Introduction
Melanoma is the most aggressive and lethal type of
skin cancer worldwide (1). Although
it accounts for only 4% of all cases of skin cancer, it causes the
highest number of cases of skin cancer-associated mortality
worldwide due to its high metastasis (2,3).
Following lung and breast cancer, melanoma is the third most common
reason for brain metastases (4).
Melanoma spreads to the brain in ≤75% of patients with melanoma
(5). Brain metastases cause
mortality in 95% of patients with high class (Class III) (6). The incidence of primary melanoma has
been increasing rapidly and doubles every 10–14 years, from 2006 to
2013 (7). Different anti-melanoma
strategies have been developed and used in clinical treatment,
including chemotherapy, immunotherapy and targeted therapy
(8). Targeted therapy is widely used
in cancer treatment due to its small side effects compared with
other strategies (9). BRAF
inhibitors, Sorafenib, Vemurafenib, Dabrafenib, mitogen-activated
protein kinase kinase inhibitor and Trametinib have been widely
used for melanoma treatment, but different inhibitors specifically
target particular mechanisms of melanoma occurrence and
progression, and one inhibitor usually only treats a small
percentage of patients with melanoma (2,10).
Therefore, novel targets for melanoma treatment are urgently
required.
Makorin ring finger protein 2 (MKRN2) belongs to the
makorin RING zinc finger family that encodes putative
ribonucleoproteins with a distinctive array of zinc finger domains
(11,12). Makorins are zinc finger proteins with
a typical C3HC4 motif (the RING domain), associated with arrays of
one to four C3H domains, and represent a type of zinc finger found
in a variety of ribonucleoproteins (13,14).
MKRN2 harbours four C3H zinc fingers and a signature C3HC4 RING
zinc finger domain (15). The RING
domain is responsible for ubiquitin ligase activity, leading to
monoubiquitination and/or the synthesis of polyubiquitin chains on
lysine residues (16). To date, two
substrates for MKRN2 have been reported on, the p65 subunit of
NF-κB (P65) and the p85α subunit of PI3K (PI3Kp85α) (12,17).
According to previous studies, MKRN2 is a novel ubiquitin E3 ligase
targeting the p65 subunit of NF-κB to negatively regulate
inflammatory responses (17), and
MKRN2 inhibits the migration and invasion of non-small-cell lung
cancer by negatively regulating the PI3K/Akt signalling pathway
(12). In the present study, MKRN2
expression was higher in human malignant melanoma cell lines
compared with that in normal skin cell lines. Downregulation of
MKRN2 inhibited melanoma cell growth accompanying P53 upregulation,
and further experiments demonstrated that MKRN2 interacted with and
ubiquitylated P53. The present study suggests that MKRN2 may act as
a potential therapeutic target for melanoma.
Materials and methods
Plasmid construction
A scramble short hairpin RNA (shRNA) and three
MKRN2-targeting shRNAs inserted into pLKO.1 plasmids were purchased
from Sigma-Aldrich; Merck KGaA; their sequences are provided in
Table I. Single guide RNA (sgRNA)
targeting TP53 was designed using an online tool (http://crispr.mit.edu/), as previously described
(18). The designed sgRNAs (Table I) were synthesized as oligos (Sangon
Biotech Co., Ltd.), annealed and inserted into a PX330 vector that
was digested with BbsI. The proteins E1 [ubiquitin like
modifier activating enzyme 1 (UBA1)-hexahistidine (His6)], E2
[ubiquitin conjugating enzyme E2 D1 (UBCH5A)-His6] and His6-Ub, and
the plasmids pET28a-TP53-His6 and pGEX4T-1-Gst-MKRN2 were provided
by Professor Ronggui Hu (Chinese Academy of Sciences, Shanghai,
China).
 | Table I.Sequences of shRNAs for MKRN2 and
sgRNA for TP53. |
Table I.
Sequences of shRNAs for MKRN2 and
sgRNA for TP53.
| shRNA/sgRNA | Target site sequence
(5′-3′) |
|---|
| MKRN2 |
|
Scramble |
GCGCGATAGCGCTAATAATTT |
|
shRNA1 |
CCTATGGAACTCGGTGCAGAT |
|
shRNA2 |
GACCTCTTCATGCACCTTTCT |
|
shRNA3 |
GTCCAGAATGCCGTGTGATAT |
| TP53 |
|
sgRNA |
CCATTGTTCAATATCGTCCGGGG |
Cell culture and transfection
Human malignant melanoma cell lines A375, SK-MEL-28
and WM-115 were purchased from the American Type Culture Collection
(ATCC). The immortal human keratinocyte cell line HaCaT was
originally purchased from the ATCC, and the primary-cultured normal
human epithelial melanocyte cell line NHEM was originally purchased
from PromoCell GmbH. These two cell lines were provided by
Professor Ronggui Hu (Chinese Academy of Sciences, Shanghai,
China). All cell lines were cultured in DMEM supplemented with 10%
FBS, 100 U/ml penicillin and 100 mg/ml streptomycin (all from
Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a humidified
atmosphere of 5% CO2. The plasmids containing MKRN2
shRNAs or TP53 sgRNA (both 3 µg) were transfected into melanoma
cells using Lipofectamine® 2000 (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol, and
screened by puromycin (Thermo Fisher Scientific, Inc.) for two
weeks before subsequent experimentation.
Cell proliferation assay
Cells (3,000 cells/well) stably transfected with the
MKRN2 shRNAs were seeded into a 96-well plate. The 0 h time-point
was defined as 6 h after the cells were seeded. After 0, 24 or 48
h, the cells were incubated with MTT solution (cat. no. C0009;
Beyotime Institute of Biotechnology) for 4 h at 37°C. Subsequently,
the product (formazan) was dissolved in DMSO and quantified
spectrophotometrically at a wavelength of 570 nm using a microplate
reader (Bio-Rad Laboratories, Inc.). Experiments were conducted
with six replicates and repeated three times.
Colony formation assay
Cells (3,000 cells/well) stably transfected with the
MKRN2 shRNAs were seeded into a 6-well plate. After 7 days, plates
were fixed with 4% paraformaldehyde (Merck KGaA) at room
temperature for 30 min, stained with 0.1% crystal violet (C0121;
Beyotime Institute of Biotechnology), at room temperature for 30
min and washed three times with PBS buffer. Images were obtained
using a camera (DSC-W800; Sony Corporation), the number of colonies
was counted, and the average number was calculated.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells (100,000
cells/well) using a total RNA kit (cat. no. DP419; Tiangen Biotech
Co., Ltd.). cDNA was synthesized using ReverTra Ace qPCR RT Master
mix (Toyobo Life Science) at 37°C for 15 min, and 95°C for 5 min,
according to the manufacturer's protocol. qPCR was performed on an
ABI 7500 fast real-time PCR system using the following conditions:
Initial denaturation at 95°C for 2 min; denaturation at 95°C for 5
sec, annealing at 60°C for 20 sec, and elongation 72°C for 34 sec,
for 40 cycles (Applied Biosystems; Thermo Fisher Scientific, Inc.)
to assess the relative abundances of TP53 and cyclin dependent
kinase inhibitor 1A (p21) mRNAs using specific primers (Table II) with staining by SYBR Green
(Toyobo Life Science). The relative abundances of TP53 and p21 were
normalized to that of the GAPDH gene, using the 2−ΔΔCq
method (19,20). All data were obtained from three
independent experiments.
| Table II.Sequences of the primers used in
reverse transcription-quantitative PCR. |
Table II.
