increasing incidence, and in the
United States it was registered as the 6th most common type of
cancer in both sexes (1). Prognosis
for patient with melanoma depends on the stage of the disease at
the time of diagnosis. The unpredictable outcome is governed by a
multitude of complex reactions between the tumor and the immune
system of the host (2,3).
Melanoma has historically been considered an
immunogenic malignancy as it contains great amount of immune cells
(different subsets of T-cells, dendritic cells, macrophages,
neutrophils, mast cells, B lymphocytes) and the presence of a dense
tumor infiltrating lymphocytes (TILs) is associated with good
prognosis (3,4). The analysis of the populations and
subsets of immune cells that form TIL and their interaction with
tumor cells led to the development of antibodies against immune
check point inhibitors, with remarkable clinical results/outcome
and new molecules are under investigation (5,6).
Currently, diagnosis, prognosis and therapy are
based on the TNM staging system which takes into account
clinico-pathological risk factors: tumor thickness, ulceration,
mitotic rate, sentinel lymph node status and presence of metastases
(7). But AJCC classification cannot
predict the distinct progression and different responses to
treatment of melanomas classified at the same stage, showing the
urgent need to decipher new parameters to better stratify them, to
identify which patient needs a certain pathway inhibitor and in
what moment, according to his particular risk. Profound evaluation
of TIL can enlighten the way to new and more effective solutions
for melanoma patients.
Melanoma: An immunogenic tumor
It is known for many years that melanoma is an
immunogenic tumor and the use of this particularity in the
development of new therapeutic strategies has been attempted.
Several clinical aspects support this theory: partial or total
regression of primary melanomas, depigmentation areas within the
tumor or sometimes depigmentation ring (‘halo’) around the primary
melanomas or cutaneous metastases, and the development of
vitiligo-like depigmentation in patients with melanoma, associated
with a good prognosis (4,8).
The histopathological evidence of lymphocytes and
other mononuclear cells in association with melanoma cells prove
the involvement of the immune system. Although it was initially
considered the cause of the malignancies, nowadays the debate is to
establish if TIL are induced by tumor cells or by other components
of the microenvironment or whether they are a host direct
immunologic reaction (2). It has been
observed that in most situations, the presence of a dense TIL is
associated with better prognosis or a better survival rates
(9,10).
Tumor infiltrating lymphocytes as a
prognostic and predictive factor in melanoma
Over time, histopathological parameters in primary
cutaneous melanoma were subject to numerous studies regarding
prognosis of the disease. Thus, multivariate analyzes have shown
that tumor thickness and ulceration are the most important
prognostic predictors in localized disease. Increased evidence
suggests that tumor TIL represent an independent prognostic marker
(9–11).
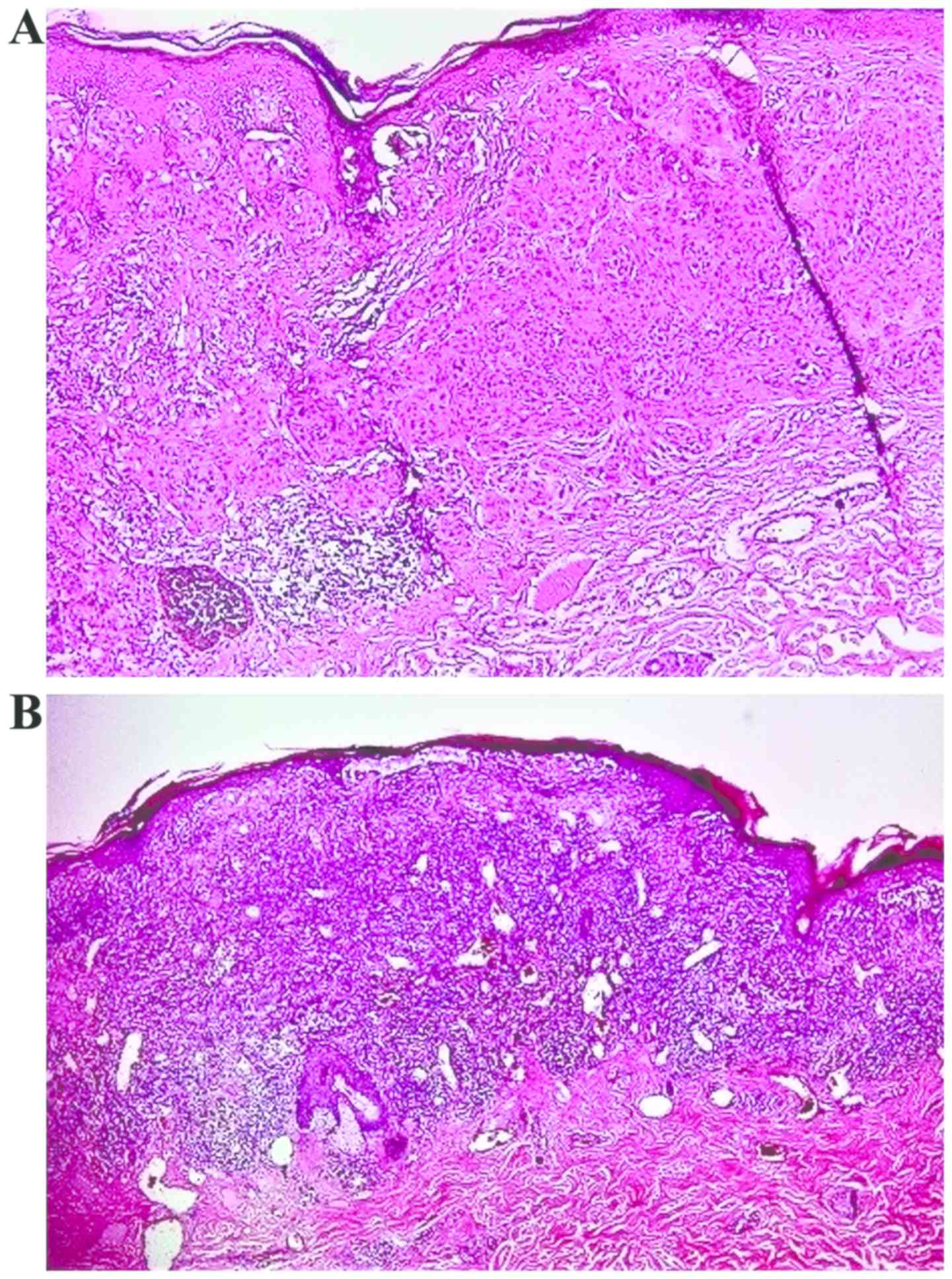
According to Clark et al (10), TIL infiltrate can be classified as:
absent, non-brisk or brisk and this quantification is still
accepted by most authors (12). TIL
are considered absent when leukocytes are absent or if they
are present, they do not infiltrate the tumor (12). Non-brisk TIL represents the infiltrate
distributed only focally and not along the entire base of the
invasive component (Fig. 1A).