Sequences of the primers used in
reverse transcription-quantitative PCR.
| Target gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| GAPDH |
GAGTCAACGGATTTGGTCGTATTG |
ATTTGCCATGGGTGGAATCATATTG |
| TP53 |
CAGACCTATGGAAACTACTTCCTGA |
CTTCATCTGGACCTGGGTCTTC |
| p21 |
CTGTCACTGTCTTGTACCCTTGT |
GGAGTGGTAGAAATCTGTCATGCT |
Co-immunoprecipitation (COIP),
immunoprecipitation and immunoblotting
For COIP, A375 cells (500,000 cells/well) were lysed
in 500 µl COIP buffer (50 mM Tris-HCl, 150 mM NaCl, 5 mM EDTA and
1% NP-40, pH 7.6) supplemented with a protease inhibitor cocktail
(Roche Diagnostics). Subsequently, the cell lysates were
centrifuged at 4°C at 12,000 × g for 10 min, incubated with P53
antibody (1:100; cat. no. 10442-1-AP; ProteinTech Group, Inc.) and
Protein G agarose beads (Merck KGaA) overnight at 4°C, then washed
three times with COIP buffer. For immunoprecipitation, cells
(500,000 cells/well) were lysed in 500 µl immunoprecipitation
buffer (50 mM Tris-HCl, 150 mM NaCl, 5 mM EDTA, 0.1% SDS and 1%
NP-40, pH 7.6) supplemented with a protease inhibitor cocktail.
Subsequently, the cell lysates were centrifuged at 4°C at 12,000 ×
g for 10 min, incubated with P53 antibody and Protein G agarose
beads overnight at 4°C, then washed three times with COIP buffer.
The immunoprecipitates were enriched and denatured at 100°C for 10
min in 2X SDS-PAGE loading buffer (50 µl). The inputs,
immunoprecipitates and other cell lysates (10 µl/lane) were then
subjected to 10% SDS-PAGE, transferred to a PVDF membrane (Bio-Rad
Laboratories, Inc.) at 200 mA for 3 h. Following which, the
membrane was blocked with 5% skimmed milk at room temperature for 1
h, then incubated with the appropriate antibodies against MKRN2
(1:500 dilution; cat. no. 12238-1-AP; ProteinTech Group, Inc.), P53
(1:2,000 dilution; as aforementioned), P21 (1:1,000 dilution; cat.
no. 10355-1-AP; ProteinTech Group, Inc.), MDM2 proto-oncogene
(MDM2; 1:500 dilution; cat. no. 19058-1-AP; ProteinTech Group,
Inc.), P65 (1:1,000 dilution; cat. no. 10745-1-AP; ProteinTech
Group, Inc.), ubiquitin (1:1,000 dilution; sc-47721; Santa Cruz
Biotechnology, Inc.) and GAPDH (1:5,000 dilution; cat. no.
60004-1-Ig; ProteinTech Group, Inc.) overnight at 4°C, Subsequently
the membranes were washed three times with TBST (50 mM Tris-HCl,
150 mM NaCl and 0.1% Tween-20, pH 7.6) then incubated with
secondary antibodies [horseradish peroxidase (HRP)-conjugated
Affinipure goat anti-mouse IgG (H+L); cat. no. SA00001-1; 1:5,000
dilution and HRP-conjugated Affinipure goat anti-rabbit IgG (H+L);
cat. no. SA00001-2; 1:5,000 dilution; ProteinTech Group, Inc.] at
room temperature for 1 h, and washed three times with TBST. The
signals were visualized using high-signal ECL western blotting
substrate (cat. no. 180-5001) and a Tanon 5200 Imaging system (both
Tanon Science and Technology Co., Ltd.). The density of the protein
bands in Fig. 1 was calculated using
ImageJ software v1.8.0 (National Institutes of Health).
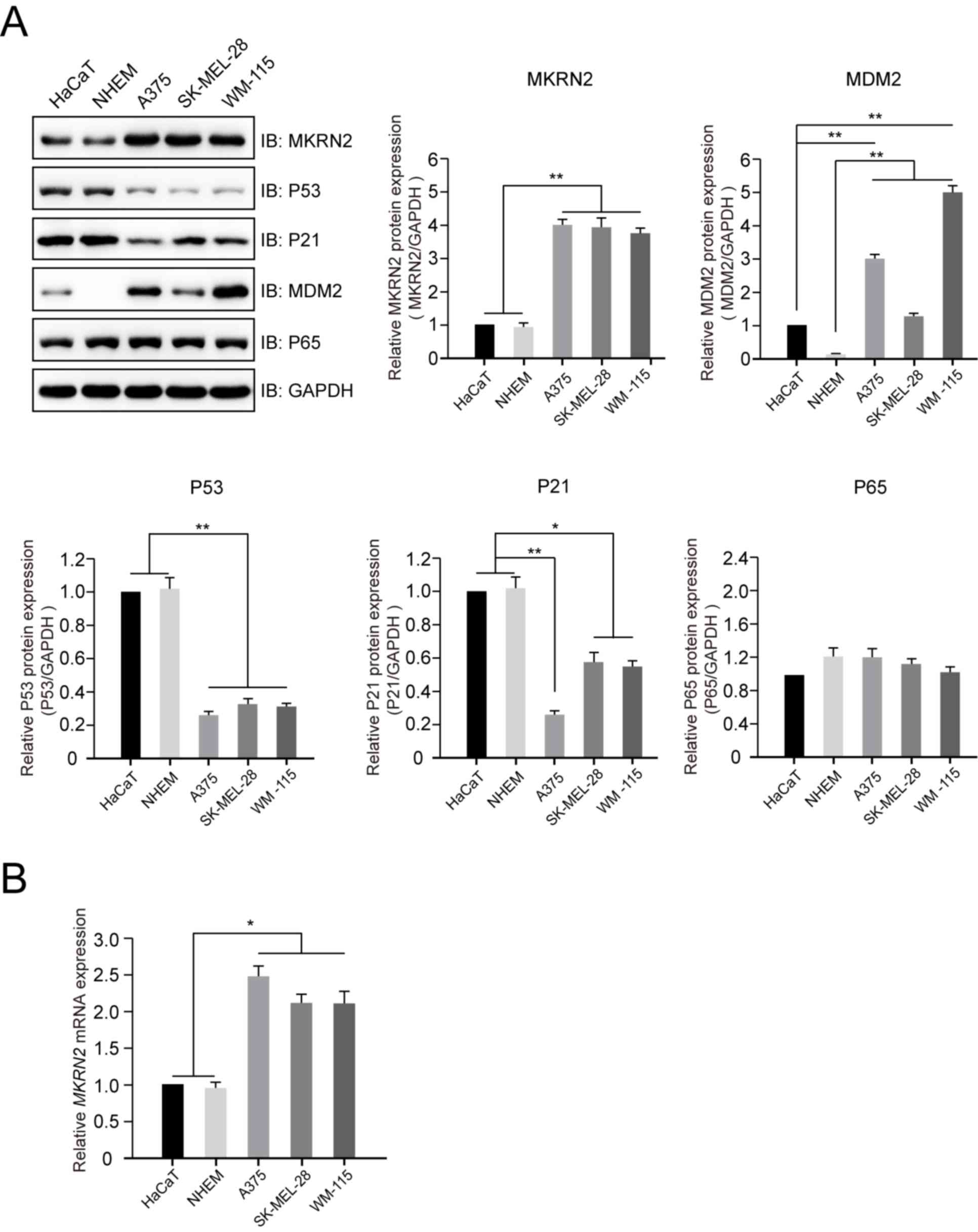 | Figure 1.Expression profile of MKRN2 in normal
human epithelial melanocyte or keratinocyte and human malignant
melanoma cell lines. (A) Protein expression profiles of MKRN2, P53,
P21, MDM2 and P65 in normal human epithelial melanocyte (A375,
SK-MEL-28 and WM-115), keratinocyte (HaCaT) and human malignant
melanoma (NHEM) cell lines detected by immunoblotting. The protein
expression levels of MKRN2, MDM2, P53, P21 and P65 in different
cell lines were compared with those of GAPDH. n=3. (B) mRNA
expression levels of MKRN2 in normal human epithelial melanocyte or
keratinocyte cell lines and human malignant melanoma cell lines
detected by reverse transcription-quantitative PCR. n=3. Data are
expressed as the mean ± standard deviation and were analysed using
one-way ANOVA with Tukey's post hoc test. *P<0.05, **P<0.01.
MDM2, MDM2 proto-oncogene; MKRN2, makorin ring finger protein 2;
P21, cyclin dependent kinase inhibitor 1A; P65, golgi reassembly
stacking protein 1; IB, immunoblot. |
Expression and purification of
recombinant proteins
Firstly, the pGEX4T-1-Gst-MKRN2 (Ampicillin
resistance) and pET28a-TP53-His6 (Kanamycin resistance) plasmids
were expressed in BL21 E. coli. Monoclones were picked,
cultured at 37°C in 2 ml lysogeny broth (LB) medium (10 g/l
tryptone, 10 g/l yeast extract and 10 g/l NaCl) with the respective
resistance overnight, then transferred to 1L LB medium and cultured
for 5 h at 37°C under normal air conditions. Following
isopropyl-β-d-thiogalactopyranoside (Sangon Biotech Co., Ltd.)
induction at 16°C overnight the cells were lysed in PBS buffer and
centrifuged at 4°C at 12,000 × g for 10 min, and incubated with
glutathione or Ni2+ TA beads (GE Healthcare) to enrich
the respective proteins. This was followed by elution with reduced
25 mM L-glutathione or 1 M imidazole dissolved in PBS buffer. The
resulting products were dialyzed in PBS buffer supplemented with
20% glycerol prior to being aliquoted and preserved at −80°C.
Glutathione S-transferase (GST)
pulldown assay
Purified Gst-MKRN2 (20 µg), P53-His6 (20 µg) and
Glutathione Sepharose 4B were incubated at 4°C overnight in 500 µl
pull-down buffer (20 mM Tris-Cl, 100 mM NaCl, 5 mM
MgCl2, 1 mM EDTA, 1 mM DTT, 0.5% (v/v) NP-40 and 10
µg/ml BSA (cat. no. SRE0096; Merck KGaA), pH 7.5). The beads were
then centrifuged at 2,000 × g at 4°C for 2 min, and washed three
times with pull-down buffer. Subsequently, the recovered beads were
denatured at 100°C for 10 min in 2X SDS-PAGE loading buffer and
subjected to immunoblotting analysis as aforementioned.
In vitro ubiquitination assay
In vitro ubiquitination assays were performed
as previously described (21).
Briefly, recombinant 200 ng His6-Ub, 200 ng UBA1-His6 (E1), 200 ng
UBCH5A-His6 (E2), 500 ng Gst-MKRN2 (E3) and 500 ng P53-His6 were
added to ubiquitination buffer (25 mM Tris-Cl, 100 mM NaCl, 1 mM
DTT, 5 mM MgCl2; pH 7.6; supplemented with 2 mM ATP)
with a final reaction volume of 50 µl and incubated at 37°C for 2
h. The ubiquitination levels of proteins were examined by an
immunoblotting assay using an anti-P53 antibody as
aforementioned.
P53-knockout cell line
The sgRNA targeting exon 1 of TP53 were designed and
inserted into PX330 vector (cat. no. 98750; Addgene, Inc.). A
P53−/− knockout cell line was established using the
CRISPR-Cas9 technique, as previously described (18). Briefly, A375 cells (100,000
cells/well) were transfected with 3 µg CRISPR-Cas9-based sgRNA
(PX330-P53-sgRNA), and monoclonal cells were chosen and detected by
immunoblotting analysis. Subsequently, genetic ablation of TP53 was
confirmed by first generation sequencing.
Statistical analysis
All experiments were performed in triplicate. All
values are presented as the mean ± SD. One-way ANOVA followed by
Tukey's post hoc multiple comparisons test was performed using
GraphPad Prism v7 (GraphPad Software, Inc.). P<0.05 was
considered to indicate a statistically significant difference,
whereas P<0.01 was considered to indicate a statistically very
significant difference.
Results
Higher expression of MKRN2 in human
malignant melanoma cell lines
To investigate the expression profile of MKRN2 in
human malignant melanoma cell lines, lysates of three melanoma cell
lines (A375, SK-MEL-28 and WM-115) and two normal cell lines
(HaCaT, immortal human keratinocyte cell line; and NHEM,
primary-cultured normal human epithelial melanocyte cell line) were
prepared. Immunoblotting analysis indicated that the protein levels
of MKRN2 were significantly higher in the three melanoma cell lines
compared with in the two normal cell lines (Fig. 1A), which was consistent with the mRNA
levels of MKRN2 detected by RT-qPCR (Fig. 1B). The protein expression levels of
P53 and P21 were higher in the two normal cell lines compared with
in the three melanoma cell lines. P65, a substrate for E3 ligase
for MKRN2, exhibited weak alterations in these five cell lines.