Brisk TIL is defined by lymphocytes that infiltrate
diffusely the entire invasive component or infiltrating across the
entire base of vertical growth phase (Fig. 1B) (12,13).
Numerous studies have repeatedly shown the density
and the pattern of distribution of TIL in the vertical growth phase
of primary melanomas has prognostic significance, but not in the
radial growth phase (9,10,14,15).
Hussein et al have shown a progressive
increase in TIL during tumorigenesis in melanocytic lesions, this
being interpreted as an effect of increasing tumor antigenicity.
Interestingly, TIL decrease in metastatic melanoma reflects defence
breakdown (16).
Moreover, it can be seen as independent predictive
factor for sentinel lymph node status (2). It was observed that the prognosis of
patients with brisk inflammatory infiltrate was significantly
better than that of patients with non-brisk or absent TIL
and correlated with prolonged disease-free and overall survival
(9,10).
Due to its reported prognostic significance, some
authors consider that this parameter should be included and
quantified in the histological report (2).
The tumor microenvironment and tumor
infiltrating lymphocytes
Tumor mass is made up of tumor cells but also
stromal cells that enable them to live and grow, endothelial cells,
infiltrating leukocytes, fibroblasts and a complex extracellular
matrix that form the local tumor environment (17).
TIL are a polymorphic group that is composed mainly
by effector T lymphocytes, regulatory T lymphocytes, natural killer
(NK) cells, dendritic cells and macrophages (18). The distribution, the density, the
profile and the activation state of the cells which constitute TIL
can be variable and modulate the clinical outcome.
Based on the current information, for a better
understanding of the phenomena, we will present the scenario of the
development of TIL, the roles of its components and how they can
modulate the progression of the disease, with the final goal to be
able to develop new immunotherapeutic strategies.
Melanoma antigens
The first step in the initiation of the host immune
response in primary cutaneous melanoma is recognition of the
antigens that will induce inflammatory intratumoral infiltrate
(19).
Tumor antigens in melanoma are of two types:
specific antigens that are expressed by tumor cells (tumor-specific
antigens, TSAs) and tumor associated antigens (TAAs), which are
predominantly found in the tumor cells. TSA are also called
‘neoantigens’ because they are newly formed antigens and they are
not found in the normal human genome. TSA exert high immunogenicity
and induce T-lymphocytes that are not affected by central tolerance
and maintain for a period of time an efficient adaptative
anti-tumor activity. There is an extremely large number of possible
mutations, so that each patient tumor is unique, tumors from the
same patient can be different, and there is also intratumoral
heterogeneity (20). On the other
hand, TAA are mainly found in tumor cells but can also be expressed
by normal melanocytes and most of them are of intracellular protein
origin. Most often melanoma cells express only MHC class I and do
not express MHC class II molecules. TAA do not induce an ideal
cytotoxic T-cell reaction and generate reduced formation of
tumor-specific CD4+ and antibodies (4,11,21).
Malignant tumor cells can employ different stratagem
to avoid T-cell intervention. In some cases, tumor cells can reduce
their expression of TAA or they can produce chemokines and
cytokines with immunosuppressive consequences (22–25).
Dendritic cells (DCs)
DC are among the first cells to reach the tumor,
recognize tumor antigens and play a pivotal role in the initiation
and regulation of both innate and adaptive immunity (17). They are the most potent
antigen-presenting cells (APCs) and after processing of
tumor-associated antigens, they induce a specific antitumor
response by cross-presentation of antigens to CD8+ T
lymphocytes by MHC class I molecules and to CD4+ by MHC
class II molecules (26). In order to
obtain a potent anti-tumoral immune response of T lymphocytes, the
antigen presentation must be efficiently done by mature dendritic
cells. DC can additionally contribute to the cytotoxic immune
reaction directly and by activating NK cells (27).
DC from the skin, Langerhans cells, dermal DC and
plasmacytoid DC may exhibit different phenotypes, with dualistic
functions (28). In the tumor
microenvironment, mediators released by tumor cells or
tumor-associated macrophages (TAMs) such as IL-8, IL-10, TGF-β1 and
VEGF limit normal DC maturation, in an attempt to evade host immune
response (20,29,30).
DCs in the immature state fail to properly stimulate
T-cells. On the other hand, by favoring proliferation of regulatory
T-cells, they block T-cell responses. In this way they are
mediators of immune tolerance despite immune activation, a
phenomenon described in patients with melanoma in 1997 by Enk et
al (31). Immature/tolerogenic DC
regulate tumor angiogenesis and favor an active tumor growth
(32). This proangiogenic effect is
no longer operative with DC maturation. The maturation process of
dendritic cells is dependent on factors encountered in the tumor
microenvironment (33).
Mature DC are distributed mainly peritumorally and
their density, together with the activation status of T lymphocyte,
correlated with melanoma tumor thickness and patient's survival,
making Kobayashi et al and Simonetti et al to
recommend the use of these parameters as a predictor of treatment
response in patients treated with immunotherapy (34,35).
An adequate number of mature DC in tumor area can
eliminate malignant cells (36).
Lotze showed that infiltration with mature DC in primary tumors is
associated with fewer metastases and better prognosis (37).
Interaction of APC and T
lymphocyte-immune checkpoints
In an ideal situation, antigen-presenting cells lead
to activation of effector memory T-cells in the lymph nodes that
mediate antitumor effects at tumor site, producing new antigens
from destroyed tumor cell and creating a tumor-immunity cycle
(2,38).
After recognizing the antigen presented by APC,
T-cells require several signals for full activation. The first
signal depends on the antigen and is represented by MHC I or MHC II
complexed with the presented peptide, binding to the T-cell
receptor (TCR) (39). Insufficiently
presented antigen on APC cannot activate T-cells and induce
immunologic ignorance (40).
Cell adhesion molecules are responsible for
maintaining the connection between the two cells and allow as many
TCRs on the T-cell as possible to become activated (41). In metastatic disease a diversity of
TCR was observed between different sites that can have different
clinical evolution, explained by the selection of T-cells directed
against different tumor antigens or against different epitopes from
the same antigen (38).
The second signal of activation requires specific
interaction between T-cell receptors from their ligands and APC
(from CD28/B7 family), and is antigen-independent (39). These receptors are of two types:
co-stimulators or co-inhibitors. Many ligands can bind to multiple
receptors, of each type (42).
The interplay between costimulatory receptors and
their ligand completely activate T-cells and induce production of
tumor-specific T-cells (43).
In normal conditions, co-inhibitor receptors
[programmed cell death-1 (PD-1), cytotoxic T lymphocyte antigen-4
(CTLA-4), B and T lymphocyte attenuator (BTLA) and T-cell
immunoglobulin and mucin domain-3 (TIM-3), known as ‘immune
checkpoint molecules’, and their ligands (respectively, programmed
death-ligand 1 (PD-L1) or PD-L2, CD80 or CD86, herpesvirus entry
mediator (HVEM) and galectin 9], inhibit T-cell activity and are
involved in maintaining self-tolerance and modulating the intensity
and duration of an anti-infectious immune response (30,42).
Activation of inhibitory immune checkpoints by cancer cells shield
them from the immune attack and allow them to proliferate further
(38,44).
APC T-cell interaction can be mediated also by tumor
necrosis factor (TNF) family members and their receptors, by the
soluble cytokines from the microenvironment (TGF-β, IL-1, IL-10,
IL-12, IL-18) and by immune-inhibitory molecules, metabolic enzymes
that deprive lymphocytes of necessary aminoacids [indoleamine
2,3-dioxygenase (IDO)] that can be new therapeutic targets
(42,45,46).
Thus, it has been observed that the immune system
has a dual role, by modifying the interactions between
co-stimulatory and co-inhibitory signals. On the one hand, it can
control malignant cells but on the other hand, it may favor tumor
progression (47).
T lymphocytes
TIL include different subsets of lymphocytes, in
different proportions, adjusted by cytokines secreted by tumor
cells or other residents: CD8+ T-cells, CD4+
T-cells, NK cells and B-cells (48,49). T
lymphocytes have a major role in the antitumor immune responses and
are the dominant elements in the tumor microenvironment.
TSA generate highly sensitive and specific
CD8+ T recognition and data suggest that TIL may
specifically target TSA (50,51). TAA can induce tumor-specific
CD8+, CD4+ T-cells and antibodies against TAA
(30).
CD8 effector T-cells (or cytotoxic T-cells) inhibit
tumor proliferation, either through direct cytolytic action on
tumor cells or by releasing interferon (IFN)-γ and TNF-α (50,52).
Therefore, a large infiltration of CD8+ T-cells in the
tumor is related to a good prognosis in patients with melanoma
(53). Protracted antigen exposure
can cause tumor antigen-specific T-cells ‘exhaustion’. PD-1 and
TIM-3 have been considered immunohistochemical markers for
‘exhaustion’ (4). Based on the
balance between co-stimulatory and co-inhibitory signals from the
micro-environment, CD8 T-cells may exert different functional
states (54).
There are four main types of CD4+ T-cells
(T helper cells), with distinct properties and in variable
percentage: Th1, Th2 Th17, and Treg.
Th1, Th2 help antitumor fight by stimulating the
activity of CD8+ T-cells through mediators such as
IFN-γ, TGF-β or IL-2 (55,56). Th1 can also favor activation of
macrophages and maturation of dendritic cells, while Th2 can use
cytotoxic weapons of eosinophils (57,58). A
consistent intratumoral infiltration of CD8+ T-cells and
Th1 cells is correlated with favorable prognosis and better
survival in most human cancers (59).
Th17 cells have two antagonist forms: in certain
cytokine milieu can convert to Th1 characteristics and exert
antitumor effects, while in other conditions can switch to
regulatory T-cells characteristics and induce tumor progression
(60).