Additionally, MDM2, an E3 ligase for P53, expression was higher in
the three melanoma cell lines compared with in the two normal cell
lines (Fig. 1A). These results
suggested that MKRN2 exhibited higher expression in human malignant
melanoma cells and its expression was negatively associated with
P53 and P21 expression.
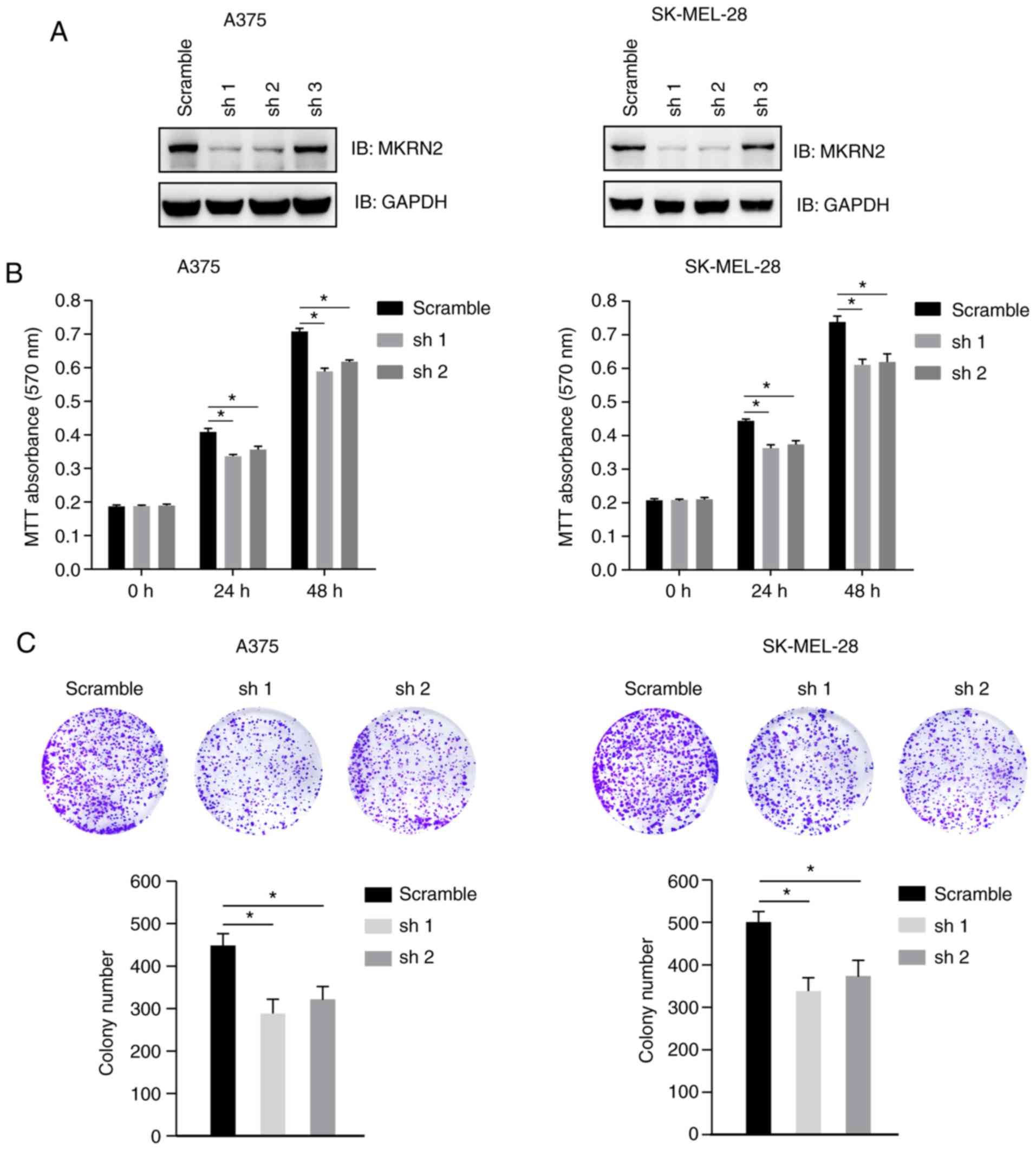
Downregulation of MKRN2 inhibits
melanoma cell growth
To investigate the effect of MKRN2 on melanoma cell
proliferation, three MKRN2 shRNAs were designed and tested in two
melanoma cell lines (A375 and SK-MEL-28). These two cell lines were
selected for further study, as the expression of MKRN2 was higher
in these two cell lines compared with that in WM-115.
Immunoblotting analysis indicated that MKRN2 expression was
markedly decreased in cells stably transfected with shRNA1 and
shRNA2, and these cells were selected for further study (Fig. 2A). Cell viability of A375 and
SK-MEL-28 cells was detected by MTT assay at 24 and 48 h post
culture, and significant cell growth arrest was observed in
MKRN2-knockdown cell lines compared with in the control group
(Fig. 2B). Decreased colony numbers
were identified in MKRN2-knockdown groups compared with in the
control group in A375 and SK-MEL-28 cells (Fig. 2C). These results suggested that
downregulation of MKRN2 inhibited melanoma cell growth.