Regulatory T-cells (CD25/FoxP3 suppressive T-cells,
Treg) are a subtype of T-cells with a key role in preventing
autoimmune diseases. Their presence in the tumor microenvironment
inhibits the antitumor immune responses. They are attracted to the
tumor microenvironment by chemokines secreted by tumor cells and
macrophages. Tregs are activated after recognizing TAA released
from destroyed tumor cells and then specifically suppress the
activation of TAA-specific effector T-cells and the efficient tumor
cell destruction by various mechanisms (IL-10, TGF-β) (41,61).
Moreover, Treg can suppress the action of several types of immune
system cells such as CD8+ T-cells, NK cells, B cells and
APC (62).
There are few studies done on the prognostic role of
Treg in melanoma. Some have noted the association between the
presence of a high Tregs infiltrate with the local recurrence of
the disease, fast tumor progression, a higher metastasis rate in
the sentinel lymph node, but without any association with tumor
thickness or patient survival (63–66).
It is considered that ratios between different
subsets of T-cells can provide more accurate information on the
local immune balance and CD8/FoxP3 (effector/regulatory) ratio and
CD8/CD4 (effector/helper) ratio are the most commonly used
(67). To skip from immune attack,
tumor cells suppress tumor-specific T effector and induce
immunosuppressive Treg cells, thus reducing CD8/Treg ratios. In
patients treated with combination checkpoint therapy, CD8/Treg
ratios increased and was associated with improving survival in
melanoma (68).
NK cells
NK are effector cells of the innate immune system
and play an essential role in cancer immune surveillance due to
their ability to destroy tumor cells independent of MHC or previous
activation (69).
NK also participate in regulation of adaptive immune
reactions through interactions with DCs, that activate NK cells and
determine a potent cytotoxic immune response against tumor cells
(27).
Tumor cells may defeat NK by releasing TGF-β, by low
antigenicity expression or by increased MHC I expression (33,69–71). Tregs
can also contribute by rivaling with NK for IL-2 (30). The activation state of NK cells is
modulated by activating and inhibitory receptors.
The prognostic role of NK cell infiltration in
melanoma has not been evaluated, but they seem to limit the
development of hematogenous metastases (28).
B cells
B cells represent 15–20% of all infiltrating
lymphocytes (16,30,66). The
exact roles of tumor-infiltrating B cells in antitumoral immune
response are not defined. A trend for their higher prevalence was
observed in thicker tumors and an increased density of B
lymphocytes infiltrating primary cutaneous melanomas was associated
with reduced risk of distant metastases and longer survival
(28,72) The roles of plasma cells are even less
understood. Bosisio et al recently reported a significantly
worse survival in primary melanomas with clusters of plasma cells
compared to a better outcome in the cases with sparse plasma cell
infiltrate. Most plasma cells were polyclonal, expressing
predominantly IgG and IgA (73).
Macrophages
TAMs are a heterogeneous group of cells with
antigen-presenting capacity and represent the predominant
inflammatory cells of the tumor infiltrating lymphocytes. They are
found in all stages of the tumor progression (30). Based on the signals from
microenvironment, macrophages can render antagonist functional
characteristics.
M1 macrophages, the classical type, have high
antigen-presenting action and also can produce Th1 cytokines and
exert antitumor effect (69).
M2 macrophages have a low antigen-presenting
activity, inhibit CD8+ T-cell and NK cell activity,
stimulate switch to Th2 and Treg predominance, stimulate antibody
production, angiogenetic effects and favor tumor cell migration
(74,75). It is the predominant profile in TAM,
induced by products secreted by dendritic cells, Treg lymphocytes
and tumor microenvironment. During tumor growth and progression,
macrophages convert from M1 to M2 phenotype (29).
Studies have shown correlation between TAM density
and tumor thickness and ulceration, but no significant correlation
with survival in patients with melanoma (76).
Immunoediting
All the above cells are players in a complex process
of immunoediting. They produce a variety of cytokines, chemokines
and growth factors with important roles in adjusting interactions
between cells, regulating cell differentiation, activation and
functioning of many types of cells (17).
Permanent interactions between the tumor and the
immune system can lead to malignant cell elimination, an
equilibrium state or malignant cell escape (77). The transition between these phases is
adjusted by the immune system status and the activity of the tumor
cells. If the innate and the adaptive immune systems successfully
remove tumor cells, the elimination is the final stage of
immunoediting (77).
In some cases, not all tumor cells can be
eliminated, but their proliferation can be handled by the immune
system. In this equilibrium state tumor cells remain latent for a
variable period of time (78).
Nevertheless malignant cells possess a wide variety
of means that in the end enable them to escape from the control of
the immune system, starting to proliferate and lead to the
appearance of the primary tumors and metastases.
Melanoma cells can passively evade or actively
suppress the antitumor immune response through several mechanisms,
deceiving and defeating one by one each of the immune system
players previously described: i) downregulate TAA (79,80); ii)
induce T-cell exhaustion by prolonged TSA exposure (81); iii) express low levels of MHC I
(82); iv) evade recognition by NK
cells and reduce their cytolytic activity (83); v) induce T-cell apoptosis by Fas-Fas
ligand pathway (49); vi) recruit
tolerogenic Treg (84); vii) produce
inhibitory signaling molecules and induce immunological tolerance
(85,86); and viii) express immune checkpoint
proteins that shelter them from T-cells or macrophages attack
(87). Despite a strong, complex
anti-melanoma immune response, melanoma cells use all the tricks to
escape without being destroyed (30,77).
Discussion
Each individual, each organ, and accordingly, each
tumor and each metastatic site form the same tumor have distinctive
immunologic particularities that are far from being deciphered.
This review wants to bring to attention the multiple
variable that modulate melanoma outcome, starting from TSA that can
be the result of a unique mutation, and induce a specific effector
T-cell, which can have a variable activity state, depending on how
the antigen was presented, depending on the specific context of
either stimulatory or inhibitory receptors, cytokines and
chemokines, that can be variable in time and favor accumulation of
different proportions of NK, macrophages or Treg, with specific
phenotypes and functions. The cumulative effect of these antagonist
forces can be either successful elimination of tumor cells,
temporary control or evasion.