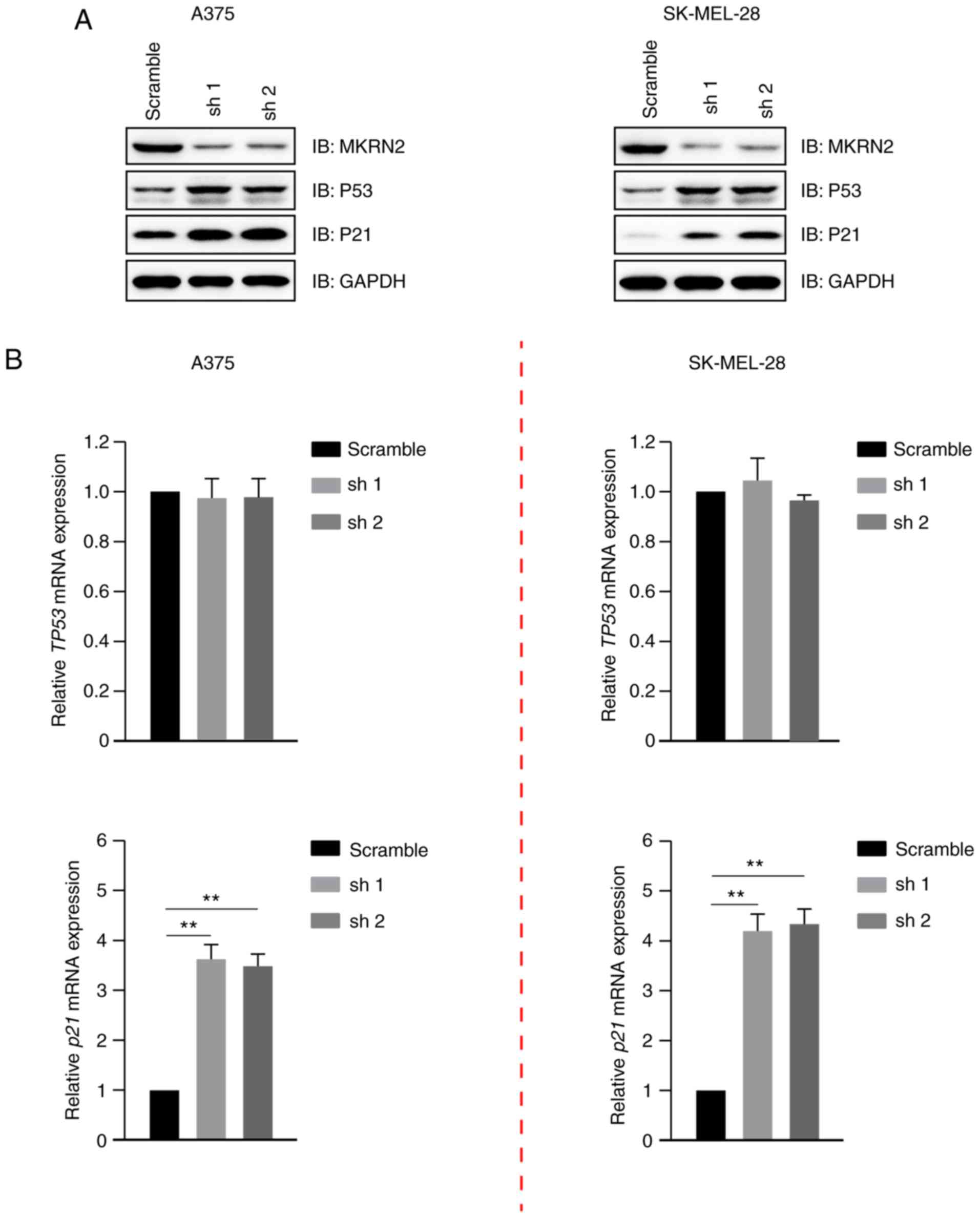
Downregulation of MKRN2 inhibits
melanoma cell growth by upregulating P53
MKRN2 expression exhibited a negative association
with P53 and P21 expression (Fig.
1A), and its effect on these was detected. Immunoblotting
analysis indicated that P53 and P21 expression were markedly
increased in MKRN2-knockdown cells compared with in the control
A375 and SK-MEL-28 cells (Fig. 3A).
It has been widely reported that P53 is a transcriptional activator
of the p21 gene (22,23). The gene expression levels of TP53 and
p21 were detected by RT-qPCR, which revealed that the mRNA levels
of p21, but not TP53, were upregulated in MKRN2-knockdown cells
(Fig. 3B). These data indicated that
MKRN2-knockdown increased the stability of the P53 protein and had
little effect on its mRNA levels.
MKRN2 interacts with and ubiquitylates
P53
The observations that MKRN2 affected the growth rate
of melanoma cells by affecting P53 protein levels prompted the
present study to determine whether P53 and MKRN2 could interact
with each other. MKRN2 expression was higher in the A375 cell line
compared with that in SK-MEL-28 and WM-115 cells; therefore A375
was selected for further study. Immunoprecipitation analyses
demonstrated that endogenous MKRN2 formed a complex with endogenous
P53 in A375 cells (Fig. 4A).
Subsequently, the present study assessed whether MKRN2 could
directly interact with P53 in vitro. Recombinant GST-tagged
MKRN2 and His6-tagged P53 were purified, and then GST pull-down
assays were performed. These assays demonstrated that MKRN2 and P53
formed a complex in vitro (Fig.
4B). An in vitro ubiquitylation assay was carried out
with recombinant proteins, and immunoblotting analysis indicated
that MKRN2 ubiquitylated P53 (Fig.
4C). Additionally, the present study indicated that ablation of
MKRN2 reduced the ubiquitylation levels of P53 in A375 cells
(Fig. 4D). Overall, these data
suggested that MKRN2 interacted with and ubiquitylated P53.
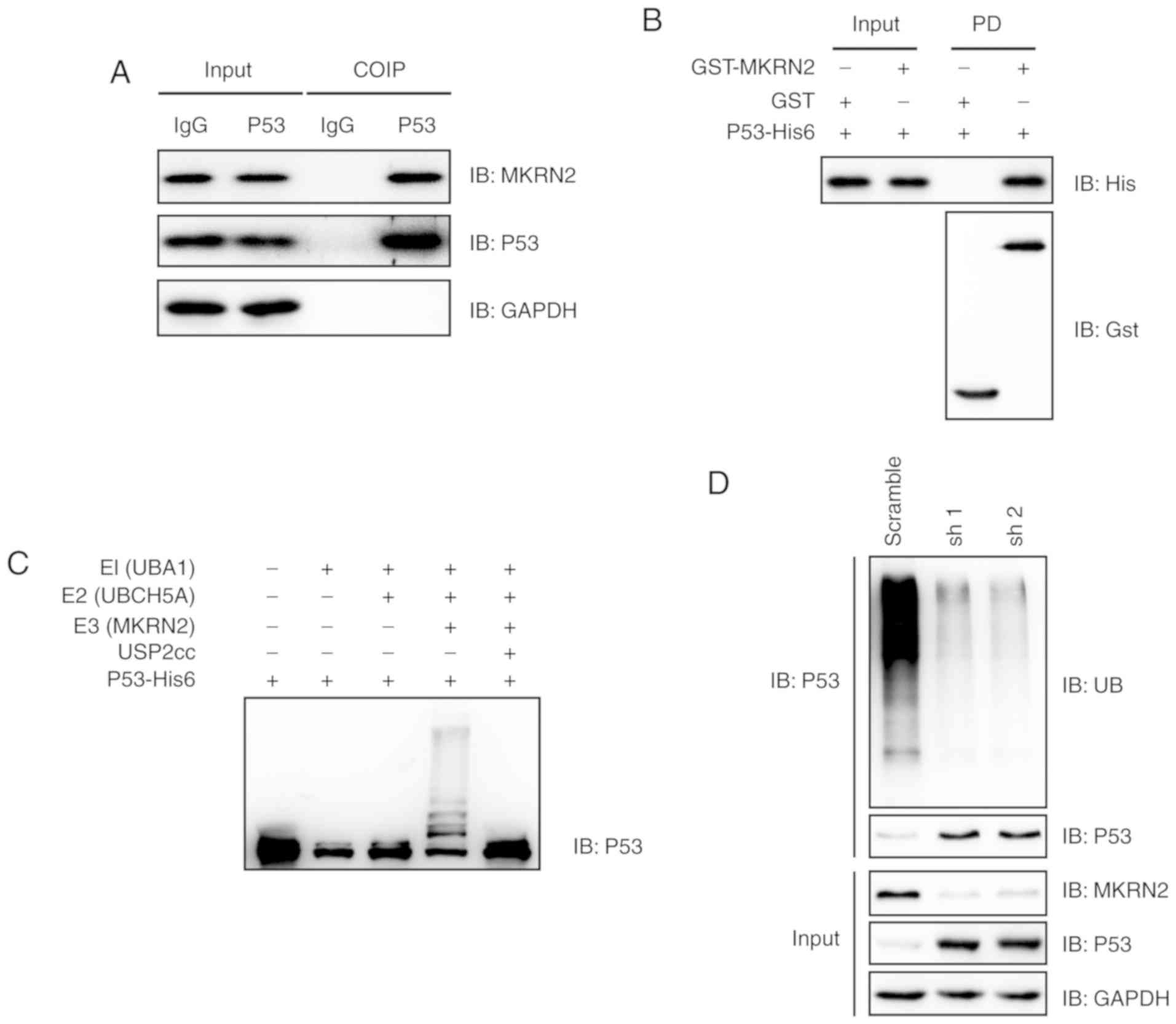 | Figure 4.MKRN2 interacts with and ubiquitylates
P53. (A) Endogenous MKRN2 and P53 formed a complex in A375 cells.