Several therapeutic approaches have been tested,
addressing different steps involved in melanomagenesis, with
unsatisfactory results. The new data on the role of the immune
system and the mechanism of immune escape led to the development of
immune checkpoint inhibitors, that unblock antitumor immunity,
considered among the most important medical findings of the recent
years, with significant increased survival for patients with stage
IV melanoma.
Even if it may represent a photo of a certain moment
in tumor evolution movie, analyzing the profile of the immune cells
from TIL may help understanding of tumor immunobiology and,
considering the already proved prognostic significance, it may help
to identify potential biomarkers for a better risk stratification,
to further devise new weapons, for new targets, for the individual
unique melanoma.
Acknowledgements
Not applicable.
Funding
This work is partially supported by a grant of
Ministry of Research and Innovation, CNCS-UEFISCDI (project no.
PN-III-P4-ID-PCE-2016-0641) within PNCDI–III. This work was
partially supported by a grant of Romanian Ministry of Research and
Innovation, CCCDI-UEFISCDI (project nο. 61PCCDI⁄2018
PN-III-P1-1.2-PCCDI-2017-0341) within PNCDI–III.
Availability of data and materials
Not applicable.
Authors' contributions
MA, RIN, GT contributed to the conception and design
of the study, interpretation of the data, and revising it
critically for important intellectual content. LN, CGP, MC, SAZ,
AB, AH, AC, MB were responsible for the acquisition and
interpretation of the data, writing and drafting the study. DAI,
CD, CB, DP had substantial contributions to the interpretation and
analysis of the data, drafting the study and revising it critically
for important intellectual content. All authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TILs
|
tumor-infiltrating lymphocytes
|
|
CTLA4
|
cytotoxic T lymphocyte antigen 4
|
|
PD-1
|
programmed cell death-1
|
|
PD-L1
|
programmed death-ligand 1
|
|
TSAs
|
tumor-specific antigens
|
|
TAAs
|
tumor-associated antigens
|
|
TCR
|
T-cell receptor
|
|
APCs
|
antigen-presenting cells
|
References
|
1
|
Sosman JA: Patient education: Melanoma
treatment; advanced or metastatic melanoma (Beyond the Basics).
Atkins MB and Vora SR (eds). https://www.uptodate.com/contents/melanoma-treatment-advanced-or-metastatic-melanoma-beyond-the-basics
|
|
2
|
Azimi F, Scolyer RA, Rumcheva P, Moncrieff
M, Murali R, McCarthy SW, Saw RP and Thompson JF:
Tumor-infiltrating lymphocyte grade is an independent predictor of
sentinel lymph node status and survival in patients with cutaneous
melanoma. J Clin Oncol. 30:2678–2683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Neagu M: The immune system: A hidden
treasure for biomarker discovery in cutaneous melanoma. Adv Clin
Chem. 58:89–140. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mukherji B: Immunology of melanoma. Clin
Dermatol. 31:156–165. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dummer R, Hauschild A, Lindenblatt N,
Pentheroudakis G and Keilholz U: Cutaneous melanoma: ESMO clinical
practice guidelines for diagnosis, treatment and follow-up. Ann
Oncol. 26:126–132. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Turcu G, Nedelcu RI, Ion DA, Brînzea A,
Cioplea MD, Jilaveanu LB and Zurac SA: CEACAM1: Expression and role
in melanocyte transformation. Disease Markers. 2016:94063192016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Coit DG, Thompson JA, Algazi A, Andtbacka
R, Bichakjian CK, Carson WE, Daniels GA, DiMaio D, Ernstoff M,
Fields RC, et al: Melanoma, Version 2.2016, NCCN Clinical Practice
Guidelines in Oncology. J Natl Compr Canc Netw. 14:450–473. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nedelcu RI, Zurac SA, Brînzea A, Cioplea
MD, Turcu G, Popescu R and Ion DA: Morphological features of
melanocytic tumors with depigmented halo: review of the literature
and personal results. Rom J Morphol Embryol. 56:659–663.
2015.PubMed/NCBI
|
|
9
|
Clemente CG, Mihm Jr MC, Bufalino R,
Zurrida S, Collini P and Cascinelli N: Prognostic value of tumor
infiltrating lymphocytes in the vertical growth phase of primary
cutaneous melanoma. Cancer. 77:1303–1310. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Clark WH, Elder DE, Guerry D, Braitman LE,
Trock BJ, Schultz D, Synnestvedt M and Halpern AC: Model predicting
survival in stage I melanoma based on tumor progression. J Natl
Cancer Inst Dec. 81:1893–1904. 1989. View Article : Google Scholar
|
|
11
|
Lee N, Zakka LR, Mihm MC and Schatton T:
Tumour-infiltrating lymphocytes in melanoma prognosis and cancer
immunotherapy. Pathology. 48:177–187. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Busam KJ, Antonescu CR, Marghoob AA, Nehal
KS, Sachs DL, Shia J and Berwick M: Histologic classification of
tumor-infiltrating lymphocytes in primary cutaneous malignant
melanoma: A study of interobserver agreement. Am J Clin Pathol.