A375 cell lysates were immunoprecipitated with anti-P53 antibody
and subjected to immunoblotting analysis. (B) MKRN2 interacted with
P53 in vitro. Recombinant GST-tagged MKRN2 and His6-tagged
P53 were purified, and then GST pull-down assays were performed and
subjected to immunoblotting analysis. (C) MKRN2 ubiquitylated P53
in vitro. An in vitro ubiquitylation assay was
carried out with the recombinant proteins P53-His6, E1 (UBA1-His6),
E2 (UBCH5A-His6), His6-Ub and Gst-MKRN2, together with the
indicated components, and subjected to immunoblotting analysis. (D)
MKRN2 ubiquitylated P53 in vivo. A375 cells stably
transfected with MKRN2 shRNAs were lysed in RIPA buffer and
immunoprecipitated with anti-P53 antibody and subjected to
immunoblotting analysis using the indicated antibodies. COIP,
co-immunoprecipitation; GST, glutathione S-transferase; His,
histidine; IgG, immunoglobulin G; MKRN2, makorin ring finger
protein 2; sh, short hairpin RNA; shRNA, short hairpin RNA; UBA1,
ubiquitin like modifier activating enzyme 1; UBCH5A, ubiquitin
conjugating enzyme E2 D1; USP2, ubiquitin specific peptidase 2; IB,
immunoblot. |
MKRN2 regulates the proliferation of
melanoma cells in a P53-dependent manner
As MKRN2 expression was higher in the A375 cell line
compared with that in SK-MEL-28 and WM-115 cells, these were
selected for further study. A TP53-knockout
(TP53−/−) A375 cell line was generated using the
CRISPR-Cas9 sgRNA-based method and was validated by immunoblotting
analysis (Fig. 5A). First-generation
sequencing indicated that the knockout cell line had a 5-bp
deletion on exon 3 of the TP53 genomic DNA (Fig. 5A). Wild type or
TP53−/− A375 cells that stably expressed MKRN2
shRNAs were seeded into a 96-well plate, and cell viability was
detected using an MTT assay at 48 h. The results indicated that
MKRN2-knockdown induced growth arrest in wild type A375 cells but
had little effect in TP53−/− A375 cells.
Additionally, P53-knockout significantly promoted A375 cell
proliferation, independent of whether MKRN2 was knocked down
(Fig. 5B).
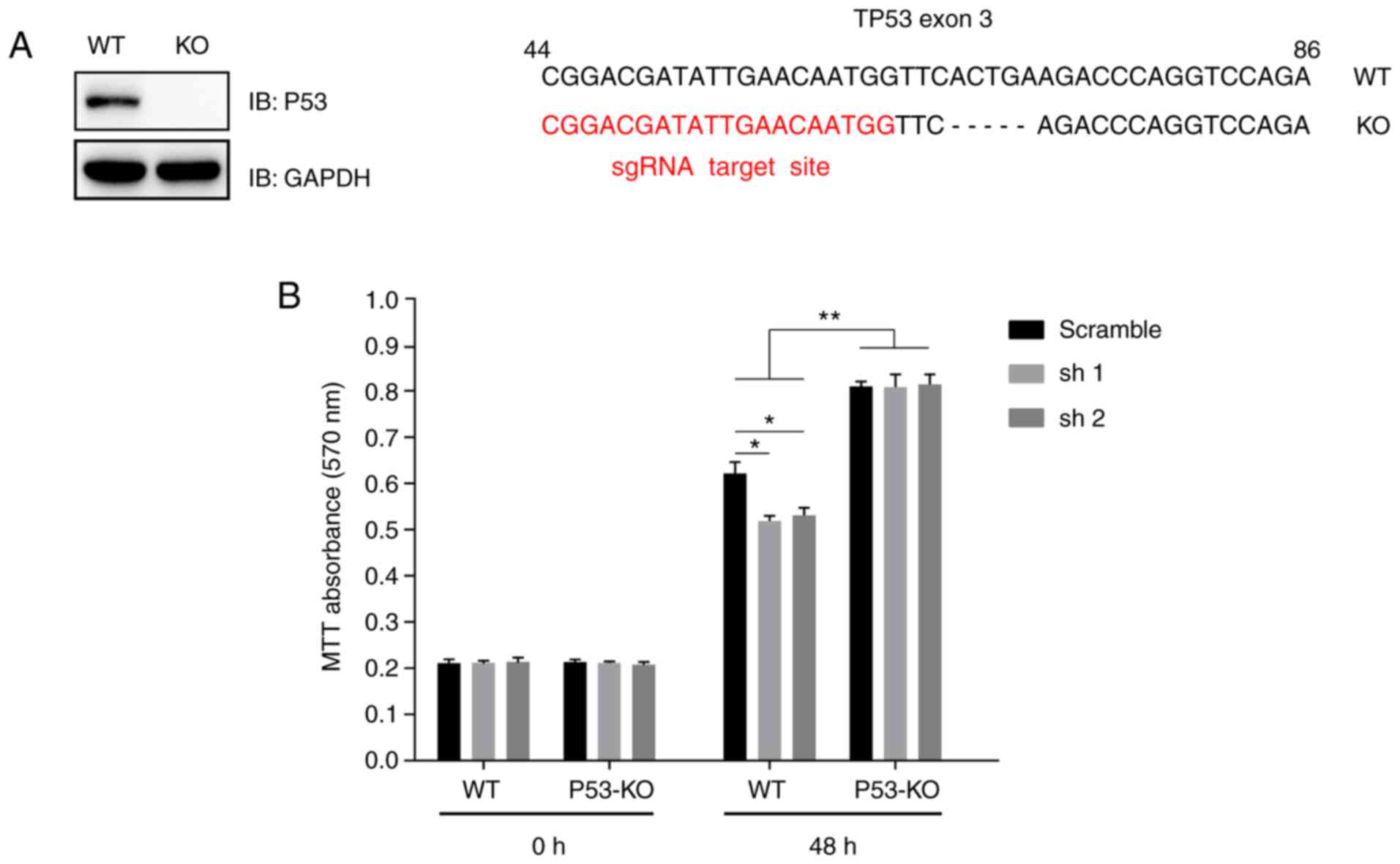 | Figure 5.MKRN2 regulates the proliferation of
melanoma cells in a P53-dependent manner. (A) A TP53−/−
A375 cell line was established using the CRISPR-Cas9 technique.
A375 cells were transfected with CRISPR-Cas9-based sgRNA indicated
in red (right panel), monoclonal cells were picked and detected by
immunoblotting analysis (left panel), and then genetic ablation of
TP53 with a 5-bp deletion was confirmed by sequencing. (B) MKRN2
knockdown induced melanoma cell growth arrest in a P53-dependent
manner. WT or TP53−/− A375 cells that stably expressed
MKRN2 shRNAs were seeded into a 96-well plate and detected by MTT
assay at the indicated time points (0 and 48 h). The 0 h time point
was defined as 6 h after the cells were seeded. Data are expressed
as the mean ± standard deviation and were analysed using one-way
ANOVA with Tukey's post hoc test. n=3. *P<0.05, **P<0.01. KO,
knockout; MKRN2, makorin ring finger protein 2; sgRNA, single guide
RNA; sh, short hairpin RNA; shRNA, short hairpin RNA; WT,
wild-type; IB, immunoblot. |
Discussion
The present study demonstrated that the protein
expression levels of MKRN2 were higher in melanoma cells and were
negatively associated with P53 and P21 expression. Furthermore,
downregulation of MKRN2 induced the arrest of melanoma cell growth,
which suggested that MKRN2 may act as an oncogene. In addition, the
present study identified that MKRN2 acted as a novel E3 ligase for
P53 and reduced the stability of the P53 protein. Finally, a
P53-knockout cell line was established, and it was demonstrated
that MKRN2 regulated the proliferation of melanoma cells in a
P53-dependent manner.
MKRN2 belongs to the MAKORIN family, which includes
three proteins: Makorin ring finger protein 1 (MKRN1), MKRN2 and
makorin ring finger protein 3 (24).
MAKORIN proteins share a highly homologous amino acid sequence,
particularly in the zinc finger domains, suggesting that they may
share similar functions or regulatory mechanisms (24). The function of MKRN1 has been
explored previously, and MKRN1 acts as an E3 ubiquitin ligase,
inducing degradation of human telomerase reverse transcriptase,
viral capsid proteins, p53 and p21 cell cycle regulators,
peroxisome proliferator activated receptor γ, adenomatous polyposis
coli and AMP-activated protein kinase α1 (24–27).
Similar to MKRN1, the present study identified that MKRN2
interacted with and ubiquitylated P53 (27); however, MKRN2 had no direct effect on
P21 protein stability.
A previous study has demonstrated that MKRN2
inhibits the migration and invasion of non-small-cell lung cancer
by negatively regulating the PI3K/Akt signalling pathway (12), suggesting that MKRN2 acts as a tumour
suppressor in lung cancer. This finding contradicts the present
study, which indicated that MKRN2 acts as an oncogene in melanoma.
However, a study by Lee et al (28) demonstrated that overexpression of
MKRN2 increases the proliferation of K562 cells, which is
consistent with the present study. Whether MKRN2 acts as a tumour
suppressor gene or oncogene may depend on the cancer type.
MKRN2 has been identified as an E3 ligase for the
p65 subunit of NF-κB and negatively regulates inflammatory
responses (17). However, P65
exhibited weak alterations in the three melanoma cell lines
compared with in the two normal cell lines used in the present
study. This result may suggest that the functions of MKRN2 depend
on cell/cancer types.
P53 is the most well-known tumour suppressor, and
>20 E3s have been reported, including MDM2, ubiquitin protein
ligase E3A and MKRN1 (27,29). The ubiquitination and degradation of
P53 serves an important role in cell cycle regulation and
tumorigenesis; the protein expression levels of MKRN2 and MDM2 were
higher in melanoma cells. Additionally, MDM2 is a well-known
proto-oncogene in numerous types of cancer, such as osteosarcoma,
glioblastoma and breast cancer, and it is an important target for
cancer therapy (30,31). Therefore, one may hypothesize that
MKRN2 may also be an important marker for melanoma and act as a
potential target for its therapy. Testing of MKRN2 function in
animal models and patient samples will be conducted in future
studies.
Acknowledgements
The authors would like to thank Professor Ronggui Hu
(State Key Laboratory of Molecular Biology, Institute of
Biochemistry and Cell Biology, Shanghai Institutes for Biological
Sciences, Chinese Academy of Sciences, Shanghai, China) for his
technical support and providing the proteins E1 [ubiquitin like
modifier activating enzyme 1 (UBA1)-hexahistidine (His6)], E2
[ubiquitin conjugating enzyme E2 D1 (UBCH5A)-His6] and His6-Ub, the
plasmids pET28a-TP53-His6 and pGEX4T-1-Gst-MKRN2, and the cell
lines (HaCaT and NHEM).
Funding
No funding was received.
Availability of data and materials
All data generated or analysed during the present
study are included in this published article.
Authors' contributions
GZ and YZ conceived and designed the experiments. YZ
and NC performed the experiments, collected the data and analysed
the results. GZ and YZ wrote the paper.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chu H, Li M and Wang X: Capsaicin induces
apoptosis and autophagy in human melanoma cells. Oncol Lett.