115:856–860. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Spatz A, Batist G and Eggermont AM: The
biology behind prognostic factors of cutaneous melanoma. Curr Opin
Oncol. 22:163–168. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tuthill RJ, Unger JM, Liu PY, Flaherty LE
and Sondak VK: Risk assessment in localized primary cutaneous
melanoma: A southwest oncology group study evaluating nine factors
and a test of the Clark logistic regression prediction model. Am J
Clin Pathol. 118:504–511. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elder DE, Guerry D, Vanhorn M, Hurwitz S,
Zehngebot L, Goldman LI, LaRossa D, Hamilton R, Bondi EE and Clark
WH Jr: The role of lymph node dissection for clinical stage I
malignant melanoma of intermediate thickness (1.51–3.99 mm).
Cancer. 56:413–418. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hussein MR, Elsers DA, Fadel SA and Omar
AE: Immunohistological characterisation of tumour infiltrating
lymphocytes in melanocytic skin lesions. J Clin Pathol. 59:316–324.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shurin MR, Shurin G V, Lokshin A and
Ferris RL: Intratumoral cytokines/chemokines/growth factors and
tumor infiltrating dendritic cells: friends or enemies? Cancer
Metast Rev. 25:333–356. 2006. View Article : Google Scholar
|
|
18
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Van Der Bruggen P, Zhang Y, Chaux P,
Stroobant V, Panichelli C, Schultz ES, Chapiro J, Van Den Eynde BJ,
Brasseur F and Boon T: Tumor-specific shared antigenic peptides
recognized by human T-cells. Immunol Rev. 188:51–64. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gerlinger M, Rowan AJ, Horswell S, Larkin
J, Endesfelder D, Gronroos E, Martinez P, Matthews N, Stewart A,
Tarpey P, et al: Intratumor heterogeneity and branched evolution
revealed by multiegion sequencing. N Engl J Med. 366:883–892. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Durrant L and Ramage J: Development of
cancer vaccines to activate cytotoxic T lymphocytes. Expert Opin
Biol Ther. 5:555–563. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ferrone S and Marincola FM: Loss of HLA
class I antigens by melanoma cells: molecular mechanisms,
functional significance and clinical relevance. Immunol Today.
16:487–494. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kageshita T, Hirai S, Ono T, Hicklin DJ
and Ferrone S: Downregulation of HLA class I antigen-processing
molecules in malignant melanoma. Am J Pathol. 154:745–754. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Al-Batran SE, Rafiyan MR, Atmaca A,
Neumann A, Karbach J, Bender A, Weidmann E, Altmannsberger HM,
Knuth A, et al: Intratumoral T-cell infiltrates and MHC class I
expression in patients with stage IV melanoma. Cancer Res.
65:3937–3941. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clancy T and Hovig E: Profiling networks
of distinct immune-cells in tumors. BMC Bioinformatics. 17:1–15.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schiavoni G, Gabriele L and Mattei F: The
tumor microenvironment: a pitch for multiple players. Front Oncol.
3:2013.doi: 10.3389/fonc.2013.00090. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tel J, Anguille S, Waterborg CEJ, Smits
EL, Figdor CG and de Vries IJM: Tumoricidal activity of human
dendritic cells. Trends Immunol. 35:38–46. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ladányi A: Prognostic and predictive
significance of immune cells infiltrating cutaneous melanoma.
Pigment Cell Melanoma Res. 28:490–500. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vitale M, Cantoni C, Pietra G, Mingari MC
and Moretta L: Effect of tumor cells and tumor microenvironment on
NK-cell function. Eur J Immunol. 44:1582–1592. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Muenst S, Läubli H, Soysal SD, Zippelius
A, Tzankov A and Hoeller S: The immune system and cancer evasion
strategies: Therapeutic concepts. J Intern Med. 279:541–562. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Enk AH, Jonuleit H, Saloga J and Knop J:
Dendritic cells as mediators of tumor-induced tolerance in
metastatic melanoma. Int J Cancer. 73:309–316. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fainaru O, Almog N, Yung CW, Nakai K,
Montoya-Zavala M, Abdollahi A, D'Amato R and Ingber DE: Tumor
growth and angiogenesis are dependent on the presence of immature
dendritic cells. FASEB J. 24:1411–1418. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Da Cunha A, Michelin MA and Murta EF:
Pattern response of dendritic cells in the tumor microenvironment
and breast cancer. World J Clin Oncol. 5:495–502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kobayashi M, Suzuki K, Yashi M, Yuzawa M,
Takayashiki N and Morita T: Tumor infiltrating dendritic cells
predict treatment response to immmunotherapy in patients with
metastatic renal cell carcinoma. Anticancer Res. 27:1137–1141.
2007.PubMed/NCBI
|
|
35
|
Simonetti O, Goteri G, Lucarini G, Rubini
C, Stramazzotti D, Lo Muzio L, Biagini D and Offidani A: In
melanoma changes of immature and mature dendritic cell expression
correlate with tumor thickness:an immunohistochemical study. Int J
Immunopathol Pharmacol. 20:325–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
El Marsafy S, Bagot M, Bensussan A and
Mauviel A: Dendritic cells in the skin-potential use for melanoma
treatment. Pigment Cell Melanoma Res. 22:30–41. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lotze MT: Getting to the source: dendritic
cells as therapeutic reagents for the treatment of patients with
cancer. Ann Surg. 226:1–5. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Boddupalli CS, Bar N, Kadaveru K,
Krauthammer M, Pornputtapong N, Mai Z, Ariyan S, Narayan D, Kluger
H, Deng Y, et al: Interlesional diversity of T-cell receptors in
melanoma with immune checkpoints enriched in tissue-resident memory
T-cells. JCI Insight. 1:e889552016.doi: 10.1172/jci.insight.88955.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pedoeem A, Azoulay-Alfaguter I, Strazza M,
Silverman GJ and Mor A: Programmed death-1 pathway in cancer and
autoimmunity. Clin Immunol. 153:145–152. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ochsenbein AF, Klenerman P, Karrer U,
Ludewig B, Pericin M, Hengartner H and Zinkernagel RM: Immune
surveillance against a solid tumor fails because of immunological
ignorance. Proc Natl Acad Sci. 96:2233–2238. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Inman B, Frigola X, Dong H and Kwon E:
Costimulation, coinhibition and cancer. Curr Cancer Drug Targets.