17:4827–4834. 2019.PubMed/NCBI
|
|
2
|
Mishra H, Mishra PK, Ekielski A, Jaggi M,
Iqbal Z and Talegaonkar S: Melanoma treatment: From conventional to
nanotechnology. J Cancer Res Clin Oncol. 144:2283–2302. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Martens MC, Seebode C, Lehmann J and
Emmert S: Photocarcinogenesis and skin cancer prevention
strategies: An update. Anticancer Res. 38:1153–1158.
2018.PubMed/NCBI
|
|
4
|
Rick JW, Shahin M, Chandra A, Dalle Ore C,
Yue JK, Nguyen A, Yagnik G, Sagar S, Arfaie S and Aghi MK: Systemic
therapy for brain metastases. Crit Rev Oncol Hematol. 142:44–50.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chowdhary M, Patel KR, Danish HH, Lawson
DH and Khan MK: BRAF inhibitors and radiotherapy for melanoma brain
metastases: Potential advantages and disadvantages of combination
therapy. Onco Targets Ther. 9:7149–7159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nicholas S, Mathios D, Jackson C and Lim
M: Metastatic melanoma to the brain: Surgery and radiation is still
the standard of care. Curr Treat Options Oncol. 14:264–279. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mandala M and Voit C: Targeting BRAF in
melanoma: Biological and clinical challenges. Crit Rev Oncol
Hematol. 87:239–255. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alexandrescu DT, Ichim TE, Riordan NH,
Marincola FM, Di Nardo A, Kabigting FD and Dasanu CA: Immunotherapy
for melanoma: Current status and perspectives. J Immunother.
33:570–590. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shimanovsky A, Jethava A and Dasanu CA:
Immune alterations in malignant melanoma and current immunotherapy
concepts. Expert Opin Biol Ther. 13:1413–1427. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Walter L and Heinzerling L: BRAF
Inhibitors and radiation do not act synergistically to inhibit WT
and V600E BRAF human melanoma. Anticancer Res. 38:1335–1341.
2018.PubMed/NCBI
|
|
11
|
Gray TA, Azama K, Whitmore K, Min A, Abe S
and Nicholls RD: Phylogenetic conservation of the makorin-2 gene,
encoding a multiple zinc-finger protein, antisense to the RAF1
proto-oncogene. Genomics. 77:119–126. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiang J, Xu Y, Ren H, Wudu M, Wang Q, Song
X, Su H, Jiang X, Jiang L and Qiu X: MKRN2 inhibits migration and
invasion of non-small-cell lung cancer by negatively regulating the
PI3K/Akt pathway. J Exp Clin Cancer Res. 37:1892018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hall TM: Multiple modes of RNA recognition
by zinc finger proteins. Curr Opin Struct Biol. 15:367–373. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Freemont PS: The RING finger. A novel
protein sequence motif related to the zinc finger. Ann N Y Acad
Sci. 684:174–192. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gray TA, Hernandez L, Carey AH, Schaldach
MA, Smithwick MJ, Rus K, Marshall Graves JA, Stewart CL and
Nicholls RD: The ancient source of a distinct gene family encoding
proteins featuring RING and C(3)H zinc-finger motifs with abundant
expression in developing brain and nervous system. Genomics.
66:76–86. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Borden KL: RING domains: Master builders
of molecular scaffolds? J Mol Biol. 295:1103–1112. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shin C, Ito Y, Ichikawa S, Tokunaga M,
Sakata-Sogawa K and Tanaka T: MKRN2 is a novel ubiquitin E3 ligase
for the p65 subunit of NF-κB and negatively regulates inflammatory
responses. Sci Rep. 7:460972017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu X, Li C, Gao X, Xia K, Guo H, Li Y, Hao
Z, Zhang L, Gao D, Xu C, et al: Excessive UBE3A dosage impairs
retinoic acid signaling and synaptic plasticity in autism spectrum
disorders. Cell Res. 28:48–68. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li C, Chen P, Zhang J, Zhang L, Huang X,
Yao Y, Che X, Fan X, Ge S and Wang Z: Enzyme-induced vitreolysis
can alleviate the progression of diabetic retinopathy through the
HIF-1α pathway. Invest Ophthalmol Vis Sci. 54:4964–4970. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Z, Chen P, Gao H, Gu Y, Yang J, Peng
H, Xu X, Wang H, Yang M, Liu X, et al: Ubiquitylation of autophagy
receptor Optineurin by HACE1 activates selective autophagy for
tumor suppression. Cancer Cell. 26:106–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dulic V, Kaufmann WK, Wilson SJ, Tlsty TD,
Lees E, Harper JW, Elledge SJ and Reed SI: p53-dependent inhibition
of cyclin-dependent kinase activities in human fibroblasts during
radiation-induced G1 arrest. Cell. 76:1013–1023. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
el-Deiry WS, Tokino T, Velculescu VE, Levy
DB, Parsons R, Trent JM, Lin D, Mercer WE, Kinzler KW and
Vogelstein B: WAF1, a potential mediator of p53 tumor suppression.
Cell. 75:817–825. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Abreu AP, Macedo DB, Brito VN, Kaiser UB
and Latronico AC: A new pathway in the control of the initiation of
puberty: The MKRN3 gene. J Mol Endocrinol. 54:R131–R139. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee HK, Lee EW, Seo J, Jeong M, Lee SH,
Kim SY, Jho EH, Choi CH, Chung JY and Song J: Ubiquitylation and
degradation of adenomatous polyposis coli by MKRN1 enhances
Wnt/β-catenin signaling. Oncogene. 37:4273–4286. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee MS, Han HJ, Han SY, Kim IY, Chae S,
Lee CS, Kim SE, Yoon SG, Park JW, Kim JH, et al: Loss of the E3
ubiquitin ligase MKRN1 represses diet-induced metabolic syndrome
through AMPK activation. Nat Commun. 9:34042018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee EW, Lee MS, Camus S, Ghim J, Yang MR,
Oh W, Ha NC, Lane DP and Song J: Differential regulation of p53 and
p21 by MKRN1 E3 ligase controls cell cycle arrest and apoptosis.
EMBO J. 28:2100–2113. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee KY, Chan KY, Tsang KS, Chen YC, Kung
HF, Ng PC, Li CK, Leung KT and Li K: Ubiquitous expression of
MAKORIN-2 in normal and malignant hematopoietic cells and its
growth promoting activity. PLoS One. 9:e927062014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sane S and Rezvani K: Essential roles of
E3 ubiquitin ligases in p53 regulation. Int J Mol Sci. 18(pii):
E4422017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gupta A, Shah K, Oza MJ and Behl T:
Reactivation of p53 gene by MDM2 inhibitors: A novel therapy for
cancer treatment. Biomed Pharmacother. 109:484–492. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Niazi S, Purohit M and Niazi JH: Role of
p53 circuitry in tumorigenesis: A brief review. Eur J Med Chem.
158:7–24. 2018. View Article : Google Scholar : PubMed/NCBI
|