7:15–30. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pardoll DM: The blockade of immune
checkpoints in cancer immunotherapy. Nat Rev Cancer. 12:252–264.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mapara MY and Sykes M: Tolerance and
Cancer: Mechanisms of tumor evasion and strategies for breaking
tolerance. J Clin Oncol. 22:1136–1151. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ancuceanu R and Neagu M: Immune based
therapy for melanoma. Indian J Med Res. 143:135–144. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Munn DH and Mellor AL: Indoleamine
2,3-dioxygenase and tumor-induced tolerance. J Clin Invest.
117:1147–1154. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ochoa AC, Zea AH, Hernandez C and
Rodriguez PC: Arginase, prostaglandins, and myeloid-derived
suppressor cells in renal cell carcinoma. Clin Cancer Res.
13:721–726. 2007. View Article : Google Scholar
|
|
47
|
Becht E, Goc J, Germain C, Giraldo NA,
Dieu-Nosjean MC, Sautès-Fridman C and Fridman WH: Shaping of an
effective immune microenvironment to and by cancer cells. Cancer
Immunol Immunother. 63:991–997. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Rosenberg S, Packard B, Aebersold P,
Solomon D, Topalian S, Toy S, Simon P, Lotze MT, Yang JC, Seipp CA,
et al: Use of tumor-infiltrating lymphocytes and interleukin-2 in
the immunotherapy of patients with metastatic melanoma. A
preliminary report. N Engl J Med. 319:1676–1680. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cohen PJ, Lotze MT, Roberts JR, Rosenberg
SA and Jaffe ES: The immunopathology of sequential tumor biopsies
in patients treated with interleukin-2. Correlation of response
with T-cell infiltration and HLA-DR expression. Am J Pathol.
129:208–216. 1987.PubMed/NCBI
|
|
50
|
Savage P, Leventhal DS and Malchow S:
Shaping the repertoire of tumor-infiltrating effector and
regulatory T-cells. Immunol Rev. 259:245–258. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Rosenberg SA and Restifo NP: Adoptive cell
transfer as personalized immunotherapy for human cancer. Science.
348:62–68. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fridman WH, Remark R, Goc J, Giraldo NA,
Becht E, Hammond SA, Damotte D, Dieu-Nosjean MC and Sautès-Fridman
C: The immune microenvironment: A major player in human cancers.
Int Arch Allergy Immunol. 164:13–26. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Schlapbach C, Shafighi M, Kiermeir D,
Hüsler R and Hunger RE: High expression of FOXP3 in primary
melanoma is associated with tumour progression. Br J Dermatol.
170:103–109. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Giraldo NA, Becht E, Remark R, Damotte D,
Sautès-Fridman C and Fridman WH: The immune contexture of primary
and metastatic human tumours. Curr Opin Immunol. 27:8–15. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kiraz Y, Baran Y and Nalbant A: T-cells in
tumor microenvironment. Tumor Biol. 37:39–45. 2016. View Article : Google Scholar
|
|
56
|
Yang ZZ and Ansell SM: The tumor
microenvironment in follicular lymphoma. Clin Adv Hematol Oncol.
10:810–818. 2012.PubMed/NCBI
|
|
57
|
Hung K, Hayashi R, Lafond-Walker A,
Lowenstein C, Pardoll D and Levitsky H: The central role of
CD4+ T-cells in the antitumor immune response. J Exp
Med. 188:2357–2368. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pereira MC, Oliveira DT and Kowalski LP:
The role of eosinophils and eosinophil cationic protein in oral
cancer (Review). Arch Oral Biol. 56:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Fridman WH, Pagès F, Sautès-Fridman C and
Galon J: The immune contexture in human tumours: impact on clinical
outcome. Nat Rev Cancer. 12:298–306. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Bailey SR, Nelson MH, Himes RA, Li Z,
Mehrotra S and Paulos CM: Th17 cells in cancer: the ultimate
identity crisis. Front Immunol. 5:1664–3224. 2014. View Article : Google Scholar
|
|
61
|
Nishikawa H and Sakaguchi S: Regulatory
T-cells in cancer immunotherapy. Curr Opin Immunol. 27:1–7. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Jiang X and Shapiro DJ: The immune system
and inflammation in breast cancer. Mol Cell Endocrinol.
382:673–682. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Miracco C, Mourmouras V, Biagioli M,
Rubegni P, Mannucci S, Monciatti I, Cosci E, Tosi P and Luzi P:
Utility of tumour-infiltrating CD25+FOXP3+
regulatory T-cell evaluation in predicting local recurrence in
vertical growth phase cutaneous melanoma. Oncol Rep. 18:1115–1122.
2007.PubMed/NCBI
|
|
64
|
Gambichler T, Bindsteiner M, Höxtermann S,
Terras S and Kreuter A: Circulating CD4+ CD25(high)
CD127(low) regulatory T-cells are an independent predictor of
advanced melanoma. Pigment Cell Melanoma Res. 26:280–283. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ma MW, Medicherla RC, Qian M, Vega-Saenz
de Miera E, Friedman EB, Berman RS, Shapiro RL, Pavlick AC, Ott PA,
Bhardwaj N, et al: Immune response in melanoma: an in-depth
analysis of the primary tumor and corresponding sentinel lymph
node. Mod Pathol. 25:1000–1010. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hillen F, Baeten CIM, Van De Winkel A,
Creytens D, Van Der Schaft DWJ, Winnepenninckx V and Griffioen AW:
Leukocyte infiltration and tumor cell plasticity are parameters of
aggressiveness in primary cutaneous melanoma. Cancer Immunol
Immunother. 57:97–106. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Gooden MJM, De Bock GH, Leffers N, Daemen
T and Nijman HW: The prognostic influence of tumour-infiltrating
lymphocytes in cancer: A systematic review with meta-analysis. Br J
Cancer. 105:93–103. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Twyman-Saint Victor C, Rech AJ, Maity A,
Rengan R, Pauken KE, Stelekati E, Benci JL, Xu B, Dada H, Odorizzi
PM, et al: Radiation and dual checkpoint blockade activate
non-redundant immune mechanisms in cancer. Nature. 520:373–377.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Bruno A, Ferlazzo G, Albini A and Noonan
DM: A think tank of TINK/TANKs: tumor-infiltrating/tumor-associated
natural killer cells in tumor progression and angiogenesis. J Natl
Cancer Inst. 106:dju2002014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Sungur CM and Murphy WJ: Positive and
negative regulation by NK cells in cancer. Crit Rev Oncog.
19:57–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Balsamo M, Vermi W, Parodi M, Pietra G,
Manzini C, Queirolo P, Lonardi S, Augugliaro R, Moretta A,
Facchetti F, et al: Melanoma cells become resistant to
NK-cell-mediated killing when exposed to NK-cell numbers compatible
with NK-cell infiltration in the tumor. Eur J Immunol.
42:1833–1842. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ladányi A, Kiss J, Mohos A, Somlai B,
Liszkay G, Gilde K, Fejös Z, Gaudi I, Dobos J and Tímár J:
Prognostic impact of B-cell density in cutaneous melanoma. Cancer
Immunol Immunother. 60:1729–1738. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Bosisio FM, Wilmott JS, Volders N, Mercier
M, Wouters J, Stas M, Blokx WA, Massi D, Thompson JF, Scolyer RA,
et al: Plasma cells in primary melanoma. Prognostic significance
and possible role of IgA. Mod Pathol. 29:347–358. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Allavena P, Sica A, Solinas G, Porta C and
Mantovani A: The inflammatory micro-environment in tumor
progression: The role of tumor-associated macrophages. Crit Rev
Oncol/Hematol. 66:1–9. 2008. View Article : Google Scholar
|
|
75
|
Sica A, Schioppa T, Mantovani A and
Allavena P: Tumour-associated macrophages are a distinct M2
polarised population promoting tumour progression: Potential
targets of anti-cancer therapy. Eur J Cancer. 42:717–727. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Storr SJ, Safuan S, Mitra A, Elliott F,
Walker C, Vasko MJ, Ho B, Cook M, Mohammed RA, Patel PM, et al:
Objective assessment of blood and lymphatic vessel invasion and
association with macrophage infiltration in cutaneous melanoma. Mod
Pathol. 25:493–504. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Mittal D, Gubin MM, Schreiber RD and Smyth
MJ: New insights into cancer immunoediting and its three component
phases - elimination, equilibrium and escape. Curr Opin Immunol.
27:16–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Koebel CM, Vermi W, Swann JB, Zerafa N,
Rodig SJ, Old LJ, Smyth MJ and Schreiber RD: Adaptive immunity
maintains occult cancer in an equilibrium state. Nature.
450:903–907. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Khong HT, Wang QJ and Rosenberg SA:
Identification of multiple antigens recognized by
tumor-infiltrating lymphocytes from a single patient: tumor escape
by antigen loss and loss of MHC expression. J Immunother.
27:184–190. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Maeurer MJ, Gollin SM, Martin D, Swaney W,
Bryant J, Castelli C, Robbins P, Parmiani G, Storkus WJ and Lotze
MT: Tumor escape from immune recognition: lethal recurrent melanoma
in a patient associated with downregulation of the peptide
transporter protein TAP-1 and loss of expression of the
immunodominant MART-1/Melan-A antigen. J Clin Invest. 98:1633–1641.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wherry EJ: T-cell exhaustion. Nat Immunol.
12:492–499. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Schatton T, Schütte U, Frank NY, Zhan Q,
Hoerning A, Robles SC, Zhou J, Hodi FS, Spagnoli GC, Murphy GF and
Frank MH: Modulation of T-cell activation by malignant melanoma
initiating cells. Cancer Res. 70:697–708. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Solana R, Casado JG, Delgado E, DelaRosa
O, Marín J, Durán E, Pawelec G and Tarazona R: Lymphocyte
activation in response to melanoma: interaction of NK-associated
receptors and their ligands. Cancer Immunol Immunother. 56:101–109.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Jandus C, Bioley G, Speiser DE and Romero
P: Selective accumulation of differentiated FOXP3+
CD4+ T-cells in metastatic tumor lesions from melanoma
patients compared to peripheral blood. Cancer Immunol Immunother.
57:1795–1805. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Gorelik L and Flavell RA: Immune-mediated
eradication of tumors through the blockade of transforming growth
factor-β signaling in T-cells. Nat Med. 7:1118–1122. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Nicolaou A, Estdale SE, Tsatmali M,
Herrero DP and Thody AJ: Prostaglandin production by melanocytic
cells and the effect of α-melanocyte stimulating hormone. FEBS
Lett. 570:223–226. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Hirano F, Kaneko K, Tamura H, Dong H, Wang
S, Ichikawa M, Rietz C, Flies DB, Lau JS, Zhu G, et al: Blockade of
B7-H1 and PD-1 by monoclonal antibodies potentiates cancer
therapeutic immunity. Cancer Res. 65:1089–1096. 2005.PubMed/NCBI
|